Introduction
The Netherlands have a long tradition in the study of microvertebrates retrieved from boreholes. Antje Schreuder, the Dutch pioneer in the study of micromammals, described many finds from boreholes in different parts of the country (Schreuder, Reference Schreuder1933, Reference Schreuder1936, Reference Schreuder1941, Reference Schreuder1943). Given the low number of natural outcrops in the flat Dutch landscape, it is hardly surprising that the number of borehole sites with mammals (e.g. Mayhew, Reference Mayhew2008; Mayhew et al., Reference Mayhew, Dieleman, Boele, Verhaard and Van den Hoek Ostende2008a; Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007; Van der Meulen & Zagwijn, Reference Van der Meulen and Zagwijn1974) exceeds the number of localities that were sampled by screen-washing (e.g. Van Kolfschoten, Reference Van Kolfschoten, Fejfar and Heinrich1990a). Even though most sites yielded a few molars only, the Zuurlandboreholes set by Leen Hordijk (Hordijk, Reference Hordijk, Van Kolfschoten and De Boer1988a,Reference Hordijkb) have shown that large amounts of vertebrate material can also be sampled from the subsurface (Reumer, Reference Reumer2001; Reumer & Hordijk, Reference Reumer, Hordijk, Reumer and De Vos1999; Reumer & Van den Hoek Ostende, 2003; Van Kolfschoten, Reference Van Kolfschoten1998; Van Kolfschoten & Tesakov, Reference Van Kolfschoten, Tesakov, Van Kolfschoten and Gibbard1998). In fact, the collection of the Zuurland boreholes is the largest micromammal collection from the Dutch subsurface.
In this paper, we describe the mammals from the Moriaanshoofd borehole (Fig. 1). This borehole is unique in that it was made from a purely scientific point of interest to solve the provenance of the mammal fauna dredged from the Oosterschelde. The borehole is situated close to the deep gullies in the estuary that yielded a rich Early Pleistocene fauna (e.g. De Vos et al., Reference De Vos, Mol and Reumer1995, Reference De Vos, Mol, Reumer, Van Kolfschoten and Gibbard1998; Mayhew et al., Reference Mayhew, De Vos and Van Veen2008b; Mol et al., Reference Mol, Van den Bergh, De Vos, Haynes, Klimowicz and Reumer1999; Reumer et al., Reference Reumer, van Veen, van der Meulen, Hordijk and de Vos1998, Reference Reumer, Mayhew and van Veen2005) (Fig. 1). For about a decade the traditional annual fishing expedition (Kor en Bot; e.g. Mol & De Vos, Reference Mol and De Vos1995; De Vos et al., Reference De Vos, Mol, Reumer, Van Kolfschoten and Gibbard1998; Slupik, Reference Slupik2004) was used to collect sediment samples for the retrieval of small mammals, an enterprise that proved to be highly successful (Reumer et al., Reference Reumer, van Veen, van der Meulen, Hordijk and de Vos1998, Reference Reumer, Mayhew and van Veen2005). Notably, the micromammal assemblage was clearly a mixture of Early Pleistocene and Late Pleistocene taxa. In order to properly assess the stratigraphic position of these finds, the Moriaanshoofd borehole was made in the summer of 2008.

Fig. 1. Location of the borehole (with detailed map) (coordinates: 3°50′09″E, 51°40′50″N; RDX = 047.740; RDY = 411.380). Dredging localities of the Oosterschelde fauna: O, Olifantenputje; G, Gastenputje.
The stratigraphical sequence obtained from the borehole (Fig. 2) was described by Slupik et al. (Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013). The position of small assemblages of micromammals was indicated in that paper. Slupik et al. (Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013) explained the mixture of taxa of different ages as a result of reworking of Early and Middle Pleistocene fossils in a Late Pleistocene stream gully.
Fig. 2. Lithostratigraphic framework of the Moriaanshoofd borehole with position of the mammal assemblages. 1, sand; 2, clay; 3, peat and wood fragments; 4, clay pebbles; 5, gravel; 6, mammal fossils; 7, shells and shell fragments; depth, m b.s. = metres below surface; 14C, radiocarbon dating; lithostratigraphy: NA, Naaldwijk Formation; NI, Nieuwkoop Formation; BX, Boxtel Formation; KR, Kreftenheye Formation; KW, Koewacht Formation; MS, Maassluis Formation; Ma, age (million years); MIS, marine isotope stage; MZ (MNR1, MNR2), mammal zone cf. Tesakov (Reference Tesakov2004).
Here, we focus on the systematic description of the micromammals and how these relate to other faunas from Europe.
Geological setting
The Moriaanshoofd record is located on the Isle of Schouwen-Duiveland (Zeeland, the Netherlands) (Fig. 1), which is situated in the southern margin of the North Sea Basin on the northern flank of the Brabant massif. In this region, the strata from the Paleogene onwards are present at shallow depths and the Neogene-Quaternary period is well represented in the geological record (e.g. Van Rummelen, Reference Van Rummelen1970, Reference Van Rummelen1978; Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007; Slupik & Janse, Reference Slupik and Janse2008; Westerhoff et al., Reference Westerhoff, Wong, De Mulder, De Mulder, Geluk, Rietsema, Westerhoff and Wong2003). The region is located in a transitional area between thick and complete Neogene and Early Quaternary successions to the north (e.g. Kuhlmann, Reference Kuhlmann2004; Kuhlmann et al., Reference Kuhlmann, Langereis, Munsterman, Van Leeuwen, Verreussel, Meulenkamp and Wong2006a,Reference Kuhlmann, Langereis, Munsterman, Van Leeuwen, Verreussel, Meulenkamp and Wongb) and incomplete, often eroded successions towards the south (De Schepper et al., Reference De Schepper, Head and Louwye2004, Reference De Schepper, Head and Louwye2008; Vandenberghe et al., Reference Vandenberghe, Laga, Steurbaut, Hardenbol, Vail, De Graciansky, Hardenbol, Jacquin and Vail1998, Reference Vandenberghe, Van Simaeys, Steurbaut, Jagt and Felder2004).
During Late Cenozoic times, the North Sea Basin was part of the northwest European Basin that covered the present-day offshore and part of onshore Netherlands, Germany and Denmark. The basin was confined by landmasses in the south and east, and the landmasses of England and Scotland in the west. Towards the northwest, it was connected to the Atlantic (Ziegler, Reference Ziegler1990) and short-lived connections through the English Channel occurred. A large clastic delta system prograded progressively into the North Sea Basin from the east; sediments originated mostly from the rivers draining the Fennoscandian Shield and the Baltic Platform (Bijlsma, Reference Bijlsma1981; Overeem et al., Reference Overeem, Weltje, Bischop-Kay and Kroonenberg2001; Kuhlmann, Reference Kuhlmann2004; Kuhlmann et al., Reference Kuhlmann, Langereis, Munsterman, Van Leeuwen, Verreussel, Meulenkamp and Wong2006a,Reference Kuhlmann, Langereis, Munsterman, Van Leeuwen, Verreussel, Meulenkamp and Wongb). Coeval subsidence created considerable accommodation space that became filled with these siliciclastic deposits, reaching a thickness of approximately 3000 m in the depocentre (Ziegler, Reference Ziegler1990; Kuhlmann, Reference Kuhlmann2004). The successions in the North Sea Basin have well-preserved signals of climatic variations and basin development. During the Neogene Quaternary, the climate in the region deteriorated from warm temperate–subtropical to the strong glacial–interglacial cycles, whose onset is estimated at c. 2.58 Ma (lower boundary of Gelasian, Early Pleistocene). Both the intensification of the associated glacioeustatic sea-level dynamics, as well as changing vegetation patterns in the hinterland drastically changed and shaped the rapid infill of the southern North Sea Basin during the Neogene and Quaternary (see Kuhlmann, Reference Kuhlmann2004 for a comprehensive approach).
The sedimentary successions towards the central parts of the basin are pretty complete and cover very long periods of time (Kuhlmann, Reference Kuhlmann2004). At the margins of a basin, the situation is different. During the sea-level high-stands, the marginal parts were under marine conditions and marine deposits were formed. During sea-level drops (due to glacio-eustacy and/or tectonics) regression followed in the marginal parts, and terrestrial conditions prevailed with, in consequence, reworking and erosion of earlier deposited sediments and/or terrestrial deposition. As a result, the successions at the margins of the sedimentary basin are less complete. Only some periods of time are reflected in the record and a number of stratigraphic hiatuses occur.
During the Neogene, marine conditions prevailed at the southern margin of the North Sea Basin in the Netherlands and a thick succession of deposits was formed. The Neogene is rather well represented in the record. However, some hiatuses are present (Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007).
During the Pleistocene, climate fluctuated significantly, which is reflected in the sedimentary record. Five major depositional environments can be distinguished that occur in repetitive successions in the subsurface of the southwest Netherlands: marine, coastal, glacial, fluvial and local terrestrial. The associated sediments are usually separated by hiatuses (see Rijsdijk et al., Reference Rijsdijk, Passchier, Weerts, Laban, Van Leeuwen and Ebbing2005 for a general approach and Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007 for a case study).
During the Early Pleistocene, marine conditions prevailed in the region, and marine sediments of the Maassluis Formation were formed. These deposits are overlain by the Late Pleistocene fluvial, terrestrial and fluvial/glacial deposits. That points to a major hiatus of about 1.8–2.0 my separating the Early Pleistocene and the Late Pleistocene successions (e.g. Van Rummelen, Reference Van Rummelen1970, Reference Van Rummelen1978; Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007, Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013). The top part of the Quaternary succession is formed by marine deposits of Holocene age. Marine deposits related to the Eemian (Early Late Pleistocene) transgression were first detected in the region by Van Rummelen (Reference Van Rummelen1970, Reference Van Rummelen1978), but this attribution was questioned by Slupik et al. (Reference Slupik, Wesselingh, Janse and Reumer2007).
The record at Moriaanshoofd comprises a 56-m thick (6.00–62.00 metres below ground level (mbgl)) complex of siliciclastic deposits representing a part of the infill of the North Sea Basin in its southernmost, marginal part during the Quaternary. The top part of the record (0.00–6.00 mbgl) is of anthropogenic origin (dike supplementation) and is not considered here.
Deposits are assigned to six formations (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013), which are from the bottom up: Maassluis Fm., Koewacht Fm., Kreftenheye Fm., Boxtel Fm., Nieuwkoop Fm. and the Naaldwijk Fm. (see TNO, 2013 for definitions of the lithostratigraphic units) (Fig. 2). Here, the local expression of the lithostratigraphic units are shortly characterised. For a comprehensive approach see Slupik et al. (Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013).
The Maassluis Formation consists of sands and silty sand with some clayey partings. In some intervals, shells and shell fragments are common and mostly strongly worn, while other intervals are barren. The Maassluis Formation is interpreted as originating in a shallow marine setting during the Gelasian (Early Pleistocene). The bottom part of the formation is not reached in the borehole.
The Koewacht Formation consists of coarse sands with gravel, abundant indurated clay pebbles and claystone clasts in the lower part. Shells, mostly well preserved, are common to very abundant. It is interpreted as being of fluvial origin and related to the Schelde river system. The basal gravelly part is interpreted as lag deposit. The Koewacht Formation is of late Late Pleistocene age. There is a major hiatus between the Maassluis Fm. and the Koewacht Fm. comprising c. 2.2 my.
The Kreftenheye Formation consists of coarse sands containing shell fragments. In the lower part, fine gravel and clay pebbles occur and the shell fragments are common. In the upper part, shell content is very low. The formation is considered to be of fluvial origin and is related to the Rhine river system. The Kreftenheye Formation is of late Late Pleistocene age.
The Boxtel Formation consists of coarse to fine sands. Some peat fragments and wood pieces occur. Shells are lacking. The deposits represent local small-scale fluvial and/or fluvio-aeolian deposition in a cold climate setting. The formation is of late Late Pleistocene age.
The Naaldwijk/Nieuwkoop Formations are of Holocene age. The Naaldwijk Formation consists of sands, sands with clayey partings and clays. The Nieuwkoop Formation consists of peat. Deposits of the Naaldwijk Formation are considered to have originated in a shallow marine setting. The formation forms the Holocene clastic coastal infill in the western Netherlands.
The origin of the Naaldwijk Formation is related to the Holocene transgression that postdated the last glacial period of the Late Pleistocene (for a detailed approach see Hijma et al., Reference Hijma, Cohen, Hoffmann, Van der Spek and Stouthamer2009, Reference Hijma, Van der Spek and Van Heteren2010; Hijma & Cohen, Reference Hijma and Cohen2010, and for paleogeographic reconstructions see Vos et al., Reference Vos, Bazelmans, Weerts and Van der Meulen2011). The Naaldwijk Formation intertongues with peat layers (Nieuwkoop Formation) that formed mainly on the landward side of the coastal plain (Weerts & Busschers, Reference Weerts and Busschers2003).
Reworking played an important role in the genesis of the stratigraphic successions in the southwestern Netherlands. Evidence for substantial reworking of older sediments and fossils into the basal parts of formations is found in the region (Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007, Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013). Reworking of older sediments and fossils during transgression, in shallow agitated coastal areas or in tidal-channel floor settings is a common phenomenon in shallow basin margins such as the southern North Sea Basin. Because of reworking, the lithoclasts and fauna elements of older lithostratigraphic units predominate in the basal parts of the overlying units. As a result, there are uncertainties and difficulties in defining lithostratigraphic boundaries and interpretation of the nature and provenance of fossil assemblages can be problematic (see Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007, Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013).
In the record at Moriaanshoofd reworking of sediments is obvious (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013). For example, the record of the fluvial Koewacht Formation that directly overlies the Maassluis Formation indicates a southern (i.e. western Belgium and northern France) origin of the material and fauna, which were deposited by the River Schelde, and also locally reworked, possibly fluvial deposits (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013).
Assemblages of small mammal remains (terrestrial animals) studied here derived from intervals in the marine Maassluis Formation and the fluvial Koewacht Formation. In particular, the basal channel lag in the latter contains a considerable concentration of bone material. The mammal assemblages are embedded in a geological succession that has an extraordinary complexity in terms of lithostratigraphy and the variety of processes that played a role in shaping the region during the Quaternary.
Material and methods
The borehole is located south of the hamlet of Moriaanshoofd on the island of Schouwen-Duiveland (Zeeland, southwest Netherlands). The borehole was located adjacent to the Oosterschelde dike south of Moriaanshoofd and reached an end depth of 62 m (TNO – Geological Survey of the Netherlands borehole number B42G0769). The surface elevation at the borehole location was 4.19 m +N.A.P. (Dutch Ordnance Level, ∼ mean sea level). Coring was performed in half-metre intervals. From each interval, about half a kilogram of raw sediment was taken for lithological description and analyses. Additionally, larger samples of approximately 2–3 kg of sediment were taken at 1-m intervals, washed and sieved (sieve mesh 1 mm), and analysed for fossil content. Fossils were picked from wash residues and identified. Molluscs were counted. All sediment samples were visually and microscopically examined.
Some sediment samples yielded teeth of small vertebrates. A total of 38 teeth and tooth fragments, and one identifiable postcranial bone were found. All tooth samples were microscopically studied using Leica 16Z stereomicroscope and associated software. The nomenclature of dental details of the arvicolid teeth is after Van der Meulen (Reference Van der Meulen1973), Rabeder (Reference Rabeder1981) and Tesakov (Reference Tesakov2004). The wear surface was measured following procedures outunderlined in Van der Meulen (Reference Van der Meulen1973) and the measurements of the enamelfree areas and the taxonomically identification follow Tesakov (Reference Tesakov2004).
All measurements are in millimetres. The following measurements were taken (as far as possible) by DFM and FED:
lower molars: AL = length of anterior loop measured from the anterior of tooth to the anterior point of the second labial re-entrant angle; L = length of wear surface; W = width of wear surface; CH = crown height from wear surface to base of enamel; ASD = height of anterior enamel free area (anterosinuid); HSD = height of buccal enamel free area (hyposinuid); HSLD = height of lingual enamel free area (hyposinulid).
upper molars: L = length of wear surface; W = width of wear surface; CH = crown height from wear surface to base of enamel; ASL = height of anterior/lingual enamel free area in M1 (anterosinulus); PRS = height of lingual enamel free area (protosinus); AS = height of buccal enamel free zone (anterosinus); DS = height of posterior enamel free zone (distosinus).
In addition, the following ratios were calculated: HH index = √(HSD² + HSLD²); PA index = √(PRS² + AS²) (Rabeder, Reference Rabeder1981). Abbreviations: BSA = buccal salient angle; LRA = lingual re-entrent angle; BRA = buccal re-entrent angle, T = dentine triangle.
Drawings (by JWFR) were made on a Wild M5 binocular microscope fitted with a drawing prism.
The sediment samples and mammal remains of the Moriaanshoofd borehole are archived at the Natuurhistorisch Museum Rotterdam (NMR) and the molluscs are archived at the Naturalis Biodiversity Center in Leiden. Six radiocarbon datings on peat and wood fragments were performed in order to obtain age indications of the deposits (see Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, Van Strydonck, Kiden, Burger and Reumer2013).
The lithostratigraphic terminology follows Westerhoff et al. (Reference Westerhoff, Wong, De Mulder, De Mulder, Geluk, Rietsema, Westerhoff and Wong2003) and TNO (2013), and the chronostratigraphy is according to definitions in Gibbard and Head (Reference Gibbard and Head2009a,Reference Gibbard and Headb), Gibbard et al. (Reference Gibbard, Head and Walker2010) and Cohen et al. (Reference Cohen, Finney, Gibbard and Fan2013). All depths are given below the ground surface (which is at 4.19 m +N.A.P.).
Results
Table 1 gives a summary of results and Table 2 shows the measurements taken.
Table 1 Summary results – occurrence of small mammal taxa in the record.
Table 2 Measurements of mammal specimens (see Material and methods) NMR, Natural History Museum Rotterdam; CH, crown height; ASD, anterosinuid; HSD, hyposinuid; HSLD, hyposinulid; AS, anterosinus; ASL, anterosinulus; PRS, protosinus; DS, distosinus; HH index, √(HSD² + HSLD²); PA index, √(PRS² + AS²). For details on terms see Van der Meulen (Reference Van der Meulen1973), Rabeder (Reference Rabeder1981) and Tesakov (Reference Tesakov2004).
Systematic section
Insectivora
Talpidae
Desmana sp.
Material
36–37 m: tibio-fibula fragment distal (NMR999100007365 [hereafter NMR7365])Footnote 1
Soricidae
Sorex sp.
Material
35–36 m: upper first left incisor (NMR7405)
Rodentia
Arvicolidae
Mimomys Forsyth Major, 1902
Mimomys pliocaenicus Forsyth Major, 1902 and Mimomys praepliocaenicus Rabeder, Reference Rabeder1981 (Figs 3 and 4)
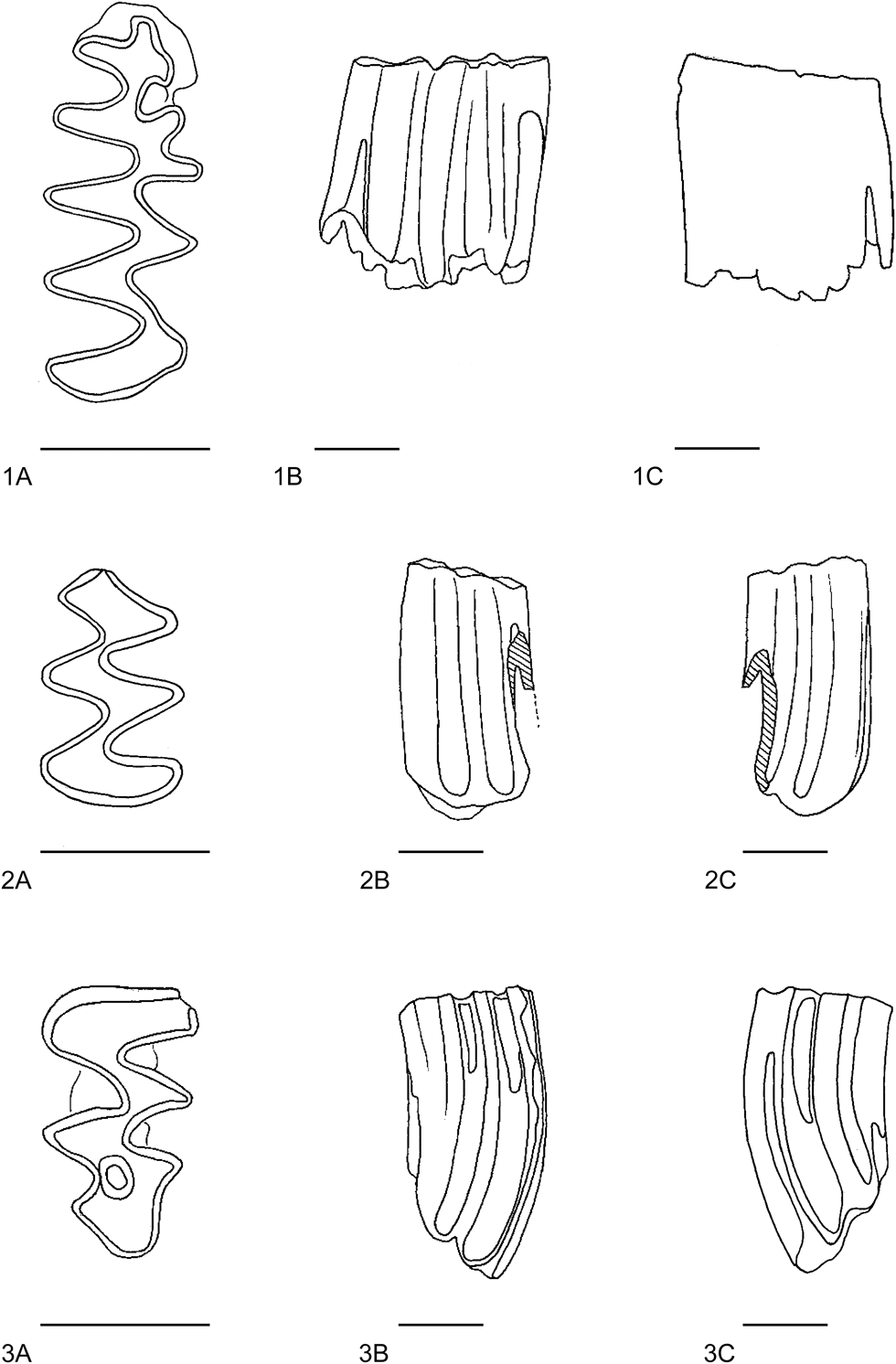
Fig. 3. 1, Mimomys praepliocaenicus M3 sin (NMR7370): A, occlusal view; B, labial view; C, lingual view; 2, Mimomys praepliocaenicus M3 dex (NMR7373): A, occlusal view; B, labial view; 3, Mimomys praepliocaenicus M3 dex (NMR7375) occlusal view, damaged; 4, Mimomys praepliocaenicus M3 sin (NMR7378): A, occlusal view; B, labial view; C, lingual view. Scale bar is 1 mm.
Fig. 4. 1, Mimomys praepliocaenicus M2 dex (NMR7374): A, occlusal view; B, labial view; C, lingual view; 2, Mimomys praepliocaenicus M2 dex (NMR7377): A, occlusal view; B, labial view; C, lingual view; 3, Mimomys praepliocaenicus m1 dex (NMR7385) occlusal view. Scale bar is 1 mm.
Material
36–37 m: M1 (NMR7380), M2 (NMR7379), M2 (NMR7371), M3 (NMR7370), m2 (NMR7372)
54–55 m: M1 (NMR7381), M2 (NMR7374), M3 (NMR7373), M3 (NMR7382), M3 (NMR7375)
55–56 m: M1 (NMR7387), M2 (NMR7376), M2 (NMR7383), M2 (NMR7377), M3 frag (NMR7386), m1 (NMR7385), m1 frag (NMR7384)
56-57 m: M3 (NMR7388), m2 (NMR7389)
37–62 m: M3 (NMR7378)
Measurements: Table 2
M2 PA index: Table 3
Table 3 Mimomys pliocaenicus/Mimomys praepliocaenicus M2 PA index = √(PRS2 + AS2).
M3 PA index: Table 4
Table 4 Mimomys pliocaenicus/Mimomys praepliocaenicus M3 PA index = √(PRS2 + AS2).
Comment
The majority of the arvicolid remains from the Moriaanshoofd borehole appear to belong to a large Mimomys species. They are characterised by large size, moderate to abundant crown cement and clearly differentiated enamel. However, the range of specific characters found in the assemblage, in particular the development of the enamel-free areas, is most probably greater than can be attributed to one species. These teeth are therefore treated as representing at least two later stages of evolution in the lineage M. hajnackensis/M. polonicus/M. praepliocaenicus/M. pliocaenicus (Tesakov, Reference Tesakov1998, Reference Tesakov2004). Certain teeth can be attributed with confidence to one of the latter two of these species. Others are too damaged or are worn so that the enamel-free area height cannot be determined: these are recorded as Mimomys pliocaenicus/praepliocaenicus.
Description
The three upper M1 are similar in size and morphology. They all have three roots. All enamel-free areas extend to the wear surface. The upper M2s (NMR7374 and NMR7377) have closed enamel-free areas (PA index = 4.5, 4.7). NMR7377 has three roots. M2 (NMR7383) is relatively large and has two roots, and the enamel-free areas extend to the wear surface. Four upper M3 (NMR7370, NMR7373, NMR7375 and NMR7378) have the enamel-free areas still closed and measurable, with PA indices 3.9, 3.3, 2.9 and 3.2.
The lower m1 (NMR7385) is moderately worn (crown height = 3.19 mm), and an enamel islet is still present. The BSA3 is prominent and posteriorly directed, nearly isolated as a fourth triangle.
Discussion
This assemblage has characteristics which span the species M. praepliocaenicus Rabeder, Reference Rabeder1981 and M. pliocaenicus Forsyth Major, 1902 as defined by Tesakov (Reference Tesakov1998, Reference Tesakov2004). The occurrence of three roots rather than two roots in M1 and M2 is a primitive character state but does not uniquely identify to species. The values of the PA index of the M2 from the lower levels of Moriaanshoofd (54–56 m) (4.5, 4.7) are outside the range of those of M. pliocaenicus from Tegelen given by Tesakov (Reference Tesakov2004). They appear to be referable to M. praepliocaenicus, slightly more advanced than material from Kryzhanovka 3, Ukraine, referred by Tesakov to M. praepliocaenicus and identical to material from the upper Norwich crag at Easton Wood. The M2 (NMR7371) from the higher level 36–37 m has an anterosinuid measuring 3.86 mm, which is much larger than in material assigned to M. praepliocaenicus and close to the lower limit of the range for M. pliocaenicus from Tegelen (Tesakov, Reference Tesakov1998, Reference Tesakov2004; Rabeder, Reference Rabeder1981).
Similarly, the M3 from the deeper levels of the borehole (NMR7373, NMR7378, NMR7375; below 37 m) have lower PA indices (2.9, 3.2, 3.3) that are, however, higher and thus more advanced than M. praepliocaenicus from Kryzhanovka 3, Ukraine (Tesakov, Reference Tesakov2004), but too low crowned for M. pliocaenicus. The M3 from the higher level (NMR7370, 36–37 m) has a higher PA index (3.9) that falls in the range of Tegelen M. pliocaenicus and outside the range of M. praepliocaenicus. Material from the upper Norwich crag at Easton Wood is identical to that from Moriaanshoofd deeper levels.
If we examine the distribution of identifiable specimens in relation to depth, the results are as follows. Specimens at depth below 37 m (see Table 1) are all referable to M. praepliocaenicus or else not identifiable to species level. Specimens above the top of the Maassluis Formation in the basal channel lag layer are attributable to M. pliocaenicus or not identifiable to species level. This is in line with the interpretation that the Maassluis Formation sediments were deposited at a time before the arrival of Mimomys pliocaenicus. However, all the material appears reworked. This is certainly the case for material retrieved from 36–37 m depth, where teeth are found together with different types of preservation, so such interpretations need to be made with caution.
One of the M3 identified as M. praepliocaenicus (NMR7378) is of interest in lacking an islet, whereas this is normally present in this species. However, it cannot be excluded that the tooth is a variant, as similar teeth have been figured by Tesakov (Reference Tesakov2004) and are said to be present in assemblages of earlier species of the lineage such as Mimomys hajnackensis (Tesakov, pers. comm.). The occurrence of M3s lacking an enamel islet (a derived character) has also been used to group together a number of medium and larger sized species as Cromeromys Zazhigin, 1980. M3 with a similar morphology occur in the Zuurland borehole (undescribed material, Mayhew, pers. obs.) at a depth of c. 92–93 m. An M3 lacking an islet, from the Norwich Crag of Covehithe, Suffolk, UK, similar in general size and morphology to NMR7378 but with higher enamel-free areas, was shown in Mayhew and Stuart (Reference Mayhew and Stuart1986) as Mimomys sp. It remains to be seen if this represents individual variability or a phylogenetic relationship.
Mimomys reidi Hinton, 1910
Fig. 5, parts 1 and 2.
Fig. 5. 1, Mimomys reidi m1 dex (MNR7390): A, occlusal view; B, labial view; C, lingual view; 2, Mimomys reidi m2 sin (MNR7402): A, occlusal view; B, labial view; C, lingual view; 3, Mimomys tigliensis M3 sin (NMR7399): A, occlusal view; B, labial view; C, lingual view. Scale bar is 1 mm.
Material
36–37 m: m1 (MNR7390), m2 (MNR7402)
54–55 m: m1 frag (NMR7391), m1 frag. (NMR7392)
55–56 m: m2 (NMR7403)
57–58 m: m2 (NMR7404)
Measurements: Table 2
Description
The m1 MNR7390 (Fig. 5, parts 1a, 1b and 1c) is the unrooted crown of a juvenile tooth. The islet is visible, but not yet fully isolated. The enamel-free areas are all closed. A clear Mimomys ridge runs anteriorly towards the base of the crown. NMR7391 represents part of the anterior loop of a small arvicolid. Lower m2 (MNR7402) is rooted, the hyposinulid and hyposinuid are closed, the anterosinuid is open at the wear surface (HH index = 2.8). T1 and T2, and T3 and T4 are broadly confluent. The enamel is relatively thin and undifferentiated. NMR7403 consists of the anterior half of an m2. The anterosinuid is open, the enamel thin and undifferentiated. NMR7404 consists of the anterior fragment of an m2, with open anterosinuid and thin enamel.
Discussion
MNR7390 demonstrates the presence of a small Mimomys species with enamel islet. It can be matched by specimens of M. reidi from the Tegelen assemblage. The m2s seem to differ from material of M. reidi from Tegelen in their smaller size and straighter margins to the triangles. However, they are similar in this respect to material referred to M. reidi found in the Norwich crag (Mayhew, unpublished). The m2 differs from the similar sized Clethrionomys kretzoii in having greater confluence of T1 and T2, and straighter margins of the triangles. Compared with material of P. pitymyoides from the type locality Osztramos-3, the m2 (MNR7402) is of smaller size and lower crown height, and more curved laterally. Mimomys reidi is a widely distributed Gelasian arvicolid species.
Mimomys tigliensis Tesakov, Reference Tesakov1998
Fig. 5, parts 3a, 3b and 3c, and Fig. 6, parts 1, 2 and 3.
Fig. 6. Occlusal views: 1, Mimomys tigliensis m1 sin (NMR7396); 2, Mimomys tigliensis m1 dex (NMR7397); 3, Mimomys tigliensis m1 dex (NMR7400); 4, Pitymimomys sp. M1 sin (NMR7401); 5, Microtus sp. M3 dex (NMR7369); 6, Microtus agrestis M1 dex (NMR7368). Scale bar is 1 mm.
Material
36–37 m: M1 frag (NMR7395), M2 (NMR7393), M3 (NMR7399), m1 (NMR7396), m1 (NMR7397), m2 (NMR7398), m3 (NMR7394)
54–55 m: m1 (NMR7400)
Measurements: Table 2
Description
M1 (NMR7395) is rooted and lacks the anterior loop. The triangles are closed, with very thick enamel on the anterior margins and the re-entrant folds filled with crown cement. The M2 (NMR7393) is high crowned, and all enamel-free areas are open at the wear surface. There is a moderate amount of crown cement. The triangles are only slightly confluent. The M3 (NMR7399) has closed roots, the parasinus and distosinus are closed, and the anterosinus is open at the wear surface. There is abundant crown cement in the buccal re-entrants. An islet is present.
The lower m1s (NMR7396 and NMR7397, Fig. 6, parts 1 and 2) are rooted, T2 is small and there is abundant crown cement also in the folds of the anterior cap (LRA4, BRA3). The m2 (NMR7398) is rather worn. The enamel is thick and the cement abundant. The enamel is clearly thicker on the posterior margins of the triangles.
The lower first molar (NMR7400) is from a young animal in which the roots are not fully formed. All enamel-free areas are open to the wear surface. The tooth is slightly curved laterally. The dentine is partly dissolved, especially in the anterior loop. There is no trace of an enamel islet or its former position. The triangles are closed and there is abundant cement. The enamel is clearly differentiated, thicker on the posterior sides of the triangles. T2 is relatively small compared with T1 and T3. The anterior cap is relatively compressed. This tooth appears more rolled than those from the higher levels 36–37 m.
Discussion
These teeth belong to a small-sized rooted vole species identified as Mimomys tigliensis Tesakov, Reference Tesakov1998. The enamel differentiation, abundant crown cement and absence of islet in m1 are characteristic. The morphology and size of the specimens fall within the range of the material from Tegelen described by Tesakov (Reference Tesakov1998). This is a widely distributed species previously mentioned in much of the older literature under the name Mimomys newtoni (name not available, see Mayhew and Stuart, Reference Mayhew and Stuart1986).
Mimomys tigliensis appears to have arrived in Western Europe by immigration in the later part of the Villanyian stage, since no species which could be ancestral are known from earlier deposits in this area. This immigration event is potentially important for biostratigraphy (Mayhew, Reference Mayhew2013). In the Stranzendorf loess profile of Austria, the species appears for the first time at the level Stranzendorf K (Mayhew, pers. obs.). In the faunas of eastern Europe, they appear for the first time at Liventsovka X placed by Tesakov (Reference Tesakov2004) at the upper boundary of zone MNR2; estimate absolute age c. 2.25 Ma. In the UK, M. tigliensis arrives (and is rare) in the upper Norwich Crag faunas, similar in age to Liventsovka X (Mayhew, Reference Mayhew2013).
The Mimomys tigliensis/Mimomys tornensis clade immigration event in biozone MNR2 (2.25–2.45 Ma, event probably towards latest part of the zone) indicates the earliest possible date for the Maassluis Formation deposits at 54–55 m in the Moriaanshoofd borehole. However, a younger age is possible in view of the reworking of material evidenced by the preservation and facies (a marine formation). These deposits are now considered to be of similar age to the Norwich Crag (age 2.25–2.35 Ma) underlying the Baventian clays of the Easton and Covehithe areas on the Suffolk, UK, coast (Mayhew, Reference Mayhew2013).
Pitymimomys Janossy & Van der Meulen, 1975
Pitymimomys sp.
Fig. 6, part 4.
Material
36–37 m: M1 (NMR7401)
Measurements: Table 2
Description
The M1 (NMR7401) is of small size, with a large anterior and smaller posterior root. The enamel-free areas are open. There is a small amount of crown cement in the folds. The triangles T1 and T2, T3 and T4 are confluent. The anterior margin of BSA2 is straight. The enamel appears thicker on the anterior margins of the triangles.
Discussion
This tooth is assigned to the small-sized rooted vole genus Pitymimomys in view of its characteristic enamel pattern. It resembles material of P. pitymyoides from the type locality Osztramos-3, Hungary, but it is slightly smaller.
The presence of Pitymimomys in the Moriaanshoofd borehole is not surprising since the species was previously recorded from Tegelen (a single tooth, Tesakov, Reference Tesakov1998) and Zuurland 63–66 m (Van Kolfschoten, Reference Van Kolfschoten, Van Kolfschoten and De Boer1988), as well as from the UK crags (Mayhew & Stuart, Reference Mayhew and Stuart1986). Pitymimomys disappears in the early Pleistocene soon after the arrival of rootless voles of the genus Allophaiomys (Tesakov, Reference Tesakov2004).
Microtus Schrank, 1798
Microtus sp.
Fig. 6, part 5.
Material
33–34 m: M3 (NMR7369)
Measurements: Table 2
Description
The upper M3 is rolled and damaged. The tooth is highly asymmetrical in shape, with three deep lingual re-entrants with abundant cement.
Discussion
As very similar complicated variants of the M3 are found in several different species of Microtus, including M. agrestis, this tooth cannot be identified to species level.
Microtus agrestis Linnaeus, 1761
Fig. 6, part 6.
Material
34–35 m M1 (NMR7368)
Measurements: Table 2
Description
This M1 consists of four triangles and an obliquely placed anterior cap with abundant cement. The posterior triangle has an additional appendage with a cement filled re-entrant angle.
Discussion
An appendage, called an agrestis loop, on the distal end of the M2 is usually regarded as diagnostic for Microtus agrestis. In Dutch M. agrestis extant populations 99% of M2s bear this feature (Dienske, Reference Dienske1969). The M1 of M. agrestis can also have a distal appendage, called the exsul loop, after the subspecies Microtus agrestis exsul Miller, 1908 from the Hebrides (UK). When present, an exsul loop on the M1 is also diagnostic for M. agrestis, but this presence in M. agrestis populations varies geographically from 0.3% (Schleswig-Holstein) to 100% (Islay, Hebrides, UK; North Ural, Russia) (Niethammer & Krapp, Reference Niethammer and Krapp1982). An exsul loop is present in 13% of M1 of Dutch extant populations (Dienske, Reference Dienske1969). The earliest fossil finds of M. agrestis in the Netherlands are described from three localities of early Saalian/Holsteinian age: Maastricht Belvédère 4 (Van Kolfschoten, Reference Van Kolfschoten1985), Wageningen Fransche Kamp and Leccius de Ridder near Rhenen (Van Kolfschoten, Reference Van Kolfschoten1981, Reference Van Kolfschoten1990b). The Microtus agrestis finds from Maastricht Belvédère 4 are of particular interest because the percentage of M1s with an exsul loop is twice as high as in Dutch extant populations, suggesting that the incidence of this feature was higher in populations from the late Middle Pleistocene. Recently a M1 with an exsul loop was also found in the Oosterschelde dredgings (Dieleman, pers. obs., 2013). It is unlikely that this molar originates from a recent specimen because extant Microtus agrestis populations are not recorded in the area (Bekker, Reference Bekker, Calle, Dobbelaar, Fortuin, Jacobusse and De Kraker2010).
General discussion and conclusions
The arvicolid remains from the Maassluis Formation marine sediments extending downwards below the base of the channel lag at c. 37 m represent a Gelasian assemblage with several Mimomys species, including Mimomys praepliocaenicus, Mimomys reidi and Mimomys tigliensis. In particular the occurrence of Mimomys tigliensis at 55 m is of interest as this species is now considered (Mayhew & Stuart, Reference Mayhew and Stuart1986; Mayhew pers. obs., 2010) to have immigrated into Western Europe during the time of formation of the upper Norwich Crag of the UK (age c. 2.25–2.35 Ma) (Mayhew, Reference Mayhew2013, submitted) and before the Weybourne Crag (age c. 2.1–2.25 Ma) (Mayhew, submitted). The complete absence of the marine bivalve Macoma balthica below the level of 37 m in the Moriaanshoofd borehole might indicate that these deposits antedate the level of 63 m in the Zuurland borehole, where this mollusc is first recorded (Meijer, Reference Meijer, Van Kolfschoten and De Boer1988), although shell material below this depth is scarce at Zuurland and Moriaanshoofd. This may support definition of a biostratigraphic stage in the North Sea Basin area characterised by the presence of the vole Mimomys tigliensis and absence of the marine mollusc Macoma balthica. Both of these species are abundantly present in the UK Weybourne Crag assemblages, such as Sidestrand, West Runton Crag and East Runton (Mayhew & Stuart, Reference Mayhew and Stuart1986) and Weybourne (Mayhew & Gibbard, Reference Mayhew and Gibbard1998).
The arvicolid assemblage from the channel lag deposits (depth 35 to c. 37 m) resting on the Maassluis Formation is of interest in consisting only of rooted arvicolids similar to the underlying Gelasian assemblage, but with the presence also of Mimomys pliocaenicus. Some specimens have a relatively fresh unrolled and less mineralised appearance. This suggests an origin from Early Pleistocene freshwater sediments, indicating reworking and redeposition of material from elsewhere. No typical Middle or Late Pleistocene vole species were recorded in these layers.
The deposits from 33 to 35 m, i.e. above the derived Gelasian assemblage mentioned above, yielded only two teeth, both of the genus Microtus: the morphology of these teeth indicates a late Middle Pleistocene, Late Pleistocene or Holocene age.
Local reworking of material from Early (and Middle) Pleistocene suggested by the coarse lag deposits above c. 37 m could explain recovery of a Trogontherium tibial fragment of large size, suggesting a Middle Pleistocene age, from the deep channel of the Oosterschelde, close to the Moriaanshoofd borehole position (Mayhew et al., Reference Mayhew, Dieleman, Boele, Verhaard and Van den Hoek Ostende2008a). Another find from the Oosterschelde dredgings concerns an M2 from Arvicola terrestris with undifferentiated enamel (Dieleman, pers. obs.), suggesting a late Middle Pleistocene age for this molar.
Compared to other localities in the region, the Moriaanshoofd assemblage from the lower part (54–56 m) of the borehole appears similar to Zuurland level 11 (Tesakov, Reference Tesakov1998; Van Kolfschoten, Reference Van Kolfschoten, Van Kolfschoten and De Boer1988), the Oosterschelde material described by Reumer et al. (Reference Reumer, van Veen, van der Meulen, Hordijk and de Vos1998, Reference Reumer, Mayhew and van Veen2005) and the lower levels of the Schelphoek borehole (Slupik et al., Reference Slupik, Wesselingh, Janse and Reumer2007). The Moriaanshoofd material from the Maassluis Formation is certainly older than the Tegelen material (age c. 2.0–2.2 Ma) described by Tesakov (Reference Tesakov1998). Compared with the UK crags, it appears to be equivalent to the younger part of the Norwich crags and older than the Weybourne Crag.
In terms of other biostratigraphic systems, the Moriaanshoofd Maassluis Formation assemblage represents Late Villanyian (in the sense of Repenning et al., Reference Repenning, Fejfar, Heinrich, Fejfar and Heinrich1990), Zone MN 17 the later part of Zone MNR 2 (Tesakov, Reference Tesakov2004), with an absolute age estimated at c. 2.25–2.35 Ma).
Acknowledgments
This paper has benefited from comments and suggestions made by two reviewers: Dr W. Wessels (Utrecht University) and Prof. Dr Th. Van Kolfschoten (Leiden University). Mr R. Last, Mr M. van den Bosch and Mr J. Elburg assisted during the boring. Mr C. Barnard (Naturalis) assisted with sample processing.