


The eye disorder cataract has been a relatively common observed symptom in the fish farming history, related to environmental, genetic, infectious, toxicological and nutritional factors(Reference Bjerkås, Breck and Waagbø1, Reference Hargis2). Cataracts in Atlantic salmon smolts have been observed in European aquaculture for the last 20 years, and an epidemiological survey performed in 1998 in seawater farms along the Norwegian coast found a cataract prevalence as high as 82 %(Reference Ersdal, Midtlyng and Jarp3). The losses for the aquaculture industry vary and can be considerably high in more severe seasons(Reference Menzies, Crockford and Breck4). Cataracts have lately also been observed in salmon growers (>1·5 kg) in their second year in seawater. Besides being an important ethical welfare issue for the fish farming industry, cataract development in the final phase of salmon production may also represent economical losses from reduced growth, reduced feed efficiencies, secondary infectious diseases and quality down grading at slaughter(Reference Menzies, Crockford and Breck4).
The amino acid histidine has been identified as a key preventative factor in the development of cataract in Atlantic salmon (Salmo salar L.) smolts(Reference Bjerkås and Sveier5–Reference Breck, Bjerkås and Campbell7). The type of cataract that occurred in the nineties at high incidences and severities in farmed salmon smolts was probably caused by the omission of blood meal from commercial salmon diets(Reference Wall8). Blood meal was banned as feed ingredient as a measure to prevent the spread of bovine spongiform encephalopathy(9). Blood meal is especially rich in histidine (approximately 50 g histidine/kg in spray-dried products)(10). Breck et al. (Reference Breck, Bjerkås and Campbell6) showed that feed could be supplemented with histidine either in the form of blood meal, high histidine fishmeal or as a pure histidine additive, and that all forms prevented cataract development in salmon smolts equally well.
The exact mechanism for the cataract mitigation by elevated dietary histidine level is not completely understood. It has become clear that histidine and related compounds (imidazoles) cover many important biochemical roles in the salmon tissues besides its function as essential amino acid in protein synthesis, such as components related to lens osmoregulation(Reference Baslow11, Reference Breck, Bjerkås and Sanderson12), muscle pH buffering(Reference Ogata13, Reference Suzuki, Hirano and Suyama14), tissue antioxidation(Reference Kim, Park and Choi15, Reference Wade and Tucker16) and detoxification of cytotoxic-reactive carbonyl species(Reference Aldini, Carini and Beretta17, Reference Hipkiss18).
The smoltification phase in salmonids has been considered as a risk period for cataract development(Reference Breck, Bjerkås and Campbell7, Reference Wall8). Muscle histidine concentrations recorded during smoltification in wild salmon demonstrated changes in the pattern of imidazoles at or after the transfer from freshwater to seawater. This was assumed to reflect the introduction of anaerobic burst swimming in seawater with increased need for muscle pH buffering(Reference Ogata, Konno and Silverstein19, Reference Munakata, Aida and Amano20). The major histidine compound in salmonid white muscle is the dipeptide anserine, which occurs at high concentration after smoltification(Reference Breck, Bjerkås and Campbell7, Reference Munakata, Aida and Amano20). The biosynthesis of anserine (β-alanyl-3-methyl-histidine) includes methylation of the precursor carnosine (β-alanyl-l-histidine), which occurs at much lower concentrations in the muscle tissue. The cataract preventative action of histidine in salmon smolts seems, however, to be mediated through the derivative N-acetyl-histidine (NAH) in the lens(Reference Baslow11, Reference Breck, Bjerkås and Sanderson12). Lens NAH occur in high concentration ranges as same as anserine in white muscle, and the concentration depends on the dietary histidine levels before and during smoltification. Low lens NAH concentration correlated with the development of severe cataract after transferring to seawater(Reference Breck, Bjerkås and Campbell7). From the results obtained in the subsequent feeding experiments, Breck et al. (Reference Breck, Bjerkås and Sanderson12) suggested a rapid synthesis and turnover of NAH in the lens to cope with osmotic challenges, supporting a proposed model of this histidine compound as an important lens osmolyte(Reference Baslow11, Reference Baslow21).
Histidine deficiency has not been regarded as an area of concern, beyond the observed reductions in growth and feed efficiency in young Atlantic salmon fed histidine below the established minimum requirement of 7 g/kg(Reference Hardy, Lim and Webster22). The underlying experiments did not, however, report on free imidazoles in tissue as markers for histidine status. Feeding experiments with Atlantic salmon smolts fed diets with normal (11·7 g/kg) and high (17·6 g/kg) levels of histidine showed that both muscle anserine and lens NAH concentrations depended directly on dietary histidine inclusion level, despite the fact that these dietary histidine levels far exceeded the established requirement for salmonids(Reference Breck, Bjerkås and Campbell7). The requirement of histidine for adult Atlantic salmon growers under practical conditions is not known, but it is normally suggested to be lower than for smolts, in line with a general lower relative protein requirement in older fish.
The main hypothesis of the present experiment was that dietary histidine exerts cataract preventive effect in Atlantic salmon growers depending on both the dietary concentration and feeding period. A feeding study with Atlantic salmon during their second year in seawater was performed at an experimental sea farming site in Southern Norway with a previous history of cataract development. The experimental design included three dietary histidine levels. Two changes in diet during the experimental period (crossover design) were included to identify critical periods of histidine feeding. The experiment was supported by assessment of somatic data, eye inspection using a slit lamp biomicroscope and analyses of free amino acids in muscle and lens tissues.
Materials and methods
Adult Atlantic salmon (Salmo salar L.) were acclimatised for 2 weeks in two seawater net pens at Lerang Research Station (Skretting ARC, Rogaland, Norway), before being divided randomly into six experimental net pens (5 m2). Mean body weight of the 1834 individually passive integrated transponder-tagged (Electronic I, Inc., Dallas, TX, USA) salmon at the start of the experiment was 1662 (sd 333) g. Three fishmeal and fish oil-based extruded high-energy diets (protein: 375 g/kg and fat: 342 g/kg), differing only in histidine content (low (L) 9·3, medium (M) 12·8 and high (H) 17·2 g histidine/kg diets), were fed to duplicate net pens by the use of feeding automats and hand feeding according to in-house tables considering biomass and water temperature. The three experimental diets were produced by Skretting Aquaculture Research Center for the whole feeding period in one production using commercially available feed ingredients (Table 1). Vitamins and minerals were supplemented as a commercial premix fulfilling the requirements for salmonids given by the Norwegian Research Council(10). Proximate composition (Table 1) was determined by an in-house near-IR spectroscopy analysis. The data obtained from near-IR spectroscopy methods were calibrated and checked by accredited conventional Association of Official Analytical Chemists methods (1995). Feed amino acid profiles (Table 1) were analysed at Skretting ARC according to an European Union standard method(23).
Table 1 Ingredients, proximate composition and amino acid profiles (g/kg) of the extruded experimental diets, varying only in histidine (low (L): 9·3, medium (M): 12·8 and high (H): 17·2 g histidine/kg diet)

DE, digestible energy; NA, not analysed.
* LT Skandinavia, Chr Holtermann AS, Norway.
† Statkorn, Oslo, Norway.
‡ Denofa, Fredrikstad, Norway.
§ Nordsildmel, Bergen, Norway.
∥ Vitamin and mineral premix; Skretting, Stavanger, Norway (fulfilling recommendations for salmonids given by Norwegian Research Council(Reference Breck, Bjerkås and Campbell10)).
¶ Kyowa Hakko, Tokyo, Japan.
** Calculated DE (protein: 18 kJ/g; lipid: 33·5 kJ/g; carbohydrates: 12·5 kJ/g).
The experimental period from June to October was divided into three seasons ((June 6–July 14 (6 weeks), July 17–September 6 (13 weeks) and September 7–October 12 (18 weeks)), each starting and ending with eye examination for cataract (slit lamp biomicroscopy), assessment of somatic data (body weight, length and condition factor), sampling of lens and muscle tissues for analysis of histidine and histidine derivatives and gene expression analyses.
Approximately, 200 individuals were allocated to each net pen belonging to L and H histidine groups, while the replicates of M histidine groups were housed each with eighty fish. Within respective replicates, approximately thirty fish fed L-histidine feed were transferred (crossed over) to M (group only receiving fish) and H histidine dietary groups at the samplings in July and September. Similarly, thirty fish fed high-histidine feed were transferred (crossed over) to M and L-histidine dietary groups at the samplings in July and September; ending up with seven dietary groups in the second period (LL, LM, LH, MM, HL, HM and HH) and eleven experimental groups in the third period (LLL, HLL, HHL, LLM, LMM, MMM, HMM, HHM, LLH, LHH and HHH) of approximately similar size, but varying in histidine level and feeding period. Fig. 1 details the experimental design with movement of fish. Since the M dietary groups received fish only during the two crossovers, they were housed with fewer fish at the start; meaning that all the net pens would have approximately equal biomass in the course of the trial.
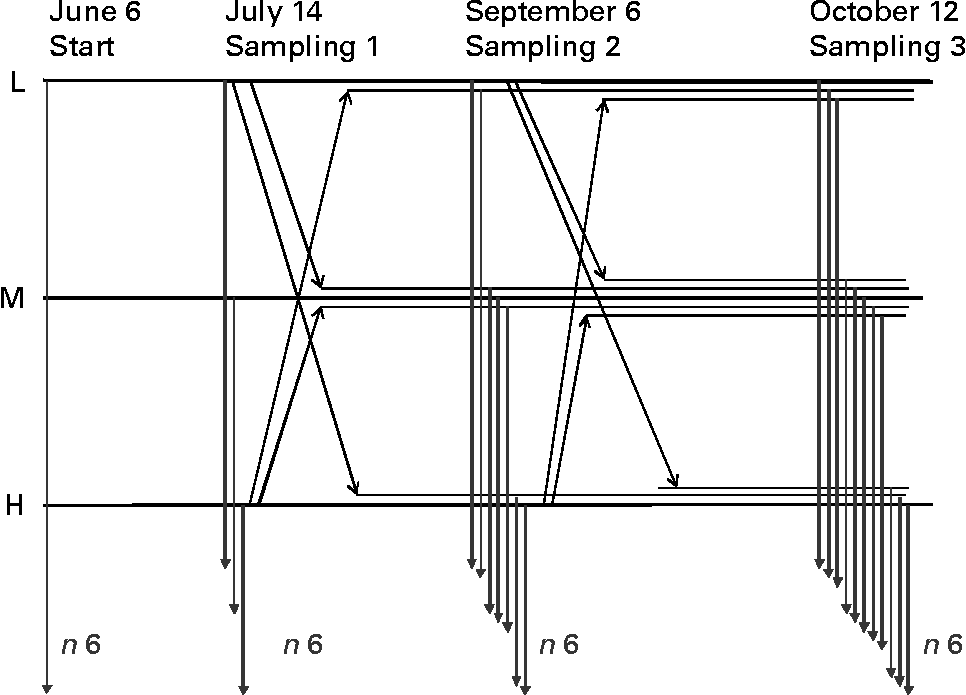
Fig. 1 Details of the experimental design run in duplicate net pens in seawater. Experimental feeds with low (L: 9·3 g/kg), medium (M: 12·8 g/kg) and high (H: 17·2 g/kg) histidine were fed during three periods, each ending with assessment of biological somatic performance data (thirty individuals per feed replicate), cataract examination (thirty per feed replicate) and tissue sampling (six per feed replicate). At samplings in July and September, individually marked fish from L and H were transferred to respective replicates of L, M and H according to a crossover design, ending up with combinations of seven (LL, LM, LH, MM, HL, HM and HH) and eleven (LLL, LLM, LLH, LMM, LHH, MMM, HLL, HMM, HHL, HHM and HHH) dietary groups.
The fish were exposed to a natural light regime in seawater (mean salinity of 30 g/l). Water temperature (4 m depth) rose from 12 to 18·5°C from June to July, and declined slowly to 16°C in the end of September and further declined to 14·4°C at the end of October. Water oxygen concentration measured at regular intervals was never below 8·2 mg/l.
The Lerang Research Station is approved, and the trial was conducted according to the guidelines of the Norwegian State Commission for Laboratory Animals.
Cataract examination
When establishing the six experimental groups, sixty fish from the two acclimatisation net pens were anaesthetised (metacain was added to the sampling tray according to an in-house protocol), weighed, measured and examined for cataract using a slit lamp biomicroscope at 15 × magnification. The fish were inspected and scored individually for changes (0–4 per eye; i.e. 0–8 for both the eyes per fish) according to the previously described procedure(Reference Wall and Bjerkås24). Fish with visual physical eye damages were excluded from the experiment. At later examinations (July 15, September 5 and October 12), thirty fish per replicate (n 60 per dietary treatment) from all the experimental groups were examined according to the same procedure. The examined fish, except six taken for sampling, were returned to their respective net pens.
Samplings
Initially (June 6), and at July 15, September 5 and October 12, muscle and lens tissues of six fish were sampled per replicate (n 12 per dietary group) for analysis of histidine compounds and gene expression profiling(Reference Tröße, Waagbø and Breck25). The fish were fasted for 1 d before sampling. Individual fish were anaesthetised, inspected for cataract, measured for weight and length (n 30 per replicate), and sampled for lens and muscle tissues (n 6 per replicate). Skin-free epaxial white muscle tissue was sampled from behind the dorsal fin and was frozen directly on dry ice. Lenses were carefully dissected, gently rolled on clean filter paper to remove liquid and muscle attachments, immediately frozen on liquid nitrogen/dry ice, and stored at − 80°C until analysis.
Analysis of free amino acids
Individual muscle samples of six fish were analysed for free amino acids, while individual lenses were analysed for histidine and NAH. Free amino acids were analysed using a Biochrom 20 Plus Amino Acid Analyser (Amersham, Cambridge, UK), according to a standardised procedure from the manufacturer (Biochrom AAAFAQ08), with post-column ninhydrin derivatisation and colorimetric detection at 570 and 440 nm. NAH and free histidine in the lens were analysed by reverse phase HPLC and UV detection at 210 nm(Reference O'Dowd, Cairns and Trainor26), with modifications according to Breck et al. (Reference Breck, Bjerkås and Campbell7).
Statistical analyses
The data were analysed by the CSS:Statistica for Windows™ Statistical Software (version 9.0; Statsoft, Inc., Tulsa, OK, USA, 1984–9). Differences between groups were assessed by nested ANOVA (net pen individuals nested in dietary groups) and Tukey's honestly significant difference post hoc test at a level of P < 0·05. All values are given as mean values with their standard errors if not stated otherwise.
Results
Growth and mortality
There were no systematic differences in somatic data related to dietary histidine regimes at any samplings (Tables 2 and 3). In the first period, the fish grew from 1662 g (mean values; n 56–58 fish from two replicates) to 2251 (sem 62), 2135 (sem 61) and 2200 (sem 59) g in the L, M and H histidine groups, respectively (nested ANOVA, P>0·05). Similarly, no differences in condition factor were observed, ranging from 1·17 to 1·18; neither were any difference in growth and somatic data observed in the second period, with mean body weight ranging between 2361 and 2652 g. At the conclusion of the trial, fish continuously fed on the high-histidine diet (HHH) were significantly heavier than fish fed continuously on the M-histidine diet (MMM) and the crossover diet (HMM) (nested ANOVA, P < 0·05). There were, however, no differences in the length (range of means from 60·7 to 63·1 cm) and condition factor (range of means from 1·18 to 1·26) between the groups. Excluding the sexually mature individuals recorded at the samplings (18·5 % in September and 28 % in October) improved the mean individual specific growth rate (0·4–0·5 %/d) in the dietary groups, but did not change the growth results relative to dietary histidine regimes.
Table 2 Somatic data from duplicate groups of adult Atlantic salmon fed three levels of dietary His for 5 weeks before a massive outbreak of cataract
(Mean values with their standard errors)

L, low; M, medium; H, high.
* The single capital letter codes indicate the use of diets L, M or H in the period from June to July.
† Initial values: weight, 1662 g (n 1834).
‡ The data were evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test (NS: P>0·05).
Table 3 Somatic data from duplicate groups of adult Atlantic salmon fed three levels of dietary His levels for 5 months*
(Mean values with their standard errors)

L, low; M, medium; H, high.a,b Mean values within a column with unlike superscript letters were significantly different (P < 0·05).
* The experimental period was divided into three seasons (the three capital letter code indicates diets L, M and H used in June–July, July–September and September–October, respectively).
† Three-letter codes indicate the use of diets L, M or H in the first, second and third periods.
‡ The data were evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test (NS: P>0·05).
Cataract assessment
At the start of the experiment, minor changes in one eye (score 1) was observed in sixteen fish, two fish showed score 2, and one individual was totally blind (score 8), ending with a cataract prevalence of 30 % and a mean score of 0·4 of sixty-three inspected fish. A similar overall outcome was observed in July, with approximately 30 % cataracts in the fish population. All the groups showed very mild cataract changes, with no difference in incidence and mean cataract scores ( ≤ 0·7; Fig. 2).

Fig. 2 Development of cataracts (mean cataract score of both eyes; 0–8) in Atlantic salmon fed the experimental histidine diets: low (L), medium (M) and high (H) (see Fig. 1) during three periods from June to October (mean of thirty examined fish per experimental duplicate). a,b,c Mean values with unlike letters were significantly different (P < 0·05; nested ANOVA and Tukey's honestly significant difference post hoc test). ![]() , LLL;
, LLL; ![]() , LLM;
, LLM; ![]() , LLH;
, LLH; ![]() , LMM;
, LMM; ![]() , LHH;
, LHH; ![]() , MMM;
, MMM; ![]() , HLL;
, HLL; ![]() , HMM;
, HMM; ![]() , HHL;
, HHL; ![]() , HHM;
, HHM; ![]() , HHH.
, HHH.
From July to September, a major outbreak of cataracts was recorded, with mean cataract scores reflecting the previous dietary histidine feeding regimes. Fish groups fed a diet that was not supplemented with histidine (L) from June to July developed significantly more cataract (mean score of 4·4 (sem 0·2)) than fish fed M- or H-histidine diets, both with scores < 1·5 (P < 0·05). The cataract scores were also influenced by the feeding levels of histidine from July to September, where fish crossed over from L to H histidine diet (LH) showed significantly lower cataract score (2·9 (sem 0·2)) than the L histidine group (LL), while fish crossed over from L to M histidine diet (LM) showed an intermediate higher score (3·7 (sem 0·2)). No further development in cataract was observed from September to October in any group, ending with approximately similar cataract severities as observed in September. All the groups fed diets supplemented with histidine during the first feeding periods (H, M and respective crossover diets) showed low but moderately increased cataract scores from the start (0·4) to the end ( ≤ 2·0; Fig. 2).
Considering all individually examined fish at the end of the experiment, no difference in cataract scores was observed between sexually mature fish and immature fish.
Free amino acid composition in muscle
Free histidine in white muscle tissue reflected systematically the feed histidine regimes, both in dietary concentration and feeding time (Fig. 3). In July, before the cataract outbreak, there were clear differences between the groups fed the L (0·13 μmol/g)-, M (0·70 μmol/g)- and H (1·86 μmol/g)-histidine diets (P < 0·05), while in September after the outbreak, muscle histidine levels were closer in concentration with significantly higher concentrations in fish continuously fed H-histidine diet (HH) than in fish continuously fed the L-histidine diet (LL). Fish crossed over to the H (LH)- and L-histidine diets (HL) showed intermediate muscle histidine levels. In October, after a second crossover, larger and systematic variations were observed among the eleven dietary groups; largely dividing into three concentration levels and reflecting the dietary histidine levels during the final period (Fig. 3).

Fig. 3 Development of white muscle histidine in Atlantic salmon fed the experimental histidine diets: low (L), medium (M) and high (H) during three periods from June to October. Data are expressed as means with their standard errors (six per experimental duplicate; see Fig. 1). a–d Mean values with unlike letters were significantly different (P < 0·05; nested ANOVA and Tukey's honestly significant difference post hoc test). ![]() , LLL;
, LLL; ![]() , LLM;
, LLM; ![]() , LLH;
, LLH; ![]() , LMM;
, LMM; ![]() , LHH;
, LHH; ![]() , MMM;
, MMM; ![]() , HAA;
, HAA; ![]() , HMM;
, HMM; ![]() , HHL;
, HHL; ![]() , HHM;
, HHM; ![]() , HHH.
, HHH.
White muscle anserine concentration constituted between 45 and 50 % of the free amino acids and was less variable (6–8 % difference) among the dietary histidine groups (Table 4; Fig. 4). Muscle anserine was significantly higher in fish fed M (17·0 μmol/g)- and H (16·7 μmol/g)-histidine diets, than in fish fed the unsupplemented histidine diet (15·7 μmol/g). Muscle anserine increased moderately from July to September in all the groups, while for fish crossed over from L to H histidine diets (LH), this increase was significant. Moderate declines in muscle anserine were apparent from September to October, mostly in fish continuously fed the L-histidine diet (LLL; Fig. 4).
Table 4 Free amino acids in white muscle tissue (μmol/g) of adult Atlantic salmon fed low (L; 9·3 g/kg), medium (M; 12·8 g/kg) and high (H; 17·2 g/kg) levels of histidine sampled in July (week 6) before a serious outbreak of cataract*

ND, below the detection limit.a,b,c Mean values within a row with unlike superscript letters were significantly different (P < 0·05).
* Six samples per duplicate; evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test at a level of P < 0·05.
† Single-letter codes indicate the use of diets L, M or H in the first feeding period.
‡ Pooled sem < 0·005.

Fig. 4 Development of white muscle anserine in Atlantic salmon fed the experimental histidine diets: low (L), medium (M) and high (H) during three periods from June to October. Data are expressed as means with their standard errors (six per experimental duplicate; see Fig. 1). a,b,c Mean values with unlike letters were significantly different (P < 0·05; nested ANOVA and Tukey's honestly significant difference post hoc test). ![]() , LLL;
, LLL; ![]() , LLM;
, LLM; ![]() , LLH;
, LLH; ![]() , LMM;
, LMM; ![]() , LHH;
, LHH; ![]() , MMM;
, MMM; ![]() , HLL;
, HLL; ![]() , HMM;
, HMM; ![]() , HHL;
, HHL; ![]() , HHM;
, HHM; ![]() , HHH.
, HHH.
Muscle carnosine was significantly higher in the M than in the L histidine group at the first sampling in July (Table 4), and declined in all the groups in September (Table 5) except in fish crossed over from L to M histidine diet (LM), however, with no significant differences between the groups. Finally, carnosine increased during the last feeding period in all the groups except those fed the L-histidine diet during the final period ( < 0·2 μmol/g). Muscle carnosine was four times higher in fish crossed over from L to H histidine diet after the second period (LLH; P < 0·05).
After 6 weeks of feeding, several free amino acids in white muscle tissue, including some essential amino acids (threonine, isoleucine, leucine and lysine), were significantly higher in salmon fed the L-histidine diet compared with fish fed histidine-supplemented diets (M and H; Table 4). The sum of free amino acids was, however, the same in all the dietary groups. After 13 weeks and following the cataract outbreak, there were minor differences among the other free amino acids in muscle than the imidazoles, with non-systematic differences in O-phospho-ethanolamine, aspartic acid, glutamine, alanine, tyrosine and ammonia (P < 0·05) among the groups (Table 5). Again, total amino acid concentration was equal in the samplings, between 34 and 35 μmol/g. At the end of the trial, a profile similar to previous free amino acid profile in muscle was observed (data not shown), with significant higher concentration of selected amino acids (hydroxyproline, proline, threonine, glutamate, tyrosine, phenylalanine, ornithine, 1-methyl histidine) in salmon fed the L-histidine diet during the two last feeding periods (HLL and LLL groups) compared with the other groups. Total amino acid concentration was equal within the sampling, but somewhat lower than that observed previously (31–33 μmol/g).
Table 5 Free amino acids in white muscle tissue (μmol/g) of adult Atlantic salmon fed low (L; 9·3 g/kg), medium (M; 12·8 g/kg) and high (H; 17·2 g/kg) levels of His in two periods and sampled in September after a serious outbreak of cataract*

ND, below the detection limit.a–e Mean values within a row with unlike superscript letters were significantly different (P < 0·05).
* Six samples per duplicate; evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test at a level of P < 0·05.
† Two-letter codes indicate the use of diets L, M or H in the first and second feeding periods.
‡ Pooled sem < 0·005.
Lens amino acid composition
As in the white muscle, lens histidine and NAH reflected dietary histidine concentrations strictly systematically relative to both concentration and feeding period (Figs. 5 and 6). Large differences in the concentration of NAH between the feeding groups was observed in July before the cataract outbreak, with significant differences (P < 0·05) between all the three groups (Fig. 6). All the dietary groups had increased lens NAH from July to September, except fish crossed over from a H to L histidine diet (HL). The lens NAH increases were most significant in fish with a low NAH status in July, and depended on dietary histidine concentration in this second feeding period. In August, the lens NAH status correlated negatively with severity of cataract given as cataract score (Fig. 7). At the end of the trial, lens NAH showed a 3-fold difference between the groups, depending on dietary histidine feeding regimes (Fig. 6). The H-histidine diet obviously supported saturation level of lens NAH at 11–12 μmol/g.

Fig. 5 Development of lens histidine in Atlantic salmon fed the experimental histidine diets: low (L), medium (M) and high (H) during three periods from June to October. Data are expressed as means with their standard errors (six per experimental duplicate; see Fig. 1). a,b,c,d Mean values with unlike letters were significantly different (P < 0·05; nested ANOVA and Tukey's honestly significant difference post hoc test). ![]() , LLL;
, LLL; ![]() , LLM;
, LLM; ![]() , LLH;
, LLH; ![]() , LMM;
, LMM; ![]() , LHH;
, LHH; ![]() , MMM;
, MMM; ![]() , HLL;
, HLL; ![]() , HMM;
, HMM; ![]() , HHL;
, HHL; ![]() , HHM;
, HHM; ![]() , HHH.
, HHH.

Fig. 6 Development of lens N-acetyl-histidine (NAH) in Atlantic salmon fed the experimental histidine diets: low (L), medium (M) and high (H) during three periods from June to October. Data are represented as means with their standard errors (six per experimental duplicate; see Fig. 1). a,b,c,d,e Mean values with unlike letters were significantly different (P < 0·05; nested ANOVA and Tukey's honestly significant difference post hoc test). ![]() , LLL;
, LLL; ![]() , LLM;
, LLM; ![]() , LLH;
, LLH; ![]() , LMM;
, LMM; ![]() , LHH;
, LHH; ![]() , MMM;
, MMM; ![]() , HLL;
, HLL; ![]() , HMM;
, HMM; ![]() , HHL;
, HHL; ![]() , HHM;
, HHM; ![]() , HHH.
, HHH.

Fig. 7 Correlation between individual lens N-acetyl-histidine (NAH) and cataract scores in adult Atlantic salmon fed experimental diets differing in histidine from June to September and examined after the second period with a serious outbreak of cataract (n 60). y = − 0·1439x 2 − 0·1624x+10·913; R 2 0·3556.
Concentrations of most free amino acids in the lens were significantly higher in the group fed L histidine compared with the group fed M and H histidine before the cataract outbreak (Table 6). In same manner as the sum of free amino acids, concentration of selected free amino acids like hydroxyproline, glutamate, alanine, isoleucine, leucine, tyrosine, phenylalanine and 1-methyl histidine gradually decreased in fish fed L-, M- and H-histidine diets. Adding NAH to the other lens amino acids gave, however, a total amino acid level of 24·3 μmol/g in L and H histidine groups, and somewhat lower level in lenses from the M histidine group (20·1 μmol/g). In September, after the outbreak of cataract, a similar elevated concentration of major amino acids were observed in the L histidine group (LL) compared with the groups fed histidine-supplemented diets in either of the two periods (Table 7). Again, the total amino acid, including NAH, in lens was similar among the groups within the range of 23·3–25·4 μmol/g. At the conclusion of the trial in October, free amino acids like glutamate, glycine, phenylalanine, taurine and 1-methyl histidine in lens were still elevated in salmon fed continuously on the L-histidine diet (LLL), especially compared with fish fed the H-histidine diet (HHH) (P < 0·05; data not shown).
Table 6 Free amino acids in lens tissue of adult Atlantic salmon fed low (L; 9·3 g/kg), medium (M; 12·8 g/kg) and high (M; 17·2 g/kg) levels of His sampled in July (week 6) before a serious outbreak of cataract*

ND, below the detection limit; NAH, N-acetyl-histidine.a,b,c Mean values within a row with unlike superscript letters were significantly different (P < 0·05).
* Six samples per duplicate; evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test at a level of P < 0·05.
† Single-letter codes indicate the use of diets L, M or H in the first feeding period.
‡ Pooled sem < 0·005.
Table 7 Free amino acids in lens tissue of adult Atlantic salmon fed low (L; 9·3 g/kg), medium (M; 12·8 g/kg) and high (M; 17·2 g/kg) levels of His sampled in September (week 13) after the outbreak of cataract*

NAH, N-acetyl-histidine.a,b,c Mean values within a row with unlike superscript letters were significantly different (P < 0·05).
* n 6 samples per duplicate; evaluated by nested ANOVA and Tukey's honestly significant difference post hoc test at a level of P < 0·05.
† Two-letter codes indicate the use of diet L, M or H in the first and second feeding periods.
‡ Pooled sem < 0·005.
Discussion
The present study demonstrates for the first time that adult Atlantic salmon have a practical requirement for dietary histidine levels above present recommendations by Norwegian Research Council(10) to prevent cataract development during their second year in seawater. Low prevalence and severity of cataracts were recorded at the start of the experiment. A massive outbreak of cataracts was recorded in September, probably initiated by a period of increased water temperature in July. Moderately elevated and fluctuating temperatures have been demonstrated to represent risk factors for cataract development, both experimentally and in field(Reference Bjerkås and Bjørnestad27, Reference Bjerkås, Bjørnestad and Breck28), probably related to increased growth and metabolism. The cataract severity among the dietary groups was systematically related to the dietary histidine level and histidine feeding period. All the groups of salmon fed the unsupplemented diet with 9 g histidine/kg during the first period developed severe cataract, while histidine-supplemented diets ≥ 13 g histidine/kg, as in the M- and H-histidine diets prevented cataract. However, fish crossed over from low to both histidine-supplemented diets (LM and LH groups) developed intermediate severities of cataract, suggesting that feeding a histidine-supplemented diet from onset July was too late for a maximum cataract preventive effect. On the other hand, fish crossed over from the H-histidine diet in the first period to a L-histidine diet in the actual period with cataract outbreak (HL group) were fully protected. In summary, both dietary histidine supplementation and histidine feeding period were critical for cataract prevention. Since the present histidine supplementation levels ( ≥ 13 g histidine/kg) prevented cataract in adult Atlantic salmon after a relatively short period of feeding, this represents an effective counter measure in periods with increased risk for cataract, like rises in water temperature during the summer season.
In a previous study with Atlantic salmon smolts, dietary histidine levels of 12 and 18 g/kg were compared, and showed that the former was not able to prevent cataract during a 26-week feeding period(Reference Breck, Bjerkås and Campbell7). This was a first indication that histidine requirement to support growth and health in Atlantic salmon parr and smolts under practical conditions may be higher than the given minimum requirement of 7 g/kg(10). Salmonids undergo smoltification to prepare for a migration from fresh water to seawater, and muscle and eye histidine metabolism seems to be a part of these adaptations(Reference Breck, Bjerkås and Campbell7, Reference Munakata, Aida and Amano20). Feeding studies with Atlantic salmon throughout the parr–smolt transformation indicated that the histidine requirements may vary according to the physiological status of the fish. After transferring to seawater and when provided with sufficient dietary His support, Atlantic salmon seems to build up tissue-specific histidine compounds, anserine and NAH in muscle and lens, respectively, thereby ‘trapping’ histidine intracellularly and making it unavailable for protein synthesis(Reference Breck, Bjerkås and Campbell7). These specific histidine compounds are believed to have important physiological and biochemical stabilising functions, related to cataract development. The underlying historical studies for establishing the histidine requirement in salmonids(10) did not consider these tissue imidazoles as markers for histidine status.
With feed target composition to fulfil the requirements for optimal growth and protein utilisation, amino acid deficiencies have not been considered to be an area of concern with respect to fish health(Reference Hardy, Lim and Webster22). Somatic growth seemed not to vary according to the present dietary histidine regimes. All the diets had levels well above the minimum requirement for histidine, and all the dietary groups showed reasonable growth during the experiment (specific growth rate ≥ 0 40 %/d). Low temperature fishmeal and soyabean protein concentrates represent high protein qualities with highly digestible histidine(10). Likewise, the free supplemented histidine salt is highly digestible and shows superior absorption(Reference Breck, Bjerkås and Campbell6, Reference Breck, Bjerkås and Campbell7, Reference Breck, Bjerkås and Sanderson12). However, the present study shows that the use of a fishmeal-based diet without histidine supplementation did not supply sufficient histidine to fulfil essential functions of histidine beyond protein synthesis.
A significant part of the fish population (28 %) in all net pens had initiated sexual maturation during the trial. Excluding these individuals at the final sampling generally improved growth rates, but did not affect the overall outcome of growth among the experimental groups. Breck et al. (Reference Breck, Bjerkås and Campbell7) suggested a growth-promoting effect of histidine in salmon smolts, which was argued to be related to direct effects on amino acid metabolism or by improved physiological conditions of the tissue. A similar growth promotion could not be confirmed in adult salmon in the present study. The growth and macronutrient deposition differ, however, between smolts and growers, which may mask the potential modest effects of histidine on protein synthesis(Reference Breck, Bjerkås and Sanderson12).
White muscle anserine, representing the major imidazole in salmon body, was affected significantly by dietary histidine regimes but within a narrow concentration range. According to the present high muscle concentrations and minor changes between groups and samplings, anserine synthesis seems to be prioritised in all the groups, even in groups with critical histidine supply and status during the outbreak of cataract. Between September and October, muscle anserine generally declined moderately, with a concomitant rise in precursors to anserine biosynthesis (carnosine and β-alanine) in most groups, except those fed on a L-histidine diet. This indicates a compensatory up-regulation of anserine synthesis in the final period, however, again depending on available free histidine as substrate. While elevated muscle anserine during parr–smolt transformation reflects a need for buffering capacity relative to increased anaerobic burst swimming activity in seawater, anserine has also been suggested to be a more important antioxidant in rainbow trout growers than juveniles(Reference Kim, Park and Choi15). Maximal muscle tissue concentration of anserine was achieved by use of the M-histidine diet with 12·8 g histidine/kg. Muscle carnosine, representing a low concentration imidazole intermediate in anserine biosynthesis was generally higher in concentration before than after the outbreak of cataract. In July, there seemed to be a saturation in anserine synthesis in both fish fed M and H histidine-supplemented diets, as evaluated by significantly lower β-alanine and carnosine and higher anserine concentrations in these two groups compared with fish fed the L-histidine diet. At later samplings, both anserine precursors were lower in all the groups.
Low concentration levels of NAH in the lens have been considered a direct risk factor for cataract development(Reference Breck, Bjerkås and Campbell7). Dietary levels of 17·6 g histidine/kg mitigated cataract formation in salmon smolts in their first season in seawater compared with a control diet with 11·7 g histidine/kg, representing lens NAH concentrations of>10 and < 2 μmol/g, respectively. In the present study, the lens NAH reflected the L-, M- and H-histidine diets by showing mean values of 1·1, 3·1 and 10·8 μmol/g before the outbreak of cataract. During the outbreak of cataract, the concentrations increased to 5·3, 10·9 and 11·8 μmol/g, respectively, illustrating an active build-up of lens NAH in all the groups; however, the concentrations were not increased sufficiently in the L histidine groups. This confirms the applicability of NAH as an indicator for cataract susceptibility, in adult salmon. The intermediate histidine feed level (M), increasing lens NAH from 3·1 to 10·9 μmol/g from July to September was sufficient to prevent cataract during the outbreak. Similarly, fish crossed over from H to L histidine diet (HL), with lens NAH between 9 and 11 μmol/g in September showed equal cataract prevention. In contrast, fish crossed over from a low to a histidine-supplemented diet (LM or LH) showed more severe cataract development; despite lens NAH levels>8 μmol/g at the time of sampling. Clearly, the actual status of NAH in these groups during the first and second feeding periods may have changed between the samplings, and for the latter groups, this represented a critical low NAH concentration which consequently led to cataract development. Since the status of lens NAH was not recorded exactly at the outbreak of cataract, it is not possible to estimate a critical value of this indicator to prevent cataract. Mean cataract scores did not increase much from September to October, suggesting that the major outbreak was over in September. For individual fish sampled in September, a negative correlation was apparent between lens NAH concentration and cataract scores (Fig. 7), as was observed previously in salmon smolts after transferring to seawater(Reference Breck, Bjerkås and Campbell7). Generally, a low level of NAH in the lens seems to represent a lower defence towards fluctuations in both external and biotic factors, which eventually Pb to cataract changes. The relatively wide concentration ranges at each score level indicates that the individual fish may be at different phases of cataract development and repair at the time of sampling.
The mechanisms behind the cataract alleviating effect of histidine are not fully understood. Histidine and its related imidazoles have been suggested to cover many biochemical roles in the salmon tissues besides being an essential amino acid in protein synthesis, such as lens osmoregulation(Reference Breck, Bjerkås and Campbell7). The former has been given support by recent studies on salmon lens ex vivo cultures(Reference Tröße, Rhodes and Sanderson29), where exposure of lenses to a hypoosmotic medium caused an efflux of NAH from the lens and into the medium. This finding was interpreted as a survival mechanism by the lens; to adjust osmolality according to the surrounding medium to prevent swelling and eventually rupture of the lens capsule. In the present study, free amino acid profiles in muscle and lens were recorded both before and after the cataract outbreak to determine if any other amino acid was influenced by the histidine feeding regimes. These data indicate first that the muscle and lens did not suffer from deficiency of any other essential amino acids (methionine, cystine, tryptophan) that have been related to cataract development(Reference Bjerkås, Breck and Waagbø1). Secondly, the concentrations of many free amino acids were significantly elevated in the L histidine groups compared with the histidine-supplemented groups, most probably to equalise osmolality or the total concentration of amino acids in deficiency of histidine for NAH synthesis. This latter argument is also supporting the role of NAH in lens osmoregulation. Obviously, NAH synthesis represents trapping of high concentrations of histidine for use as an effective osmolyte, compared with a less efficient mixed battery of other amino acids, among others taurine, glutamic acid, glutamine, glycine, valine, methionine, leucine, tyrosine and phenylalanine; all with concentrations>1 μmol/g in the L histidine group before the cataract outbreak and somewhat less after the outbreak. This is summarised in the Tables 6 and 7 as a fairly equal sum of total amino acids (including NAH) in the lens and supports our earlier finding in salmon smolts(Reference Breck, Bjerkås and Campbell7).
In summary, the present study demonstrates for the first time a histidine-related cataract in adult Atlantic salmon in their second year in seawater. Low lens NAH concentrations in the first two feeding periods coincided with increased severity of cataract formation. Feeding a diet with 12·8 g histidine/kg throughout the experimental periods prevented cataract development. Alternatively, it also seemed possible to prevent cataract by pre-feeding a H-histidine diet in risk periods. Even lately initiated cataract counter measures seem to reduce cataract outbreaks, where a diet supporting 17·2 g histidine/kg added more protection compared with a diet with 12·8 g histidine/kg. A risk factor in the present study was probably a rapid rise in the sea temperature during summer. The outcome of the present histidine feeding study may be used as a nutritional guideline for cataract prevention in adult salmonids in periods of increased risks.
In a separate communication, we aimed to explore potential molecular mechanisms for the present histidine-related cataract, both with respect to acute cataractogenesis from July to September and a suggested repair phase from September to October. Cataract v. non-cataract lenses from selected groups were screened for differentially expressed genes using a microarray experiment(Reference Breck, Bjerkås and Campbell25). The expression levels of a subset of genes coding for proteins involved in antioxidation, osmoregulation, carbohydrate metabolism and apoptosis were identified, all representing the mechanisms involved in cataractogenesis and potentially influenced by histidine status.
Acknowledgements
The authors wish to thank research engineer Johnny Elvrum at Skretting ARC, Lerang Research station, and engineer Anita Birkenes at National Institute of Nutrition and Seafood Research, for their valuable technical assistance at the rearing site and the laboratory, respectively. This research was supported by the Norwegian Research Council (#168435/S40), Marine Harvest Norway, Skretting ARC, Norway, Skretting Ireland and National Institute of Nutrition and Seafood Research. There are no conflicts of interest that the authors should disclose. Each co-author has assisted the first author in preparing the manuscript, although they participated differently in designing the research (O. B., R. W., W. K. and R. F.), making the feeds (W. K. and R. F.), conducting the experiment and sampling (C. T., O. B., R. W., W. K. and R. F.), and performing chemical (C. T.) and statistical analyses (C. T., O. B. and R. W.).














