



INTRODUCTION
Despite evidence for continuous human presence in the Maya Lowlands for at least 12,000 years (see Wrobel et al. Reference Wrobel, Hoggarth and Marshall2021), most bioarchaeological research in the region is dominated by a focus on the most visible and accessible contexts that date to the Classic period (a.d. 300–900/1000). Most of what we know about the biological variability of ancient populations, therefore, reflects the specific patterns of social, economic, political, and environmental complexity that characterized the large and highly stratified communities of this era. While studies of the Classic Maya provide foundational knowledge about these preindustrial state-level societies, as well as important lessons about the dire biological consequences of stresses placed on populations from environmental instability, political factionalization, and social inequality, equally fascinating is the story of the rise of large and complex Maya societies during the Preclassic (Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019; Scherer Reference Scherer2017; Somerville et al. Reference Somerville, Fauvelle and Froehle2013; Wright and White Reference Wright and White1996). Bioarchaeological studies in other areas of the world that have considered a deeper history have demonstrated that significant biocultural changes occurred as a result of growing social, economic, and technological complexity (e.g., Cohen and Armelagos Reference Cohen and George J1984; Cohen and Crane-Kramer Reference Cohen and Crane-Kramer2007; Larsen et al. Reference Larsen, Knüsel, Haddow, Pilloud, Milella, Sadvari, Pearson, Ruff, Garofalo, Bocaege, Betz, Dori and Glencross2019; Pihasi and Stock Reference Pihasi and Stock2011; Redfern Reference Redfern, Betsinger and DeWitte2020). It is widely accepted that a pan-Maya identity had arisen in eastern Mesoamerica by at least 1200/1000 b.c. (if not before), and that by the beginning of the Late Preclassic period, around 300 b.c., intensive agriculture, hierarchical rulership, and complex social and economic institutions were already well-established (Doyle Reference Doyle, Hutson and Ardren2020). Unfortunately, we know very little about how this long process unfolded in the Maya area from a biological perspective.
WHY STUDY EARLY SKELETONS?
Studies of human skeletons dating to the earliest phases of settled village life provide an essential source of biocultural data to address a broad range of questions posed by archaeologists, including those related to economies and subsistence, changing political and social conditions, and individual health and well-being. These have most often been framed through the lens of cultural evolutionary models, with an eye towards identifying and timing the precursors of the cultural and biological features characteristic of later time periods. In these contexts, bioarchaeological research tends to highlight behavioral correlates with the establishment and development of social, economic, and political complexity in the Maya region as reflected by skeletal remains and mortuary practices. This review identifies and discusses the picture provided by relevant skeletal data reported from the earliest lowland Maya communities. In support of this effort, we have created a table listing 398 individuals from 40 sites that are assigned Early and Middle Preclassic dates (1800–300 b.c.; Figure 1, Supplementary Table 1). These remains were identified through an extensive literature review, consultations with archaeologists focused on Preclassic contexts, and use of a database of Maya burials created and maintained by Vera Tiesler. While this list is not a completely exhaustive dataset, it represents the largest compilation to date of published Early and Middle burial information from the Maya region, broadly construed. The scope of this review is purposely limited to focus on our emerging picture of biology and mortuary behavior in the earliest Maya contexts, primarily in the Southern Lowlands, though we discuss relevant trends from adjacent regions when appropriate. For broader reviews of Maya bioarchaeology that include the more extensive mortuary record for the Late Preclassic and subsequent eras, we direct the reader to an excellent recent synthesis by Scherer (Reference Scherer2017; see also Spence and White Reference Spence and White2009; Wright and White Reference Wright and White1996).
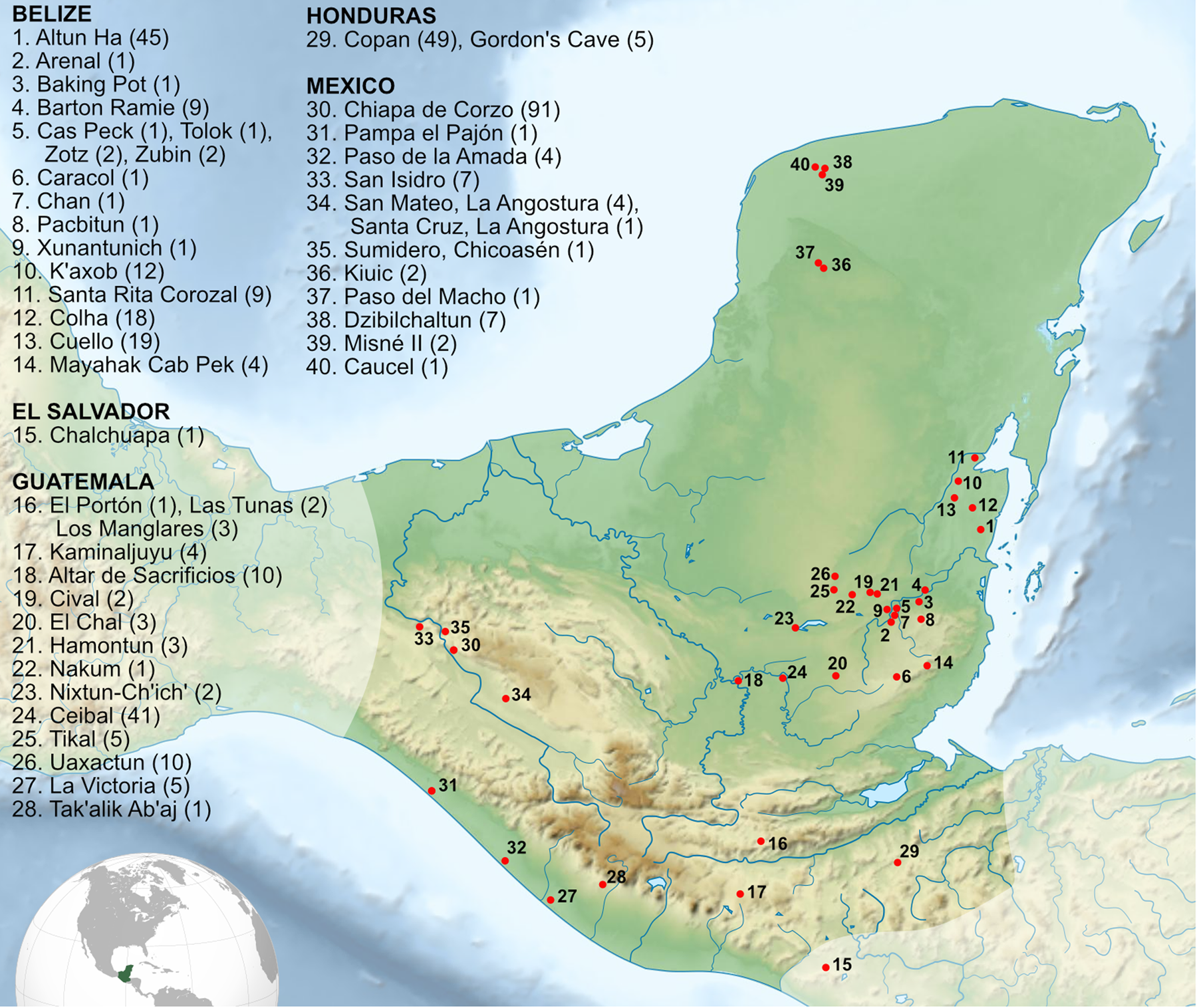
Figure 1. Map of sites in the Maya region with burials dating to the Early and/or Middle Preclassic period. Number of early burials for each site is in parentheses. Map created by Wrobel based on map by Sémhur, https://commons.wikimedia.org/wiki/File:Maya_civilization_location_map-blank.svg.
Our ability to synthesize the Early and Middle Preclassic mortuary and skeletal data is limited by a number of obstacles to bioarchaeological analyses, stemming from small and biased samples, limited geographic representation, lack of chronological control, and poor preservation, all of which are discussed in more detail at the end of the article. However, despite the frustrating nature of these assorted challenges, an increasing focus by archaeologists on early periods has yielded relevant skeletal data that, with careful consideration of confounding variables, provide us with an emerging picture of biological diversity of these poorly understood periods.
SKELETAL STUDIES OF THE EARLIEST MAYA
The study of early Maya skeletons has the potential to address several essential questions related to both biological and cultural processes, as well as foundational issues about how these sets of factors are intimately entwined. Thus far, bioarchaeological investigations of early Maya communities have focused on changes in diet and health that relate to increasing reliance on domesticated plants, population density, and trade and exchange, and on diversification of mortuary practices that relate to increasing social complexity and ritual elaboration.
THE MORTUARY RECORD
Mortuary treatment is the main source of data reported for Early and Middle Preclassic Maya burials. Specifically, there have been a number of studies that have used structured burial data to document changes in mortuary ritual over the course of the Preclassic, interpreting these changes within the context of increasing social complexity and broader integration of the Maya with each other and with other Mesoamerican groups (see recent review by Horn et al. Reference Horn, Powis and Pendergast2020; Ebert and Awe Reference Ebert and Awe2021). The Middle Preclassic is often identified as a precursor to the fluorescence of Maya civilization, hallmarks of which are established in the Late Preclassic (see discussion by McAnany Reference McAnany and McAnany2004:4–5). From a mortuary perspective, this transition is usually demonstrated by highlighting examples of new forms of behavioral complexity, including standardization of burial practices between sites and regions, and the appearance of wealthy and/or exotic grave good assemblages, specialized mortuary facilities in both public and residential settings, and a diverse array of funerary and non-funerary body treatments. Beyond the constructed environments of early population centers, skeletons from caves and rock shelters in Honduras (Brady Reference Brady1995), southern Belize (Hill et al. Reference Hill, Moes, Trask, Kate, McKenzie, Edgar, Kennett and Prufer2018; Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020), and the Yucatan (González et al. Reference González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2013; Wrobel, Hoggarth, and Marshall Reference Wrobel, Hoggarth and Marshall2021, this Special Section) show that use of these mortuary locations, which was widespread in the region by the Late Preclassic and Protoclassic (Garza et al. Reference Garza, Brady and Christensen2001; Wrobel et al. Reference Wrobel, Freiwald, Michael, Helmke, Awe, Kennett, Gibbs, Ferguson and Griffith2017a, Reference Wrobel, Morton, Michael, Biggs and Andres2017b), are consistent with traditions that have origins at least as far back as the Late Pleistocene. Middle Preclassic mortuary behavior does appear to be generally less complex than that of the Late Preclassic, especially in earlier contexts. As noted in other articles in this Special Section, however, communities established prior to 300 b.c. were highly variable in their sociopolitical and economic systems. Accordingly, the mortuary record also demonstrates intriguing indicators of complexity emerging earlier.
One of the main uses of mortuary data from early contexts has been to complement archaeological studies investigating the origins of the large and elaborate public architecture that dominates Classic-period site cores. These early buildings, which begin to be constructed in earnest during the Middle Preclassic (~800–700 b.c.), are typically low platforms and often contain cached objects assumed to be ritual paraphernalia (Inomata et al. Reference Inomata, Triadan, Aoyama, Castillo and Yonenobu2013, Reference Inomata, MacLellan and Burham2015; Rice Reference Rice2015, Reference Rice2021). Specifically, placement of burials within these non-residential structures is assumed to broadly reflect public social roles within the community (rather than identification with household contexts in which most others are buried). Thus, public burial location is seen as a valuable means of identifying evidence for social differentiation of individuals or groups responsible for their construction and the public activities performed there.
Within settlement centers, the placement of the earliest funerary contexts is often generalized as being largely restricted to residential architecture. Burial under house floors and in nearby structures is a widespread practice that appears to be established early and persists, alongside a diversity of other mortuary practices, throughout the pre-Contact period (Joyce Reference Joyce, Grove and Joyce1999; McAnany Reference McAnany1995). At sites such as Cuello and K'axob in northern Belize, for instance, almost all individuals dating to the Early and Late Middle Preclassic were found in houses or ancillary structures (Hammond Reference Hammond and Grube1995:50, 52; Henderson Reference Henderson2003; McAnany et al. Reference McAnany, Storey and Lockard1999:134). While it is true that most burials are found in residential contexts (in the Classic period as well), it seems likely that our picture is restricted by where we dig, and that relatively few individuals were buried in association with architecture. Even at sites like Cuello and K'axob, where burials are found in residential buildings, it should be noted that these make up only a tiny fraction of the total population, suggesting that most community members were buried elsewhere; thus, while some rituals were focused on social reproduction of household groups, residential burial was only a small part of the mortuary program. There also appear to be many exceptions to residential placement, even among the earliest burials, suggesting that mortuary practices among early Maya communities were already quite diverse, likely reflecting social identities beyond associations with specific households or local corporate groups (Joyce Reference Joyce, Grove and Joyce1999; McAnany and López Reference McAnany and López1999; McAnany et al. Reference McAnany, Storey and Lockard1999). For instance, no burials were found within residential structures in the earliest habitation phases at Chiapa de Corzo (prior to 550 b.c.) or Ceibal (prior to 770 b.c.), indicating the presence of some other, as yet unidentified non-residential interment areas (Lowe Reference Lowe and Agrinier1964; Palomo et al. Reference Palomo, Inomata and Triadan2017:310). At Ceibal (formerly Seibal), very few burials were found in residential contexts until the Late Preclassic, prior to which they instead appear in public ceremonial complexes and large platforms. Other sites also demonstrate early mortuary use of public architecture, including round structures at Altun Ha (Pendergast Reference Pendergast1982:Figure 98) and Cahal Pech (Aimers et al. Reference Aimers, Powis and Awe2000; Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019), an E-Group at Ceibal (Palomo et al. Reference Palomo, Inomata and Triadan2017:310), and platforms at Cuello (Hammond et al. Reference Hammond, Clarke and Robin1991, Reference Hammond, Clarke and Belli1992), Santa Rita Corozal (Chase and Chase Reference Chase and Chase2006:89; Chase et al. Reference Chase, Chase and Chase2018:160), and Ceibal (Palomo et al. Reference Palomo, Inomata and Triadan2017:310–311).
Mortuary patterning and analyses of grave furnishings have been useful in exploring status differentiation and the development of social hierarchy within early communities, and the presence of exotic items within graves may indicate that the source of wealth is tied to trade with other regions. Overall, the earliest burials include few, if any, grave goods, and when present they tend to be relatively simple and sourced locally (Hammond et al. Reference Hammond, Clarke and Belli1992:963, Reference Hammond, Clarke and Donaghey1995:127; McAnany et al. Reference McAnany, Storey and Lockard1999:134). These items include personal adornments, such as earrings, necklaces, pectorals, and wristbands, as well as ceramic vessels and figurines, animal bones, and stone artifacts. While most (but not all) Middle Preclassic graves are furnished modestly or not at all, over time the presence of grave goods and examples of ostentation become more common, though the extent of this varies greatly between communities.
At Altun Ha, the earliest burials, dated sometime before 600 b.c., were found in Structure C-13 and mostly have few or no associated artifacts. Pendergast (Reference Pendergast1982:200) notes a hiatus in mortuary use of the building, after which it was refurbished into a round structure that was used intensively for burial. This later phase of use likely dates to 500–400 b.c. and several of the associated burials have rich furnishings, including trade items like jadeite, spondylus, and obsidian. Similarly, at Cuello, grave goods were common by the end of the Early Preclassic, and by the Bladen phase (900–600 b.c.) of the Middle Preclassic were also included with children, suggesting the establishment of affiliative status (Hammond et al. Reference Hammond, Clarke and Robin1991:362, 1992:961). The appearance of long-distance trade items (jade and greenstone) in the burial assemblage by ~700 b.c. coincides with other indicators of social complexity at the site (Hammond Reference Hammond and Grube1995:50). Burials at Ceibal dating to the eighth century b.c. include elaborate grave wealth (Palomo et al. Reference Palomo, Inomata and Triadan2017:310–311), and at the Puuc site of Paso del Macho, an Early Middle Preclassic adolescent male was buried with numerous ceramic vessels in the center of a plaza and in close association with several jade caches (Parker et al. Reference Parker, Bey and Negrón2020). Finally, there are a number of examples of particularly wealthy graves, likely indicative of the individuals’ leadership roles. In particular, Cuello's B160, dated to ~500 b.c., includes ceramics, jade and shell beads, carved bone tubes, and a gorget with a mask design (Hammond et al. Reference Hammond, Clarke and Belli1992:961–963). At Santa Rita Corozal, Burial SD P24A-1 contained three pottery vessels and four small tubular jadeite beads (Chase and Chase Reference Chase and Chase2006:93). At K'axob, Burial 43 was interred with more than 2,000 small, marine-shell beads (arranged as bracelets and upper arm bands) and two ceramic vessels (McAnany and López Reference McAnany and López1999:155). By the Late Preclassic, there appears to be a shift in elaborate burial activity from domestic contexts to public/communal ones (Hammond Reference Hammond and Grube1995:57).
Changes in early mortuary ritual within some communities include increased standardization of mortuary treatment, which may indicate more rigid and defined social messaging, perhaps related to group identity. For instance, at Santa Rita Corozal, Middle Preclassic burials often contain a single small vessel placed upright near or on the chest (Chase et al. Reference Chase, Chase and Chase2018:160). At Cuello, the earliest burials are diverse, while later burials in more formal Middle Preclassic buildings have more uniform grave goods and are treated more consistently, such as having vessels covering their skulls and standardized body positioning (Hammond Reference Hammond and Grube1995:50; Hammond et al. Reference Hammond, Clarke and Robin1991:Figure 5, 1992).
Body treatment can also reflect aspects about social organization at the level of the community and broader region. The vast majority of burials from the Early and Middle Preclassic are primary interments in simple pits, with no evidence of intensive or extended mortuary ritual (McAnany et al. Reference McAnany, Storey and Lockard1999:134). The most elaborate funeral architecture consists of cists in which slab stones surround the body (Hammond et al. Reference Hammond, Clarke and Robin1991:Figure 5, 1992). Many of the more complex mortuary practices that are ubiquitous in the Late Preclassic, such as secondary and multiple burials, appear to be almost completely absent in the Middle Preclassic. However, there are notable exceptions that demonstrate the presence of mortuary complexity beginning in some communities much earlier than 300 b.c. For instance, at Santa Rita Corozal, Chase and colleagues (2018:164) note a link in the Middle Preclassic between fire pits and interments, which is perhaps a precursor to ritual burning commonly found at many sites throughout the Classic period. The burials from Altun Ha's Structure C-13, dating to the later Middle Preclassic, include secondary interment (Pendergast Reference Pendergast1982:178) of five individuals placed atop two primary individuals (Burial C-13/5 [RP-610]). This structure also contained a large number of caches, some with human bones, perhaps indicating more elaborate and extended mortuary practices. Similarly, Cuello and K'axob each have secondary burials dating to the Early Middle Preclassic (Robin and Hammond Reference Robin, Hammond and Hammond1991:208; Storey Reference Storey and McAnany2004). Several burnt long bone fragments were found at Cahal Pech dating to the Terminal Early Preclassic (Cunil phase), and Awe (Reference Awe1992:335) suggests that these likely reflect ritual practices. As early as 775–700 b.c., Ceibal's burials include examples of multiple and secondary burial, and burials found in public areas, including infants (Palomo et al. Reference Palomo, Inomata and Triadan2017:310–311). Cut marks are visible on the chin of Middle Preclassic Burial 9 at Cuello, which was also the only Middle Preclassic individual at the site buried in a seated position (Saul and Saul Reference Saul, Saul and Hammond1991:155; Hammond Reference Hammond and Grube1995:50). Perhaps most dramatic is the evidence of postmortem modification of a cranium from Cuello. The frontal had holes drilled in it to allow suspension, perhaps as a personal adornment, and was found in what may be a feasting deposit. Hammond and colleagues (2002:952) interpret this object as likely relating to ancestor veneration, and note the presence of a separate Preclassic primary inhumation whose skull had been removed secondarily.
Studies of ancient social systems have utilized demographic data to test whether specialized social or economic roles were assigned based on age and/or sex. This practice could be indicated by differential access to specific mortuary locations or mortuary treatments. While poor preservation limits the reliability of sex and age estimation, and therefore the scope of interpretations, what is clear is that these early Maya burial contexts are not true cemeteries, and instead comprise groups of individuals selected for some reason as yet unclear. For example, Middle Preclassic skeletal assemblages are dominated by males and by adults, suggesting that the skeletal series are not representative of the general population (Saul and Saul Reference Saul, Saul, Whittington and Reed1997; Thompson Reference Thompson2005). Out of the 130 sexed individuals in Supplementary Table 1, 84 were male or likely male, while only 46 were female or likely female. Of the rest, 90 were not sexed because they were infants or juveniles, while the remaining 179 were adolescents or adults with no sex estimate reported. The presence of adults of both sexes as well as children in most contexts suggests that selection criteria were likely based primarily on family or corporate group membership, rather than on other specific achieved social roles. These roles may instead be reflected in other variations in burial treatment or furnishings. It is certainly likely that taphonomy partially accounts for subadults and females being underrepresented (Gordon and Buikstra Reference Gordon and Buikstra1981), but this pattern is consistent with some (Hernández Espinoza and Márquez Morfín Reference Espinoza, Olga and Márquez Morfín2015; Wright Reference Wright2006), but not all (Wrobel et al. Reference Wrobel, Morton, Michael, Biggs and Andres2017b:312) later mortuary contexts from both public and residential contexts in which preservation is better, suggesting that cultural practices are also at work in at least some instances. Among the earliest burials at Cuello, for example, Hammond (Reference Hammond and Grube1995:50) notes no correlations between age and sex categories and access to multiple grave goods or long-distance trade items. Thus, while the small groups of burials do not provide data useful in paleodemographic reconstructions, these data instead provide a perspective on social organization through the lens of differential access to mortuary locations.
In sum, it does appear that mortuary complexity increases over time at sites, and that this is related to broader social changes within these communities. However, it is also clear that there is a lot of variation in mortuary behavior between communities and that aspects of the more standardized forms of mortuary complexity characterizing the Late Preclassic have their roots in the Middle Preclassic, appearing in different ways and at different times. It stands to reason, then, that many of the aspects of mortuary complexity that are well-documented for the Late Preclassic have their origins in the Middle Preclassic. In part, this issue of timing is clouded by confusion about the ceramic sequence. Chase and Chase (Reference Chase and Chase2006:96) note that ceramic complexes usually considered to be sequential in fact co-occur in stratigraphic levels and even within individual graves, suggesting that there is temporal overlap between them. Furthermore, it is worth noting that the dating and ceramic forms for the Middle Preclassic are not uniform throughout the Maya area, and the 300 b.c. date that typically marks the break between the Middle and Late Preclassic ceramic forms is simply a convention that was established at Uaxactun (Arlen Chase, personal communication 2020). Thus, the complicated nature of the mortuary record undoubtedly demonstrates that the transition to complexity does not occur everywhere at the same time. This should come as no major surprise, as many of the more complex mortuary practices occur first in larger communities, such as Ceibal and Altun Ha.
MOBILITY
While mobility studies using strontium, oxygen, and, more recently, sulfur and lead isotopes, have seen an explosion in the recent Maya bioarchaeology literature (e.g., Ebert et al. Reference Ebert, Rand, Green-Mink, Hoggarth, Freiwald, Awe, Trask, Yaeger, Kathryn Brown, Helmke, Guerra, Danforth and Kennett2021; Freiwald Reference Freiwald, Hutson and Ardren2020; Price et al. Reference Price, Burton, Sharer, Buikstra, Wright, Traxler and Miller2010, Reference Price, Burton, Fullagar, Wright, Buikstra, Tiesler and Cucina2015; Sharpe et al. Reference Sharpe, Kamenov, Gilli, Hodell, Emery, Brenner and Krigbaum2016), there has been a clear dearth of analyses focused on the earliest skeletal remains from the region, inhibiting our ability to make any type of definitive statement about movement and migration during the Early and Middle Preclassic. Available strontium (87Sr/86Sr) data, which can reflect migration from isotopically distinct regions when 87Sr/86Sr values in tooth enamel differ from the geologic 87Sr/86Sr values of the burial location (Price et al. Reference Price, Burton and Bentley2002), primarily come from a recent study by Palomo (Reference Palomo2020) at Ceibal. Values for 87Sr/86Sr, combined with lead and oxygen from enamel, show that at least one individual (Burial 132C) from the Early Middle Preclassic Real 3 phase (1000–700 b.c.) was non-local to Ceibal, possibly migrating from the Usumacinta or Peten regions of Guatemala (Palomo Reference Palomo2020:183). The 87Sr/86Sr data from individuals dating to the Late Middle Preclassic Escoba (1/2) phase (700–300 b.c.), show that all individuals were local to the Ceibal region, despite many burials representing children found in a sacrificial context on the E-Group (Palomo Reference Palomo2020:187). Isotopic studies of mobility using 87Sr/86Sr analyses are not well-represented from other parts of the Maya region, though available data from the Belize Valley region (see Freiwald Reference Freiwald, Hutson and Ardren2020) document limited migration during the Middle Preclassic. In another study, Wright and colleagues (Reference Wright, Valdés, Burton, Price and Schwarcz2010) used δ18O values from tooth enamel for examining movement and migration from the Middle Preclassic through Late Postclassic at the highland site of Kaminaljuyu. The single Middle Preclassic individual in this study had a δ18O value that was low, but still consistent with a local origin.
Mobility studies represent an important topic for future research, as much of the interest in early sites is focused on determining the extent of political and economic connections between early population centers within the Lowlands and between other regions (e.g., Gulf Coast Olmec, central Mexico). Additional studies seeking to identify migrants by their non-local isotopic signatures could help to clarify these broader questions, determining whether the presence of exotic trade items at sites was accompanied by the arrival of new groups moving into the area, or whether initiation of large-scale construction at sites resulted from the expansion of populations from established centers or was spurred internally. Studies of later Maya groups have shown that long-distance movement of individuals was quite common throughout the Classic and Postclassic periods (for examples, see Freiwald Reference Freiwald2011, Reference Freiwald, Ebert, Hoggarth and Awe2018, Reference Freiwald, Hutson and Ardren2020; Hodell et al. Reference Hodell, Quinn, Brenner and Kamenov2004; Price et al. Reference Price, Burton, Sharer, Buikstra, Wright, Traxler and Miller2010, Reference Price, Burton, Fullagar, Wright, Buikstra, Tiesler and Cucina2015; Trask Reference Trask2018), and it seems likely that the same would be true for the earliest communities.
DIET AND HEALTH
In other areas of the Americas (and elsewhere in the world), there has been a major emphasis on documenting the transition to agriculture as it relates to the rise of social complexity. In the Maya Lowlands, and the rest of Mesoamerica, the adoption of domesticates predates the Early Preclassic, but is still relevant for understanding the formation of early Maya culture. Skeletal studies from other world regions have traditionally focused on characterizing health, dietary, and demographic shifts that accompanied changing subsistence patterns by comparing data from hunter-gatherers, horticulturists, and agriculturists (Cohen and Armelagos Reference Cohen and George J1984; Cohen and Crane-Kramer Reference Cohen and Crane-Kramer2007; Larsen et al. Reference Larsen, Knüsel, Haddow, Pilloud, Milella, Sadvari, Pearson, Ruff, Garofalo, Bocaege, Betz, Dori and Glencross2019). This body of work has documented a variety of complex biocultural effects related to the transition to agriculture, demonstrating how it related to phenomena like population growth and movement, intentional and unintentional changes of local environments, the rise of new infectious and noninfectious diseases, and changes in social organization and complexity. In Mesoamerica, most archaeological studies focused on this transition have come from southwest and central Mexico, where teosinte was first domesticated around 9,000 years ago, as evidenced by changes in corn cobs found in stratified deposits (Blake Reference Blake and Blake2015) and modern genomic studies (Doebly Reference Doebly2004; Kistler et al. Reference Kistler, Maezumi, De Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Gilbert, Freitas and Allaby2018). Even in the homeland of maize, however, human skeletal remains dating to this period (>3,000 years ago) are quite rare and analysis is further limited by poor preservation and contamination issues (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). In the Maya Lowlands, the presence of maize microfossils, including phytoliths and starch, indicate its dispersal across Mesoamerica by ~5000 cal b.c., along with other domesticates (e.g., Curcurbita; Piperno Reference Piperno2011). Evidence for maize cultivation in the form of microfossils first appears around ca. 4500 b.c. in northern Belize (Rosenswig et al. Reference Rosenswig, Pearsall, Masson, Culleton and Kennett2014), with more intensive agriculture associated with dramatic deforestation and erosion across the Lowlands between 2500 and 1500 cal b.c. (Anselmetti et al. Reference Anselmetti, Hodell, Ariztegui, Brenner and Rosenmeier2007; Pohl et al. Reference Pohl, Pope, Jones, Jacob, Piperno, deFrance, Lentz, Gifford, Danforth and Josserand1996). Direct evidence for maize consumption through isotopic paleodietary studies, however, is limited in central Mexico and the Maya Lowlands by the paucity of skeletal remains associated with early agricultural contexts, biasing studies about the rise of agriculture in Mesoamerica to focus on later intensification rather than its initial establishment.
Recent work in the Maya Mountains of southern Belize has provided a rare glimpse of the transition to agriculture from a skeletal perspective (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). An analysis of stable carbon (δ 13C) and nitrogen (δ 15N) isotope measurements focuses on 52 radiocarbon-dated individuals interred in two rock shelters over the course of around 10,000 years. When combined, δ 13C and δ 15N isotope measurements on human bone collagen are a well-established proxy for the types of dietary proteins consumed by individuals (Ambrose and Krigbaum Reference Ambrose and Krigbaum2003; DeNiro and Epstein Reference DeNiro and Epstein1978, Reference DeNiro and Epstein1981). Values for δ 13C in plants are determined by photosynthetic pathways used by C3 (Calvin-Benson; trees, shrubs) and C4 (Hatch-Slack; grasses) species (Smith and Epstein Reference Smith and Epstein1971; van der Merwe Reference van der Merwe1982). While most plant foods consumed by the prehistoric Maya were C3 plants, the most frequently consumed C4 plant was maize, allowing δ 13C values to track the increasing dietary importance of maize as a staple crop. Nitrogen isotope (δ 15N) values in human bone vary with trophic level, increasing stepwise by approximately +3 to +5‰ between trophic levels (Bocherens and Drucker Reference Bocherens and Drucker2003). Because the well-preserved skeletal series from the Maya Mountains rock shelters is the only one in the lowland neotropics that spans the transition to maize-based food production, it provides unique and important information about the nature and timing of agricultural adoption in the region. Based on shifts in δ 13C values, Kennett and colleagues (Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020) found that maize became a significant dietary staple (~30 percent of total diet) starting around 2700–2000 b.c. After this time, maize contributed over 70 percent of the total dietary protein for most individuals, resulting in a dietary significance for maize comparable to Classic-period diets (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020).
Dietary and health studies of the Early and Middle Preclassic periods have been mainly descriptive due to small sample sizes and poor preservation of the human skeletal series. We provide a few examples of existing studies characterizing health and dietary indicators among the earliest Maya based on isotopic geochemical analyses and pathological lesions visible on bones and teeth. In general, early data are discussed in the context of agricultural intensification and increasing social differentiation. It should be noted that data from the small numbers of early burials are often grouped together with Late Preclassic datasets. Diachronic dietary developments have therefore been examined by assigning individuals to different periods (Preclassic versus Classic), so that discussion tends to focus on the nature of the Preclassic to Early Classic transition, while nothing can said specifically about Early or Middle Preclassic individuals. These studies generally show a trend of increasing reliance on domesticates from the Preclassic through the Postclassic, though this is not always consistent. Generalized Preclassic data from the Pasión subregion of Guatemala is available for the sites of Altar de Sacrificios (n = 9), Ceibal (n = 7), and Tamarindito (n = 1) (Wright Reference Wright2006). While there is variability between sites, with individual collagen δ 13C values ranging from –8.6‰ to –13.4‰, the Pasión dataset provides evidence for the highest level of maize consumption for the Preclassic. On the other hand, these sites exhibit some of the lowest mean δ 15N values for the Preclassic, suggesting only moderate terrestrial meat consumption (Palomo Reference Palomo2020:123–124).
Early and Middle Preclassic datapoints are relatively sparse, but present a picture of diet that points to diversity among early sedentary communities in the Belize Valley, northern Belize, the Maya Mountains, and the Pasión regions of the Lowlands, where published data are available (n = 65 individuals; Figure 2 and Table 1). The lowest δ 13C values for the Early and Middle Preclassic have been reported for the Maya Mountains region of the Lowlands. Nevertheless, the study by Kennett and colleagues (Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020) suggests that maize composed at least 70 percent of dietary protein, indicating that most lowland subregions where higher δ 13C values are reported were dominated by agricultural economies during the Early and Middle Preclassic, though with less emphasis on maize agriculture compared to the Classic period (Figure 2).
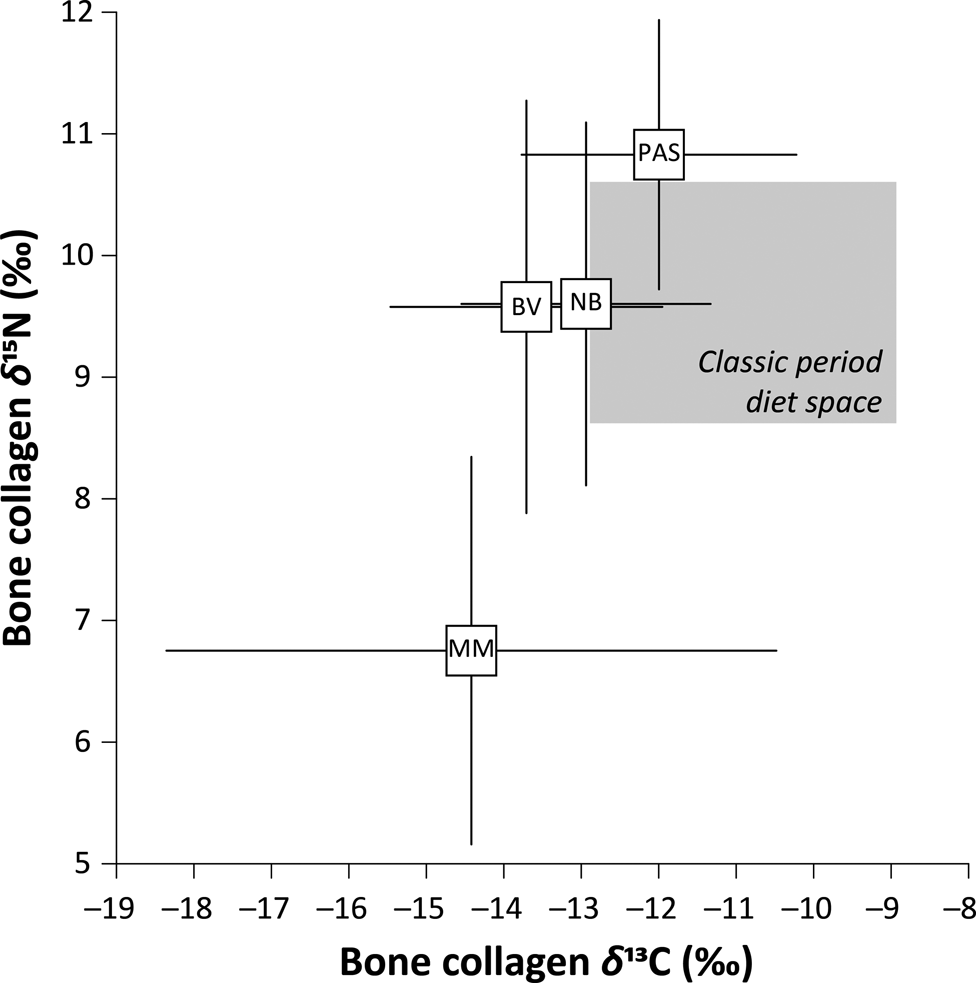
Figure 2. Bivariate plots of mean carbon (δ 13C) and nitrogen (δ 15N) isotope values for the Early and Middle Preclassic Maya lowlands presented in Table 1. Boxes represent mean values with 2σ standard deviation from the sample mean shown with error bars. BV, Belize Valley; NB, Northern Belize; PAS, Pasión; MM, Maya Mountains. Classic-period diet space after data presented by Somerville et al. Reference Somerville, Fauvelle and Froehle2013. Figure by Ebert.
Table 1. Mean δ 13C and δ 15N values from bone collagen for the Early and Middle Preclassic by lowland region. Bold indicates the summary values for each region. SD, standard deviation.
In the Belize Valley region, stable isotopic data show lower maize consumption earlier compared to northern Belize and the Pasión zones. Analyses of a small number of Middle Preclassic burials from Cahal Pech and its surrounding settlement have been subjected to isotopic analyses, with results documenting relatively low mean δ 13C and δ 15N values (–13.1‰ and +8.4‰) compared to other lowland regions (Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019, Reference Ebert, Rand, Green-Mink, Hoggarth, Freiwald, Awe, Trask, Yaeger, Kathryn Brown, Helmke, Guerra, Danforth and Kennett2021; Powis et al. Reference Powis, Stanchly, White, Healy, Awe and Longstaffe1999; see also White et al. Reference White, Pendergast, Longstaffe and Law2001). Though the sample size prohibits definitive statements about diet composition, isotopic data for Cahal Pech individuals, as well as one individual from Baking Pot (Hoggarth et al. Reference Hoggarth, Culleton, Awe and Kennett2014), generally reflect a broad diet that included the consumption of a range of domesticated plants, including maize, in addition to wild plants and animals from riparian zones along the Belize River and its tributaries.
The Pasión subregion possesses one of the largest datasets for the Middle Preclassic (n = 28), with all individuals from the site of Ceibal (Palomo Reference Palomo2020). While an average δ 13C value of –12.0 ± 1.8‰ suggests similar proportions of maize in the diet compared to the Belize Valley, a much higher mean δ 15N value +10.8‰ likely reflects more meat or freshwater foods in local diets compared to other subregions (Palomo Reference Palomo2020:120).
In northern Belize, stable isotope data for a total of 30 individuals is available from three sites (Altun Ha, n = 12, White et al. Reference White, Pendergast, Longstaffe and Law2001; Cuello, n = 14, Tykot et al. Reference Tykot, van der Merwe, Hammond and Orna1996; K'axob, n = 4, Henderson Reference Henderson2003). A mean δ 13C value of –12.9‰ for this subregion possibly suggests more investment in maize production compared to the Belize Valley, with either wild or domesticated C3 foods being less important protein sources than maize. The mean δ 15N value for northern Belize is comparable to that of the individuals from the Belize Valley, perhaps indicating that similar amounts of animal protein, likely from hunting, were consumed. Nevertheless, dietary differences are documented between northern Belize sites. K'axob, for example, has the lowest mean δ 13C and δ 15N values for northern Belize, possibly reflecting less reliance on C4 foods, as well as a subsistence economy likely dominated by terrestrial animal resources (Henderson Reference Henderson2003). At Altun Ha, isotopic analyses by White and colleagues (2001:381–382) of Middle Preclassic individuals buried in Structure C-13 also show less focus on the consumption of C4 foods compared to later (mostly Late Classic) burials at the site. On the other hand, Altun Ha has the highest δ 15N values of post-weaning individuals (n = 8; +10.6‰) for early Maya burials. Zooarchaeological data indicate that consumption of freshwater fish and fauna, and possibly marine foods, was higher compared to other northern Belize sites like Cuello and K'axob. While the authors caution that these high values may indicate that the Structure C-13 individuals came from another location closer to marine habitats, Altun Ha's close proximity (~13 km) to the Caribbean coast and barrier reef, as well as its rich marine shell assemblage (Horn et al. Reference Horn, Powis and Pendergast2020:438), indicates that marine food resources were easily accessible for the site's population. A more interesting question is perhaps why marine food declined during the Classic period.
Cuello has also seen some of the most intensive isotopic investigation of early Maya skeletal remains to date. While both δ 13C and δ 15N values have been presented as aggregate results for the combined Preclassic sample (Tykot Reference Tykot and Jakes2002; Tykot et al. Reference Tykot, van der Merwe, Hammond and Orna1996), the large numbers of Middle Preclassic individuals in the dataset warrant review. A mean bone collagen δ 13C value of −12.6‰, combined with δ 13C data from bone and enamel apatite, indicate that the Middle Preclassic population was not as dependent on maize compared to Classic period groups. Van der Merwe and colleagues (Reference van der Merwe, Tykot, Hammond, Oakberg, Ambrose and Katzenberg2002:29–30) estimated that maize accounted for between 30 and 55 percent of the Preclassic human dietary protein at Cuello, though consumption of dog and armadillo, species which potentially also consumed maize, may skew these estimates.
Within the Middle Preclassic sample at Cuello, dietary differences based on sex were also documented. While females and males consumed similar types of protein, higher δ13C values for males could suggest that C4 foods comprised a larger proportion of their diets (Tykot et al. Reference Tykot, van der Merwe, Hammond and Orna1996:359). This pattern mirrors that for the Preclassic Maya lowlands more generally, with males having higher average δ 13C values (−12.1 ± 1.5‰) compared to females (−12.8 ± 1.5‰). This difference may be associated with higher maize consumption, but has also been interpreted as protein input from marine foods (White Reference White2005). Male δ 15N values (+9.8 ± 1.2‰) are also higher than those for females (+8.9 ± 1.2‰), further suggesting increased meat/marine protein consumption (i.e., males were more carnivorous).
Skeletal markers of diet and health also shed light on the biological experience of Maya individuals. Porotic hyperostosis and cribra orbitalia, which manifest as localized areas of spongy or porous bone tissue on the surfaces of the parietals, occipital, and frontal and on the roof of the eye orbit, represent metabolic disturbances typically caused by anemia. In the Maya region, as elsewhere, signs of anemia on ancient skeletons are often attributed to dietary deficiencies related to agricultural intensification—specifically, to overreliance on maize, and iron deficiency caused by a lack of protein (Hooton Reference Hooton, Hay, Linton, Lothrop, Shapiro and Vaillant1940). However, the etiology of these lesions is far more complex than this, necessitating deep contextualization of the conditions in which they occurred. While anemia can be the direct result of poor nutrition or parasites, it can also be exacerbated when it co-occurs with other metabolic diseases, such as scurvy (White et al. Reference White, Maxwell, Dolphin, Williams and Longstaffe2006; Wright and Chew Reference Wright and Chew1998). Furthermore, the porous bone is caused by an expansion of the diploe, indicating increased red blood cell production, which Walker and colleagues (Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009) have pointed out cannot occur in situations of iron deficiency.
At Altar de Sacrificios, among the nine individuals dated to the San Felix phase (600–300 b.c.) were two infants with active porotic hyperostosis. One of the oldest human skeletons from Altar de Sacrificios (Burial 135), dated between 900 and 600 b.c. (Xe phase), is a young adult male who exhibits a healed porotic hyperostosis lesion (Saul Reference Saul1972). At Cuello, porotic hyperostosis is notably reported as being absent in the Middle Preclassic crania, though the number of observable crania is not reported (Saul and Saul Reference Saul, Saul, Whittington and Reed1997).
Linear enamel hypoplasias (LEH) are visible as narrow, sunken bands on the tooth surface that indicate a physiological stress such as illness or malnutrition (Cucina Reference Cucina2011; Hillson Reference Hillson1996; Wright Reference Wright2006; Wright and White Reference Wright and White1996). The position of the line on the tooth can be measured to estimate the timing of the event during development, and it is common to find stress events around the time of weaning (Michael et al. Reference Michael, Wrobel, Biggs, Willermet and Cucina2018). In most cases, researchers simply focus on their presence and number, relating these to differential stress loads within and between groups. At Cuello, LEHs were found in relatively high frequencies in the combined Swasey–Bladen (1200–600 b.c.; 47 percent) and Mamon (600–350 b.c.; 65 percent) phases, with females showing higher frequencies in both periods (Saul and Saul Reference Saul, Saul, Whittington and Reed1997). Saul and Saul (Reference Saul, Saul, Whittington and Reed1997:154) also argue that the Cuello population had better health and nutrition during the Middle Preclassic compared to the Late Preclassic Chicanel phase (350 b.c.–a.d. 250).
Other dental pathologies are often noted as present within Middle Preclassic populations, including caries, calculus, abscesses, antemortem tooth loss, and lingual tooth surface attrition, all of which are expected in agricultural populations. Saul (Reference Saul1972) describes the Xe phase (900–600 b.c.) individual at Altar de Sacrificios as having most of the hallmark dental pathologies of a high carbohydrate diet, including calculus formations, caries, and antemortem tooth loss, along with healed porotic hyperostosis. At Cuello, Saul and Saul (Reference Saul, Saul, Whittington and Reed1997) note fluctuations in caries rates between groups and over time, suggesting variation within the community. It is impossible to confidently attribute these to changes in specific social or environmental factors, given the very small sample sizes, the long temporal span they cover, and the wide range of ages of the individuals being studied. Instead, variations in oral health should be expected, as they are attributable to an array of factors that vary between environments, cultural groups, and individuals (Hillson Reference Hillson1996).
Other skeletal indicators of health, diet, and trauma have been reported sporadically in individuals from early sites. Saul and Saul (Reference Saul, Saul, Whittington and Reed1997) describe cases of subperiosteal hemorrhages resulting from infections and osteoarthritis at Cuello. They also report high frequencies of tibial abnormalities, including anterior-posterior bowing and cortical expansion, which they attribute to endemic syphilis. It should be noted, however, that while relatively common in other areas of the Americas (Powell and Cook Reference Powell and Cook2005), there has been very little evidence of syphilis in the Maya area prior to European contact, and the few possible cases identified thus far are most likely yaws (T. pallidum pertenue), a related treponemal disease (Wright Reference Wright2006:197). No trauma was reported among the Middle Preclassic individuals, in contrast to a variety of healed postcranial fractures and two cases of healed cranial trauma from Late Preclassic individuals. At Altar de Sacrificios, the Middle Preclassic Burial 135 had a healed transverse fracture on the left clavicle (Saul Reference Saul1972). Among the San Felix phase (600–300 b.c.) burials at Altar de Sacrificios, two adult males were found with ossified subperiosteal hemorrhages, and one elderly adult had a healed cranial lesion, all indicating some form of trauma in life.
Another study of Mesoamerican biocultural dynamics during the Preclassic period by Rodríguez (Reference Rodríguez2017) focused on comparisons of several physiological and nutritional indicators in small samples of Middle and Late Preclassic skeletons from Chiapa de Corzo and Caucel, Mexico; Ceibal and Altar de Sacrificios, Guatemala; and Barton Ramie, Belize. Rodríguez found no significant differences in porotic hyperostosis, non-specific periosteal reactions, linear enamel hypoplasias, caries, abscesses, and dental wear.
Due to the typically poor bone preservation at Early and Middle Preclassic sites, reporting of skeletal pathology data is inconsistent. Thus, lesions resulting from disease and trauma are often reported when present, but we lack appropriate frequency data to carry out any sort of comparative analyses. Instead, these data are most useful in carrying out osteobiographies, which utilize contextualized skeletal data to describe the life experiences of individuals living in the past. This approach has a long history in Mesoamerica (Romero Molina Reference Romero Molina1934, Reference Romero Molina1937; Rubín de la Borbolla Reference de la Borbolla, Daniel and Caso1969; Saul Reference Saul1972; Saul and Saul Reference Saul, Saul, İşcan and Kennedy1989) and recently has seen a resurgence of interest in the bioarchaeological community, thanks to new methodologies and theoretical frameworks that allow more complete and meaningful reconstruction and contextualization of life histories (Hosek and Robb Reference Hosek and Robb2019).
BODY MODIFICATION
As discussed above, there have been few syntheses of skeletal data that distinguish between the Early, Middle, and Late Preclassic periods. A notable exception is Tiesler's (Reference Tiesler2012, Reference Tiesler2014) work on cranial and dental modification. In her nearly exhaustive synthesis of cranial modification practices from across Mesoamerica, Tiesler discusses its ancient origins, citing previous studies that describe the presence of tabular erect modifications on six individuals from greater Mesoamerica dating to 8800–7000 years b.p. (Tiesler Reference Tiesler2014:163). Following Romano, she posits that the practice among hunter-gatherers likely stemmed from cradleboard use. In the Preclassic, with the rise of agriculture and settled villages, as well as territoriality and social stratification, a diversity of forms arose through the use of both compression cribs and head devices. Two Middle Preclassic individuals from Kaminaljuyu, for instance, showed frontal flattening that appears to have been an inadvertent consequence of tumpline use (Tiesler Reference Tiesler2014:172).
The earliest skeletal evidence of cranial modification in the Maya area comes from Middle Preclassic contexts at the sites of Cuello, Belize (Hammond et al. Reference Hammond, Saul and Saul2002) and Altar de Sacrificios (Saul Reference Saul1972); however, despite a lack of evidence for it among the few, poorly preserved remains dating to the Early Preclassic, these groups almost certainly practiced cranial modification, given its ubiquity in the region before and after that period. In general, Tiesler notes that cranial modification in the Preclassic appears to be less frequent and diverse, increasing dramatically in the Classic period because of new techniques. Though Tiesler's Maya Middle Preclassic sample is quite small (n = 17), she notes the presence of ten individuals with “pseudo-circular fronto-occipital” modifications, a style that Romano attributed to the Olmec, based on its resemblance to the forms of the anthropomorphic Olmec head sculptures and figurines (Romano Reference Romano and Paillés1980; Tiesler Reference Tiesler2010, Reference Tiesler2014). Among the 1,242 crania included in Tiesler's study, she found only ten examples of this form, all of which predate a.d. 250 and are from geographically dispersed major Preclassic political or exchange centers in the northern Yucatán, the Southern Lowlands, and Chiapas. The four Middle Preclassic examples (predating a.d. 450) come from Altar de Sacrificios (Burial 124), Ceibal (Burial 11), Caucel (Burial 1), and Pampa el Pajón (Burial 1). They have no clear association with sex or social status, and Tiesler (Reference Tiesler2014:179) argues that the presence of these forms in the Maya area, most examples of which post-date the fall of the Olmec (ca. 400 b.c.), “speaks more of syncretism and reinterpretation of Olmec heartland ideology than direct imposition.”
The earliest known dental modifications, in the form of simple filing, appear in the Maya area during the Early Bladen phase (900–800 b.c.) of the Middle Preclassic period at Cuello Belize (Saul and Saul Reference Saul, Saul, Whittington and Reed1997:45–46). Burial 11 at Ceibal, dated to the Middle Preclassic, displays Ik form (T-shaped) dental filing (Tiesler Reference Tiesler2010:296), which remains common throughout time across the Maya area. Javier Romero Molina (Reference Romero Molina1958) describes early material evidence of Maya dental practices, such as inlays in the drilled teeth of an individual who lived at the central lowland site of Uaxactún (Peten, Guatemala) during the Mamon phase (600–350 b.c.).
LIMITATIONS OF THE DATASET
A review of the dataset presented in Supplementary Table 1 reveals several impediments to creating meaningful syntheses of the available skeletal data, so it is perhaps worthwhile to include a discussion of the biases in our dataset, which will be familiar to bioarchaeologists working in the region. First and foremost, Maya burials dating to the Early and Middle Preclassic period are relatively rare. In part, this reflects biases by archaeologists towards researching later time periods, but also the fact that Preclassic contexts (and the burials contained within them) are often deeply buried beneath (and sometimes destroyed by) later Classic period (~a.d. 250–900/1000) monumental architecture, making them relatively inaccessible. As a result, excavations targeting early architecture often necessitate time-consuming documentation and removal of overlying layers, making large-scale exposure of these contexts problematic (Chase and Chase Reference Chase and Chase2006:96; Hammond et al. Reference Hammond, Clarke and Donaghey1995:121). At many sites, the earliest deposits are known only by what appears at the bottom of narrow test pits. Of the 40 sites with Early or Middle Preclassic burials, only eight had ten or more individuals. Even at sites with larger sample sizes, burials tend to come from a limited number of discrete contexts; for instance, the relatively large sample of 45 Middle Preclassic individuals from Altun Ha all come from a single non-residential structure (Pendergast Reference Pendergast1982:170). Thus, these cannot be treated as representative of a general population, but instead are likely some specific, culturally defined social subunit within the larger community.
Second, our current models of mortuary and biological variability in these early contexts are greatly biased by the limited research that is published. Our relatively small samples are spread across a significant geographic and temporal range, and this makes it difficult to generate a coherent picture of diversity among Early and Middle Preclassic communities. Of all the sites with mortuary contexts dating to the Middle Preclassic, Cuello has received by far the most attention, and for that reason our picture of the entire period is largely influenced by data from this site, as well as a few other Belizean sites, including K'axob, Santa Rita Corozal, and Cahal Pech. However, recent research at Ceibal, located in the Pasión region of Guatemala, has overturned previous models by demonstrating a much greater level of complexity, highlighting the importance of regional variation in mortuary practices, diet, and mobility (Palomo Reference Palomo2020; Palomo et al. Reference Palomo, Inomata and Triadan2017).
Third, much of the data from early skeletons is frustratingly difficult to synthesize, since interpretations are restricted by temporal associations. Analyses of skeletal remains for the Preclassic in general have rarely been combined with radiocarbon dating, impeding precise temporal assignments necessary to look at changes over time. Instead, almost all dates for these burials are based on ceramic and/or stratigraphic associations, rather than AMS radiocarbon dates. Burials are therefore lumped into ceramic phases encompassing several hundred years, preventing adequate contextualization of skeletal data.
Fourth, skeletal data for these early individuals are generally descriptive in nature, in large part due to the poor preservation that hampers consistent visual assessment of indicators of sex, age, and cranial/dental modification. Paleopathological and stable isotopic data, most often reported as summary data (e.g., averages of values, aggregates of all Preclassic individuals), have been collected over a span of many decades by investigators using a variety of different methodologies, thus making comparisons difficult or impossible. Bioarchaeologists walk a fine line, where we try to move beyond simple description of individuals by amalgamating them into groups for comparative analyses, while also attempting to maintain the social, environmental, and geographic distinctions among our samples that form the basis of anthropological inquiry. For the purposes of this review, we have discussed general trends, using examples where they are available, and focusing primarily on highlighting the significant variability present within and among these communities.
CONCLUSIONS AND FUTURE DIRECTIONS
This review identifies some intriguing themes among Early and Middle Preclassic skeletons in the Maya area. Unfortunately, it is impossible at this point to make definitive statements about the nature of biological and mortuary variability of these early groups, due in large part to a lack of chronological resolution, poor preservation, and reliance on small sample sizes that reflect diverse mortuary customs (rather than being representative of a general population). We note with great enthusiasm, however, that bioarchaeology is currently benefiting from rapid methodological advances and that several ongoing projects are actively working to aid in providing a wider range of biological data. These new data will help to overcome some of the limitations inherent in current samples and increase our ability to reach beyond the Late Preclassic to investigate the origins of the transition to social complexity. While a full review of such research within the broader field of bioarchaeology is beyond the scope of this article, we highlight a couple of examples below to demonstrate how new data will impact the study of the earliest Maya groups.
First, as AMS radiocarbon dating methods and calibration curves are continually being improved (e.g., Price et al. Reference Price, Capriles, Hoggarth, Kyle Bocinsky, Ebert and Jones2021; Reimer et al. 2020), early burial dates assigned based on archaeological associations can be tested. There is now underway a project (Maya Bone Project) aimed at directly dating large numbers of burials throughout the Maya region. Dates generated from this project and others have already helped to confirm the early associations of some burials, as well as to identify burials that were incorrectly assigned Middle Preclassic dates (e.g., Ebert and Awe Reference Ebert and Awe2021). While many early burials have no preserved collagen, thus preventing 14C dating, data generated from well-preserved burials are helping to refine ceramic chronologies based on ceramic types found in association with the skeletons, thus facilitating comparative analyses (Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019, Reference Ebert, Rand, Green-Mink, Hoggarth, Freiwald, Awe, Trask, Yaeger, Kathryn Brown, Helmke, Guerra, Danforth and Kennett2021; Hoggarth et al. Reference Hoggarth, Culleton, Awe and Kennett2014). This work will no doubt impact many of the burial dates presented in our summary table, so investigators utilizing this list in the future should be alert to subsequent research that we anticipate will result in revisions.
Another potentially transformational source of new data is ancient DNA (aDNA). As methods for extraction, sequencing, and analysis are rapidly improving, prices for this work are plummeting, allowing sampling from a greater number of individuals and more complete sequencing of individual genomes. Currently, most genomic research in the Maya area has focused on the initial peopling of the region during the Late Pleistocene and Archaic (e.g., Chatters et al. Reference Chatters, Kennett, Asmerom, Kemp, Polyak, Blank, Beddows, Reinhardt, Arroyo, Bolnick, Malhi, Culleton, Luna, Rissolo, Morell-Hart and Stafford2014; Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nägele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwängler, Haak, Harkins, Harper, Hünemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-García, Chim, Müller Plumm Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Méndez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018; Roca-Rada et al. Reference Roca-Rada, Souilmi, Teixeira and Llamas2020). The creation of large reference samples will allow comparative analyses that will explore a range of other specific questions about Maya history and social organization. There has been a number of recent critiques of aDNA research that have accompanied the hype surrounding it, and we agree that these new data are only as good as the questions we ask of them, necessitating deep contextualization provided by archaeology to examine the meaning of patterns of genomic variability (Michael et al. Reference Michael, Bengtson and Blatt2021). New aDNA research is increasingly focused on specific social questions, including how patterns of genetic relatedness correlate to specific cultural practices, such as body modifications and mortuary rituals (Verdugo et al. Reference Verdugo, Kassadjikova, Washburn, Harkins and Fehren-Schmitz2017, Reference Verdugo, Zhu, Kassadjikova, Berg, Forst, Galloway, Brady and Fehren-Schmitz2020a, Reference Verdugo, Zhu, Prout, Broomandkhoshbacht, Galloway, Fehren-Schmitz and Brady2020b), as well as exploring and mapping patterns of relatedness within and between communities that can also be used to estimate effective population sizes. Used in combination with stable isotope analyses, aDNA can help to identify patterns of migration and mobility (Freiwald Reference Freiwald, Hutson and Ardren2020). While many early individuals will not have preserved tissues suitable for DNA extraction, modeling based on data from later groups will provide valuable information about the genetic make-up of ancestral groups (González-Oliver et al. Reference González-Oliver, Pineda-Vázquez, Garfias-Morales, De La Cruz-Laina, Medrano-González, Márquez-Morfín and Ortega-Muñoz2019).
Finally, perhaps the largest issue limiting bioarchaeological investigation of these early communities is the relative paucity of early burials. However, there are several archaeological projects under way that are focusing on excavations of Early and Middle Preclassic contexts, at the sites of Xunantunich (Brown et al. Reference Brown, Awe, Garber, Brown and Bey2018), Cahal Pech (Ebert and Awe Reference Ebert and Awe2021; Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019), and Pacbitun (Powis Reference Powis, Powis, Skaggs and Micheletti2020) in the Belize River Valley; rock shelter sites in the Maya Mountains of southern Belize (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020; Prufer et al. Reference Prufer, Robinson and Kennett2020); Nixtun-Ch'ich' in Peten, Guatemala (Pugh and Rice Reference Pugh and Rice2017; Rice Reference Rice2020); various sites in the Puuc region of Mexico (Andrews et al. Reference Andrews, Wyllys, George, Gunn, Brown and Bey2018); and Ceibal in the Pasión region of Guatemala (Palomo Reference Palomo2020; Palomo et al. Reference Palomo, Inomata and Triadan2017). Guided by the preliminary studies reviewed here, these new investigations can begin to document the sources of variations in skeletal data, asking critical questions about the biocultural forces that shaped Maya civilization.
RESUMEN
Este capítulo presenta una revisión de los restos óseos maya tempranos encontrados hasta ahora. Nosotros hemos compilado una lista de más de 398 entierros mayas que fechan de los períodos preclásico temprano y medio. Estos sitios están distribuidos por toda la región maya y los datos permiten realizar síntesis descriptivas básicas sobre el comportamiento mortuorio temprano, así como aspectos de salud y dieta. La mala preservación y el análisis e informe diferente limitan severamente el alcance de la posible interpretación, pero se espera que esta revisión estimule la investigación coordinada de la biología de los grupos tempranos.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/S0956536121000456
ACKNOWLEDGMENTS
Many thanks to our generous colleagues for their help with the burial inventories and with comments on early drafts: Jaime Awe, Kat Brown, Adrian Chase, Arlen Chase, Andrea Cucina, Carolyn Freiwald, Julie Hoggarth, Aubree Marshall, Juan Manuel Palomo Mijangos, Katherine Miller Wolf, Vera Tiesler, and two anonymous reviewers.





 Open access
Open access
{kind=link}