




Introduction
The ‘Two Layer Hypothesis’ in Southeast Asia describes the expansion of farming communities southward from East Asian centres of rice and millet domestication into a region occupied for millennia by foragers (Higham et al. Reference Higham, Douka and Higham2015). Foragers formed the first layer of occupation in Southeast Asia, whereas farmers from China, who are presumed to have cultivated rice as the dominant domesticate, moved south into territories occupied by these foragers as a second layer of occupation c. 2000 BC. Initial archaeobotanical research in eastern-central Thailand, however, has revealed that the earliest farming in Southeast Asia involved millet (Weber et al. Reference Weber, Lehman, Barela, Hawks and Harriman2010). Employing data from one of the largest collections of archaeobotanical material in Southeast Asia, we examine over 3000 years of changing farming practices in eastern-central Thailand, a region that played an important role in the development of metallurgy and social complexity in Southeast Asia (see Higham et al. Reference Higham2020). Our research focuses on four sites: Non Pa Wai, Nil Kham Haeng and Non Mak La, located in the Khao Wong Prachan Valley, and Phromtin Tai to the north-west, on the Lopburi Plain (Figure 1).

Figure 1. Location of sites mentioned in the text. A: 1) Non Pa Wa; 2) Nil Kham Haeng; 3) Non Mak La; 4) Phromtin Tai; 5) Nong Ratchawat; 6) Khok Phanom Di 7) Ban Non Wat; 8) Noen U-Loke; 9) Non Ban Jak; 10) Khao Sam Kaeo; 11) Phu Khao Tong; 12) Rach Nui; 13) Haimenkou. B: 1) Non Pa Wai; 2) Nil Kham Haeng; 3) Non Mak La; 4) Phromtin Tai; 5) Nong Ratchawat; 6) Khok Phanom Di; 7) Ban Non Wat; 8) Noen U-Loke (figure made by C.F.W. Higham using GeoMapApp (https://urldefense.com/v3/__http://www.geomapapp.org__;!!Mih3wA!QFNtjNA_zEWfMv4616kXxCuTgH_PRNyh_Y8I6Gxe0Vg_SopNytzjTNbCJHmsVvg$); CC BY (Ryan et al. Reference Ryan2009)).).
Following a number of scholars (Mudar Reference Mudar1995; Mudar & Pigott Reference Mudar, Pigott, Karlstrom and Kallen2003; Pigott et al. Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006; Weber et al. Reference Weber, Lehman, Barela, Hawks and Harriman2010), we argue that the first farming communities in the Khao Wong Prachan Valley practised dryland/upland millet-based agriculture, supplemented by a substantial element of wild plant foraging. Mudar and Pigott (Reference Mudar, Pigott, Karlstrom and Kallen2003) and Pigott et al. (Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006) have postulated that this strategy was employed only during the earliest phases of occupation in the Khao Wong Prachan Valley sites, and that the later higher productivity of wet rice agriculture (i.e. planting seeds on dryland and transferring the seedlings to a flooded field) was necessary to meet increasing population pressure. Furthermore, they argued that, from c. 1100 BC, the inhabitants of the Khao Wong Prachan Valley began to mine, smelt and cast copper to exchange agricultural products with those living in areas more suited to wet rice agriculture.
Our results concur with previous phytolith studies in the Khao Wong Prachan Valley that demonstrate the existence of upland farming regimes (Kealhofer Reference Kealhofer2002). We argue that sufficient agricultural production of upland (dryland) rice cultivation and small amounts of millet were integrated into local systems of foraging, and later combined with swiddening in ancient central Thailand. These non-irrigated systems of farming continued to play an important part in subsistence for millennia, persisting into the present day.
While the Two Layer Hypothesis is now widely accepted, based on archaeological, linguistic and human biological evidence (Bellwood & Renfrew Reference Bellwood and Renfrew2003; Bellwood Reference Bellwood and Stark2006; Rispoli Reference Rispoli2007; McColl et al. Reference McColl2018; cf. White Reference White and Enfield2011), the finer details of the migratory routes taken by the first millet and rice farmers in Southeast Asia are only now being identified. Some scholars have highlighted coastal routes starting in Taiwan and Fujian (e.g. Bellwood Reference Bellwood1991; Bellwood & Dizon Reference Bellwood, Dizon, Sanchez-Mazas, Blench, Ross, Peiros and Lin2008), while others have suggested routes from the Middle Yangtze to Lingnan, Guangdong and Guangxi, with populations entering Southeast Asia through Vietnam (e.g. Rispoli Reference Rispoli2007; Fuller et al. Reference Fuller, van Etten, Manning, Castillo, Kingwell-Banham, Weisskopf, Qin, Sato and Hijmans2011). Still others have highlighted the role played by higher-altitude areas to the west, such as the Yunnan Guizhou Plateau (e.g. Higham Reference Higham2002). A similar debate has involved the direction of transmission of copper/bronze metallurgy, with some authors viewing the ultimate origin of this technology in the Seima-Turbino horizon of the Altaï region, which then travelled along the Eastern Himalayas via Gansu, Sichuan and Yunnan (White & Hamilton Reference White, Hamilton, Roberts and Thornton2014). The alternative mode sees the Steppic technology reaching the Chinese Central Plain and being transmitted south along lowland routes in the Central Yangtze region into Lingnan and Yunnan and then on to Southeast Asia (Pigott & Ciarla Reference Pigott and Ciarla2007; Higham et al. Reference Higham, Higham, Ciarla, Douka, Kijngam and Rispoli2011, Reference Higham, Douka and Higham2015). Here, we apply a computational model to examine how the agricultural niche was affected by changes in ancient climate and to determine which areas of early agricultural systems may have come under adaptive stress.
Sites and excavations
Data from Non Pa Wai, Non Mak La and Nil Kham Haeng were obtained by the Thailand Archaeometallurgy Project, and those from Phromtin Tai were acquired during excavations by Thanik Lertcharnrit. We combined these data to evaluate how farming strategies were modified from the earliest Neolithic to the late historic period.
Phromtin Tai, a moated settlement and burial site with evidence for copper production occupied from the Iron Age to the Dvaravati period (c. 500 BC to tenth century AD), is located close to the floodbanks of the Chao Phraya River (d'Alpoim Guedes et al. Reference d'Alpoim Guedes, Hanson, Higham, Higham and Lertcharnrit2019). Non Pa Wai, Non Mak La and Nil Kham Haeng are located in the Khao Wong Prachan Valley in Lopburi Province on the eastern margin of the Chao Phraya River Valley (Figure 1) (Pigott et al. Reference Pigott, Weiss, Natapintu, Ciarla and Rispoli1997; Rispoli et al. Reference Rispoli, Ciarla and Pigott2013; Pigott Reference Pigott, White and Hamilton2019). They all lie close to valley copper ore sources, and have yielded evidence for settlement and copper smelting. Non Pa Wai and Nil Kham Haeng, both approximately 5ha in size, are among the largest known prehistoric copper-smelting sites in Asia. Non Mak La, an even larger multi-purpose settlement and burial site, has yielded only modest copper-smelting evidence. Our current understanding is that the earliest occupation in the Khao Wong Prachan Valley dates to the late third millennium BC. Copper-base metallurgy began towards the end of second millennium BC, with intensive production in the following millennium. Together, these sites are one of only two major prehistoric copper-production centres known in Southeast Asia (Pigott Reference Pigott, White and Hamilton2019; Higham et al. Reference Higham2020). Although the final reports of these sites remain unpublished, significant research is in print (see Pigott et al. Reference Pigott, Weiss, Natapintu, Ciarla and Rispoli1997; Weber et al. Reference Weber, Lehman, Barela, Hawks and Harriman2010; Rispoli et al. Reference Rispoli, Ciarla and Pigott2013; Higham et al. Reference Higham2020).
We report here the results of the analysis of 247 samples containing charred seeds for which we have contextual information. Seeds were found in approximately 30 per cent of floated samples (see the online supplementary material (OSM)). We focus our analysis on the temporal distribution of these remains over several phases (Table 1; full results in the OSM, Table S1).
Table 1. Phases present at the sites discussed.

Results
Although we recovered only small quantities of archaeobotanical remains from Neolithic phase 1, no rice caryopses, husks or spikelets were found. Remains from phase 1 consist entirely of foxtail millet (Setaria italica), confirming patterns previously noted by Weber et al. (Reference Weber, Lehman, Barela, Hawks and Harriman2010). Only a few samples from the later Neolithic (phase 2) components of Non Pa Wai and Non Mak La contained seeds. While rice remains are present, foxtail millet remains the numerically dominant crop throughout the Early Bronze Age (phases 3 and 4). Rice progressively increases throughout the assemblage. By phase 5 (the Late Bronze Age), rice comprises half the raw seed count, and by the Iron Age (phase 6) and following the Dvaravati phase (phase 7), it dominates the raw count at Phromtin Tai (Figure 2).

Figure 2. Grain-crop proportions by phase from Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai. Note that phase 6 represents a combination of samples from both Phromtin Tai and the Thailand Archaeometallugy Project sites; N = number of samples; numbers on histogram are numbers of charred seeds (figure by J. d'Alpoim Guedes).
Rice spikelets from assemblages in China after 4000 BP show high proportions of domesticated spikelet bases. At Neolithic Baiyangcun in Yunnan (third millennium BC), for example, 76 per cent of spikelet bases were from domesticated rice (Dal Martello et al. Reference Dal Martello, Min, Stevens, Higham, Higham, Qin and Fuller2018). In our samples, domesticated spikelets account for only 20 per cent of the total assemblage, suggesting that local varieties of wild rice (Oryza rufipogon) were gathered alongside introduced and domesticated rice. Although the numbers of rice spikelets are too low in phases 2–4 for meaningful statistical analyses, large quantities of wild spikelets are present in phases 4/5–6 (Figure 3). An increase in the number of domesticated spikelets in the Dvaravati phase (phase 7) is only seen at Phromtin Tai. Many spikelets in our assemblage, however, were poorly preserved, and it is possible that this has skewed our results.
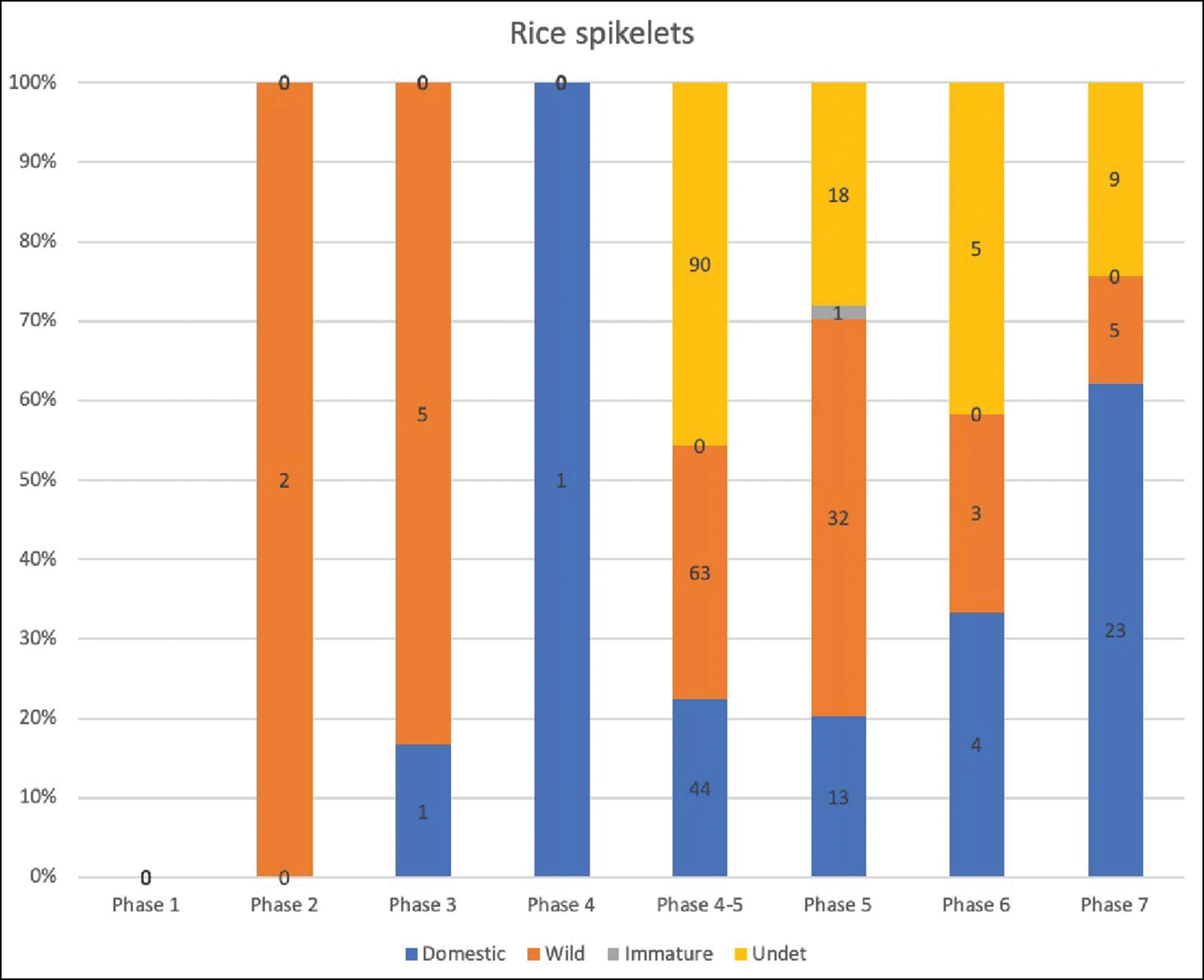
Figure 3. Proportions of rice spikelets by phase at Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai; N = number of samples; numbers on histogram are numbers of seeds (figure by J. d'Alpoim Guedes).
One specimen of pigeon pea (Cajanus cajan) appears during phase 3 at Non Pa Wai (Figure 4). The natural range of Cajanus sp. is in South Asia, where some of the earliest domesticated examples date to c. 1500 BC (Fuller et al. Reference Fuller, Murphy, Kingwell-Banham, Castillo and Naik2019). Pigeon pea has been found at other sites in Southeast Asia, including Khao Sam Kaeo and Phu Khao Tong (Figure 1), although its chronological context is unclear (Castillo et al. Reference Castillo, Bellina and Fuller2016a). Depending on the age of these specimens, the pigeon pea at Non Pa Wai may be one of the earliest in Southeast Asia. Several fragments of another unidentified Fabaceae, possibly Vigna sp., appear in phases 4–5 at Non Pa Wai. The presence of cotton (Gossypium sp.) at Phromtin Tai in phase 7 also suggests contact with South Asia (Figure 4) (see d'Alpoim Guedes et al. Reference d'Alpoim Guedes, Hanson, Higham, Higham and Lertcharnrit2019).

Figure 4. Key domestic pulse and grain taxa from Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai: A) rice (Oryza sativa); B) foxtail millet (Setaria italica); C) cotton (Gossypium sp.) funicular cap; D–E) pigeon pea (Cajanus cajan); F) unidentified Fabaceae (figure by J. d'Alpoim Guedes).
Fruit remains were also present at our sites. Citrus rind begins to appear in phase 1 of Non Pa Wai. Taxa include Citrus medica (lemon or lime), potentially native to the southern flanks of the Himalayas and Citrus reticulata (oranges or mandarins), also possibly native to South Asia. The rind of Citrus maxima (pomelo), whose potential centre of domestication may be in Southeast Asia, has been identified at Khao Sam Kaeo and Phu Khao Tong (Castillo et al. Reference Castillo, Bellina and Fuller2016a & b). Citrus maxima has marked raised pusticulae (warts) on the rind's surface (Fuller et al. Reference Fuller, Castillo, Weisskopf, Kingwell-Banham, Qin, Zech-Matterne and Fiorentino2017), a feature not visible on the specimen we recovered in phase 4 at Non Mak La. This specimen appears to conform most closely to C. medica, which has fewer visible oil glands that are angular and squarish in appearance (Fuller et al. Reference Fuller, Castillo, Weisskopf, Kingwell-Banham, Qin, Zech-Matterne and Fiorentino2017) (Figure 5). This may show further connections with South Asia (Rispoli et al. Reference Rispoli, Ciarla and Pigott2013: 150–57). Two additional, tiny and unidentifiable citrus rind fragments were found; one in phase 2 of Non Mak La, and one in phase 3 of Nil Kham Haeng.

Figure 5. Key fruit taxa unearthed at Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai: A) Indian jujube (Ziziphus mauritiana); B–C) cf. Citrus maxima (pomelo) rind; D–E) pygmy date palm (Phoenix roebelenii); F) Cassia sp. (figure by J. d'Alpoim Guedes).
Several other wild foraged fruit remains were also found in our assemblage. The number of seeds of a wild date palm fruit, probably of the genus Phoenix because of its oval shape and large groove running along the ventral side (Figure 5), increased over time in the assemblage. While some Phoenix species are native to the Indian subcontinent, Phoenix roebelenii is native to south China and Southeast Asia, and its fruits are edible (Barrow Reference Barrow1994; Frodin & Govaerts Reference Frodin and Govaerts2002). At 5mm in length, our specimens are smaller than examples reported in the literature (Frodin & Govaerts Reference Frodin and Govaerts2002). Palm leaf epidermis phytoliths were also recovered at Non Mak La, suggesting that palm trees were encouraged to grow near the site, or that palm leaves were regularly employed for economic purposes (Kealhofer Reference Kealhofer2002).
Other fruits in the assemblage include one specimen of the Indian jujube (Ziziphus mauritiana) in phases 4–5 at Nil Kham Haeng, and a single specimen of Cassia (Cassia sp.) from phase 4 at Non Pa Wai (Figure 5). Both species are probably native to Southeast Asia. Indian jujube is an important economic fruit; as for Cassia sp., its leaves and flowers are edible, and its wood may have also been used.
Seeds traditionally considered to be weeds were identified in substantial numbers, including aster (Acmella paniculata), Chenopodium-amaranth perisperms, Trianthema sp. and a wild member of the Aveneae tribe (Figure 6). All these ‘weedy’ taxa are found primarily in dryland habitats (Caton et al. Reference Caton, Mortimer and Hill2004). Despite the increasing importance of rice, these dryland weeds form the majority of the assemblage, suggesting that the rice at these central Thailand sites was grown in a rain-fed upland and not in an irrigated paddy environment. Acmella paniculata was identified in Bronze Age contexts at Ban Non Wat, where it was considered as evidence for dryland (i.e. rain-fed) rice cultivation (Castillo et al. Reference Castillo, Fuller, Piper, Bellwood and Oxenham2018a).

Figure 6. Weed seed proportions at Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai by phase (figure by J. d'Alpoim Guedes).
A few wetland weeds such as Fimbristylis sp. (sedge) are present, but in small amounts (Figure 7), a result that is consistent with previous research showing very low numbers of sedges but high numbers of panicoid and bambusoid grasses and members of the Asteraceae family (Kealhofer Reference Kealhofer2002). All these weeds are common in open woodlands maintained by regular burning regimes (Kealhofer Reference Kealhofer2002; Pigott et al. Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006). Many of these ‘weedy’ taxa also serve important functions in modern Thai cuisine: leaves of Acmella and Chenopodium (goosefoot), for example, are important greens in stir-fries, and hence mechanisms other than land management could explain their presence in our assemblages (see d'Alpoim Guedes et al. Reference d'Alpoim Guedes, Hanson, Higham, Higham and Lertcharnrit2019).

Figure 7. Key weedy taxa recovered at Non Pa Wai, Non Mak La, Nil Kham Haeng and Phromtin Tai: A) sedge (Cyperaceae); B) Chenopodium-amaranth perisperm; C) Aveneae tribe; D) aster (Acmella paniculata); E) Trianthema sp.; F) Panicoid grass (figure by J. d'Alpoim Guedes).
Field ecology and cultivation practices in central Thailand
The weedy taxa in our assemblage come from a primarily upland (or rain-fed) environment. As foxtail millet is a rain-fed crop, it is possible that some of the weedy taxa from these sites came from millet fields. Rice can be grown in both upland (rain-fed) and lowland (irrigated, or paddy) habitats (Fuller & Weisskopf Reference Fuller and Weisskopf2011). Despite an increasing proportion of rice in our assemblage, the high proportion of upland weeds and almost negligible amount of wetland weeds suggests that rice may have been cultivated as a dryland, rain-fed crop. Irrigated rice requires plants to be in water during the establishment of shoots, and dry at the time of flowering and grain production (Fuller & Weisskopf Reference Fuller and Weisskopf2011). This demands either an artificial or natural paddy environment: one that can maintain stands of water around plants, but also allows for this water to be drained at specific times of the crop growth cycle. Rain-fed rice, particularly wild varieties, rely on monsoonal flooding of lake edges, followed by drying (White Reference White1995).
The large quantity of silicified (i.e. turned to silica) rice husks in our samples suggest that rice-crop-processing probably took place onsite, the rice being cultivated nearby and transported to the site in sheaths. It is therefore unlikely that this rice was imported in exchange for metal goods (Mudar & Pigott Reference Mudar, Pigott, Karlstrom and Kallen2003).
Rain-fed (upland) varieties of rice generally grow only in areas where annual rainfall exceeds 800–1000mm (Yoshida Reference Yoshida1981; Jacquot & Courtois Reference Jacquot and Courtois1987). Today, the Lopburi Plain has sufficient rainfall (averaging 1200–1300mm annually) to support both rain-fed millet and rice. A reduction in rainfall of between 200 and 300mm would render the cultivation of rain-fed rice marginal. Frequent droughts occur in the area today and may have made rice cultivation less reliable than millets in the past. This may explain the continued presence of foxtail millet in our assemblage, until at least the Iron Age (Figure 4). Millets may have been attractive crops for Southeast Asian foragers, who had begun to interact with shifting cultivators, because millets have short growing seasons (as little as 60–90 days) and do not require investment in irrigation (d'Alpoim Guedes Reference d'Alpoim Guedes2011; d'Alpoim Guedes & Butler Reference d'Alpoim Guedes and Butler2014).
The high proportion of wild-type spikelets in our samples may indicate that a significant number of wild plants (O. rufipogon) were growing in the fields. This may have reduced the yield of rice, reinforcing the importance of millet for consistently reliable yields. Indeed, White's (Reference White1995) ethnographic work in north-east Thailand has demonstrated that wild varieties of rice still grow along streams and lake edges during the rainy season. It is possible that this wild rice was gathered by local foragers prior to the arrival of domesticated rice into the Khao Wong Prachan Valley. The inhabitants of the sites discussed here may have continued this tradition.
Compared to other sites in Southeast Asia and China, both our weed flora data and previous phytolith work (Pigott et al. Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006) from central Thailand suggest that the inhabitants invested less labour in the construction and maintenance of irrigated paddy fields. Instead, they continued to focus on less intensive dryland rice and millet cultivation systems. Pigott et al. (Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006) and Rispoli et al. (Reference Rispoli, Ciarla and Pigott2013) have noted that plough animals, such as cattle and water buffalo, are rare in the Non Pa Wai faunal assemblages well into the Iron Age, suggesting that local agriculture did not involve much use of animal traction or ploughs. This is particularly evident when compared to other Iron Age sites in Thailand, such as Noen U-Loke, where an iron ploughshare was found as a funerary offering, and a palimpsest of cattle and water buffalo hoof prints have been interpreted as draught animal enclosures (Castillo et al. Reference Castillo2018b). The presence of sickles and hoes at Tha Kae near the Khao Wong Prachan Valley (Rispoli et al. Reference Rispoli, Ciarla and Pigott2013) and at Phromtin Tai suggests less intensive modification of the local landscape. We argue that rice-cultivation practices in early central Thailand mimicked the habitat and intensity with which indigenous wild rices were probably gathered.
This pattern of low-intensity gathering and upland farming is also reflected in our sites’ faunal records, which show high proportions of wild animals compared to domesticated taxa, such as pigs and dogs (Pigott et al. Reference Pigott, Mudar, Agelarakis, Kealhofer, Weber, Voelker, Bacus, Glover and Pigott2006), suggesting a continuation of traditions begun by local foragers.
Climatic factors and the spread of agriculture to central Thailand
The direction from which both early farming and metallurgy spread from East Asia into central Thailand has long been unclear, although certain pottery styles in the earliest layers at sites in central Thailand (e.g. Non Pa Wai and Tha Kae) suggest strong connections with sites such as Baiyangcun and Dadunzi in Yunnan (Rispoli Reference Rispoli2007; Rispoli et al. Reference Rispoli, Ciarla and Pigott2013)—connections that continued into the Bronze Age. A deep-socketed axe from a burial at Non Pa Wai, for example, shares similarities with axe moulds from Haimenkou in Yunnan Province (Higham et al. Reference Higham, Higham, Ciarla, Douka, Kijngam and Rispoli2011; Chiou-Peng Reference Chiou-Peng, Linduff and Rubinson2018).
Our analysis confirms that millets were probably the earliest crops introduced to the Khao Wong Prachan Valley, arriving by c. 2300 cal BC (following Weber et al. Reference Weber, Lehman, Barela, Hawks and Harriman2010). Foxtail millet cultivation was practised along the margins of the Tibetan Plateau in China as early as c. 3500 BC (d'Alpoim Guedes Reference d'Alpoim Guedes2011; d'Alpoim Guedes et al. Reference d'Alpoim Guedes, Manning and Bocinsky2016). Millets appear in our samples by c. 2300 cal BC, shortly after appearing on the Chengdu Plain (c. 2700 BC) and in Yunnan Province (c. 2600 cal BC), where they are coupled with finds of rice (d'Alpoim Guedes et al. Reference d'Alpoim Guedes, Jiang, He, Wu and Jiang2013; Jin et al. Reference Jin, Xu, Rui, Li, Wu and Hein2014; Dal Martello et al. Reference Dal Martello, Min, Stevens, Higham, Higham, Qin and Fuller2018). Both crops also appear together in archaeobotanical assemblages in south-eastern China, c. 2200 BC (Deng et al. Reference Deng, Hung, Fan, Huang and Lu2017). The initial introduction of millets without rice into the Khao Wong Prachan Valley suggests that millets may have been part of an early spread of domesticates transmitted via the Eastern Himalayas through the Yunnan-Guizhou Plateau, a transmission that took place before that of rice farming.
It has been argued that early rice in Thailand was primarily Oryza sativa, subspecies japonica, which moved into Southeast Asia from East Asia (Castillo & Fuller Reference Castillo, Fuller and Bellina2010; Castillo et al. Reference Castillo2016b). Recent genetic data on rice from Khao Sam Kaeo, Phu Khao Tong, Ban Non Wat and Noen U-Loke show that early archaeobotanical rice in Thailand conforms to a japonica cultivar (Castillo et al. Reference Castillo2016b), rather than Oryza sativa subspecies indica. The latter was probably domesticated in South Asia following introgression with japonica species (Fuller et al. Reference Fuller, Sato, Castillo, Qin, Weisskopf, Kingwell-Banham, Song, Ahn and van Etten2010). Our samples contain too few measurable grains to confirm this for the earliest examples of rice at the sites examined here. Following Castillo and Fuller (Reference Castillo, Fuller and Bellina2010), however, we presume that the rice found in central Thailand was a japonica rather than an indica cultivar. Both tropical and temperate varieties of O. japonica exist, and genetic evidence reveals that tropical O. japonica is probably ancestral to temperate O. japonica (Garris et al. Reference Garris, Tai, Coburn, Kresovich and McCouch2005; Sweeney & McCouch Reference Sweeney and McCouch2007). Today, the range of tropical O. japonica is limited to Southeast Asia and far southern China. In the warmer Early Holocene (before 2000 cal BC), however, this crop may have ranged as far as north-eastern China (d'Alpoim Guedes & Bocinsky Reference d'Alpoim Guedes and Bocinsky2018).
After 2000 cal BC, increasingly cooler temperatures led to a retreat of the area available for cultivating tropical varieties of O. japonica to south-eastern China and Southeast Asia (Figure 8). By 1590 cal BC, farmers of tropical rice varieties in both Yunnan and the Central Yangtze region would have already begun to experience difficulties in cultivating this crop (d'Alpoim Guedes & Bocinsky Reference d'Alpoim Guedes and Bocinsky2018; Gutaker et al. Reference Gutaker2020) (Figure 8). While large parts of Zhejiang and Hunan lost their suitability for tropical O. japonica, the higher-altitude Yunnan-Guizhou Plateau suffered the largest loss of area suitable for this type of rice. Temperate varieties of O. japonica appear to have fared better, even if their range was compromised, particularly in Yunnan and Guizhou (Figure 8).

Figure 8. Changes in the probability of rice being in the thermal niche at key time intervals throughout the Late Holocene: blue) areas where growing rice is below the 70 per cent threshold of being in the niche; red) areas where rice is above this threshold (map created using methodology published in d'Alpoim Guedes and Bocinsky (Reference d'Alpoim Guedes and Bocinsky2018); figure by Jade d'Alpoim Guedes).
It is possible that the reduced territory suited to rice farming may have led Yunnan farmers who practised metallurgy in the later second millennium BC to either intensify their trade relationships, or to seek warmer areas in which to cultivate tropical O. japonica. Wild varieties of tropical japonica rice could also have spread south as their range was affected at higher altitude and latitude. Rice falls out of the diet at sites such as Haimenkou c. 1590 BC to be replaced by cold-hardy Western Eurasian domesticates, such as wheat and barley (Jin et al. Reference Jin, Xu, Rui, Li, Wu and Hein2014).
The substantial decrease in the thermal niche for tropical rice following the second millennium BC on the Yunnan-Guizhou Plateau may have contributed to the intensification of trade networks or influenced farmers to migrate south in search of more suitable environments to grow their crops. The timing of this climatic downturn may explain the intensified connections between Yunnan and Southeast Asia, and possibly also the spread of people, rice and tin-bronze technology. This was not, however, the only factor that contributed to the spread of crops into Southeast Asia. Millet, for example, spread earlier in ways unrelated to climatic factors, and the initial spread of rice by 2000 cal BC to sites such as Rach Nui in southern Vietnam does not appear to have been precipitated in this way (Castillo et al. Reference Castillo, Higham, Miller, Chang, Douka, Higham and Fuller2018b), perhaps indicating the existence of coastal routes into Southeast Asia.
Conclusion
In this article, we have documented an early arrival of millet into the Khao Wong Prachan Valley of central Thailand (Weber et al. Reference Weber, Lehman, Barela, Hawks and Harriman2010). The presence of millet, initially without rice, suggests that it may have formed part of an early transmission of domesticates via the Eastern Himalayas through the Yunnan-Guizhou Plateau highlands. Over the 3000 years of occupation recorded at the sites examined here, rice replaced millet as the dominant crop, but, up to and including the historic Dvaravati period, there is no evidence for intensifying rice production through ploughing or irrigation. Rather, dryland, rain-fed cultivation dominated, representing a continuation of local forager subsistence practices. This contrasts sharply with the sequence at Khok Phanom Di, where a coastal saline environment generally militated against rice cultivation (Thompson Reference Thompson1992). It also contrasts with the sequences in the Upper Mun Valley occupation sites of Ban Non Wat, Noen U-Loke and Non Ban Jak, which have not yielded a single grain of millet, despite intensive flotation. There, rice dominated from initial settlement in the seventeenth century BC until the end of the prehistoric period. Moreover, a sharp increase in aridity from c. AD 200 stimulated an agricultural revolution involving the construction of reservoirs and the reticulation of water into permanent, ploughed rice fields (Wohlfarth et al. Reference Wohlfarth, Higham, Yamoah, Chabangborn, Chawchai and Smittenberg2016; Castillo et al. Reference Castillo, Higham, Miller, Chang, Douka, Higham and Fuller2018b).
Ongoing research in the Khao Wong Prachan Valley, and more generally in Southeast Asia, continues to reveal the story of its human past. The analysis of further data may provide a clearer understanding of the development of the cultivated landscape of Southeast Asia and add detail to the ‘Two Layer Hypothesis’ about the expansion of millet- and rice-farming communities into a region occupied for millennia by foragers.
Acknowledgements
This paper is in loving memory of Steven Weber, an inspirational archaeobotanist and ethnobiologist who passed away during the process of its writing. We thank the Marsden Fund of New Zealand for supporting the Thailand Archaeometallugy Project AMS-dating programme, and the Oxford Radiocarbon Accelerator Unit for dating this material. The Thailand Archaeometallugy Project, co-directed by Vincent C. Pigott and Surapol Natapintu and co-sponsored by the Thai Fine Arts Department and the Penn Museum, has been supported, in part, by the National Science Foundation, the National Geographic Society and the American Philosophical Society.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2020.8.












 Open access
Open access