


Introduction
In many places outside protected areas ecosystem/habitat connectivity across the landscape is characterized by distinct pathways connecting important resources (Blake & Inkamba-Nkulu, Reference Blake and Inkamba-Nkulu2004; Von Gerhardt et al., Reference Von Gerhardt, Van Niekerk, Kidd, Samways and Hanks2014). Studies have shown that encroachment of development onto wildlife paths is an important driver of human–wildlife conflict (Guerbois et al., Reference Guerbois, Chapanda and Fritz2012; Songhurst & Coulson, Reference Songhurst and Coulson2014). Although temporal and spatial patterns of crop raiding by wildlife appear to be influenced by a variety of other factors associated with the location of fields in relation to landscape features such as water availability, protected areas and daytime refuges for wildlife (Naughton-Treves, Reference Naughton-Treves1998; Linkie et al., Reference Linkie, Dinata, Nofrianto and Leader-Williams2007; Graham et al., Reference Graham, Notter, Adams, Lee and Ochieng2010; Guerbois et al., Reference Guerbois, Chapanda and Fritz2012), appropriate land-use planning is advocated as a requirement to protect wildlife pathways and alleviate human–wildlife conflict (Weaver et al., Reference Weaver, Paquet and Ruggiero1996; Osborn & Parker, Reference Osborn and Parker2003; Douglas-Hamilton et al., Reference Douglas-Hamilton, Krink and Vollrath2005; Fernando et al., Reference Fernando, Wikramanayake, Weerakoon, Jayasinghe, Gunawardene and Janaka2005; Linnell et al., Reference Linnell, Nilsen, Lande, Herfindal, Odden, Skogen, Woodroffe, Thirgood and Rabinowitz2005; Von Gerhardt et al., Reference Von Gerhardt, Van Niekerk, Kidd, Samways and Hanks2014). Few studies, however, have examined elephant Loxodonta africana behaviour at a population level to help justify pathway protection as an important strategy in conflict intervention. If protecting pathways is to be effective in reducing human–wildlife conflict we need a greater understanding of how a population at large is using these critical movement pathways, and the extent to which the movement of wildlife along the pathways is influenced by the presence and activity of people.
How elephants utilize various landscapes is determined by a combination of environmental (e.g. Hanks, Reference Hanks1979; Viljoen & Bothma, Reference Viljoen and Bothma1990; Redfern et al., Reference Redfern, Grant, Biggs and Getz2003; Murwira & Skidmore, Reference Murwira and Skidmore2005; Chamaillé-Jammes et al., Reference Chamaillé-Jammes, Valeix and Fritz2007; Loarie et al., Reference Loarie, van Aarde and Pimm2009; Young et al., Reference Young, Ferreira and van Aarde2009) and social factors (e.g. Moss & Poole, Reference Moss, Poole and Hinde1983; Stokke, Reference Stokke1999; Wittemyer et al., Reference Wittemyer, Getz, Vollrath and Douglas-Hamilton2007). Movements also vary temporally according to seasons (Douglas-Hamilton et al., Reference Douglas-Hamilton, Krink and Vollrath2005; Young et al., Reference Young, Ferreira and van Aarde2009), time of day, the speed of movement and the length of time spent in any one location (Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009). In areas where home ranges overlap with human habitation and infrastructure, animals also have to adapt their behaviour to avoid disturbance by people (Martin et al., Reference Martin, Basille, Moorter, Kindberg, Allainé and Swenson2010). Disturbances to animals may arise as a result of physical impediments to movement (i.e. fences, roads, cultivated land), or human behaviour such as hunting, farming and wildlife conflict mitigation activities. Animals have been found to adapt to risks associated with humans; for example, elephants tend to move under cover of darkness and travel more quickly in areas close to human habitation (Douglas-Hamilton et al., Reference Douglas-Hamilton, Krink and Vollrath2005; Galanti et al., Reference Galanti, Preatoni, Martinoli, Wauters and Tosi2006; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009). Studies have also reported elephants avoiding unprotected areas with high human population density and activity (Parker & Graham, Reference Parker and Graham1989; Happold, Reference Happold1995; Hoare, Reference Hoare1999; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009), and moving in high-risk areas such as cultivated fields predominantly at night (Osborn, Reference Osborn1998; Sitati et al., Reference Sitati, Walpole, Smith and Leader-Williams2003). Carnivores appear to respond in a similar way to the presence of humans (Martin et al., Reference Martin, Basille, Moorter, Kindberg, Allainé and Swenson2010). Guerbois et al. (Reference Guerbois, Chapanda and Fritz2012) argued that for improved management of human–elephant conflict, elephants need to associate human landscapes with perceived risk. Risk avoidance is fundamental to the design of various mitigation techniques to alter movement behaviour and deter elephants from individual fields and settlements (Conover, Reference Conover2002), but it is not clear how such behavioural adaptations can be incorporated into the design of practical conservation management strategies at a landscape level.
Here we examine the fine-scale movement behaviour of a population of free-ranging elephants that share their range with humans in a multi-use heterogeneous landscape, and the extent to which adaptations in the elephants’ movement that define risk avoidance behaviour contribute to spatial separation in land use.
Study area
The study was conducted during January 2008–July 2010 on the eastern side of the Okavango Panhandle, where the Okavango River reaches the Okavango Delta in Botswana (Fig. 1). The eastern Panhandle covers 8,732 km2 and its northern and southern boundaries are delineated by the Namibian border and the northern buffalo fence, respectively. There are protected areas to the north, south and east of the area, which are utilized by tourism operations.

Fig. 1 Location of 12 villages in the study area in the Okavango Panhandle, with the main elephant Loxodonta africana pathways shown. Paths are colour coded to show low (blue), medium (orange), and high (red) utilization gradients, based on the number of herds crossing the road over 3 years. Pathways were numbered from east to west (i.e. the path furthest to the east is EP1). The area shaded in black on the inset indicates the location of the Okavango Delta in Botswana.
Mean annual rainfall is 360–500 mm, and the rainy season is generally mid October–March. The mean monthly maximum temperature is 26.1–35.1°C, with July being the coldest month and October the hottest. Deep Kalahari sands are predominant throughout the study area but the area can be divided broadly into dry woodland savannah in the wildlife management areas NG11 & 13 and the Okavango wetland in NG12 (Fig. 1). Elevation is 880–1,110 m (Songhurst, Reference Songhurst2012).
The 2011 census recorded 16,306 people living in the eastern Panhandle (CSO, 2011), with 12 villages (population > 500), and additional settlements occurring between villages. Subsistence agriculture is an important livelihood strategy and, depending on rainfall, the planting of crops occurs during November–January and harvesting occurs during April–June. During the crop season many people move to their agricultural lands to guard their fields. The elephant population in the eastern Panhandle is estimated to be c. 15,000 and is considered trapped by the fences to the north, east and south, and the Okavango River to the west (Songhurst, Reference Songhurst2012). It has been observed that elephants use distinctive pathways to move between the Okavango Delta and foraging areas in dry woodland habitat in the study area (Jackson et al., Reference Jackson, Mosojane, Ferreira and van Aarde2008). These pathways are bisected by the main road that services all villages in the eastern Panhandle, which are distributed in a ribbon type development pattern along the edge of the wetland. Elephants damage crops, break fences, damage property and chase, injure and sometimes kill livestock and people. Meanwhile, people are modifying the elephants’ habitat through cultivating land and developing new settlements, and they also chase, injure and occasionally kill elephants. Crop damage is the most prevalent cause of human–elephant conflict in the area, but the number of crop raiding incidents (frequency) per month and the amount of damage incurred (intensity) per month vary annually (Songhurst, Reference Songhurst2012). During 2008–2010 788 fields were raided and a total of 162 ha of crops, predominantly millet, were damaged by elephants in the eastern Panhandle (Songhurst & Coulson, Reference Songhurst and Coulson2014).
Methods
It is difficult to observe elephants directly in the Panhandle, and therefore we used an indirect method based on examining footprints (Western et al., Reference Western, Moss and Georgiadis1983; Lee & Moss, Reference Lee and Moss1995) to estimate group size and composition, similar to the studies of Chiyo & Cochrane (Reference Chiyo and Cochrane2005) and Von Gerhardt et al. (Reference Von Gerhardt, Van Niekerk, Kidd, Samways and Hanks2014). Male and female elephant groups were distinguished by the presence or absence of footprints of elephants < 8 years old (footprint length ≤ 30 cm), which has been used successfully by Balasubramanian et al. (Reference Balasubramanian, Bhaskaran, Swaminathan, Desai, Daniel and Datye1995), Chiyo & Cochrane (Reference Chiyo and Cochrane2005) and Von Gerhardt et al. (Reference Von Gerhardt, Van Niekerk, Kidd, Samways and Hanks2014). Groups whose footprints were all > 30 cm were categorized as male-only groups (Lee & Moss, Reference Lee and Moss1995; Whitehouse & Hall-Martin, Reference Whitehouse and Hall-Martin2000; Moss, Reference Moss2001).
Ground surveys of the main road (180 km) were conducted on dry mornings (06.00–09.00), bi-monthly over 3 years (2008–2010) to assess recent elephant activity. Transects were driven in different directions each time to avoid bias. We recorded all elephant footprints crossing the transect, counted the number of elephant groups at each intersection between a pathway and the main road, and estimated the number of individuals per group, and their direction of travel. Group size was estimated by counting the number of different footprints present. This was achieved, in a similar way to Chiyo & Cochrane (Reference Chiyo and Cochrane2005), by following elephant tracks at crossings back along pathways to see how many elephants utilized those paths. Data were collected with the assistance of an experienced local tracker, who identified only fresh footprints and separated groups of elephants per observation. Stander et al. (Reference Stander, Ghau, Tsisaba, Oma and Ui1997) showed that local trackers, with their indigenous knowledge, were able to identify individuals by spoor with 94% accuracy. All footprints were cleared after counting to avoid double counting in subsequent assessments. Locations of the start and end of footprints on each intersection were recorded using a global positioning system, and elevation and vegetation type were recorded at the location of each footprint.
Although the substrata in the area comprise mostly fine Kalahari sands and alluvial clays, which show clear footprints, the identification of individual elephant tracks is difficult and therefore calculations of numbers of elephants per group are considered to be estimates. Subsequent analyses therefore used two response variables in generalized linear mixed-effect models: the number of herds using pathways, and the number of individuals using pathways.
Aerial orthophotographs (1 : 50,000) and Google Earth (Google Inc., Mountain View, USA) were used to digitize well-worn elephant paths between the dry mixed woodland habitat and the Okavango River and Delta, as well as human settlements, fences and cultivated land, using ArcMap v. 9.3 (ESRI, Redlands, USA). These maps were verified using a combination of methods: ground surveys; comparisons with existing maps based on data from collared elephants (Jackson et al., Reference Jackson, Mosojane, Ferreira and van Aarde2008), management plans (ODMP, 2002), and reports (Albertson, Reference Albertson1998); and consultations with local community members. Only permanent pathways, with floors devoid of vegetation and used regularly by elephants, were selected (Blake & Inkamba-Nkulu, Reference Blake and Inkamba-Nkulu2004). Regular use of pathways by elephants was confirmed through visual observation of dung, feeding damage and footprints (Von Gerhardt et al., Reference Von Gerhardt, Van Niekerk, Kidd, Samways and Hanks2014).
Data on human population numbers in the study area were collated from the Botswana Central Statistics Office (CSO, 2011). Songhurst (Reference Songhurst2012) showed that fields < 1 km from an elephant pathway are 50% more likely to be raided. A 1 km buffer was therefore calculated and drawn around each pathway, and the area of cultivated land within the buffer was calculated. The minimum distance between each pathway and a settlement or village was measured using the proximity buffer tool in ArcMap.
Statistical analysis
All analysis was carried out using R v. 2.11.1 (R Development Core Team, 2010). The characteristics of path use by herds of elephant (i.e. herd composition, direction of movement, favoured paths) were explored using χ2 tests, with significance determined at P < 0.05. Bi-monthly surveys of elephant pathway road crossings may have resulted in temporal pseudoreplication in the data, causing non-independence in errors and a higher risk of spuriously significant results (Hurlbert, Reference Hurlbert1990; Crawley, Reference Crawley2007). Generalized linear mixed-effect models are a useful tool for the analysis of such data (Pinheiro & Bates, Reference Pinheiro and Bates2000; Crawley, Reference Crawley2007). Models were fitted to the response variables ‘elephants present on pathway or not’ (coded 1 or 0) with binomial errors, ‘if elephants present, estimated number of herds per crossing’, and ‘if elephants present, estimated number of elephants per crossing’ with Poisson errors. The models were fitted with two crossed random effects: elephant pathway and month. Continuous explanatory variables included the area of cultivated land within a 1 km buffer of an elephant path, the distance from a path edge to the nearest settlement, and the elevation at the intersection between an elephant path and the main road. Continuous response and explanatory variables were square root transformed to conform better to a normal distribution where appropriate. Significance tests of the fixed effects were limited to two-way interactions, to avoid overfitted models, and model selection was based on the Akaike information criterion (AIC), as suggested by Whittingham et al. (Reference Whittingham, Stephens, Bradbury and Freckleton2006). All possible combinations of fixed effects and two-way interactions were explored, and models were compared using Akaike weights. The model with the highest Akaike weight (i.e. the weight of evidence in its favour) was considered to be the best model within the set (Burnham & Anderson, Reference Burnham and Anderson2002). Interactions were plotted to aid interpreting model predictions. A variance components analysis of the final mixed-effects models was carried out (Pinheiro & Bates, Reference Pinheiro and Bates2000; Crawley, Reference Crawley2007) to estimate the variation explained by the crossed random effects: elephant pathway and month.
Results
During 3 years we identified 106 elephant pathways crossing the main road transect (Fig. 1). A total of 12,836 elephant herd crossings were recorded during March 2008–June 2010, with a mean of 119 ± SD 255 herds crossing per pathway. We recorded significantly more family groups than male-only herds (χ2 = 171.7, df = 1, P < 0.001).
Both male herds and female-led family groups used certain pathways more frequently than others. One pathway (EP40) was utilized significantly more frequently throughout the year than all other pathways combined (χ2 = 8.8, df = 1, P < 0.001; Figs 1 & 2). Analysis was conducted with and without this pathway to assess its effect on results.
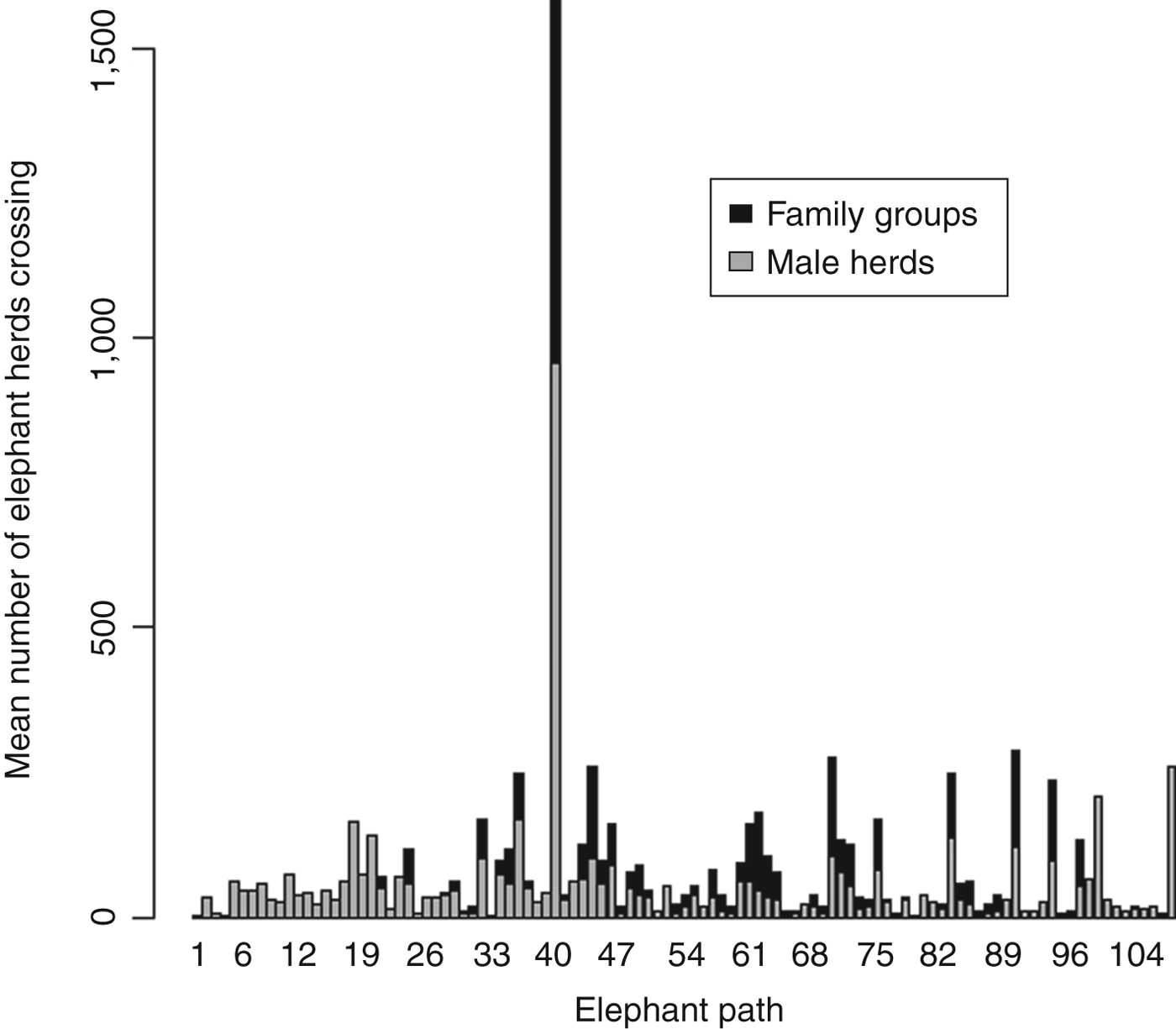
Fig. 2 Number of herds of family groups and male elephants that utilized each elephant pathway in the Okavango Panhandle (Fig. 1) during 2008–2010.
More crossings where elephants were travelling away from the river were recorded (χ2 = 84.7, df = 1, P < 0.001). To avoid double counting crossings by the same herd (i.e. counting their going to and coming from the river as two separate groups) only data from groups travelling away from the river (n = 6,757) were used in subsequent analyses.
The generalized linear mixed-effect model of whether elephants used a pathway or not, with the smallest AIC value (AIC = 3,010) and highest model weight (w i = 0.34) retained the main fixed effects elevation (maximum likelihood estimate = −5.8 ± SE 2.1) and human population (maximum likelihood estimate = − 4.7 ± SE 1.2), and the two-way interaction between these two effects (maximum likelihood estimate = 0.18 ± SE 0.07; Tables 1 & 2). This model shows there are likely to be elephants present when the human population in the nearest settlement is small, but the probability of elephants occurring when human populations are larger increases at higher elevation. The variance components of the random effects show that 38.8% of explained variation is between elephant paths and 61.2% is between months at each pathway.
Table 1 Fixed effects of the mixed effects model of whether elephants Loxodonta africana used a pathway or not, the number of elephant herds (if present) using a pathway, and the number of elephants (if present) using a pathway, with mean estimate ± SE, Z and P.
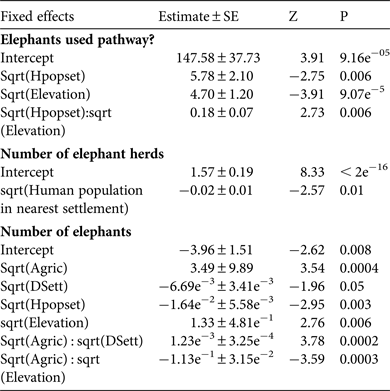
Table 2 Akaike information criteria (AIC) and Akaike model weights (w i, estimated probabilities of model truth) relating the probability of elephants using a path or not (coded 1 or 0), the likelihood of a larger number of herds (> 0) present at a pathway crossing, and the likelihood of a larger number of elephants (> 0) present at a pathway crossing to combinations of environmental and human habitat modification variables, for the top five models ranked according to AIC model weights.
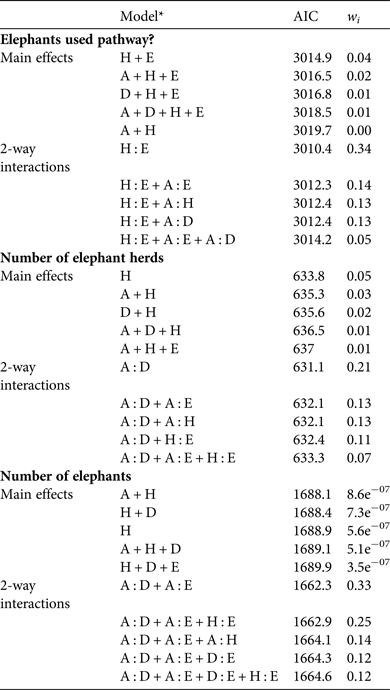
* A, sqrt(Area of agriculture within 1 km of path); E, sqrt(Elevation); D, sqrt(Distance to nearest settlement); H, sqrt(Human population in nearest settlement)
The model of the square root of the number of elephant herds (if present) using a pathway, with the smallest AIC value (AIC = 631.1) and highest model weight (w i = 0.21) retained the main fixed effects human population in the nearest settlement (maximum likelihood estimate = −0.01 ± SE 0.005), area of agriculture within a 1 km buffer of the path (maximum likelihood estimate = −0.03 ± SE 0.01), and distance to nearest human settlement (maximum likelihood estimate = −0.003 ± SE 0.003), and the two-way interaction between the latter two effects (maximum likelihood estimate = 0.0008 ± SE 0.0003; Tables 1 & 2). This model shows more herds are likely to cross on pathways near less populated settlements. There are also likely to be fewer herds crossing where there is a larger area of agriculture near a pathway and where pathways are close to human settlements, but the likelihood of more herds crossing when there are larger areas of agriculture increases with increasing distance to human settlements. Variance components show that 67.8% of variation is between pathways and 32.2% is between months at each pathway.
The model of the estimated number of elephants (if present) using a pathway, with the smallest AIC value (AIC = 1662.3) and highest model weight (w i = 0.33) retained the main fixed effects human population in nearest settlement (maximum likelihood estimate = −0.02 ± SE 0.006), elevation (maximum likelihood estimate = 1.33 ± SE 0.48), area of agriculture within a 1 km buffer of the path (maximum likelihood estimate = 3.49 ± SE 0.99), and distance to nearest human settlement (maximum likelihood estimate = −0.007 ± SE 0.003), and the two-way interaction between elevation and area of agriculture (maximum likelihood estimate = −0.11 ± SE 0.03) and distance to nearest settlement and area of agriculture (maximum likelihood estimate = 0.001 ± SE 0.0004; Tables 1 & 2). This model shows that, similar to number of herds, more elephants are more likely to cross on pathways near less populated settlements. There are also likely to be more elephants using a path if it is near larger areas of agriculture and further from settlements. Although it is likely that more elephants will use a path if it is near large areas of agriculture, this likelihood decreases with increasing elevation. Graphs of these interactions show a small amount of curvature, however, indicating that such relationships are unlikely to be biologically important. Variance components show that 53% of variation is between elephant pathways and 47% is between months at each pathway.
When all the models were fitted to a subset of data where crossings on pathway EP40 were removed, the model estimates for the fixed effects were not significantly affected.
Discussion
By mapping localized elephant pathways and monitoring fine-scale movement behaviour of an elephant population, rather than individual elephants, by means of detailed ground surveys we were able to record varying behavioural strategies adopted by the elephant population that contributed to risk avoidance, including avoiding pathways near human settlements, particularly those close to larger settlements, avoiding pathways close to large cultivated areas of land, and adopting a safety-in-numbers strategy when moving through large areas of cultivated land close to human settlements. Elephants in the eastern Panhandle of the Okavango Delta appear, therefore, to adapt their behaviour in response to associated levels of risk connected with humans, which influences their preferred choice of movement pathways.
Elephants generally avoided highly populated settlements and large areas of cultivated land, indicating a certain amount of risk associated with these areas of high human activity, which is consistent with the findings of other similar studies (Hoare & du Toit, Reference Hoare and du Toit1999; Douglas-Hamilton et al., Reference Douglas-Hamilton, Krink and Vollrath2005; Graham et al., Reference Graham, Douglas-Hamilton, Adams and Lee2009). The number of herds crossing in highly cultivated areas increased with distance from settlements, indicating elephants may be avoiding people. Areas of agricultural land appear to present a lower risk of disturbance than areas near settlements, thereby providing a safer route of passage and hence attracting larger numbers of elephant herds. The variance components analysis showed that the variation between the numbers of herds using various paths was affected more by the path itself (67.8%) than by the month (32.2%), indicating that the location of the path had a larger influence than seasonal variation on the number of herds utilizing a pathway. There were also differences in path use between female and male elephants, with some paths closer to areas of high human activity used only by groups of male elephants, indicating a higher level of risk taking by male-only groups (Sukumar, Reference Sukumar1991). We investigated a limited number of possible drivers of elephant movement, and differences in observed path use between the sexes warrant further investigation to determine the impact of other social and environmental factors. We found that path use by elephants was influenced by elevation, but a greater understanding of how resource availability affects the movement of elephants in the Panhandle is also needed.
We found a clear example of elephants aggregating in larger numbers near human activity, which could be a strategy to minimize risk (safety in numbers; Hamilton, Reference Hamilton1971). Pathways near cultivated land attracted fewer groups but larger numbers of elephants per group. This could be a result of group size being fluid, with families (or males) gathering into larger groups when entering high-risk areas, or perhaps only larger groups enter high-risk areas. Further investigation is needed to account for such differences but both scenarios suggest a behavioural adaptation to avoid risk.
Larger numbers of elephants per group using paths near cultivated land could also be a result of elephants being attracted to the high nutritional content in crops compared with natural forage, as surmised in previous studies (Sukumar, Reference Sukumar1989; Osborn, Reference Osborn1998; Chiyo et al., Reference Chiyo, Cochrane, Naughton and Basuta2005). If this were the case, however, we would have expected more elephant groups to have crossed in these areas, not necessarily more elephants per group. The temporal patterns of path use by elephants indicate that significantly more herds used paths closer to cropland after the crop season during May–November (Fig. 3), which indicates that many elephants may be avoiding cultivated areas during the ploughing season, thus adapting their temporal movement behaviour to avoid increased disturbance from human activity. Apparent seasonal variation in pathway use close to cultivated lands was consistent with our variance components analysis, which showed a higher percentage of variation between whether elephants used a path or not explained by month (61.2%) rather than pathway (38.8%). Not all elephants will be deterred from human disturbance, but there is no clear evidence to suggest that the elephant population is generally attracted to cultivated land in the eastern Panhandle. The likelihood of a field being raided by an elephant increases significantly when the field is within 1 km of a main pathway, suggesting that crop raiding incidents are predominantly opportunistic (Songhurst & Coulson, Reference Songhurst and Coulson2014). Affording elephant pathways protection with a 1 km development-free buffer zone could, therefore, play an important role in reducing crop raiding by elephants.

Fig. 3 Total number of elephant herds that crossed the main road in the Okavango Panhandle each month during 2008–2010. The horizontal lines show the median number of elephant herds crossing per month, the whiskers show 1.5 times the interquartile range, and the crosses are outliers.
Our results suggest that elephants have the ability to learn and adapt to varying levels of disturbance from humans, as other animals do in response to varying levels of predation risk (Laundré et al., Reference Laundré, Hernández and Ripple2010). Our findings support the suggestion by Guerbois et al. (Reference Guerbois, Chapanda and Fritz2012) to incorporate the so-called ecology of fear in the design of mitigation strategies for human–elephant conflict. According to Schoenfeld (Reference Schoenfeld1969) any avoidance response may be conditioned or controlled by an appropriate selection of values of three parameters, among others: operant level of the response, stimulus frequency, and stimulus intensity in both positive and negative reinforcement schedules. The various types of avoidance behaviour we observed suggest a response to negative stimulus from human encounters, and therefore risk avoidance by elephants could be conditioned or controlled by increasing or intensifying a negative stimulus associated with human inhabitation or development zones. Appropriate deterrent techniques, such as the use of capsicum oleoresin (Osborn, Reference Osborn2002; Sitati & Walpole, Reference Sitati and Walpole2006; Hedges & Gunaryadi, Reference Hedges and Gunaryadi2010) or bees (King et al., Reference King, Douglas-Hamilton and Vollrath2011), could intensify the negative stimulus to deter elephants at interfaces between pathways and arable fields. We argue that in combination with appropriate land use zones, such negative stimuli could be effective in alleviating spatial and resource use overlap at a landscape level, reinforcing boundaries around human-inhabited areas and ensuring safe passage for elephants.
Correctly managed buffers or adaptive management zones outside protected areas may be as important as wildlife reserves for the long-term viability of wide-ranging species (Noss et al., Reference Noss, Quigley, Hornocker, Merrill and Paquet1996). Such strategies require creating zones within a multi-use landscape to minimize negative interactions between people and wildlife (Fernando et al., Reference Fernando, Wikramanayake, Weerakoon, Jayasinghe, Gunawardene and Janaka2005; Linnell et al., Reference Linnell, Nilsen, Lande, Herfindal, Odden, Skogen, Woodroffe, Thirgood and Rabinowitz2005). Knowledge of their risk avoidance behaviour may be used to encourage elephants to use certain pathways when moving through human inhabited areas, and deter them from fields and settlements. Landscape-level management interventions could help define boundaries at the human–wildlife interface, minimizing opportunistic crop raiding by elephants, while protecting critical movement paths and habitat connectivity. We recommend monitoring elephant movements at a population level, as we have done, in further studies of the effectiveness of protecting elephant pathways in reducing human–elephant conflict. Long-term monitoring of path use with and without negative stimuli at the interface between development-free buffer zones and agricultural or settlement zones is necessary to reveal the potential of combining pathway buffers and mitigation techniques as a strategy to avoid human–elephant conflict. Such investigations could inform and motivate appropriate land use planning initiatives, but it will be essential to galvanize the political will among local and national land authorities to implement such measures.
Acknowledgements
We thank the Botswana Ministry of Environment, Wildlife & Tourism for granting permission for this research, and the Wildlife Conservation Society and Wilderness Wildlife Trust for providing funding. We also thank the ground surveying team. Tim Coulson acknowledges support from an ERC Advanced Grant, LEED.
Biographical sketches
Anna Songhurst is a conservation biologist interested in the conservation and management of wildlife, in particular elephants. Graham McCulloch is an ecologist and biodiversity specialist with particular interest in the sustainable management of biodiversity inside and outside protected areas. Tim Coulson is a quantitative ecologist, and his research interests are conservation, wildlife management and population biology.





