


The gut-associated lymphoid tissue (GALT) is the first line of defence against a variety of antigens (Ag) in the intestine. Structurally, the GALT is divided into organised and diffuse compartments. The organised or inductor GALT is formed by isolated lymphoid follicles (ILF) and Peyer’s patches (PP), whereas the diffuse or effector GALT is formed by lymphocyte populations scattered across the epithelial layer or in the lamina propria. Moreover, the mesenteric lymph nodes (MLN) are part of the intestinal immune system, although they are not referred to as GALT as they do not sample intestinal Ag directly( Reference Brandtzaeg 1 ).
The main humoral immune product in the GALT, as well as in the other mucosal tissues, is the polymeric IgA, which represents approximately 80–90 % of the total secreted IgA (S-IgA) of the body( Reference Mora and von Andrian 2 , Reference Brandtzaeg 3 ). In a lower proportion, IgM (6–19 %) and IgG (4–5 %) are also present in the mucosal surfaces( Reference Brandtzaeg 4 ). The high content of S-IgA in the intestine plays a key role in the maintenance of gut homoeostasis and oral tolerance( Reference Cerutti and Rescigno 5 ).
M cells from PP are specialised cells in luminal Ag uptake that transport luminal Ag from the intestinal lumen towards Ag-presenting cells. These cells interact with interfollicular T lymphocytes( Reference Cerutti and Rescigno 5 ), which once activated bring about the differentiation and maturation of B cells, inducing them to become IgA+ cells and later IgA-secreting cells (IgA-SC). This process depends on determined cytokines such as transforming growth factor β1 (TGF-β1), IL-5, IL-6, IL-10 and IL-21( Reference Kunisawa and Kiyono 6 – Reference Ramsay, Husband and Ramshaw 9 ). B cell differentiation occurs in PP and, to a lesser extent, in ILF and MLN( Reference Mora and von Andrian 2 ). From these inductive sites, the activated B cells reach the peripheral blood stream and migrate to secretory effector sites where they extravasate depending on adhesion molecules and chemokine–chemokine receptor pairs( Reference Brandtzaeg 1 , Reference Agnello, Denimal and Lavaux 10 ). The interaction between several regulated endothelial adhesion molecules and the corresponding ligands expressed on the memory/effector B cells’ surface provokes a successful homing.
Although GALT is thought to constitute the major part of mucosal-associated lymphoid tissue (MALT), the induction of mucosal immune responses can take place in other mucosal compartments that also belong to the MALT( Reference Brandtzaeg 3 ). In fact, studies point to the possibility that other mucosal compartments may be even more important than the GALT compartments for S-IgA production. In this sense, salivary glands are considered important effector sites in the mucosal immune network( Reference O’Sullivan, Skandera and Montgomery 11 ).
Previous studies carried out in our laboratory have demonstrated the ability of a 10 % cocoa (C10) diet to modulate the immune system in vitro and in vivo ( Reference Pérez-Cano, Massot-Cladera and Franch 12 ). In particular, a down-regulation of the serum and intestinal Ig contents was observed in young rats fed a cocoa diet( Reference Ramiro-Puig, Pérez-Cano and Ramos-Romero 13 – Reference Massot-Cladera, Pérez-Berezo and Franch 16 ). As cocoa contains mainly flavonoids such as (−)-epicatechin, (+)-catechin and their polymeric forms called procyanidins( Reference Hammerstone, Lazarus and Mitchell 17 ), most of these effects of cocoa have been attributed to its high content of polyphenols( Reference Massot-Cladera, Abril-Gil and Torres 18 ). However, it must be added that cocoa is also an important source of dietary fibre (DF) (26–40 % of its content, mostly insoluble fibre), and therefore DF seems to strengthen or to be in part responsible for the effects described above( Reference Massot-Cladera, Costabile and Childs 19 ).
On the basis of this background, the aim of this study was to investigate in depth the impact of cocoa on IgA and IgM production in the intestinal and extra-intestinal (salivary glands) mucosal compartments, establishing the particular involvement of cocoa fibre (CF) in such effects. Moreover, some of the mechanisms by which cocoa intake may affect the expression of molecules involved in IgA+ B cell homing and IgA synthesis in the salivary glands were also studied.
Methods
Animals and diets
Female Wistar rats (3 weeks old) were obtained from Janvier Labs and housed in cages under conditions of controlled temperature and humidity in a 12 h light–12 h dark cycle. The rats were randomly distributed into four dietary groups: the reference (REF), C10, CF and inulin (I) groups (ten per each group). The REF group was fed the standard diet AIN-93M (Harlan); the C10 group received chow containing 10 % cocoa, which finally provided 0·4 % of polyphenols, 0·85 % of soluble fibre and 2·55 % of insoluble fibre; the CF group received a diet with the same proportions of soluble and insoluble CF as the C10 group but with a very low amount of polyphenols (<0·02 %); and the I group received the AIN-93M standard diet to which the same amount of soluble fibre provided to the C10 group (0·85 %) was added, but as I, in order to distinguish the particular effect of soluble CF (Table 1). Natural forastero cocoa and CF powders (provided by Idilia Foods S.L. (formerly Nutrexpa S.L.)) with 4·02 and 0·35 % of polyphenols, respectively, were used to elaborate the C10 and CF diets. I from chicory roots (Fibruline® Instant; InnovaFood 2005, S.L.) was used as a reference soluble fibre. The three experimental diets were elaborated on the basis of the AIN-93M formula by subtracting the amount of carbohydrates, proteins, lipids and insoluble fibre provided by the corresponding supplements. The resulting chows were isoenergetic and had a similar proportion of macronutrients (carbohydrates, proteins and lipids) and insoluble fibre as the REF diet (Table 1). Animals were given free access to water and chow. The diets lasted for 3 weeks. In addition, taking into account the obtained results, a similar design but with just 1 week of diet duration was executed; the results are included in the online Supplementary Material.
Table 1 Composition and content of macronutrients and micronutrients of the experimental diets

REF, reference; C10, cocoa; CF, cocoa fibre; I, inulin.
* Total polyphenol compounds were quantified using the Folin–Ciocalteu method.
Studies were performed according to criteria outlined by the Guide for the Care and Use of Laboratory Animals. Experimental procedures were reviewed and approved by the Ethical Committee for Animal Experimentation of the University of Barcelona (ref no. 358/12).
Sample collection and preparation
At the end of the nutritional intervention, mucosal and blood samples were collected for Ig quantification. In brief, animals were anaesthetised intramuscularly with ketamine (90 mg/kg) (Merial Laboratorios, S.A.) and xylazine (10 mg/kg) (Bayer A.G.). The small intestine, MLN and the left submaxillary salivary gland (SMG) and the left parotid salivary gland (PSG) were collected. A small piece of SMG was immediately immersed in RNAlater® (Ambion, Life Technologies), incubated at 4°C overnight and stored at −20°C until PCR analysis. The distal half of the small intestine was carefully rinsed with cold 0·9 % NaCl saline solution in order to remove faecal content, it was opened lengthwise and visible PP were excised. The intestinal tissue without PP was used to obtain the gut wash (GW) for Ig quantification by ELISA, as established previously in our laboratory( Reference Ramiro-Puig, Pérez-Cano and Ramos-Romero 13 ). The MLN, PP and both salivary glands were processed for Ig quantification. For this purpose, MLN, PP, SMG and PSG homogenates were obtained as described previously( Reference Massot-Cladera, Pérez-Berezo and Franch 16 ).
Ig quantification
IgA and IgM concentrations in GW, MLN, PP, SMG and PSG homogenates and serum IgA, IgM and IgG were quantified at the end of the nutritional intervention by ELISA following the manufacturer’s instructions for IgA (Bethyl Laboratories Inc.) and IgM (BD Biosciences). Absorbance was measured using a microplate photometer (LabSystem Multiskan, LabX) and data were interpolated using ASCENT version 2.6 software (Thermo Fisher Scientific) into standard curves, and expressed as ng/mg of tissue in homogenates and GW and as µg/ml for the serum results.
Assessment of RNA gene expression by RT-PCR
To assess the gene expressions of IgA, TGF-β1, retinoic acid receptor (RAR)α, RARβ, chemokine(C-C motif) ligand 28 (CCL28), polymeric immunoglobulin receptor (pIgR) and IL-6, RNA was isolated from SMG samples using the RNeasy® Mini Kit (Qiagen) following the manufacturer’s recommendations. The NanoDrop spectrophotometer and NanoDrop IVD-1000 version 3.1.2 software (NanoDrop Technologies) were used to quantify the amount of RNA obtained. The Agilent 2100 Bioanalyzer with the RNA 6000 LabChip Kit (Agilent Technologies) was used to assess the RNA integrity for each sample. All samples used for further experiments showed an RNA integrity number ≥9 and purity between 1·814 and 2·033 using the A260/A280 ratio. A quantity of 2 μg of total RNA was converted to complementary DNA. Specific PCR TaqMan® primers and probes (Applied Biosystems) were used to measure selected targets: Iga (331943, made to order), Ccl28 (Rn00586715_m1, inventoried), Tgfb1 (Rn00572010_m1, inventoried), Rara (Rn00580551_m1, inventoried), Rarb (Rn01537835_m1, inventoried), Pigr (Rn00562362_m1, inventoried) and Il6 (Rn01410330_m1, inventoried). Quantitative real-time PCR assays were performed in duplicate for each sample using an ABI PRISM 7900HT Sequence Detection System (Applied Biosystems). Quantification of the studied genes was normalised to the housekeeping Gusb (β-glucuronidase gene, Rn00566655_m1, inventoried), which showed constant level of expression in our experimental conditions, similar to the level of expression of the studied genes. SDS version 2.4 software (Applied Biosystems) was used to analyse the expression data. The amount of target mRNA relative to the endogenous control expression was calculated for the three nutritional intervention groups relative to values from the REF group, which represents 100 % gene expression, using the standard
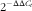 $$2^{{{\minus}\Delta \Delta C_{t} }} $$
method, as previously described(
Reference Pérez-Cano, Ramírez-Santana and Molero-Luís
20
). Results are expressed as mean values with their standard errors of the percentage of these values.
$$2^{{{\minus}\Delta \Delta C_{t} }} $$
method, as previously described(
Reference Pérez-Cano, Ramírez-Santana and Molero-Luís
20
). Results are expressed as mean values with their standard errors of the percentage of these values.
Statistical analysis
Levene’s and Kolmogorov–Smirnov tests were applied to assess variance equality and normal distribution, respectively. Conventional one-way ANOVA followed by the Bonferroni’s post hoc significance test was applied when normal distribution and equality of variance existed. Otherwise, non-parametric tests (Kruskal–Wallis and Mann–Whitney U ranked-sum tests) were used to assess significance. Statistical analysis was performed using the software package SPSS 22.0 (SPSS Inc.), and significant differences were established at P<0·05.
Results
Body weight and chow intake
Throughout the study, no changes in the increasing body weight pattern were found as a result of CF and I diet intakes. However, the C10-fed animals showed a slower body weight increase, with this being significantly lower in comparison with other groups throughout the study (up to 31·7 % lower body weight increase in the C10 group compared with the rest of the groups at day 7, 72·2 % at day 14 and 103·2 % at day 21). This effect was not related to lower chow intake, which was similar throughout the study in all the experimental groups (data not shown). Similar results on body weight by C10 diet have been already reported( Reference Ramiro-Puig, Pérez-Cano and Ramos-Romero 13 , Reference Pérez-Berezo, Franch and Castellote 15 , Reference Massot-Cladera, Abril-Gil and Torres 18 ).
Ig content in the inductor intestinal compartments
After 3 weeks of nutritional intervention, changes in IgA and IgM concentrations in both the studied inductor compartments in the intestine (PP and MLN) were observed (Fig. 1). In PP, the C10 diet significantly reduced IgA and IgM concentrations (Fig. 1(a) and (b)) compared with those observed in the rest of the groups (P<0·05), whereas the CF increased the IgM concentration with respect to the I-fed animals (P<0·05). The I diet did not significantly modify IgA and IgM concentrations in PP. The same changes on PP IgA and IgM concentrations were already observable after just 1 week of nutritional intervention with the C10 diet (P<0·05) (online Supplementary Table S1).

Fig. 1 Effects of cocoa (C10) diet on IgA and IgM content in the inductor intestinal compartments after 3 weeks of diet. IgA (a) and IgM (b) concentrations in Peyer’s patches, and IgA (c) and IgM (d) concentrations in mesenteric lymph nodes. Values are means (n 10), with their standard errors represented by vertical bars. * Mean value was significantly different from that of the reference (REF) group (P<0·05); † mean value was significantly different from that of the group consuming the C10 diet (P<0·05); ‡ mean value was significantly different from that of the group consuming the cocoa fibre (CF) diet (P<0·05); § mean value was significantly different from that of the group consuming the inulin (I) diet (P<0·05). ![]() , REF;
, REF; ![]() , C10;
, C10; ![]() , CF;
, CF; ![]() , I.
, I.
In MLN, C10 intake did not significantly modify IgA concentration (Fig. 1(c)) but resulted in a significant reduction in IgM concentration in comparison with those in the REF, CF and I groups (P<0·05) (Fig. 1(d)). No statistical differences in MLN IgM or IgA were observed after just 1 week of C10 diet (online Supplementary Table S1). Moreover, in the same tissue, the CF diet increased the IgA concentration compared with the other groups (P<0·05) and also IgM concentration compared with the I group (P<0·05) (Fig. 1(c) and (d)). This increase in MLN IgA and IgM was already observed after 1 week of diet supplementation (online Supplementary Table S1). The I diet did not significantly modify IgA and IgM concentrations in MLN (Fig. 1(c) and (d)).
Ig content in the effector intestinal compartments
IgA and IgM concentrations were also determined in GW as representative of the production of Ig in the effector tissues (Fig. 2). Although all three experimental diets significantly reduced the GW IgA concentration compared with the REF group (P<0·05), it was the C10 diet that produced the most profound reduction in comparison with the CF and I diets (P<0·05) (Fig. 2(a)) that was already observed after 1 week of nutritional intervention (P<0·05) (online Supplementary Table S1). The decrease in IgA after the I diet intake was also more pronounced than that observed in the CF group (P<0·05). With regard to IgM in GW, the C10 diet reduced its concentration compared with the rest of the groups (P<0·05) (Fig. 2(b)). The CF diet increased the IgM concentration compared with the I diet (P<0·05), but not when compared with the REF diet.

Fig. 2 Effects of cocoa (C10) diet on IgA and IgM contents in the effector intestinal compartments after 3 weeks of diet. IgA (a) and IgM (b) concentrations in gut washes. Values are means (n 10), with their standard errors represented by vertical bars. * Mean value was significantly different from that of the reference (REF) group (P<0·05); ‡ mean value was significantly different from that of the group consuming the cocoa fibre (CF) diet (P<0·05); § mean value was significantly different from that of the group consuming the inulin (I) diet (P<0·05). ![]() , REF;
, REF; ![]() , C10;
, C10; ![]() , CF;
, CF; ![]() , I.
, I.
Ig content in the effector extra-intestinal mucosal compartments
Regarding the salivary glands, a similar pattern was observed in the SMG and PSG for both Ig (Fig. 3). The C10 diet significantly reduced IgA and IgM contents in both salivary glands compared with the rest of the diets (P<0·05). The CF diet also reduced IgA concentration in both salivary glands in a more modest way in comparison with the REF diet (Fig. 3(a) and (c)), without affecting the IgM concentration (Fig. 3(b) and (d)). The same C10 and CF effects were already observed after 1 week (online Supplementary Table S1). No changes in IgA and IgM were observed in the salivary glands after I diet intake.

Fig. 3 Effects of cocoa (C10) diet on IgA and IgM contents in the effector extra-intestinal mucosal compartments after 3 weeks of diet. IgA (a) and IgM (b) concentrations in the submaxillary salivary gland and IgA (c) and IgM (d) concentrations in the parotid salivary gland. Values are means (n 10), with their standard errors represented by vertical bars. * Mean value was significantly different from that of the reference (REF) group (P<0·05); ‡ mean value was significantly different from that of the group consuming the cocoa fibre (CF) diet (P<0·05); § mean value was significantly different from that of the group consuming the inulin (I) diet (P<0·05). ![]() , REF;
, REF; ![]() , C10;
, C10; ![]() , CF;
, CF; ![]() , I.
, I.
Systemic Ig
Serum concentrations of IgA, IgM and IgG from all the experimental groups after 3 weeks of diet are summarised in Fig. 4. With regard to IgA concentration, the CF diet intake resulted in higher levels of IgA compared with the rest of the diets (P<0·05) (Fig. 4(a)). IgM concentration was reduced by the C10 diet when compared with the REF diet after 3 weeks of intervention (P<0·05) (Fig. 4(b)). Serum IgG concentration was diminished by both the C10 and the CF diets compared with the REF diet at the end of the 3rd week of study (P<0·05) (Fig. 4(c)).

Fig. 4 Effects of cocoa (C10) diet on serum IgA (a), IgM (b) and IgG (c) concentrations after 3 weeks of diet. Values are means (n 10), with their standard errors represented by vertical bars. * Mean value was significantly different from that of the reference (REF) group (P<0·05); † mean value was significantly different from that of the group consuming the cocoa (C10) diet (P<0·05); § mean value was significantly different from that of the group consuming the inulin (I) diet (P<0·05). ![]() , REF;
, REF; ![]() , C10;
, C10; ![]() , CF;
, CF; ![]() , I.
, I.
Submaxillary gland gene expression
Gene expressions of IgA, CCL28, TGF-β1, RARα, RARβ, pIgR and IL-6 were assessed in the SMG at the end of the study to examine in depth the mechanism involved in the Ig modulation activity of cocoa products (Fig. 5). IgA expression was significantly down-regulated only in the C10 group compared with the REF and CF diets, whereas CCL28 was significantly up-regulated by this diet compared with the CF diet (P<0·05). In addition, a tendency to down-regulate TGF-β1 gene expression has been observed in the C10 diet group. On the other hand, the CF diet-fed animals showed significantly lower expressions of TGF-β1 and RARα after 3 weeks of nutritional intervention compared with REF animals (P<0·05). Moreover, the expression of pIgR in animals fed CF was also significantly lower than those fed the I diet (P<0·05). The I diet did not modify the expression of any of the studied genes. Il6 expression was not detected in the SMG from any of the studied groups.

Fig. 5 Expressions of genes associated with IgA synthesis, secretion, switching and intestinal homing in submaxillary salivary gland after 3 weeks of diet. Expression levels were normalised using the expression of Gusb (β-glucuronidase) as the endogenous housekeeping gene. Values are means (n 5–6), with their standard errors represented by vertical bars. * Mean value was significantly different from that of the reference (REF) group, which represents 100 % gene expression (P<0·05); ‡ mean value was significantly different from that of the group consuming the cocoa fibre (CF) diet (P<0·05); § mean value was significantly different from that of the group consuming the inulin (I) diet (P<0·05). ![]() , REF;
, REF; ![]() , cocoa;
, cocoa; ![]() , CF;
, CF; ![]() , I; CCL28, chemokine(C-C motif) ligand 28; TGF-β1, transforming growth factor β1; RAR, retinoic acid receptor; pIgR, polymeric immunoglobulin receptor.
, I; CCL28, chemokine(C-C motif) ligand 28; TGF-β1, transforming growth factor β1; RAR, retinoic acid receptor; pIgR, polymeric immunoglobulin receptor.
Discussion
In previous studies, we reported that the intake of a C10 diet down-regulates the Ig content at systemic and intestinal levels in young rats( Reference Ramiro-Puig, Pérez-Cano and Ramos-Romero 13 – Reference Massot-Cladera, Pérez-Berezo and Franch 16 ). However, the impact of C10 at the extra-intestinal mucosal level, the mechanism by which it may act and the involvement of CF in such effects are unknown to date. This study compares the effect of whole cocoa intake with that produced by CF extract (with a lower amount of polyphenols) on Ig production and that produced by a soluble fibre such as I. The results showed that the diet containing whole cocoa induced the most pronounced effects on the synthesis of Ig in intestinal and extra-intestinal mucosal compartments. In addition, we found that most of the C10 effects observed at the end of the 3rd week were already significant after just 1 week of the diet. The impact of the CF on Ig production was lower than that produced by whole cocoa in some tissues (GW and salivary glands for IgA), whereas in other compartments it had no effect (PP for both Ig and GW, MLN and serum for IgM) or it even exerted the opposite effect (MLN and serum for IgA). Finally, the soluble fibre-enriched diet only modified GW IgA levels, decreasing its concentration, similar to those effects resulting from CF intake. The consequence and meaning of these results in each compartment are discussed next.
At the GALT inductor sites (PP and MLN), C10 intake attenuated the production of IgM in MLN and that of IgA and IgM in PP after 3 weeks of diet, the latter effects already being observed after 1 week of the diet. On the contrary, the CF diet (containing a very low amount of polyphenols) exerted the opposite effect of the C10 diet by increasing the IgA content in both PP and MLN (only significant in the latter). These results are in line with those previously reporting a higher proportion of IgA-coated bacteria in faeces from CF-fed animals( Reference Massot-Cladera, Costabile and Childs 19 ). To our knowledge, this is the first report about the immune-potentiating effect of CF. In line with these results, a rise in IgA concentration has also been described for well-known substances with prebiotic effects( Reference Vogt, Meyer and Pullens 21 ). Contrary to that, the prebiotic substance used in the present study (I) did not modulate Ig production, as reported in other studies( Reference Seifert and Watzl 22 ), which could be attributed to several factors, such as the methodologies, population groups, length of intervention, as well as the type and dose of I used( Reference Kolida, Meyer and Gibson 23 ). Likewise, the CF diet increased the IgM content in PP and MLN with respect to the I group, suggesting that CF has particular compounds and/or composition that are not in common with I, which could contribute to these effects. In addition, it is possible that the prebiotic effects attributed to the CF( Reference Massot-Cladera, Costabile and Childs 19 ) may be involved in the mechanism by which CF increased the IgA concentration in this compartment. This enhancement was also reflected in the serum from the CF group, in which IgA concentration was also increased, behaviour that is not always present after prebiotic intake( Reference Delgado, Thomé and Gabriel 24 , Reference Yamamoto, To and Hayashi 25 ). Moreover, this means that the fibre present in cocoa can be discarded as being the main factor responsible for the down-regulation caused by the whole cocoa, and this allows us to suggest that other compounds present in cocoa may be masking and counteracting the immune-potentiating effect of the CF.
Regarding the effector intestinal compartment (GW), previous studies carried out in our laboratory reported a reduction in IgA and IgM concentrations after a 6-week cocoa diet( Reference Pérez-Berezo, Franch and Castellote 15 ). In line with those results, in the present study, we have observed that this reductive effect of C10 is already evidenced after 3 weeks and 1 week of diet, thus showing the early down-regulatory effects of C10. The CF- or I-fed animals also presented lower IgA concentrations in this compartment. Therefore, the CF effect on inductor sites is not reproduced at the effector level. However, as the CF influence was not as pronounced as that produced by the C10 diet, CF might only be partially contributing to the down-regulatory effects of cocoa, and other cocoa compounds such as flavonoids might be potentiating the effect of fibre or even be the key factor responsible for this effect. Nonetheless, studies using three different polyphenol-enriched cocoa extracts evidenced that, although they were also able to modulate Ig production, they were not the main compounds in charge of such an effect( Reference Massot-Cladera, Abril-Gil and Torres 18 , Reference Massot-Cladera, Franch and Castellote 26 ). Therefore, further studies should be carried out in order to fully elucidate the contribution of other cocoa components such as methylxanthines on these intestinal effects.
A third compartment included in the present study was the salivary glands. Despite the existing evidence about the protective effect of flavonoid consumption from medication and/or age-related dysfunction of salivary glands( Reference Yamamoto, Staples and Wataha 27 – Reference Ryo, Ayako and Yoh 29 ), there is no information about the impact of polyphenols on salivary glands’ Ig production. In fact, to our knowledge, this is the first report describing the impact of whole cocoa (containing polyphenols and fibre) on Ig production in the extra-intestinal mucosal compartments. We observed that the C10 diet diminished IgA and IgM concentrations in the salivary glands at the end of just 1 week of dietary intervention. Similarly, there is scarce information about the influence of indigestible carbohydrates on IgA response in the salivary glands, this being the first time the impact of fibre from cocoa or as I on Ig production in these tissues has been described. In this study, we have reported that the CF-fed animals, although in a more moderate way than the C10 group, also presented lower concentrations of IgA in this compartment at the same time points. These results are not in line with those observed in animals fed fructo-oligosaccharides (FOS) and a mixture of polydextrose and lactitol, which induced higher submandibular IgA concentrations( Reference Yamamoto, To and Hayashi 25 ). Unlike the prebiotic substances and the results mentioned above, the CF did not exert immune-potentiating effects in the salivary glands, thus suggesting that not only the fibre but also other compounds present in cocoa might be interfering with Ig production in the salivary glands.
In order to obtain insights into the underlying mechanisms associated with the reduction in IgA concentration in the salivary glands due to the diets, the gene expressions of some key molecules involved in IgA synthesis and regulation, as well as IgA-SC mucosal homing, have been evaluated in this compartment. In this study, we have observed that IgA gene expression was drastically down-regulated in the C10 group but not in the CF and I groups. These results agree with the most pronounced effects of the C10 diet on SMG IgA content, which could be due to the presence of a lower number of IgA-SC cells in this tissue and/or a lower ability to secrete it. Some of the molecules involved in the maturation and differentiation of B cells in becoming IgA-SC are IL-6 and TGF-β1. Although IL-6 mRNA was not detected in the SMG, the TGF-β1 gene expression tended to be lessened by all three experimental diets, only being statistically significant in the CF diet group. These results suggest the attenuating effect of soluble fibre in the differentiation of B cells into the salivary glands and could partially explain the decreased IgA content found after C10 and CF diets. Similar results have been found in the small intestine from animals fed a cocoa diet for 3 and 7 weeks( Reference Pérez-Berezo, Franch and Castellote 15 , Reference Pérez-Berezo, Franch and Ramos-Romero 30 ).
Once the IgA-secreting B cells are activated, they migrate to the blood and later return to the mucosal effector compartments( Reference Cerutti and Rescigno 5 ). Mucosal homing is mainly regulated by adhesion molecules and chemokine-mediated interaction. To date, CCL28 chemokine and its corresponding receptor are crucial in the recruitment of IgA+ B cells to the extra-intestinal as well as intestinal mucosa( Reference Lazarus, Kunkel and Johnston 31 , Reference Kunkel, Kim and Lazarus 32 ). In this study, similarly to what has been reported in the small intestine( Reference Pérez-Berezo, Franch and Ramos-Romero 30 ), C10 intake, but not the CF, up-regulated the expression of CCL28. These results may point to a compensatory mechanism to strongly attract the IgA + B cells to avoid the down-modulation on IgA concentration. Other molecules involved in the intestinal cell homing mechanism are those that constitute the RAR family( Reference Sirisinha 33 ). After evaluating the expression of both RARα and RARβ in the salivary gland, we found that the C10 intake for 3 weeks did not modify either the expression of RARα or RARβ at this level, which agrees with those results obtained in the small intestine and MLN after 3 weeks of diet( Reference Pérez-Berezo, Franch and Castellote 15 ). However, the CF diet significantly reduced the expression of RARα but not RARβ, which could contribute to the IgA-decreasing effect of the CF on salivary glands. Finally, in order to reach the mucosal lumen, the IgA requires the association with the transmembrane epithelial protein pIgR( Reference Brandtzaeg 4 ). We found that pIgR gene expression was not modified by the C10 diet at the extra-intestinal level or in the intestine( Reference Pérez-Berezo, Franch and Castellote 15 ), thus indicating that the reduction of IgA concentration associated with the C10 intake is not due to a lower transport across the epithelium. Little is known about the impact of fibre on pIgR gene expression. To date, an increase in its expression after the intake of FOS and a mixture of polydextrose and lactitol in rodents has been reported( Reference Yamamoto, To and Hayashi 25 , Reference Nakamura, Nosaka and Suzuki 34 ). However, in the present study, the CF intake resulted in a lower pIgR gene expression, suggesting that the reduction in IgA content in the salivary glands as a result of CF intake also takes place by means of IgA transport.
In summary, we have demonstrated that a C10 diet led to a lower IgA and IgM secretion in both inductor and effector intestinal compartments as well as in the extra-intestinal mucosal compartments in young Wistar rats. Most of these effects could be already observed after 1 week of nutritional intervention. Soluble fibre seems to be partially responsible for the lower IgA secretion into the intestinal lumen. In spite of the results produced by the C10 diet, it is worth noting that the enhancement by the CF diet of IgA synthesis in the inductor intestinal compartment was also detectable at the systemic level, where higher IgA concentration was quantified. Both the C10 and CF diets produced a down-regulatory effect on IgA secretion in the extra-intestinal tissues. The C10 diet interacted with the mechanisms involved in IgA synthesis in a similar way to that observed in the small intestine, whereas the CF showed particular effects on homing and transcytosis of IgA across the epithelial gland.
Overall, CF has IgA immune-potentiating effects that are counteracted when it is present in the whole cocoa product. This fact suggests that other cocoa components are involved in such an effect. This immunomodulation was not found in the I diet. Further studies must be considered to ascertain the mechanism by which CF is able to act in up-regulating IgA production at the intestinal inductor compartments, whereas it down-regulates its production in those of the mucosal effectors.
Acknowledgements
The authors thank Judith González, Ignasi Azagra, Nadir Rodríguez, Mònica Solà and Silvia Plana for their technical assistance. The authors also thank the genomic services of the ‘Centres Científics i Tecnològics’ of the University of Barcelona (CCiT-UB); Idilia Foods S.L. (formerly Nutrexpa S.L.) and InnovaFood 2005 S.L. for providing the conventional cocoa and cocoa fibre powders and the inulin extract, respectively.
The present study was supported by a grant from the Spanish Ministry of Economy and Competitiveness (AGL2011-24279). M. M.-C. holds a fellowship from the Generalitat de Catalunya (grant no. 2014FI_B2 00048). None of the funders had a role in the design or analysis of the study or in the writing of this article.
The authors’ contributions were as follows: M. M.-C., À. F., F. J. P.-C. and M. C. conceived and designed the research; M. M.-C., F. J. P.-C. and M. C. carried out the experiments, the data analysis and were involved in the interpretation of the data; M. M.-C. contributed to the initial draft of the manuscript; À. F., F. J. P.-C. and M. C. contributed to the critical revision of the manuscript; M. C. has primary responsibility for the final content. All authors have read and approved the final version of the manuscript for publication.
The authors declare that there are no conflicts of interest.
Supplementary Material
For supplementary material/s referred to in this article, please visit http://dx.doi.org/doi:10.1017/S000711451600074X








