
25 results
Interacting with others while reacting to the environment
-
- Journal:
- Behavioral and Brain Sciences / Volume 45 / 2022
- Published online by Cambridge University Press:
- 07 July 2022, e106
-
- Article
- Export citation
SIMPLY CONNECTED, SPINELESS 4-MANIFOLDS
- Part of
-
- Journal:
- Forum of Mathematics, Sigma / Volume 7 / 2019
- Published online by Cambridge University Press:
- 17 May 2019, e14
-
- Article
-
- You have access
- Open access
- Export citation
-
We construct infinitely many compact, smooth 4-manifolds which are homotopy equivalent to
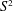 $S^{2}$ but do not admit a spine (that is, a piecewise linear embedding of
$S^{2}$ but do not admit a spine (that is, a piecewise linear embedding of 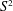 $S^{2}$ that realizes the homotopy equivalence). This is the remaining case in the existence problem for codimension-2 spines in simply connected manifolds. The obstruction comes from the Heegaard Floer
$S^{2}$ that realizes the homotopy equivalence). This is the remaining case in the existence problem for codimension-2 spines in simply connected manifolds. The obstruction comes from the Heegaard Floer 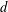 $d$ invariants.
$d$ invariants.
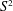
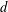
NONSURJECTIVE SATELLITE OPERATORS AND PIECEWISE-LINEAR CONCORDANCE
- Part of
-
- Journal:
- Forum of Mathematics, Sigma / Volume 4 / 2016
- Published online by Cambridge University Press:
- 23 December 2016, e34
-
- Article
-
- You have access
- Open access
- Export citation
-
We exhibit a knot
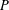 $P$ in the solid torus, representing a generator of first homology, such that for any knot
$P$ in the solid torus, representing a generator of first homology, such that for any knot 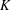 $K$ in the 3-sphere, the satellite knot with pattern
$K$ in the 3-sphere, the satellite knot with pattern 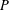 $P$ and companion
$P$ and companion 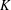 $K$ is not smoothly slice in any homology 4-ball. As a consequence, we obtain a knot in a homology 3-sphere that does not bound a piecewise-linear disk in any homology 4-ball.
$K$ is not smoothly slice in any homology 4-ball. As a consequence, we obtain a knot in a homology 3-sphere that does not bound a piecewise-linear disk in any homology 4-ball.
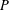
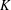
Optical properties of defects in nitride semiconductors
-
- Journal:
- Journal of Materials Research / Volume 30 / Issue 20 / 28 October 2015
- Published online by Cambridge University Press:
- 23 September 2015, pp. 2977-2990
- Print publication:
- 28 October 2015
-
- Article
- Export citation
Contributors
-
-
- Book:
- The Cambridge Dictionary of Philosophy
- Published online:
- 05 August 2015
- Print publication:
- 27 April 2015, pp ix-xxx
-
- Chapter
- Export citation
Notes on contributors
-
-
- Book:
- William Wordsworth in Context
- Published online:
- 05 February 2015
- Print publication:
- 12 February 2015, pp ix-xvi
-
- Chapter
- Export citation
Early Treatment of a Progressive Rasmussen's Like Syndrome with Ganciclovir
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 38 / Issue 2 / March 2011
- Published online by Cambridge University Press:
- 02 December 2014, pp. 296-298
-
- Article
-
- You have access
- Export citation
Contributor affiliations
-
-
- Book:
- Textbook of Neural Repair and Rehabilitation
- Published online:
- 05 May 2014
- Print publication:
- 24 April 2014, pp ix-xvi
-
- Chapter
- Export citation
Contributor affiliations
-
-
- Book:
- Textbook of Neural Repair and Rehabilitation
- Published online:
- 05 June 2014
- Print publication:
- 24 April 2014, pp ix-xvi
-
- Chapter
- Export citation
In-Focus Electrostatic Zach Phase Plate Imaging for Transmission Electron Microscopy with Tunable Phase Contrast of Frozen Hydrated Biological Samples
-
- Journal:
- Microscopy and Microanalysis / Volume 20 / Issue 1 / February 2014
- Published online by Cambridge University Press:
- 02 January 2014, pp. 175-183
- Print publication:
- February 2014
-
- Article
- Export citation
Social-ecological systems as complex adaptive systems: modeling and policy implications
-
- Journal:
- Environment and Development Economics / Volume 18 / Issue 2 / April 2013
- Published online by Cambridge University Press:
- 27 November 2012, pp. 111-132
-
- Article
- Export citation
Contributors
-
-
- Book:
- Trauma
- Published online:
- 07 September 2011
- Print publication:
- 16 June 2011, pp ix-xiv
-
- Chapter
- Export citation
Psychiatric symptoms in Ground Zero ironworkers in the aftermath of 9/11: prevalence and predictors
-
- Journal:
- Psychiatric Bulletin / Volume 33 / Issue 2 / February 2009
- Published online by Cambridge University Press:
- 02 January 2018, pp. 49-52
- Print publication:
- February 2009
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Five - Falling Between Two Stools: How womens land rights are lost between state & customary law in Apac District, Northern Uganda
-
-
- Book:
- Women's Land Rights and Privatization in Eastern Africa
- Published by:
- Boydell & Brewer
- Published online:
- 03 March 2023
- Print publication:
- 20 November 2008, pp 101-120
-
- Chapter
- Export citation
Afterword: Securing womens land rights
-
-
- Book:
- Women's Land Rights and Privatization in Eastern Africa
- Published by:
- Boydell & Brewer
- Published online:
- 03 March 2023
- Print publication:
- 20 November 2008, pp 158-175
-
- Chapter
- Export citation
Economic growth, carrying capacity, and the environment*
-
- Journal:
- Environment and Development Economics / Volume 1 / Issue 1 / February 1996
- Published online by Cambridge University Press:
- 31 July 2008, pp. 104-110
-
- Article
- Export citation
Introduction: Infectious diseases
-
- Journal:
- Environment and Development Economics / Volume 12 / Issue 5 / October 2007
- Published online by Cambridge University Press:
- 04 October 2007, pp. 625-626
-
- Article
- Export citation
6 - Variation in Susceptibility: Lessons from an Insect Virus
-
-
- Book:
- Adaptive Dynamics of Infectious Diseases
- Published online:
- 15 January 2010
- Print publication:
- 11 April 2002, pp 74-84
-
- Chapter
- Export citation
Use of a routine, self-report outcome measure (HoNOSCA–SR) in two adolescent mental health services
-
- Journal:
- The British Journal of Psychiatry / Volume 180 / Issue 3 / March 2002
- Published online by Cambridge University Press:
- 02 January 2018, pp. 266-269
- Print publication:
- March 2002
-
- Article
-
- You have access
- HTML
- Export citation
Anticipating environmental disasters
-
- Journal:
- Environment and Development Economics / Volume 3 / Issue 4 / October 1998
- Published online by Cambridge University Press:
- 02 April 2001, pp. 491-537
-
- Article
- Export citation
