

In marine turtles the selection of oviposition habitat is a key determinant of reproductive success because it can affect embryo survival, body size at hatching, sex ratio and the orientation of offspring (Musseau & Fox, 1998; Kamel & Mrosovsky, Reference Kamel and Mrosovsky2005). Studies of the factors influencing the distribution of turtle nests along beaches have in several cases identified nest site selection that has been interpreted as attempts by females to increase the survival of their offspring (Mortimer, Reference Mortimer and Bjorndal1982; Kamel & Mrosovsky, Reference Kamel and Mrosovsky2005; Karawas et al., 2005). However, in other studies no nest site selection has been detected or selection was equivocal (Hays et al., Reference Hays, MacKay, Adams, Mortimer, Speakman and Boerema1995; Kamel & Mrosovsky, Reference Kamel and Mrosovsky2004), or no relationship was identified between nest site selection and reproductive success (Mortimer, Reference Mortimer and Bjorndal1982). Such results suggest that differences among species, areas or seasons can be strong, but also that our knowledge of beach features necessary for turtle nesting is incomplete (Miller, Reference Miller, Lutz and Musick1997).
An important analytical issue in the study of species-habitat relationships is spatial autocorrelation, which occurs when the environmental similarity among points increases as the spatial distance decreases. The presence of spatial autocorrelation can violate the assumption of independence required for statistical analyses such as regression and ANOVA (Lichstein et al., Reference Lichstein, Simons, Shriner and Franzreb2002; Wagner & Fortin, Reference Wagner and Fortin2005). Spatial autocorrelation is frequently overlooked in ecological studies, and analyses of species-habitat relationships ignoring the effects of spatial autocorrelation can result in the over- or underestimation of the effect of environmental variables, the detection of spurious relationships, and misinterpretation (Lichstein et al., Reference Lichstein, Simons, Shriner and Franzreb2002; Wagner & Fortin, Reference Wagner and Fortin2005). In any analysis of the relationship between turtle nesting and habitat it is thus necessary to consider spatial autocorrelation.
The hawksbill turtle Eretmochelys imbricata is Critically Endangered because of the exploitation of adults for food and trade, egg poaching, and the alteration of habitat, particularly nesting beaches (Meylan, Reference Meylan1999; Meylan & Donnelly, 1999; IUCN, 2006; Koch et al., Reference Koch, Nichols, Peckhamb and de la Toba2006). Females may lay eggs at a specific distance from or elevation above the shoreline to avoid the risk of nest destruction during storms and to reduce the risk of nest depredation, and frequently nest in shaded areas, which produces a more balanced sex ratio (Horrocks & Scott, Reference Horrocks and Scott1991; Kamel & Mrosovsky, Reference Kamel and Mrosovsky2005, Reference Kamel and Mrosovsky2006). However, although the hawksbill turtle has a worldwide distribution most of these studies were in the Caribbean, and it is unclear how widely applicable they are.
I evaluated the influence of beach features on the nesting activity of hawksbill turtles in the Arabian Gulf, which has important populations of this species but is undergoing rapid coastal development. The study area is located in Rass Laffan, north-eastern Qatar (25.9° N, 51.5°E). Climate is hyperarid and vegetation on the dunes is limited to a few xerophitic species of grasses and bushes; maximum semidiurnal tidal excursion is c. 1.5 m. Rass Laffan has 14 km of coastline with both rocky and sandy areas; offshore there are submerged coral patches and seagrass beds. A programme of nest monitoring and protection since 2001 has identified c. 300 turtle nests per year. The hawksbill is the only turtle known to nest in this area (Tayab & Quiton, Reference Tayab and Quiton2003).
I monitored 1.3 km of coastline daily during the nesting season (April–September 2005) for turtle nesting activity the previous night. Monitoring was by walking immediately after dawn, for reliable identification of tracks (Mortimer, Reference Mortimer and Bjorndal1982). A successful nesting was recorded if there was evidence of covering the nest with the front flippers; the track was otherwise considered an unsuccessful nesting attempt. The location of each nest or failed attempt was recorded using a Global Positioning System. In a minority of cases females may dig a nest without laying but nests were not opened to confirm egg presence and therefore it is possible that the nest count slightly overestimates the actual number of nests with eggs.
To measure beach features I divided the area into 35 m sections and recorded six environmental features in each section: beach angle (averaged from three equally spaced measurements per section recorded at the high tide mark); height of the dune above the nightly low tide marks; distance between the top of the dune and the low tide; percentage rock cover; abundance of vegetation on the dune (measured using the following scale; 0: vegetation absent; 1: discontinuous vegetation cover; 2: continuous cover); dominant hardness of the dune soil (in this area there is a strong difference between zones where the soil is composed of soft sand, i.e. hardness = 0, and zones where the soil is compact and difficult to excavate, i.e. hardness = 1).
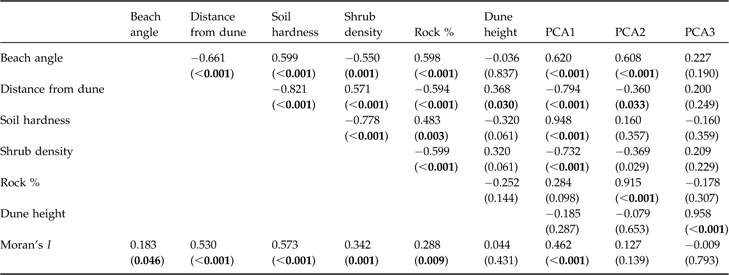
Beach features were strongly correlated (Table 1) and therefore they could not be used together as independent variables. A principal component analysis (PCA) was used to reduce the six variables to uncorrelated factors. The resulting axes were rotated using a Varimax rotation to aid interpretation. The first three factors were retained, explaining 86.3% of variation (PCA1 42.7%; PCA2 25.1%; PCA3 18.5%). PCA1 was strongly correlated with soil hardness and beach angle and negatively correlated with shrub density and distance between low tide and dune, PCA2 was positively correlated with rock cover and beach angle, and PCA3 was positively correlated with dune height.
Table 1 Correlations (Pearson’s correlation coefficient, r, with P in parentheses) between the six beach environmental variables, and between the six variables and the three factors extracted by principal component analysis (PCA, see text for further details), and the spatial autocorrelation (Moran’s I) of each variable and the three PCA factors. Significant results are in bold.
The influence of the PCA axes, as independent variables, on three dependent variables was tested using regression. The dependent variables were number of nests per section (nest density), number of unsuccessful nesting attempts (failure density), and overall turtle activity per sector (i.e. the sum of successful and unsuccessful nesting attempts). Overall activity is obviously not independent of nest or failure density but is meant to evaluate the coastline features that allow turtles to arrive on the beach independently of the outcome of the nesting attempt.
I used Moran’s I to measure the spatial autocorrelation of independent and dependent variables. Most habitat features displayed strong spatial autocorrelation (Table 1). Traditional regression models used in preliminary analyses resulted in violations of the assumptions of spatial independence (Lagrange diagnostics: nest density χ2 = 3.27, P = 0.07; failure density χ2 = 5.15, P = 0.02; Anselin et al., Reference Anselin, Bera, Florax and Yoon1996). I therefore used spatial simultaneous autoregressive lag model estimation (lagSAR) to evaluate which attributes of the coastlines are related to nesting activity. LagSAR includes a maximum likelihood estimation of the spatial autoregressive coefficient in multiple regression; these models are therefore suitable to evaluate the relationship between species distribution and environmental features when spatial autocorrelation may bias the results of regression (Anselin, Reference Anselin and Baltagi2001). The goodness of fit of models was evaluated by calculating the squared Pearson’s correlation coefficient between fitted and observed values. Spatial analyses were performed using spdep 0.3-12 in the statistical system R v. 2.2 (R, 2007); count data were square-root transformed prior to analysis.
In total 20 successful and 23 unsuccessful turtle nests were found. The study site is c. 9% of the Rass Laffan coastline and the observed nests represent c. 10% of the 170–200 nests observed during 2005 in the Rass Laffan area (authorities of Rass Laffan City, unpubl. data). The distribution of nests and unsuccessful attempts was spatially autocorrelated (Moran’s I = 0.250, P = 0.003). In some areas almost all turtles that approached the beach excavated a nest, whereas in other areas nesting was very scarce but many unsuccessful attempts were observed.
Turtle activity on the beach was negatively related to PCA2 (Table 2), indicating that turtles were less likely to arrive on beaches with a steep coastline and many rocks. Nest density was negatively related to both PCA1 and PCA2, indicating that turtles arrived and dug nests in areas with low rock cover and soft soil, with dunes far from the shoreline and high shrub abundance. Failure density was positively related to PCA1 and negatively related to PCA2, indicating that in areas with low rock cover but hard soil, with dunes close to the shoreline, and low shrub abundance, turtles arrived but returned to the sea without nesting. Despite the relatively small sample size these results demonstrate the influence of coastline features on turtle nesting activity and confirm the key importance of certain aspects of the terrestrial environment for aquatic reptiles that nest on land (Ficetola & De Bernardi, Reference Ficetola and De Bernardi2006; Kemel & Mrosovsky 2006).
Table 2 Results of lag spatial autoregressive models (see text for details) relating the nesting activity of hawksbill turtles to beach features, as summarized by the three independent axes of a principal component analysis (see text for details). Significant factors are in bold.
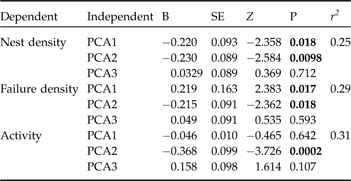
The coastline has to allow easy access to the beach, with a low rock cover and not steep, but this accessibility only determines where turtles can access the beach and not the outcome of nesting attempts. PCA1 is the factor that discriminated between areas suitable and unsuitable for reproduction: in areas with compact soil, with dunes close to the shoreline and with low shrub density turtles generally returned to the sea without nesting. Kamel & Mrosovsky (Reference Kamel and Mrosovsky2005) observed that hawksbill turtles have a preference for nesting in areas shaded by vegetation, and recorded higher hatch success in vegetated than in open areas. The selection for areas with vegetation could limit the temperature and influence the moisture and water potential of the nest chamber, and these features affect the survival and body size of offspring, and also their sex determination (Ackerman, Reference Ackerman, Lutz and Musick1997; Kamel & Mrosovsky, Reference Kamel and Mrosovsky2006). The selection of areas with vegetation is particularly important in the tropics where insolation and temperature can be extreme, and could become critical in the future to buffer increases in nest temperatures caused by global warming (Kamel & Mrosovsky, Reference Kamel and Mrosovsky2006). Nesting activity was also observed only in areas with a gentle slope and with dunes far from the coastline, in contrast to the observations of Horrocks & Scott (Reference Horrocks and Scott1991) who observed a preference for steeper coastlines. These differences are probably caused by differences in topography. In Qatar the steepest sections of coast have high rock cover and are inaccessible to turtles.
This study successfully used models that account for the spatial structure of the data to estimate the effects of habitat variables on hawksbill turtle nesting. Such spatially explicit models can help to ease the difficulty of taking management actions in places where detailed local studies are not available. The features identified as important in this study should be prioritized in coastline management for hawksbill turtles. Accessibility of the beach and the presence of soft sand should be preserved to allow turtles to reach nesting sites and excavate nests. Coastal development should not take place in areas suitable for turtle nesting but when beaches are altered, suitability needs to be promptly restored. It is also important to preserve the natural vegetation along the coastline. Both the removal of natural vegetation and substitution with non-native species can have critical consequences (Kamel & Mrosovsky, Reference Kamel and Mrosovsky2006).
Acknowledgements
I thank the Rass Laffan City authorities for logistical support, and W. Thuiller and two anonymous reviewers for comments on earlier versions of the manuscript. This study was funded by ERM Italy.
Biographical sketch
Francesco Ficetola’s research interests include the conservation, evolutionary ecology and population genetics of herpetofauna. He combines academic research with the practical implementation of monitoring and management plans. This study is part of a collaboration with the authorities of Qatar to evaluate plans for the conservation of marine turtles in areas where coastal development is ongoing.


