


Introduction
Zygodont proboscideans (Tobien, Reference Tobien1975) have been referred to the family Mammutidae Hay, Reference Hay1922. Shoshani and Tassy (Reference Shoshani and Tassy2005) have reported this family containing two subfamilies, namely Eozygodontinae Mckenna and Bell, Reference McKenna and Bell1997 (including only Eozygodon Tassy and Pickford, Reference Tassy and Pickford1983, from the lower Miocene bed of Africa) and Mammutinae Hay, Reference Hay1922 (including Zygolophodon Vacek, Reference Vacek1877 from the middle-upper Miocene and its descendant Mammut Blumenbach, Reference Blumenbach1799 from the Plio-Pleistocene). However, the fossils of palaeomastodont–like proboscideans that first expressed mammutid characters (Sanders et al., Reference Sanders, Kappelman and Rasmussen2004) were discovered from the late Oligocene at Chilga, Ethiopia as well as Rasmussen and Gutierrez (Reference Rasmussen and Gutiérrez2009) also reported a new taxon of Mammutidae from the upper Oligocene bed at the Losodok (Lothidok), Kenya and named it Losodokodon losodokius. In this case, the Losodok mammutid is more primitive and older than Eozygodon so that the subdivision of Eozygodontinae and Mammutinae need to be revised or expanded. In addition, the late Oligocene palaeomastodont-like fossils from Chilga, which suggest the first expression of mammutid features, and possibly that Palaeomastodontidae is not monophyletic but instead is composed of ancestral gomphotheriids (Phiomia) and ancestral mammutids (Palaeomastodon) (Sanders et al., Reference Sanders, Kappelman and Rasmussen2004). Hence, the first mammutids occur in the late Oligocene of Africa and then they immigrated into Europe by the end of the early Miocene, and by the middle Miocene had spread throughout Eurasia and into the New World (Tassy, Reference Tassy1986; Mazo, Reference Mazo1996; Sanders, Reference Sanders1996; Tobien, Reference Tobien1996; Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa and Delmer2010).
Zygodont proboscidean fossils have flourished at middle to rather high northern latitudes, in the early Miocene to Pleistocene bed of North America, Europe, northern Africa, and Asia (China and Pakistan). However, there are some reported in low latitude regions such as in the late Oligocene to middle Miocene bed of Kenya, Uganda and Namibia (Pickford, Reference Pickford2007) that are important for the origin of mammutids. In particular, zygodonts have never been reported from Southeast Asia, although zygodont material referred to Zygolophodon is known from Lufeng and Kaiyuan in Yunnan Province, southern China (Chow et al., Reference Chow, Zhang and You1978; Zhang, Reference Zhang1982).
Although zygodonts have not been reported from Thailand, 10 proboscidean genera ranging in age from the middle Miocene to Holocene have been discovered in that country over the past several years. They include Archaeobelodon sp., Prodeinotherium pentapotamiae, Gomphotherium sp., cf. Protanancus macinnesi, Tetralophodon cf. xiaolongtanensis, Tetralophodon sp., Stegolophodon nasaiensis, Sl. cf. latidens, Sl. praelatidens, Sl. cf. stegodontoides, Stegodon elephantoides, S. insignis, S. cf. insignis, S. cf. orientalis, Anancus sp., Sinomastodon aff. yangziensis, and Elephas (von Koenigwald, Reference Koenigswald1959; Buffetaut et al., Reference Buffetaut, Helmcke–Ingavat, Jaeger, Jongkanjanasoontorn and Suteethorn1988; Tassy, Reference Tassy1990; Tassy et al., Reference Tassy, Anupandhanant, Ginsburg, Mein, Ratanasthien and Suteethorn1992; Saegusa et al., Reference Saegusa, Ratanasthien and Nakaya1999; Nakaya et al., Reference Nakaya, Saegusa, Ratanasthien, Kunimatsu, Nagaoka and Suganuma2002, Reference Nakaya, Saegusa, Ratanasthien, Kunimatsu, Nagaoka, Chintaskul, Suganuma and Fukuchi2003; Chaimanee et al., Reference Chaimanee, Suteethorn, Jintasakul, Vidthayanon, Maradat and Jaeger2004; Kunimatsu et al., Reference Kunimatsu, Ratanasthien, Nakaya, Saegusa and Nagaoka2004; Pickford et al., Reference Pickford, Nakaya, Kunimatsu, Saegusa, Fukuchi and Ratanasthien2004; Suganuma et al., Reference Suganuma, Hamada, Tanaka, Okada, Nakaya, Kunimatsu, Saegusa, Nagaoka and Ratanasthien2006; Thasod, Reference Thasod2007; Duangkrayom et al., Reference Duangkrayom, Thasod and Jintasakul2010; Thasod et al., 2012).
In 2011, new materials representing a zygodont proboscidean were discovered by workers at the sand pit no.10 of Tha Chang sand pits (belonging to Takut Khon Village), Nakhon Ratchasima Province, northeastern part (Fig. 1). The Tha Chang sand pits were given the numbers 1 through 10 by the Thailand–Japan team, with the Takut Khon Village sand pit being number 10. A lower molar was collected by the Northeastern Research Institute of Petrified Wood and Mineral Resources, Nakhon Ratchasima Rajabhat University, and an upper molar was collected by the Sukhothai Airport Natural History Museum. Although the two teeth did not come from the same individual skeleton, their morphology suggests that they belong to the same zygodont proboscidean species. The specimens described in this paper are the first record of any zygodont proboscidean for Thailand, and indeed for Southeast Asia. They have important implications for understanding zygodont evolution and dispersal patterns.
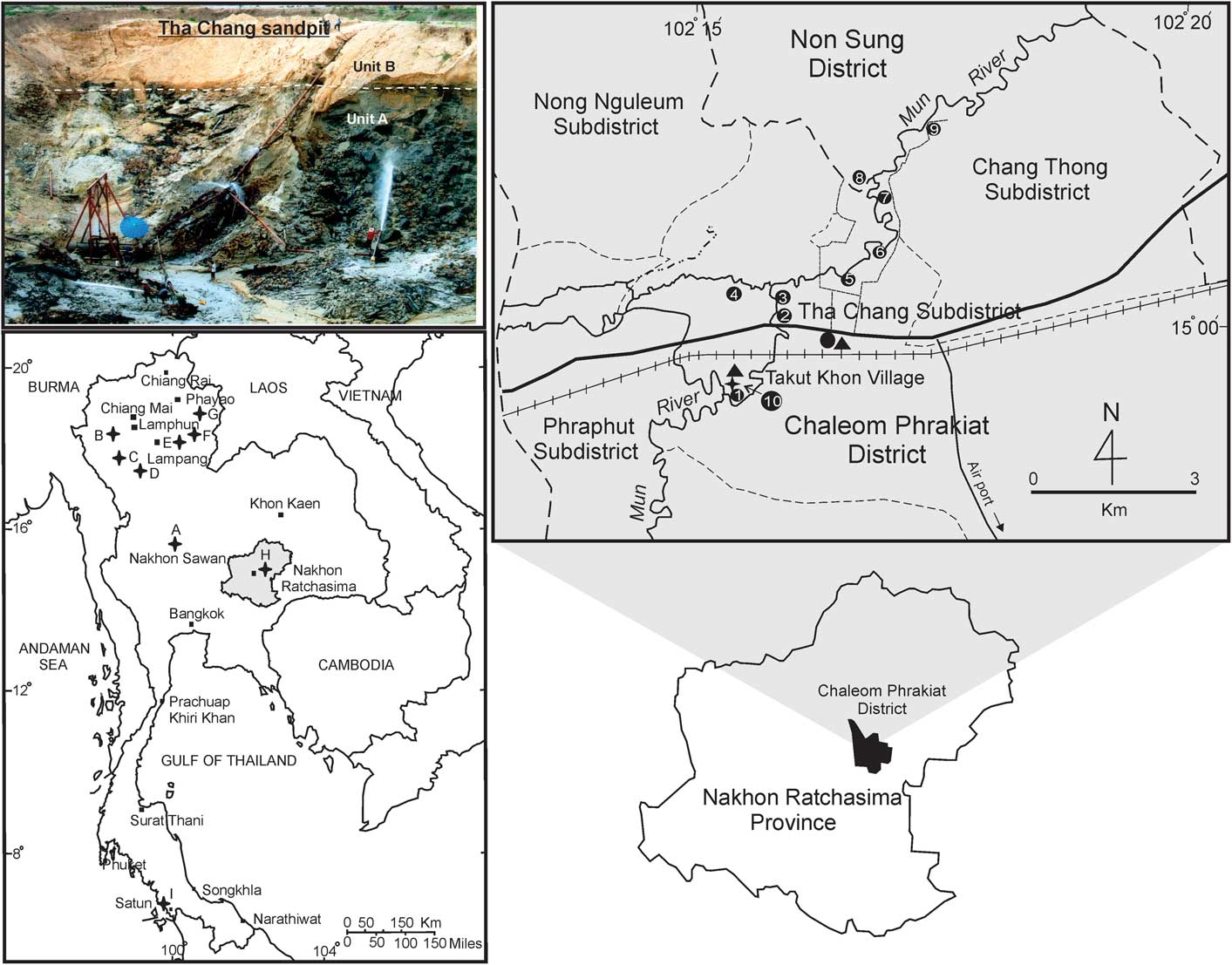
Figure 1 Map of Thailand showing fossil proboscidean localities. Lower left: A, Dechatiwongse Bridge, Nakonsawan Province; B, Mae Soi locality in Chom Thong District, Chiang Mai Province; C, Ban Na Sai coal mine, Lumphun Province; D, Mae Teep coal mine, Lumphun Province; E, Mae Moh coal mine, Lumpang Province; F, Sop Mae Tham locality, Lumpang Province; G, Chiang Muan coal mine, Proyao Province; H, Tha Chang sand pit, Nakhon Ratchasima Province (shaded); I, Wang Kuai cave, Thungwa District, Satun Province. Top left: Photo of the Tha Chang sand pit is marked to indicate the extent of the lower reduced unit (Unit A) and upper oxidized unit (Unit B). Top right: Map of Chaleom Phrakiat District shows the Tha Chang sand pits. The Takut Khon Village sand pit is in number 10.
Materials and methods
Materials for study include the right lower third molar (m3), NRRU-TKK-001 and the left upper third molar (M3), PRY–200 from Tha Chang sand pits (Figs. 2–4). The materials for comparison include AMNH19414, AMNH8137, AMNH26461, AMNH26476, DPC9009, DPC12598, DPC5920, F:AM23339, GIU, IVPP-V2485, IVPP-V5584, IVPP-V5585, IVPP-V4688.1, IVPP-RV77001, IVPP-unnumbered, KNM-LS18244, KNM-ME17, KNM-ME7545, KNM-ME7547, no. 2280/2, PIN2202-4, PIN2202-5, PIN2202-6, RAM908, THP-00079, THP-10022, THP-10031, THP-18908, THP-18903, YV031, and YV0132 (Table 1).
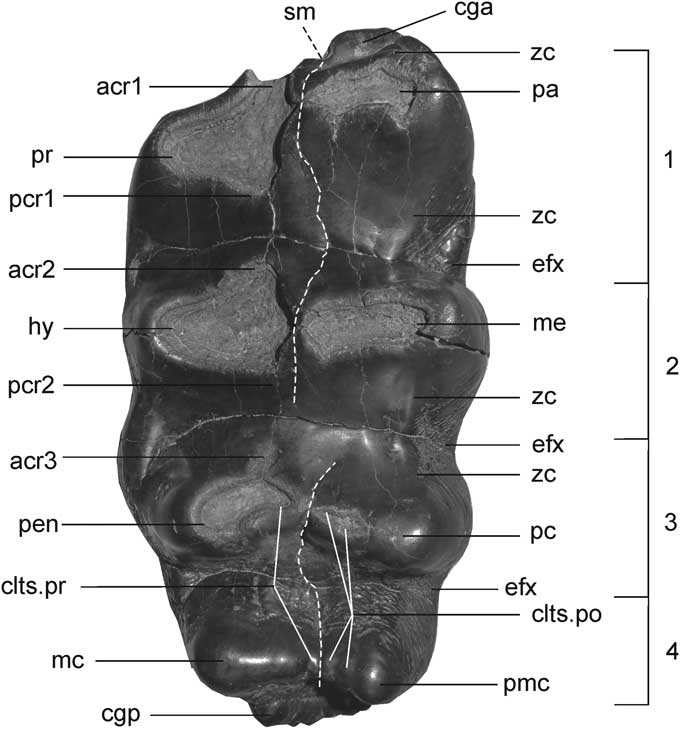
Figure 2 Dental nomenclature: (acr1, 2, 3), anterior pretrite crescentoids of the first, second and third loph(id)s; (cga), anterior cingulum; (cgp), posterior cingulum (talon(id)); (clts.pr), pretrite conelets; (clts.po), posttrite conelets; (efx), ectoflexus; (hy), hypocone (pretrite main cone of the second loph(id)); (mc), main cone (pretrite main cone of the fourth loph(id)); (me), metacone (posttrite main cone of the second loph(id)); (pa), paracone (posttrite main cone of the first loph(id)): (pc), third posttrite cusp (posttrite main cone of the third loph(id)); (pcr1, 2), posterior pretrite crescentoids of the first and second loph(id)s; (pen), postentoconule (pretrite main cone of the third loph(id)): (pmc), posttrite main cone of the fourth loph(id); (pr), protocone (pretrite main cone of the first loph(id)); (sm), median sulcus; (zc), zygodont crest.
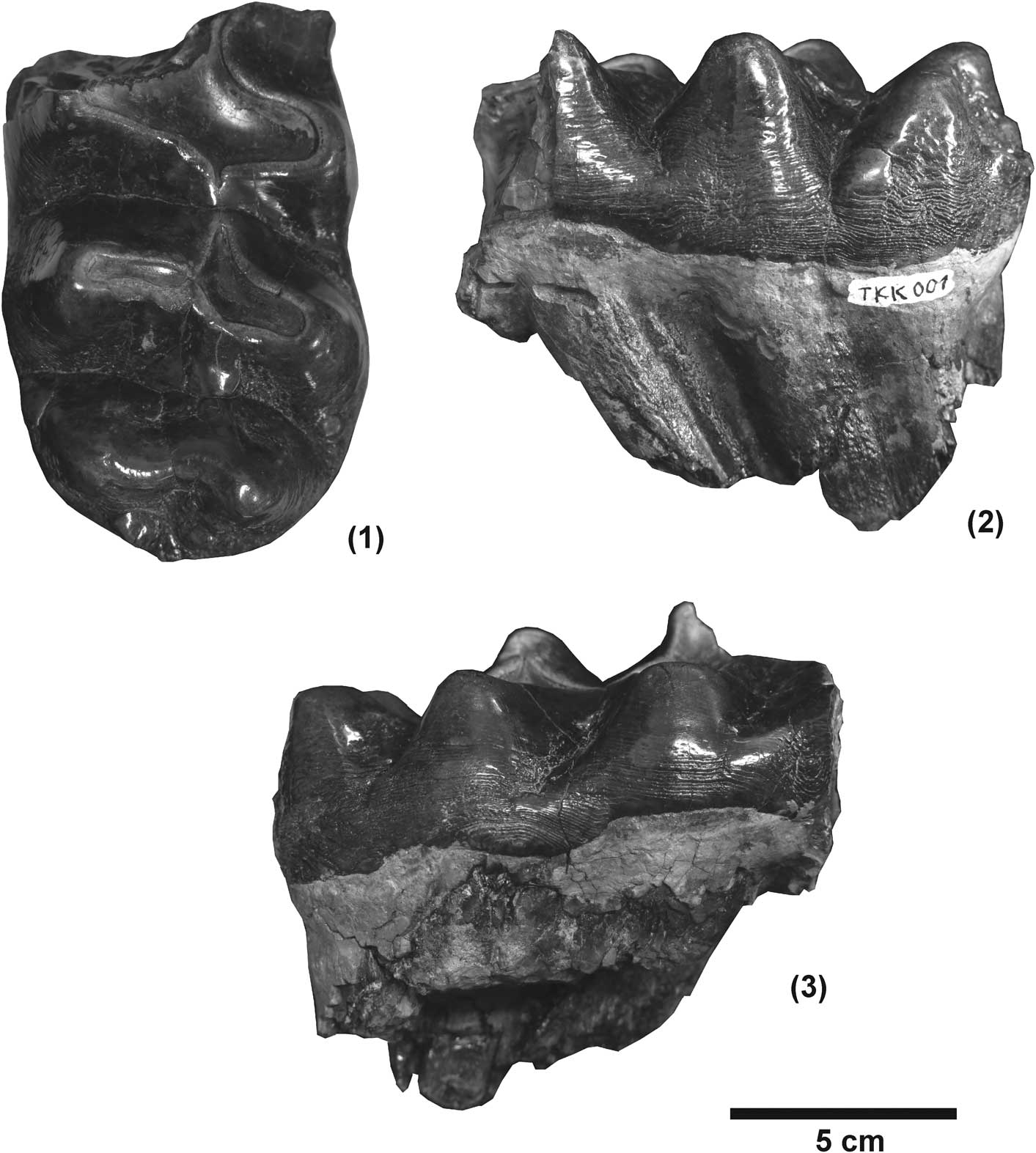
Figure 3 The lower third molar of Zygolophodon sp. from Tha Chang sand pit no. 10, NRRU-TKK001: (1) occlusal view, (2) lingual view, (3) buccal view.
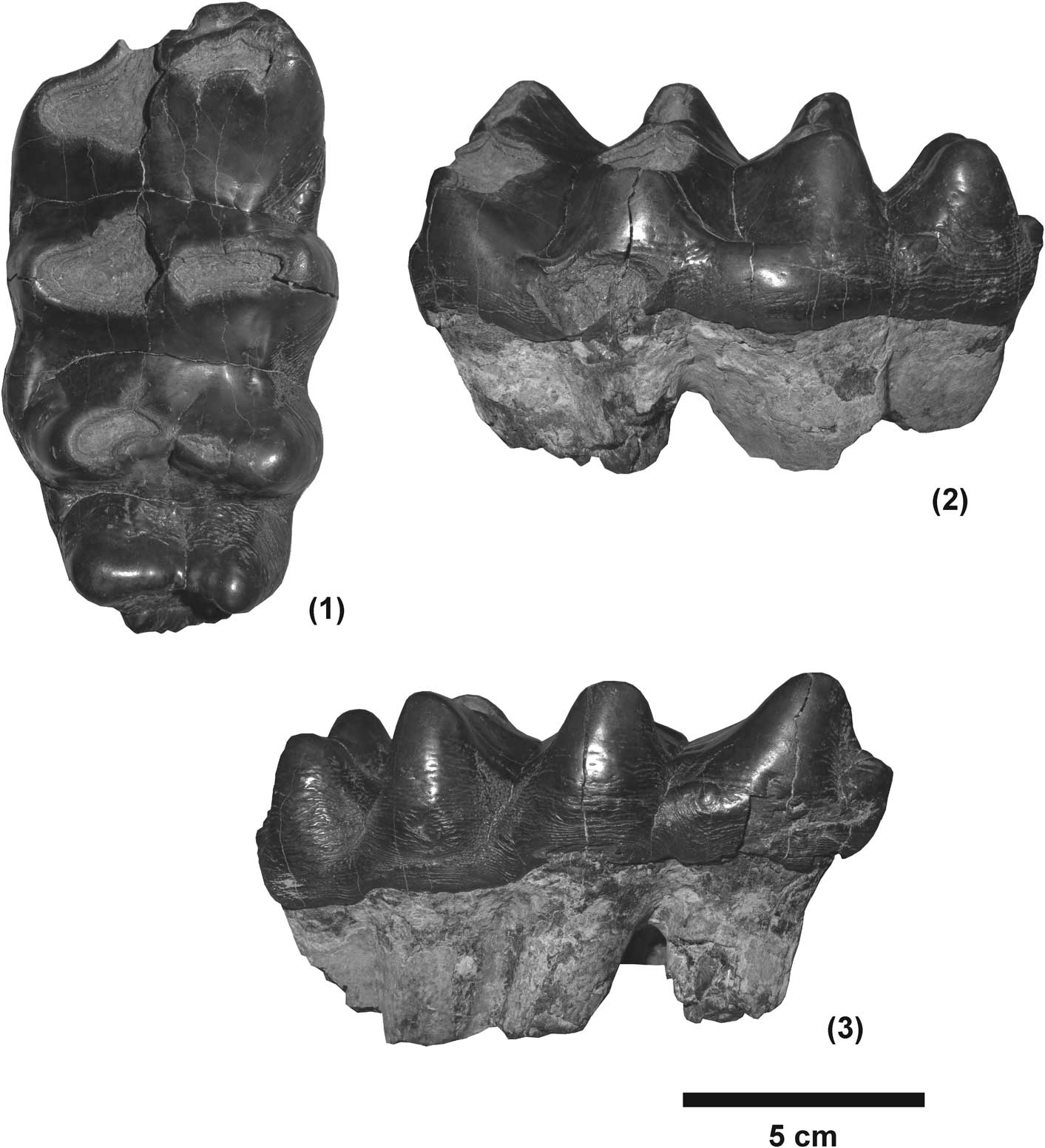
Figure 4 The upper third molar of Zygolophodon sp. from Tha Chang sand pit no. 10, PRY200: (1) occlusal view, (2) lingual view, (3) buccal view.
Table 1 Third molars measurement of the zygodont proboscideans (in mm), Rm, Lm = right and left lower molars, RM, LM = right and left upper molars, R = amount of loph(id) (ridge), L = total of molar leangh, W = width of the crown, H = high of the crown, W1 = width of the 1st lophid, Wx = width of the talonid, HI = hypsodonty index, in which the height of individual lophid is expressed as a percentage of the width (HFootnote *100/W), WI = width–length index, ratio between the maximum length and the maximum width (width/(length/100)), ET = enamel thickness.
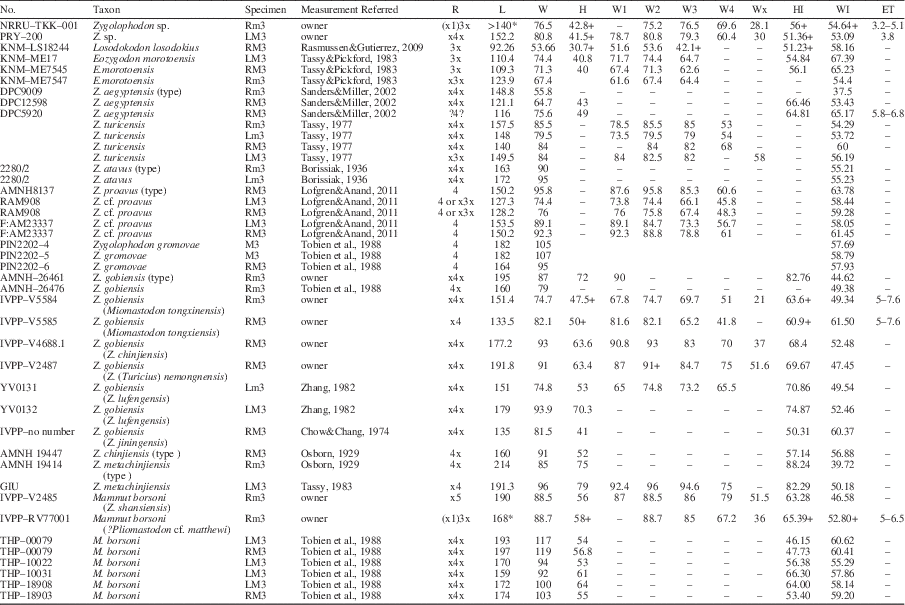
* Estimate measured.
Repositories and institutional abbreviations
AMNH, American Museum of Natural History, New York; DPC, Duke Primate Center, North Carolina; F:AM, Frick Collection, American Museum of Natural History, New York; IVPP-V, Institute of Vertebrate Paleontology and Paleoanthropology, vertebrate collection, Beijing; NRRU-TKK, Northeastern Research Institute of Petrified Wood and Mineral Resources, Nakhon Ratchasima Rajabhat University, Takut Khon Village collection, Nakhon Ratchasima; PIN, Paleontological Institute, Moscow; PRY, Sukhothai Airport Natural History Museum, Sukhothai, Mr. Piriya Vachajitpan collection; RAM, Raymond M. Alf Museum of Paleontology, Claremont, California; THP, Natural History Museum, Tianjin (Museum of Huangho and Paiho).
Systematic paleontology
Dental nomenclature follows Tassy (1996). Description of occlusal features of zygodont cheek tooth M3 follows Tobien (Reference Tobien1975) (Fig. 2) and measurements are in millimeters.
Order Proboscidea Illiger, 1811
Family Mammutidae Hay, Reference Hay1922
Genus Zygolophodon Vacek, Reference Vacek1877
Type species
Mastodon turicensis Schinz, Reference Schinz1824 (for more details including the non-validity of “Mastodon tapiroides Desmarets, Reference Desmarest1822”, sometimes used by earlier authors, see Tassy, Reference Tassy1985).
Diagnosis
A mammutid with intermediate and third molars in which the pretrite and posttrite elements are of bunodont character and arranged in clearly yoke-like transverse crests. D4 and all molars are zygodont, having a transverse or slightly oblique loph. In lateral view, the main cusps are wide at the base but taper to an apical point. Transverse valleys have few and small accessory cusps. Worn loph(id)s become sharpened at their summits and show a more zygodont character; zygodont crests show a sharp angle between the lingual and posterior surfaces of the principal lingual cuspids in the lower molars, and between the corresponding surfaces of the buccal cusps in the upper molars; the accessory conules are not developed but rather have crescentoids descending into the transverse valley between loph(id)s on the pretrite side at both upper and lower molars; lophs of upper molars are normal to the long axis of the crown; lophids of lower molars are more or less oblique (i.e., postero-external to antero-internal); upper third molar bears three lophs plus a strong talon or four lophs plus a weak cingulum; and lower third molar bears four lophids plus a more or less strong cingulid (Tobien in Shoshani and Tassy, Reference Shoshani and Tassy1996; Mazo and van de Made, Reference Mazo and van der Made2012).
Zygolophodon sp.
Occurrence
Both specimens were found in sand pit no.10 of Tha Chang sand pits (14°59'50.27"N, 102°14'47.50"E) belonging to Takut Khon Village, Tha Chang Subdistrict, Chaloem Prakiat District, Nakhon Ratchasima Province (Fig. 1). Howard et al. (Reference Howard, Haines, Burrett, Ali and Bonopas2003) and Haines et al. (Reference Haines, Howard, Ali, Burrett and Bunopas2004) subdivided the sediments in the sand pits along the Mun River into a lower reduced unit (Unit A) and an upper oxidized unit (Unit B) (Fig. 1). The lower reduced unit is composed of reduced gray sediment in the form of cross–bedded, stratified to massive sand and gravel, with mud lenses. Abundant Neogene faunal and floral fossils were discovered in this unit (Suteethorn et al., Reference Suteethorn, Chaimanee and Khansupa1997; Sato, Reference Sato2002; Nakaya et al., Reference Nakaya, Saegusa, Ratanasthien, Kunimatsu, Nagaoka and Suganuma2002; Howard et al., Reference Howard, Haines, Burrett, Ali and Bonopas2003). The materials described in this study were found in the lower reducing unit. However, they were discovered by sand pit workers, and their detailed lithological context is unclear because the water jets used during the sand excavation process caused extensive damage to the strata and fossils collapsed.
Description
m3, NRRU-TKK001 (Fig. 3). This right lower third molar is missing the first lophid. There are (×1)3× lophids plus a weak talonid (posterior cingulid). A crescentoid is present and the two half lophids are arranged as yoke-like transverse crests. The second lophid is oblique to the long axis of the crown, and the third lophid is slightly oblique. The lophids become more anteroposteriorly constricted at their apices. The first lophid is broken out. The second lophid is severely worn. The third lophid is also worn, bearing a narrow, transversely elongated wear facet. There are anterior and posterior crescentoids (acr and pcr) on the pretrite side that show narrow and elongated wear surfaces. The third posttrite half lophid has a main cone and smaller adaxial conelets (clts.po), but conelets are not clearly evident on the pretrite half lophid. The unworn fourth lophid has a main cone and smaller adaxial conelet on the pretrite half lophid while the posttrite has a large and transversely elongate main cone, however the conelet is furrow–like. Zygodont crests (zc) are present, but not so strong. The cingulids are distinctly reduced. A median sulcus divides the lophids. The talonid (posterior cingulid) is shelf–like and thicker on the pretrite side than on the posttrite side. No cementum is present (Fig. 3).
M3, PRY200. This upper left molar has four lophs, in addition to anterior and posterior cingula. The lingual cingulum is quite strong, and the buccal cingulum is only evident in the vicinity of the first valley. The crowns of the first and second lophs are deeply worn, and the third loph is also somewhat worn. The crescentoids are present in the pretrite side and arranged in straight lines that run perpendicular to the anteroposterior axis of the crown, and appear to be yoke–like in this molar. The first and second lophs bear clear zygodont crests. The pretrite and posttrite half lophs are separated by a strong median sulcus. The molar is wider anteriorly than posteriorly. In lateral view, the posterior pretrite crescentoid of the first loph (pcr1) is well-developed and extends downward to the base of the adjacent valley, where it joins the anterior pretrite crecentiod of the second loph (acr2). A similar feature is also present in the valley between the second and the third lophs. However, the valley between the third and fourth lophs is open. No cementum remains in the valleys.
The pretrite of the first loph is displaced slightly posteriorly relative to the posttrite part. The zygodont crest of the first loph is stronger than those of the other lophs, and continuous with the anterior cingulum. Anterior and posterior pretrite crescentoids (acr1 and pcr1) are present. However, their surfaces are deeply worn. The acr1 is connected to the anterior cingulum.
The second loph is aligned with the transverse axis of the crown, and is deeply worn. An anterior pretrite crescentoid (acr2) is present, and extends downward to the base of the adjacent valley, whereas the posterior pretrite crecentoid (pcr2) is not prominent. The pretrite of the third loph is slightly posteriorly, while the posttrite is straight. The wear surface of the third loph is moderately worn. The posttrite half loph has a rather large main cone and small adaxial conelets (clts.po), which appear furrow–like. However, the morphology of the conelets is not clear in the pretrite half loph because of surface wear. The anterior pretrite crescentoid (acr3) is slightly worn and narrow in shape. The posterior pretrite crescentoid (pcr3) is very weak in this loph. The unworn fourth loph shows two small adaxial conelets (clts.po) on the posttrite side, which appear furrow-like. The main cone is quite large. The pretrite half loph has a large and transversely elongate main cone, and a small conelet near the median sulcus. The anterior crescentoid (acr4) is clearly and extends down to the base of the nearby valley. The talon (posterior cingulum) is shelf-like, and better developed on the pretrite side than on the posttrite side (Fig. 4).
Materials
The right lower third molar (m3) is stored at the Northeastern Research Institute of Petrified Wood and Mineral Resources, Nakhon Ratchasima Rajabhat University, as NRRU-TKK-001. The left upper third molar (M3) is stored at the Sukhothai Airport Natural History Museum, as PRY–200 (Figs. 2–4).
Remarks
The lophids of the specimen NRRU-TKK-001 (right m3) are more oblique than the loph of PRY-200 (left M3). This is a generally different pattern between the lower and upper molar, especially in the zygodont proboscideans. The enamel wear surfaces are narrow, transversely elongated and yoke–like (Tobien, Reference Tobien1973b). Both of specimens from Tha Chang have clear zygodont crests. The conelets on the posttrite side (clts.po.) are better developed than those on the pretrite side (clts.pr.). The conelets and half loph(id)s are somewhat less regularly arranged than in the bunodont molars of proboscideans such as Gomphotherium. In addition, the mesoconelets of Gomphotherium are blunter and more morphologically distinct from the zygodont proboscideans (Tobien, Reference Tobien1973b). The pretrite central conules of bunodonts are arranged in a conule–like pattern, rather than a yoke-like pattern as in the specimens from Tha Chang. These characters above indicate that the Tha Chang specimens are the zygodont proboscideans.
Compared with the oldest zygodont proboscidean, Losodokodon losodokius, from the late Oligocene of Losodok (Lothidok), Kenya (Rasmussen and Gutierrez, Reference Rasmussen and Gutiérrez2009), and the early to middle Miocene species, Eozygodon morotoensis, from Moroto, Uganda (Pickford and Tassy, Reference Pickford and Tassy1980; Tassy and Pickford, Reference Tassy and Pickford1983; Pickford, Reference Pickford2007) that they have three loph(id)s in the third upper and lower molars. The fourth loph(id)s in m3/M3 are absent or poor developed. On the contrary, the Tha Chang specimens have four loph(id)s with the posterior cingula and larger in molar size (Tables 1–3; Fig. 5).
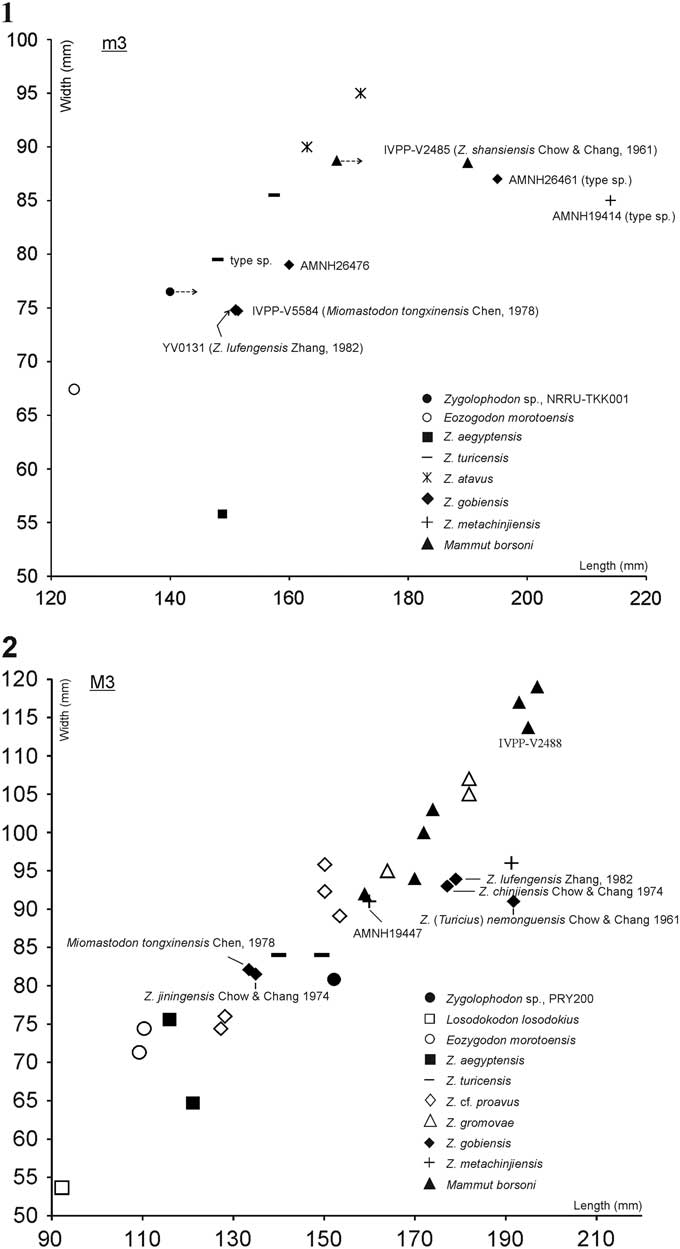
Figure 5 Scatterplot of third molar proportions for various zygodont proboscideans, including Zygolophodon sp. from Tha Chang sand pit no. 10: (1) lower molar, (2) upper molar.
Table 2 The lower third molars (m3) morphology of the Old Word zygodont proboscideans.

Table 3 The upper third molars (M3) morphology of the Old Word zygodont proboscideans.

The zygodont crests of the Tha Chang specimens are not strongly developed, in contrast to the condition in Mammut. In addition, the lateral cingula of the Tha Chang specimens are less well developed than in Mammut, whereas the crescentoids of the Tha Chang specimen are more distinct than in Mammut. According to the differentiation between the Tha Chang specimens and the oldest and youngest zygodont proboscideans, Losodokodon losodokius, Eozygodon morotoensis, and Mammut, the dental features such as molar size and well-developed anterior and posterior ridges of Tha Chang specimens indicate that it belongs to Zygolophodon.
Comparison between the Tha Chang specimens and the Egyptian early Miocene zygolophodont Zygolophodon aegyptensis (see Sander and Miller, Reference Sanders and Miller2002, p. 398, fig. 9) reveals that the third and fourth lophids of m3 are strongly convex anteriorly in the Egyptian species but quite straight in the Tha Chang specimen. In lateral view, the lophids of Z. aegyptensis are more widely separated than those of the Tha Chang specimens. However, the anterior and posterior pretrite crescentoids of the m3 of Z. aegyptensis are clearly in contact with each other according to wear, which is also the case in the Tha Chang specimen. The complete M3 of Z. aegyptensis possesses three lophs or variably a fourth loph, a thin anterior cingulum composed of numerous tiny tubercles, and a moderate-sized heel; whereas the Tha Chang M3 possesses four lophs and a tiny posterior cingulum. In addition, the lower m3 from Tha Chang is wider but shorter than that of the Egyptian species, whereas the upper M3 from Tha Chang is both wider and longer than its Egyptian counterpart (Tables 1–3; Fig. 5.1, 5.2).
The M3 of Zygolophodon cf. proavus, RAM 908 (see Lofgren and Anand, Reference Lofgren and Anand2011, p. 1393, fig. 4) from the middle Miocene of the Mud Hills, Mojave Desert, California, North America resembles the Tha Chang M3 in several respects. In Z. cf. proavus central conules are not present; however, the wear facet on the protocone extends onto thickened enamel that is present on both anterior and posterior slopes. The posterior and anterior extensions of this wear facet give a false impression that the central conules are only weakly developed. These characters are reminiscent of the Tha Chang M3, but in other important respects this tooth differs from its equivalent in Z. cf. proavus. The fourth loph is much less well developed in Z. cf. proavus than in the Tha Chang M3. In Z. cf. proavus, the first three lophs reduce their transverse dimension anteroposteriorly, unlike in the Tha Chang M3, and an ectoflexus is present on all three interlophs in Z. cf. proavus but only on the first interloph of the Tha Chang M3. Moreover, the labial cingulum is better–developed in Z. cf. proavus than in the Tha Chang M3. However, the upper molars of Z. cf. proavus described by Lofgren and Anand (Reference Lofgren and Anand2011) are smaller than other specimens of the Z. proavus. The Tha Chang M3 is longer and wider than RAM908, but narrower than other specimens of Z. proavus (Tables 1, 3; Fig. 5.2).
The furrow-like posttrite conelets of the fourth loph(id)s of both Tha Chang specimens resemble the equivalent structures in the middle Miocene European zygolophodont Zygolophodon turicensis Schinz, Reference Schinz1824. The Tha Chang specimens are only slightly narrower than the equivalent molars of Z. turicensis but slightly longer in upper molar and slightly shorter in lower molar (in the lower molar case, the length is probably a bit longer than it estimated) (Table 1; Fig. 5.1, 5.2). However, the zygodont crests and cingula of the Tha Chang specimens are reduced, whereas very strong cingula are present in Z. turicensis. In addition, a well-developed lingual cingulum is also present in the upper third molar of Z. turicensis from the middle Miocene of Jebel Cherichera, Tunisia (Tassy, Reference Tassy1985, pp. 509–510, fig. 213; Pickford, Reference Pickford2007, p. 30, fig. 2A) and the upper second molars from the Grildain, Ngorora Formation, Member A, Tugen Hills, Kenya (Pickford, Reference Pickford2007, p. 31, fig. 3). Moreover, the crescentoids of Tha Chang specimens are blunter than those of Z. turicensis (see Tobien, Reference Tobien1975, p. 196, fig. 1; Tassy, Reference Tassy1977, fig. 3, p. 659; Tables 2, 3).
A closer inspection of the molars of the zygodont proboscidean “Mastodon” atavus Borissiak, Reference Borissiak1936, from the lower Miocene Djilancik beds of Kazakhstan, will be needed in order to determine whether this specimen is assignable to Z. gobiensis, Z. turicensis, or neither. However, the symphysis is longer, and the incisors more strongly developed, than in Z. gobiensis (Tassy, Reference Tassy1985, fig. 201). These differences seem consistent with the somewhat earlier age (Shanwangian/Orleanian) of the Djilancik site. The shortening of the symphysis and incisors seen in Z. gobiensis represents an evolutionary step towards Mammut, in which the symphysis and incisors are reduced (Tobien et al., Reference Tobien, Chen and Li1988). The m3 of Z. atavus is larger and much wider than the Tha Chang m3 (Tables 1, 2; Fig. 5.1).
Previously described Chinese zygolophodonts show varying degrees of similarity to the Tha Chang specimens. The type specimen of Zygolophodon gobiensis Osborn and Granger, Reference Osborn and Granger1932 (AMNH 26461) is a hemimandible with a complete m2 and m3 from the Tunggur locality, Nei Mongol. The m3 of this specimen resembles the Tha Chang m3 NRRU-TKK001 in having small conelets on both pretrite and posttrite sides, a thick talonid on the pretrite, lophids with a low degree of obliquity, weakly developed zygodont crests, a reduced lateral cingulum, and valleys that lack cementum. The m3 of the type specimen of Z. gobiensis is larger than the Tha Chang specimen; however, the specimen AMNH 26467 is nearly equal in size to Tha Chang m3. In addition, molar has a narrower crown and the main cone and conelets of the fourth lophid are blunter in Z. gobiensis (Tables 1, 2; Fig. 5.1).
Zygolophodon gromovae Dubrovo, 1970 is represented by an upper M2 and M3 (PIN. no. 2002-5, 2002-6 respectively), from the middle Miocene in age of the Tunggur locality, Nei Mongol. In these specimens, small pretrite conelets are present. Upper M3 has three conelets on the posttrite side. The fourth loph is crest-like rather than conule-like (Tobien et al., Reference Tobien, Chen and Li1988). In these characters, the M3 of Z. gromovae resembles the Tha Chang M3, PRY200. However, the Tha Chang M3 is smaller than the M3 of Z. gromovae and has a better-developed fourth loph (Tobien et al., Reference Tobien, Chen and Li1988) (Tables 1, 3; Fig. 5.2). However, the type specimen of Z. gromovae is so badly damaged that its M3 is difficult to compare to the Tha Chang M3 in other respects.
The only representative of Zygolophodon nemonguensis Chow and Chang, Reference Chow and Zhang1961, an M3 (IVPP-V2487) from Nei Mongol, has four lophs. The fourth loph is as well developed as that of the Tha Chang M3, PRY200. However, the fourth loph is equal in width to the third loph in IVPP-V2487, but clearly narrower than the third loph in PRY200. The talon of IVPP-V2487 is stronger than that of the Tha Chang M3. Nevertheless, the zygodont crest of the first and second posttrite half lophs, the lingual beaded antero-external cingula, and the crescentoids are more prominent in IVPP-V2487 than in PRY200. The pretrite main cone of the fourth loph of PRY200 is larger and more transversely elongate than in IVPP-V2487, and the posttrite half lophs are divided into more conelets. Furthermore, PRY200 is smaller than IVPP-V2487 (Tables 1, 3; Fig. 5.2).
The Zygolophodon jiningensis Chow and Chang, Reference Chow and Zhang1974 is an unnumbered M3 from Jining, Nei Mongol. This tooth is unworn, but the summits of the three most anterior lophs are so badly broken that cusps cannot be distinguished. This specimen has four lophs and a nodular talon. The posttrites are bicuspid, with small adaxial conelets. In these features, the Jining specimen resembles the Tha Chang M3, PRY200. In particular, the bicuspid nature of the posttrite and the presence of small adaxial conelets can be clearly recognized in the fourth loph of PRY200. However, the fourth loph of PRY200 is better developed than that of the Jining specimen. In addition, the pretrite of the fourth loph of the Jining specimen has two subequal larger mammillae separated by two smaller squeezed ones (see Tobien et al., Reference Tobien, Chen and Li1988, p. 151, fig. 42), while the main cone of the Tha Chang specimen is large and transversely elongate with small one adaxial conelets presented (Figs. 2, 4.1). The zygodont crests of the Jining specimen are better developed than in the Tha Chang M3, and the molar is shorter (Tables 1, 3; Fig. 5.2).
Miomastodon tongxinensis Chen, Reference Chen1978 is represented by an m3 (IVPP-V5584) and an M3 (IVPP-V5585), from Gujiazhuangzi, Tongxin, Ningxia. The anterior loph of the upper third molar is strongly worn. The third loph has a small adaxial conelet and a strong anterior conule, but no posterior conule is present. The first and third posttrite half lophs have oblique, oppositely oriented long axes, while the second one is normal to the tooth midline. An antero-external cingulum is present. The pretrite side of the fourth loph has an abaxial main cone and a smaller adaxial conelet. In the above characters, the Tongxin M3 resembles the Tha Chang M3, PRY200. However, the Tongxin M3 differs from the Tha Chang M3 in that the posttrite half lophs are bicuspid, zygodont crests are present only in the first and second lophs, and the fourth loph is the narrowest one. The lower third molar is similar to the Tha Chang m3 NRRU-TKK001 in that the third and fourth lophids bear small adaxial conelets, and the upper and lower molars from Tongxin are similar in size to their counterparts from Tha Chang (Tables 1–3; Fig. 5.1, 5.2). Nevertheless, the pretrite and posttrite of the fourth loph(id)s are bicuspid in both Tongxin molars, whereas the posttrite fourth loph is tricuspid in the Tha Chang specimens.
Zygolophodon chinjiensis was named by Osborn (Reference Osborn1929) as Serridentinus chinjiensis. The type specimen (AMNH 19447), a right upper third molar (Osborn, Reference Osborn1929, p. 4, fig. 5), bears four ridges and a posterior cingulum. The anterior lophs are each divided into many conelets. We cannot determine whether this character is present in the Tha Chang M3 because of deep wear. However, the fourth loph has a tricuspid posttrite side, and a bicuspid pretrite side. The adaxial conelets are smaller than the main cone, as in the Tha Chang M3. The zygodont crests, crescentoids and lateral cingula of Z. chinjiensis are much stronger than those of the Tha Chang M3. The type specimen of Z. chinjiensis is much wider and a bit longer than the Tha Chang M3 (Table 1; Fig. 5.2).
Z. chinjiensis from China was described by Chow and Chang (1978), based on two specimens of M3 (IVPP-V4688.1 and IVPP-V4688.2) from the upper Miocene of Xiaolongtan coal mine, Kaiyuan, Yunnan. These Chinese specimens each bear four lophs with posteriorcingulum and have two conelets on the pretrite side. The third and fourth posttrites are similar to the Tha Chang M3 in being tricuspid, but the main cone of the fourth loph pretrite is less transversely elongated than in the Tha Chang M3. The crescentoids are stronger and the size is larger than in the Tha Chang M3 (Tables 1, 3; Fig. 5.2).
Zygolophodon lufengensis from the upper Miocene (Biasatti et al., Reference Biasatti, Wang, Gao, Xu and Flynn2012) of Shihuiba, Lufeng, Yunnan, was described by Zhang (Reference Zhang1982). The holotype, left lower m3 (Zhang, Reference Zhang1982, pl. 1, fig. 1), bears four lophids with small a posterior cingulum, and the antero–external and pretrite cingula are quite strong. Zygodont crests are visible on the first and second lophids. The conelets on both the pretrite and posttrite half lophids are small. Hence, this specimen can be referred to Zygolophodon gobiensis, (i.e., Tobien et al., Reference Tobien, Chen and Li1988) and is similar to the Tha Chang specimens. In addition, the Lufeng m3 is similar in molar size to Miomastodon tongxinensis and the Tha Chang specimen, but the M3 of Lufeng specimen is larger than both Tongxin and Tha Chang specimens. The m3 of Z. lufengensis shows an advanced character state not seen in M. tongxinensis and Z. gobiensis, in that the transverse lophids are rather oblique; however, it is also present in the Tha Chang m3 (Tables 1–3; Fig. 5.1, 5.2).
However, Tobien et al. (Reference Tobien, Chen and Li1988) synonymized many Chinese zygodont proboscideans with Zygolophodon gobiensis, including Zygolophodon gromovae Dubrovo, Reference Dubrovo1974, Z. (Turicius) nemonguensis Chow and Chang, Reference Chow and Zhang1961, Z. nemonguensis Yan, Reference Yan1979 (M3, IVPP2487), Z. jiningensis Chow and Chang, Reference Chow and Zhang1974 (M3, IVPP-5156), Z. chinjiensis Chow and Chang, 1978, Miomastodon tongxiensis Chen, Reference Chen1978, fragments of M1, IVPP-4685.7 and 4685.8, of Gomphotherium xiaolongtanensis Chow and Chang, 1978, from the upper Miocene of Xiaolongtan coal mine, Kaiyuan, Yunnan.
In addition, Tobien et al. (Reference Tobien, Chen and Li1988) transferred some species of Zygolophodon to Mammut borsoni, including an upper D4 (IVPP-V4685) of Zygolophodon sp. from the lower or middle Pliocene of Anle, Huoxian, Shanxi; M3 and m3 (IVPP-VM867 and IVPP-V2485) of Z. shansiensis Chow and Chang, Reference Chow and Zhang1961 and M3 (IVPP-V2488) of Zygolophodon sp. (Chow and Chang, Reference Chow and Zhang1961) from the upper Pliocene or lower Pleistocene of the Yushe Basin, Shanxi; M2 (IVPP-V4689) of Zygolophodon sp. (Chow et al., Reference Chow, Zhang and You1978) from the Pliocene of Zhaotong, Yunnan. All of the above taxa and specimens are larger in terms of molar size than the Tha Chang specimens, as well as having stronger zygodont crests and lophids that are more obliquely aligned to the long axes of the teeth (Tables 1–3; Fig. 5.1, 5.2).
Zygolophodon metachinjiensis Osborn, Reference Osborn1929 is represented by a right hemimandible with m2 and m3 (AMNH 19414) from the middle Miocene of lower Chinji Formation, Chinji, Pakistan. The m3 has four lophids, and a low posterior cingulum consisting of many accessories this cingulum. Molar is subhypsodont (75 mm height) and has no cement, and the summits of the lophids each consist of four to five conelets. Tobien (1972) allocated the holotype, lower m2 and m3, of Z. metachinjiensis to Gomphotherium by used the character of subhypsodonty and relatively large size of m3 that were mentioned by Osborn (Reference Osborn1936, p.456). However, there are a few details in this holotype. In 1983, Tassy synonymized the upper M3 of Z. chinjiensis and lower m2 and m3 of Z. metachinjiensis (Osborn, Reference Osborn1929) to Z. metachinjiensis. In this case, after considering the figures of Osborn (Reference Osborn1929, figs. 4, 5, 5A and A1) and new material of Tassy (Reference Tassy1983a, pl. 2, fig. 2), Tobien et al. (Reference Tobien, Chen and Li1988) suggested the relations are narrow in their proportions, like the lower and upper second and third molars of Z. gobiensis. The pretrite conelets are rather small in Z. metachinjiensis, suggesting this taxon may be closer to Z. gobiensis than to Z. turicensis. If Tobien et al. (Reference Tobien, Chen and Li1988) are right; the oldest name for Z. gobiensis should be Z. metachinjiensis Osborn, Reference Osborn1929. All Zygolophodon species that were synonymized by Tobien et al. (Reference Tobien, Chen and Li1988) should also be changed. In the Tha Chang m3 NRRU-TKK-001, the transverse lophids are so severely worn that the morphology of the pretrite conelets is generally unclear, except the fourth lophid. In the fourth lophid; however, very small conelets are clearly present on both the pretrite and posttrite sides. The third molars of Z. metachinjiensis are much larger, particularly with regard to the length of both the upper and lower molars, than their counterparts from Tha Chang (Tables 1–3; Fig. 5.1, 5.2).
Discussion
In the course of the “proboscidean datum event” (Madden and van Couvering, Reference Madden and van Couvering1976), which actually comprised multiple proboscidean dispersals from Africa to Asia and Europe, mammutids initially immigrated, in the end of early Miocene, into Europe, and thereafter into Asia and South Asia (Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa and Delmer2010). In Europe, Tassy (Reference Tassy1989) identified Zygolophodon turicensis in MN3b sediments in France. In Eurasia and Asia, the records of these taxa in Pakistan, Kazakhstan, and China occur in sediments that date to, or are slightly younger than, MN4 (Tassy, Reference Tassy1996b; Lucas and Bendukidze, Reference Lucas and Bendukidze1997). However, material from the Loh Formation in Central Mongolia appears to be late early to early middle Miocene in age, which would make the specimens in question, slightly younger than the first gomphothere and mammutid immigrants (Göhlich, Reference Göhlich2007). However, Mein (Reference Mein1990) thinks that Zygolophodon arrived at Europe coincident with Gomphotherium during MN4. If the Loh Formation indicates late early to early middle Miocene in age and considering the opinion of Mein (Reference Mein1990), the initially migration of mammutids is still unresolved.
The family Mammutidae is characterized by a zygodont pattern (yoke–like transverse crests) of the intermediate and third molars. The oldest, most primitive mammutid is Losodokodon losodokius, from Kenya dated to the late Oligocene (Rasmussen and Gutierrez, Reference Rasmussen and Gutiérrez2009). In Eurasia the Mammutidae is only represented by two genera, Zygolophodon and its descendant Mammut (Konidaris and Koufos, Reference Konidaris and Koufos2009). The oldest Zygolophodon, Z. aegyptensis, is from the lower Miocene of Wadi Moghara, Egypt (Sanders and Miller, Reference Sanders and Miller2002 in Konidaris and Koufos, Reference Konidaris and Koufos2009). The only European Zygolophodon representative is the abundant Z. turicensis, whose long stratigraphic range in Europe extends from the middle to the upper Miocene (Konidaris and Koufos, Reference Konidaris and Koufos2009). The “Zygolophodon turicensis group” includes four Eurasian species: the type species Z. turicensis Schinz, Reference Schinz1824 from Europe, Z. atavus from Kazakhstan, Z. metachinjiensis from Pakistan and Z. gobiensis from Mongolia and China (Tassy, 1996).
Zygolophodon has not previously been reported from Southeast Asia. The known Zygolophodon specimens closest to the region are from Lufeng, Yunnan, South China, and were identified as Z. lufengensis Zhang, Reference Zhang1982 and Z. chinjiensis (Chow et al., Reference Chow, Zhang and You1978; Zhang, Reference Zhang1982). The new specimens from the Tha Chang sand pits are the first record in Southeast Asia. Z. chinjiensis and Z. lufengensis from Yunnan were assigned to Z. gobiensis by Tobien et al. (Reference Tobien, Chen and Li1988) on the basis of dental morphology.
Tassy (Reference Tassy1985) and Tobien (Reference Tobien1996) noted two dental morphotypes within Mammutidae, robust and gracile, however, the dental morphology in this family is indeed high variable (Tables 2, 3). The zygodont specimens from the Tha Chang sand pits are close in size to the equivalent teeth of Zygolophodon gobiensis (Miomastodon tongxinensis Chen, Reference Chen1978, IVPP-V5584; Z. gobiensis, AMNH 26467, Tobien et al., Reference Tobien, Chen and Li1988) (both lower and upper molars), Z. lufengensis (lower molar) and Z. turicensis (both lower and upper molars), but they are smaller than the holotype of Z. gobiensis, Z. metachijiensis, and Mammut borsoni (Fig. 5.1, 5.2). Tha Chang specimens have small adaxial conelets on both the pretrite and posttrite sides, a feature seen in Zygolophodon specimens from both China and Pakistan. As mentioned above, Tobien et al. (Reference Tobien, Chen and Li1988) synonymized many species of Zygolophodon with Z. gobiensis, implying high variation in the dental morphology of this species. In addition, they suggested that Z. gobiensis was more closely related to Pakistan, Z. metachinjiensis, which share the small pretrite conelets seen in Z. gobiensis, than to Z. turicensis. Nevertheless, the essential diagnostic characters of Z. gobiensis are the small anterior pretrite conelets on the upper and lower second and third molars, the fact that the lophids are only slightly oblique. Tha Chang specimens share some of these characters, but lack others.
Although the Tha Chang specimens are deeply worn, it is still evident that their crescentoids are less inflated than in the type specimen of Z. gobiensis, Miomastodon tongxinensis, and Z. jiningensis but similar in their degree of inflation to late Miocene Z. lufengensis and Z. chinjiensis. However, the lophids are less oblique and the zygodont crests less well developed than in Z. lufengensis and Z. chinjiensis, as in the type specimen and Miomastodon tongxinensis. The strongly oblique loph(id)s and zygodont crests are more remarkable in the derived zygodont proboscideans such as Mammut (Osborn, Reference Osborn1936; Tobien et al., Reference Tobien, Chen and Li1988). The broken specimen of Z. gromovae has a loph–like rather than conule–like fourth loph, a point of resemblance to the Tha Chang specimen. In contrast, the fourth loph of the upper M3 of Tha Chang specimen is better developed than in Z. gromovae. South Chinese species have small and strongly divided posttrite adaxial conelets, making them more similar to species from the Pakistan (Z. metachinjiensis) than those from North China (Z. gobiensis, Miomastodon tongxinensis, Z. nemonguensis, and Z. jiningensis). The Tha Chang specimens share specific features with the zygodonts from the Pakistan and Yunnan such as small adaxial conelets, however, they are deeply worn and broken in the anterior loph(id)s and so it is difficult to establish their precise affinities though it is reasonable to designate them as Zygolophodon sp.
The faunal assemblage in the Tha Chang sand pits no. 1 to 9 includes proboscideans, anthracotheres, pigs, rhinos, bovids, giraffids, horses, apes, crocodiles, and tortoises (Nakaya et al., Reference Nakaya, Saegusa, Ratanasthien, Kunimatsu, Nagaoka and Suganuma2002; Chimanee et al., Reference Chaimanee, Suteethorn, Jintasakul, Vidthayanon, Maradat and Jaeger2004) (Fig. 1), and indicates a middle Miocene to early Pleistocene age. The middle Miocene mammalian fauna consists of amebelodontine gomphotheres (Protanancus), Gomphotherium, and Prodeinotherium. The late Miocene to early Pliocene fauna includes a new orangutan species Khoratpithecus piriyai Chimanee et al., Reference Chaimanee, Suteethorn, Jintasakul, Vidthayanon, Maradat and Jaeger2004, a new anthracothere species Merycopotamus thachangensis Hanta et al., Reference Hanta, Ratanasthien, Kunimatsu, Saegusa, Nakaya, Nagaoka and Chintasakul2008, a new rhino species Aceratherium porpani Deng, Hanta, and Jintasakul, Reference Deng, Hanta and Jintasakul2013 (from the same sand pit that yielded the Zygolophodon specimens, Tha Chang sand pit no. 10), horse Hipparion, proboscidean Stegolophodon and a primitive species of Stegodon. The early Pleistocene fauna includes an advanced species of Stegodon (Nakaya et al., Reference Nakaya, Saegusa, Ratanasthien, Kunimatsu, Nagaoka and Suganuma2002, Thasod, 2012). Furthermore, Thasod and Ratanasthien (Reference Thasod and Ratanasthien2005) reported a new species of Sinomastodon more derived than Sinomastodon intermedius from the Pliocene of China. Sinomastodon materials have also been reported from Java (Saegusa, Reference Saegusa1995 in van den Bergh, Reference Bergh1999). These specimens were referred to S. bumiajuensis, but their skull and dental features resemble those of Chinese sinomastodonts. In addition, the molars of S. bumiajuensis are more derived than those of Pliocene sinomastodonts from Japan and North China, but very similar to Sinomastodon yangziensis remains from early Pleistocene cave faunas in South China (van den Bergh, Reference Bergh1999).
Although the Zygolophodon specimens from the Tha Chang sand pits were discovered by sand pit workers and their stratigraphic positions are uncertain, their many dental features are similar to the Yunnan species such as in molar size, four lophids with small a posterior cingulum, strong antero–external and pretrite cingula, zygodont crests on the first and second lophids, and small conelets on both the pretrite and posttrite half lophids. In addition, the geographical localities are nearby each other. Furthermore, the teeth of zygodont proboscideans from South China and Pakistan are more gracile or mixed between gracile and robust forms than those of northern Chinese species, which are more robust. The Yunnan specimens have been dated to the late Miocene (Biasatti et al., Reference Biasatti, Wang, Gao, Xu and Flynn2012), and those from Pakistan occur in the lower Chinji Formation (about 14–13 Ma) (Osborn, Reference Osborn1929; Chavasseau et al., Reference Chavasseau, Chaimanee, Yamee, Tian, Rugbumrung, Marandat and Jaeger2009). The North China species have been dated to the middle Miocene (Tobien et al., Reference Tobien, Chen and Li1988), and might be older than the species from Pakistan. Because of similarity to the Yunnan species regarding both dental morphology and geographical locality, thus, the Tha Chang specimens might be late Miocene in age. However, the size and morphological variation of such teeth make stratigraphic conclusions very tentative.
Among the previously described middle to late Miocene proboscideans from northern Thailand are in Mae Moh coal mine (13.3 and 13.1 Ma), Lam Pang Province include Stegolophodon praelatiden and Gomphotherium cf. browni (Chavasseau et al., Reference Chavasseau, Chaimanee, Yamee, Tian, Rugbumrung, Marandat and Jaeger2009). Then, the Chiang Muan coal mine (12.4–12.2 Ma), Prayao Province, has produced Tetralophodon cf. xiaolongtanensis (Thasod, Reference Thasod2007; Chavasseau et al., Reference Chavasseau, Chaimanee, Yamee, Tian, Rugbumrung, Marandat and Jaeger2009). The recognition of G. cf. browni in northern Thailand implies a degree of biogeographic affinity with Pakistan, whereas the presence of Tetralophodon cf. xiaolongtanensis in Chiang Muan is a clear link with the fauna of South China. The most widespread taxa in the middle Miocene of Pakistan are the mammutid Zygolophodon metachinjiensis, the amebelodont Protanancus chinjiensis, the choerolophodont Choerolophodon corrugatus and the gomphothere Gomphotherium browni (Tassy, Reference Tassy1983a, Reference Tassy1983b, fig. 7). The first three taxa have apparently not been recorded from the middle Miocene of Thailand, which points to obvious faunal dissimilarities between Thailand and Pakistan (Chavasseau et al., Reference Chavasseau, Chaimanee, Yamee, Tian, Rugbumrung, Marandat and Jaeger2009). The middle Miocene localities of North China have yielded a very different fauna dominated by Zygolophodon, the amebelodont Platybelodon, and supposedly endemic Gomphotherium species (Tobien et al., Reference Tobien, Chen and Li1986; Tobien et al., Reference Tobien, Chen and Li1988, fig. 7). Thus, the new Zygolophodon specimens in Tha Chang sand pit might indicate a dispersal link between South and East Asian middle and late Miocene proboscideans.
The late Miocene paleoenvironmental record from the Tha Chang sand pits indicates a transition from woodland-dominated to grassland-dominated landscapes (Sepulchre et al., Reference Sepulchre, Jolly, Ducrocq, Chaimanee, Jaeger and Raillard2010). These results can be correlated to a long-term climatic change that occurred between 8.5 and 6 Ma, leading to a major environmental change with C4 savannas replacing C3 forests and woodlands (Cerling et al., Reference Cerling, Quade and Wang1994; Sepulchre et al., Reference Sepulchre, Jolly, Ducrocq, Chaimanee, Jaeger and Raillard2010). However, the late late Miocene new rhino species, Aceratherium porpani, from the same sand pit as the Zygolophodon materials, is consistent in its chronological implications with the faunal sample from Tha Chang sand pit no. 8, which includes such taxa as Khoratpithecus piriyai and Merycopotamus thachangensis (Chaimanee et al., Reference Chaimanee, Suteethorn, Jintasakul, Vidthayanon, Maradat and Jaeger2004, Reference Chaimanee, Yamee, Tian, Khaowiset, Marandat, Tafforeau, Nemoz and Jaeger2006; Hanta et al., 2005, Reference Hanta, Ratanasthien, Kunimatsu, Saegusa, Nakaya, Nagaoka and Chintasakul2008; Deng et al., Reference Deng, Hanta and Jintasakul2013). The occipital surface of the Tha Chang rhino is slightly inclined posteriorly or nearly vertical and the cheek teeth are subhypsodont (Deng et al., Reference Deng, Hanta and Jintasakul2013). Both features indicate a woodland habitat (Zeuner, Reference Zeuner1934).
The zygodont proboscidean discovered in Thailand has significant implications for understanding the dispersal of Zygolophodon. The oldest zygodont proboscidean in Africa, Losodokodon losodokius, is dated to late Oligocene (Rasmussen and Gutierrez, Reference Rasmussen and Gutiérrez2009). The oldest Zygolophodon in Europe is in the late early Miocene, either MN3b or MN4 (approximately 17 Ma) (Drake in Tassy, Reference Tassy1990) while the Zygolophodon materials from Pakistan are in the lower Chinji Formation (approximately 14–13 Ma) (Osborn, Reference Osborn1929; Chavasseau et al., Reference Chavasseau, Chaimanee, Yamee, Tian, Rugbumrung, Marandat and Jaeger2009). The North Chinese species are from the middle Miocene (Tobien et al., Reference Tobien, Chen and Li1988) and might be older than the species from Pakistan. Finally, the South Chinese species are from the upper Miocene (Biasatti et al., Reference Biasatti, Wang, Gao, Xu and Flynn2012) (Fig. 6). Zygolophodon appears to have dispersed from Africa to Europe in the early Miocene, and become widespread in Asia (especially at middle and high latitudes) in the middle Miocene. Zygolophodon then shifted to low latitude areas such as Thailand in the late Miocene (Fig. 6).
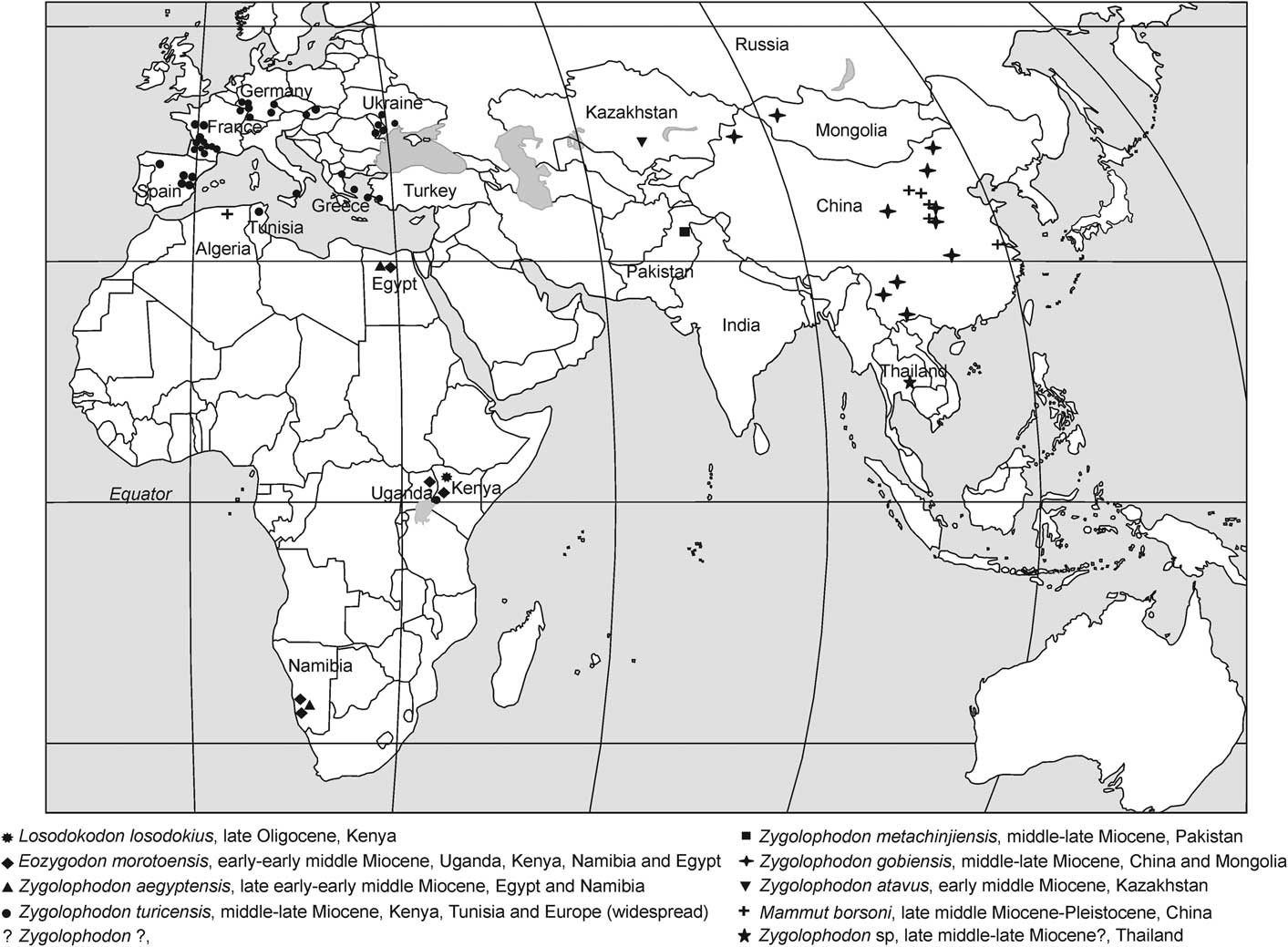
Figure 6 Distribution of zygodont proboscideans in the Old World that include the new specimens from Thailand. The oldest zygodont proboscideans have presented in the late Oligocene of Africa and they migrated to Europe and Asia in Miocene (modified data from Tobien et al., Reference Tobien, Chen and Li1988; Guan, 1996 in Shoshani and Tassy, Reference Shoshani and Tassy1996; Sanders and Miller, Reference Sanders and Miller2002; Pickford, Reference Pickford2007; NOW, 2007; Konidaris and Koufos, Reference Konidaris and Koufos2009; Rasmussen and Gutierrez, 2009).
Conclusion
The zygodont proboscidean teeth from the Tha Chang sand pits (no. 10) show the zygodont pattern. The adaxial conelets are small, more closely resembling Chinese specimens of Zygolophodon gobiensis and Pakistan species Z. metachinjiensis than the holotype species Z. turicensis. Despite deep wear, it is evident that the central conules are less inflated than those of the type specimen of Z. gobiensis and Miomastodon tongxinensis, but similar to Z. lufengensis and Z. chinjiensis from the upper Miocene of South China. However, the Tha Chang specimens are badly worn and the m3 is broken anteriorly, making it only to designate these specimens as Zygolophodon sp. This Zygolophodon is the first zygodont proboscidean recorded in Southeast Asia and is likely late Miocene in age.
Acknowledgments
This paper was completed with the help of several people. We would especially like to thank P. Vachajitpan for allowing us to study his collection. We wish to thank to people of the Institute of Vertebrate Paleontology and Paleoanthropology including T. Haowen, who gave us generous suggestions about the proboscideans morphology; C. Sullivan, who advised us about English; and C. Jin, who generously provided us with a comfortable workspace as we studied the specimens in the institute’s collections room., We would like to thank to the staffs of the Research Institute of Petrified Wood and Mineral Resources and Nakhon Ratchasima Rajabhat University, especially W. Songtham, W. Rugmai, N. Boonchai, P.J. Grote, K. Jenjitpaiboon, P. Kachenchat, S. Sodok, S. Ruanjangreed, N. Kullaprutmetha, and P. Pipatwatchara for preparing samples and documents as well as making some valuable suggestions while we were studying samples in Thailand. We thank N. Chanon, who gave us a vehicle for comfortable transportation during our work in Thailand. We are grateful for financial support from the National Natural Science Foundation of China (41430102), the Institute of Vertebrate Paleontology and Paleoanthropology, the University of Chinese Academy of Sciences, and Nakhon Ratchasima Rajabhat University.









