



Introduction
Compared with the widespread Rhuddanian brachiopod Virgiana Twenhofel, Reference Twenhofel1914 (Virgianidae, Pentamerida), which commonly occurs as regionally traceable shell beds in nearly all carbonate basins of Laurentia, Siberia, and their adjacent terranes (see summary in Jin and Copper, Reference Jin and Copper2000, Reference Jin and Copper2010; Jin et al., Reference Jin, Mikulic and Kluessendorf2019), the contemporaneous Platymerella Foerste, Reference Foerste1909 has a much more limited area of distribution, confined to the southeastern and mid-continental United States (Fig. 1), primarily Tennessee and Illinois (Foerste, Reference Foerste1909, Reference Foerste1920; Savage, Reference Savage1913; Boucot et al., Reference Boucot, Johnson and Rubel1971). Virgiana is one of the first Silurian-aspect brachiopod taxa to occur after the Late Ordovician mass extinction event and has been shown to be a useful biostratigraphic marker for the Rhuddanian Stage (Watkins and Kuglitsch, Reference Watkins and Kuglitsch1997; Jin and Copper, Reference Jin and Copper2000) because of its general abundance, shell-bed-forming tendency within a narrow stratigraphic range, and relatively wide distribution in paleotropical carbonate basins. Platymerella, however, has remained poorly understood partly because of its incompletely known morphological characteristics and relatively rare occurrences. Confusion has also arisen from the early interpretation of a “middle Llandovery” age for Virgiana and Platymerella (Berry and Boucot, Reference Berry and Boucot1970; Ziegler and Boucot, Reference Ziegler, Boucot, Berry and Boucot1970), an age designation that has persisted through many subsequent studies (e.g., Boucot, Reference Boucot1975, p. 134; Sapelnikov, Reference Sapelnikov1985; Witzke and Johnson, Reference Witzke, Johnson, Boucot and Lawson1999, p. 838), including the revised brachiopod volumes of the Treatise (Boucot et al., Reference Boucot, Rong, Blodgett and Kaesler2002). This may have obscured the paleobiogeographic significance of Platymerella as compared with its coeval Virgiana of middle–late Rhuddanian age.

Figure 1. Paleogeographic map of Laurentia showing occurrences of Virgiana (black dots) and Platymerella (red dots) during the earliest Silurian (Rhuddanian). Paleogeographic map based on Cocks and Torsvik (Reference Cocks and Torsvik2011); Sebree Trough based on Kolata et al. (Reference Kolata, Huff and Bergström2001). Platymerella localities (see text for details): 1, Trenton, Georgia; 2, Tiftonia, western Tennessee; 3, near Riverside, southwestern Tennessee (type locality of Platymerella manniensis); 4, Monterey School section; 5, National Quarry, Joliet.
Since its establishment, Platymerella has been regarded as a possible synonym of Virgiana by some authors (e.g., Ziegler and Boucot, Reference Ziegler, Boucot, Berry and Boucot1970; Boucot et al., Reference Boucot, Rong, Blodgett and Kaesler2002). Well-preserved specimens of Platymerella gathered for this study, however, demonstrate that it is not only morphologically distinct from Virgiana, but also paleogeographically separate. These observations suggest the validity of two separate genera, which were the result of earliest Silurian virgianid divergence into separate lineages in response to different environmental stressors during the early recovery of benthic marine shelly faunas following the Late Ordovician mass extinction event. The main objectives of this study, therefore, are to elucidate the morphological details of Platymerella, and explore their paleoecological and paleogeographical implications.
Geologic and stratigraphic settings
Southern Appalachian sites
Reported occurrences of Platymerella manniensis Foerste, Reference Foerste1909 in the Appalachian foreland basin of southeastern United States are limited. Previously, it was mentioned as a Georgia occurrence only once, in the “lower division” of the Silurian Red Mountain Formation of Georgia and Alabama (Butts and Gildersleeve, Reference Butts and Gildersleeve1948). Platymerella was not mentioned or illustrated by Allen and Lester (Reference Allen and Lester1954) in their summary of the paleontology of northwest Georgia. Berry and Boucot (Reference Berry and Boucot1970) reported a Platymerella zone (Llandovery) in the Brassfield Limestone of western Tennessee. The shells of Platymerella from the Southern Appalachians described in this paper were collected from two localities in the Lookout Valley on the northwest side of the Appalachian Valley and Ridge province in the vicinity of Chattanooga, Tennessee: the first at Tiftonia, Tennessee (Fig. 2), and the other at Trenton, Georgia. None of the specimens was in situ, but all were directly associated with outcrops of lower Silurian (Rhuddanian) strata assigned to the Red Mountain Formation in Alabama–Georgia and coeval Rockwood Formation in Tennessee (Hayes, Reference Hayes1894; Butts and Gildersleeve, Reference Butts and Gildersleeve1948; Milici and Wedow, Reference Milici and Wedow1977; Chowns, Reference Chowns and Broadhead1996).
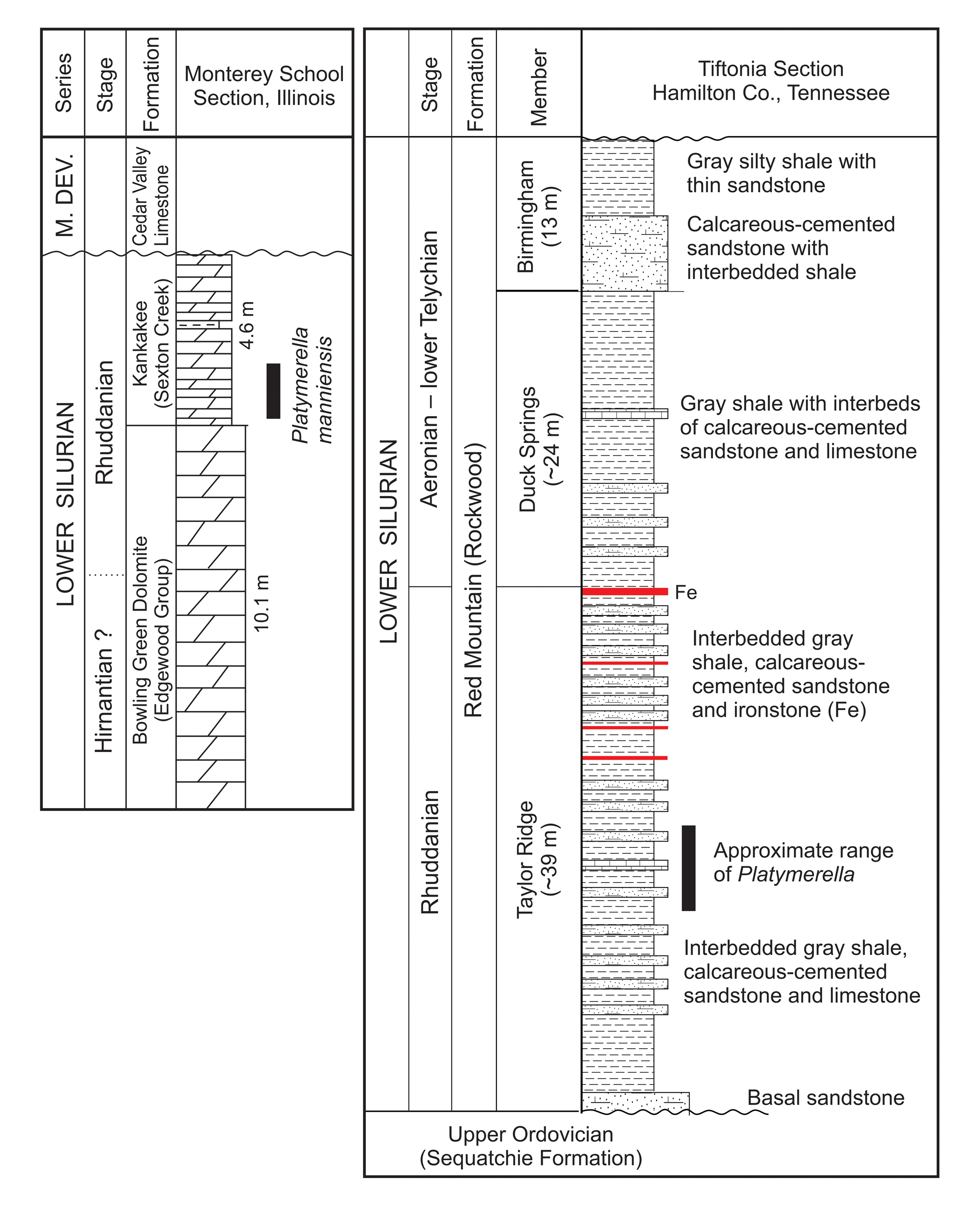
Figure 2. Stratigraphy of Platymerella-bearing Rhuddanian strata in Illinois (left) and Tennessee (right).
The Red Mountain Formation is an unconformity-bounded unit, locally underlain by the Sequatchie Fomation (Upper Ordovician) and overlain by the Chattanooga Shale (Devonian), with the type section at Birmingham, Alabama (Chowns, Reference Chowns and Chowns2006). The formation is presently divided into six members, but only the three lowest members are present around Chattanooga, higher strata having been truncated beneath the Chattanooga Shale. In southeastern outcrop belts within the Valley and Ridge, the Red Mountain Formation consists of a thick sequence of sandstone and shale (up to 360 m near Dalton, Georgia), but in Lookout Valley the section is relatively thin (80 m), dominated by shale and limestone. The formation is best known for the sedimentary iron ores, upon which Birmingham was established, and some thin hematitic limestones are present in the study area. Sedimentological studies indicate deposition on a tropical, storm-dominated shelf subject to flexural subsidence and eustatic changes in sea level driven by glaciation in Gondwana (Chowns and Rindsberg, Reference Chowns, Rindsberg and Holmes2015; Chowns, Reference Chowns2018; Chowns and Ashley, Reference Chowns and Ashley2018).
The Red Mountain Formation in Lookout Valley is divided into three members (Chowns, Reference Chowns and Broadhead1996; Chowns and Rindsberg, Reference Chowns, Rindsberg and Holmes2015), including, in ascending order, the Taylor Ridge (Rhuddanian), Duck Springs (?lower Aeronian), and Birmingham (mid-Aeronian to lower Telychian) members. Age determinations are based on brachiopod lineages investigated by Baarli (in Chowns, Reference Chowns and Broadhead1996) and conodonts (Manzo et al., Reference Manzo, Bergström, Huff and Kolata2002). In particular, the age of the Taylor Ridge Member is established by the Stricklandia lineage, while the base of the Birmingham Member is marked by the occurrence of the S. lens progressa–Pentamerus oblongus Biozone (middle–late Aeronian). According to Johnson (in Chowns, Reference Chowns and Broadhead1996), some specimens of P. oblongus Sowerby, Reference Sowerby and Murchison1839 (earliest Telychian) are transitional to Pentameroides subrectus (Hall and Clarke, Reference Hall and Clarke1893). The specimens from Tiftonia and Trenton described in this paper came from an earlier collection from the Taylor Ridge Member (Rhuddanian), and the precise horizon within the member is therefore unknown.
The Tiftonia section is located on the south side of U.S. 11, just west of the intersection of U.S. 41, Interstate I-24, and exit 174 behind a number of motels and restaurants. The collection was made in the 1990s when the site had been cleared for development, but more construction since has made the site much less accessible and less productive, and some motel properties posted signs against trespassing. At this site, fine transgressive sandstones are observed to lie unconformably over the Upper Ordovician Sequatchie Formation. Above is shale with thin, fine-grained sandstone. At the top of the exposure is a unit of very fossiliferous shale, calcareous dolomitic sandstone, and bioclastic limestone. In addition to three specimens of Platymerella collected at this site, other significant brachiopod taxa include Eoplectodonta sp., Eospirigerina sp., Hesperorthis sp., Levenea sp., and Strophonella sp. (see Chowns and Rindsberg, Reference Chowns, Rindsberg and Holmes2015). Tabulate and solitary rugose corals are common in the uppermost layers of the exposure (Beard and Holmes, Reference Beard and Holmes2010; Landis and Holmes, Reference Landis and Holmes2010).
The Trenton exposure is located on the south side of highway GA-136, approximately 0.8 km west of Highway I-59. It is composed of fine-grained calcareous sandstone and shale capped by a layer of very fossiliferous limestone. A relatively rich fauna includes solitary rugose and tabulate coral, crinoid stems, bryozoans, gastropods, two trilobite genera, and ichnofossils. In addition to six shells of Platymerella collected here, other biostratigaphically useful brachiopods include well-preserved Eospirigerina sp., Eoplectodonta sp., and Levenea sp. (see Chowns and Rindsberg, Reference Chowns, Rindsberg and Holmes2015). Limestone slabs exhibit well-preserved brachiopods in relatively undisturbed positions.
Illinois and surrounding states
Silurian strata are known from broad areas of northeastern and northwestern Illinois, along with more limited areas in the west-central and southern parts of the state. Each of these four regions is more closely related to Silurian rocks in adjacent states than to each other, but collectively they provide critical information on the distribution and character of Platymerella beds.
In northeastern Illinois, beds characterized by common to abundant specimens of Platymerella are known from numerous cores, quarries, and natural exposures over a broad area, including parts of Kankakee, Will, Cook, Du Page, and Kane counties. This interval is a prominent and easily identifiable biostratigraphic marker in the region. One of the best known and longest studied exposure of these beds occurred in the now abandoned and partially filled National Quarry (Vulcan Materials Company) near Joliet in Will County. This quarry had exhibited a 75 m thick section from the top of the Scales Formation of the Maquoketa Group (upper Katian) up to the Racine Dolomite (Wenlock). Platymerella-bearing dolomitic strata occur in the upper (1.5 m) Elwood and the basal Kankakee formations. At this location, the Elwood Formation consists mainly of a sequence of up to 10 m of cherty dolomite that becomes increasingly argillaceous downward, with the maximum thickness exposed at the National Quarry. The basal, more argillaceous part of the formation grades into strata of the underlying and very argillaceous Wilhelmi Formation (12.1 m), whereas the upper part of the Elwood grades up into the overlying clean carbonate of the Drummond Member (2.6 m) of the Kankakee Dolomite. The entire section of the Wilhelmi–Elwood–Drummond strata marks a single major transgressive flooding event with shale at the base of the Wilhelmi gradually grading up into the clean carbonate of the Drummond. The contact of the Wilhelmi with the underlying late Ordovician Maquoketa Formation marks a major regional unconformity with up to 30 m of local relief, and the Wilhelmi contains a prominent positive δ13C isotope excursion equivalent to the Hirnantian excursion (J. Kluessendorf and D. Mikulic, unpublished data). Conodonts in the upper Wilhelmi here are typical members of the Ozarkodina hassi zone (Mikulic et al., Reference Mikulic, Sargent, Norby and Kolata1985). Savage (Reference Savage1913, Reference Savage1916) described a diverse biota dominated by brachiopods, trilobites, and rugose corals from the upper Wilhelmi. The top of the Drummond marked a prominent depositional break with little relief. The overlying Offerman Member of the Kankakee Dolomite is a thin transgressive unit (2.5 m) associated with a positive carbon δ13C isotope excursion in northeastern Illinois, whereas the underlying Platymerella-bearing Elwood strata exhibit a conspicuous negative δ13C excursion (J. Kluessendorf and D. Mikulic, unpublished data).
Abundant conjoined shells and disarticulated valves of Platymerella occur in the top 1.5 m of the Elwood Dolomite without evidence of sorting or directional orientation of valves. The shells are better preserved in chert nodules than in the dolomite matrix. The Drummond Member of the basal Kankakee Formation comprises a 2.6 m thick succession of olive-grey, massive to thick-bedded, tightly cemented dolomite. The lower 1.7 m is cherty and irregularly bedded, whereas the upper 0.9 m is massive, with strong vuggy porosity. The contact between the Elwood and the Drummond is gradational over a meter of strata. The highest chert has been placed in the Drummond but contains no specimens of Platymerella.
In states adjacent to northeastern Illinois, Platymerella has been reported from cores in Kenosha County of Wisconsin. In Kenosha County, a few poorly preserved specimens have been found in the same stratigraphic position as in adjacent Illinois, but they completely disappear a short distance to the north. Northward, no related brachiopods have been observed until prominent virgianid shell beds are found in the same stratigraphic position north of Milwaukee County.
The Monterey School section is located in west-central Illinois exposing a section from the Bowling Green Dolomite (Edgewood Group, Hirnantian–basal Rhuddanian) and the overlying Sexton Creek Formation (middle–upper Rhuddanian; Fig. 2). The Sexton Creek Formation in this area is considered a facies equivalent, at least partially, to the Elwood and Kankakee formations in northeastern Illinois. The Platymerella found at this locality are probably from the transition beds between Bowling Green and Sexton Creek and not truly from the Sexton Creek.
In northwestern and southern Illinois, Platymerella has not been found in early Silurian rocks although similar depositional sequences and related lithologies are present. For example, in northwestern Illinois, the Mosalem and Tete des Morts formations are equivalent in age and show the same basic shale-to-carbonate trend as the Wilhelmi–Drummond sequence in northeastern Illinois, but Platymerella is absent. In southern Illinois, the type section of the Sexton Creek Formation is exposed at the mouth of Sexton Creek, Alexander County, consisting predominantly of dolomite, with minor silty limestone interbeds. Platymerella is not known from the Sexton Creek or any other rocks of this region.
Paleobiogeographic implications
Boucot (Reference Boucot1975, p. 134–135) commented that among the extremely abundant and widespread Virgiana and related forms in North America (e.g., Platymerella), morphological “clines may well exist from one form to another” and speculated that Virgiana and Platymerella may have composed such a continuous species cline.
The extreme abundance and wide geographic distribution of Virgiana, characterized by its shell-bed-forming tendency across most of the earliest Silurian sedimentary basins in Launrentia, “must be seen to be believed” as noted by Boucot (Reference Boucot1975, p. 134). Similar occurrences of Virgiana are also known from North Greenland (part of Laurentia), Siberia, and its adjacent terranes (for summary, see Jin and Copper, Reference Jin and Copper2000; Jin et al., Reference Jin, Mikulic and Kluessendorf2019). The Virgiana fauna represents the initial post-extinction recovery of the pentameride brachiopods (Jin et al., Reference Jin, Copper and Zhan2007; Jin and Copper, Reference Jin and Copper2010). Paleoecologically, the Rhuddanian Virgiana fauna occupied relatively shallow, tropical marine environments, especially in epeiric seas. This is similar to the latest Katian Tcherskidium fauna, although the two faunas belong to separate evolutionary lineages. Paleogeographically, the Katian Tcherskidium virgianid fauna was largely confined to the Late Ordovician Northern Hemisphere (Jin et al., Reference Jin, Blodgett, Harper and Rusmussen2022), whereas the early Silurian Virgiana fauna expanded across the paleoequator into both the northern and southern paleotropics.
In contrast to the wide paleogeographic distribution of Virgiana, Platymerella was confined to the higher southern paleotropics of southern Laurentia (Fig. 1). Available data indicate that Platymerella occurrences were associated with the southern Appalachian basin and the Sebree Trough along the southern margin of Laurentia, which were susceptible to cool-water upwelling during the Late Ordovician (Holland and Patzkowsky, Reference Holland, Patzkowsky, Witzke, Ludvigsen and Day1996; Kolata et al., Reference Kolata, Huff and Bergström2001; Ettensohn, Reference Ettensohn, Finney and Berry2010), a pattern that may have persisted at least into the earliest Silurian. In the Appalachian foreland basin, for example, phosphate-rich beds are common and laterally extensive in Rhuddanian strata, often used for regional stratigraphic correlation, such as the prominent mid-Rhuddanian Artpark Phosphate Bed and the Densmore Creek Phosphate Bed at the Rhuddanian–Aeronian boundary interval (Brett et al., Reference Brett, Baarli, Chowns, Cotter, Driese, Goodman, Johnson, Landing and Johnson1998).
Despite its relatively small shell with a much-reduced ventral umbo and a low biconvexity, Platymerella has been found to live in a vertical, beak-down position in the Sexton Creek (Bowling Green Dolomite) Formation of Illinois, similar to the vertical life positions of Virgiana and the fairly flat-shelled Microcardinalia reported from Anticosti Island (Jin, Reference Jin2008, p. 181, fig. 12I). The morphological characteristics and living positions of Platymerella may reflect its adaptation to subtropical cool-water conditions consistent with the following observations.
During the Late Ordovician and earliest Silurian, there was a general trend of increased shell size toward the equator and northern paleotropics. The largest shells of virgianids are represented by latest Katian Tcherskidium, Proconchidium, and Holorhynchus, predominantly from the Late Ordovician Northern Hemisphere, as well as the late Rhuddanian Virgiana decussata (Whiteaves, Reference Whiteaves1891) from the northern Williston Basin, slightly south of the paleoequator (Cocks and Torsvik, Reference Cocks and Torsvik2011; Jin et al., Reference Jin, Harper, Cocks, McCausland, Rasmussen and Sheehan2013). These warm-water virgianids commonly attained a maximum shell length of ~80 mm (e.g., Nikolaev and Sapelnikov, Reference Nikolaev and Sapelnikov1969; Jin et al., Reference Jin, Caldwell and Norford1993). Virgiana barrandei (Billings, Reference Billings1857) and V. mayvillensis (Savage, Reference Savage1916) from the Anticosti and Michigan basins (at mid-paleotropic to paleosubtropic latitudes) reach a maximum length of only ~50 mm (average of 30 mm and 40 mm, respectively). The smaller shell size of Platymerella (maximum length ~30 mm) from cool-water settings along the southern margin of Laurentia appears to be consistent with this trend. The paleogeographic setting of these virgianids suggests the presence of a morphocline (as suggested by Boucot, Reference Boucot1975) along an ocean temperature gradient (Fig. 1). Other paleoenvironmental conditions, such as water depth and turbulence levels, were not likely significant controlling factors for the virgianid morphocline as virgianid shell beds were usually found in similar depositional settings dominated by severe storms, as suggested by their common association with hummocky cross stratification (e.g., Jin, Reference Jin2008, p. 169, fig. 6; Jin et al., Reference Jin, Mikulic and Kluessendorf2019, p. 642, fig. 3).
In addition to greater shell size, virgianids tended to develop an increasingly thicker, deeper, and more strongly arched ventral valve and concomitantly a thinner and flatter dorsal valve toward the paleoequator and northern paleotropics. This type of heavy and deeply arched ventral valve has been shown to indicate a recumbent life position (Rong et al., Reference Rong, Jin and Zhan2007; Jin et al., Reference Jin, Harper, Cocks, McCausland, Rasmussen and Sheehan2013), partly embedded in the sediment and forming a fixed, stable base for the brachiopod shell. Opening and closing actions were performed by the light, thin, lid-like dorsal valve only, thus maximizing shell movement efficiency while conserving metabolic energy. Such shell design can be interpreted as an adaptation to living in the hurricane-free equatorial belt, where the risk of smothering by hurricane-mobilized mud sediments was minimal. In the higher tropics where hurricane-grade severe storms would have dominated, pentameride brachiopods have been found to prefer a vertical, beak-down life position, maintained by tight clustering of shells. Even relatively flat pentameride shells, such as those of various stricklandioids from the lower Silurian of Anticosti Island, have been found in vertical life position (Jin, Reference Jin2008). This type of life position would have helped the brachiopods shed muddy sediment during storms, as neither valve would serve as a receptacle, and a posterolaterally open commissure would facilitate the flushing of fine sediments from within the shell.
Materials and methods
Materials
This study is based on a total of 19 free, conjoined, calcareous shells from Tennessee, Georgia, and Illinois and numerous silicified specimens embedded in 28 small blocks of white chert samples from Illinois. The calcareous shells from Tiftonia, Tennessee, show the best-preserved pristine shell microstructures. One of these shells is selected for serial sectioning using a Croft parallel grinder and acetate peels, a well-known technique for brachiopod studies. A few silicified specimens from Illinois occur as good internal molds, which are suitable for reconstruction of internal structures via casting. A two-part silicon molding rubber (Easymold silicone putty) was used to make the rubber cast illustrated in this study.
Tiftonia collection
Three conjoined shells, with some degree of dorsoventral compaction, Taylor Ridge Member, lower Red Mountain (lower Rockwood) Formation, Tiftonia (35.0166°N, 85.3823°W), Tennessee.
Trenton collection
Six complete, conjoined shells (not showing any compression), stratigraphically equivalent to the Tiftonia collection, Trenton (34.8675°N, 85.5228°W), Georgia.
Illinois collections
Northeastern outcrop belt: 28 small blocks with silicified shells embedded in white chert matrix, Elwood Formation, National Quarry (Vulcan Materials Company, Joliet; 41.484°N, 88.0938°W), Illinois. Collection by D.M. and Joanne Kluessendorf in 1996.
West-central outcrop belt.—Ten conjoined shells (most showing minor damage anteriorly), basal Sexton Creek Formation (= Bowling Green Dolomite), Quarry above Monterey School (39°1'52.17”N, 90°36'28.45"W) ~4.8 km east of Batchtown, Calhoun County, Illinois.
Repositories and institutional abbreviations
Figured specimens used in this study were housed at Tellus Science Museum (TSM), Cartersville, Georgia, USA; Illinois Geological Survey, University of Illinois (UI), Champaign-Urbana, Illinois, USA; and the Field Museum (FMNH-PE), Chicago, Illinois, USA.
Systematic paleontology
Order Pentamerida Schuchert and Cooper, Reference Schuchert and Cooper1931
Suborder Pentameridina Schuchert and Cooper, Reference Schuchert and Cooper1931
Superfamily Pentameroidea M'Coy, Reference M'Coy1844
Family Virgianidae Boucot and Amsden, Reference Boucot and Amsden1963
Genus Platymerella Foerste, Reference Foerste1909
Type species
Platymerella manniensis Foerste, Reference Foerste1909 (see the following).
Diagnosis (emended)
Shell small to medium sized for virgianids, equibiconvex and nearly equal-sized ventral and dorsal valves; ventral and dorsal umbones rising slightly posterior of hinge line, with ventral umbo slightly higher. Costae weak posteriorly, better developed anteriorly. Fold and sulcus absent, but faint trilobation may be present. Spondylium short, broadly V-shaped, with thickened walls, supported by thick median septum. Inner hinge plates low, thick, dorsomedially inclined, forming pseudocruralium posteriorly; outer hinge plates slightly longer. Configuration of crura similar to that in Virgiana.
Occurrence
Middle–late Rhuddanian, southern Laurentia (southeastern and mid-continental United States: Georgia, Tennessee, Missouri, Ohio, Indiana).
Remarks
In the revised Treatise (Rong and Boucot, Reference Rong and Boucot1998; Boucot et al., Reference Boucot, Rong, Blodgett and Kaesler2002), Platymerella was downgraded to a subgenus of Virgiana because of its similar costae and internal structures. Detailed examination of the internal structures in this study, however, demonstrates that, despite its relatively small and delicate-looking shells, Platymerella has pronounced thickening of the shell wall and internal structures, a morphological trait that resembles more the Late Ordovician virgianids (e.g., Tcherskidium and Proconchidium) than the early Silurian Virgiana. The prominent thickening of hinge plates forms a pseudocruralium, which was described as a “cruralium resting directly upon the bottom of the interior of the valve” (Foerste, Reference Foerste1920, p. 223). Schuchert and Cooper (Reference Schuchert and Cooper1932, p. 184) interpreted the structure as a “pseudocruralium” formed by an “extra testaceous substance” that unites the hinge plates with the valve floor. The pseudocruralium interpretation agrees with the observation in this study based on serial sections—the lamellar layers of the inner hinge plates are dorsomedially inclined but remain discrete, and the cruralium-like structure was formed by the thickened prismatic shell substance. The inner hinge plates in Virgiana, although dorsomedially inclined, do not converge and unite themselves to form a cruralium, nor are there extra-thickened prismatic layers on the inner sides of the hinge plates to merge medially into a pseudocruralium. In addition, Virgiana has a pronounced tendency to become strongly ventribiconvex with a strongly convex ventral valve and a highly arched ventral umbo and beak, which contrasts sharply with the relatively flat, equibiconvex shells with both umbones barely raised above the hinge line. Among the early Silurian virgianids, Virgianoides Jin, Mikulic, and Kluessendorf, 2019 has an incipient but true cruralium supported anteriorly by a low median septum, as shown by Virgianoides major (Savage, Reference Savage1916) from the uppermost Rhuddanian Lime Island Formation of Wisconsin. In its development of a pseudocruralium, Platymerella shows a greater morphological affinity to Virgianoides than to Virgiana, which has basomedially inclined but completely discrete inner hinge plates. In light of its distinct characters, Platymerella is retained as an independent genus in this study (sensu Foerste, Reference Foerste1909; Amsden and Biernat, Reference Amsden, Biernat and Moore1965).
Platymerella manniensis Foerste, Reference Foerste1909
Figures 3–6; Table 1
- Reference Foerste1909
Platymerella manniensis Foerste, p. 70, pl. 1, figs. 1A–D.
- Reference Savage1916
Platymerella manniensis; Savage, p. 324, pl. 16, figs. 11, 12.
- Reference Foerste1920
Platymerella manniensis; Foerste, p. 223, pl. 23, figs. 5A–H.
- Reference Schuchert and Cooper1932
Platymerella manniensis; Schuchert and Cooper, p. 184, pl. 27, figs. 2, 3, 5, 11.
- Reference Amsden, Biernat and Moore1965
Platymerella manniensis; Amsden and Biernat, p. H547, figs. 408-6a–e, 414-6a–d.
- Reference Boucot, Johnson and Rubel1971
Platymerella manniensis; Boucot et al., p. 273, pl. 6, figs. 6–12.
- Reference Sapelnikov1985
Platymerella manniensis; Sapelnikov, p. 34, pl. 6, figs. 4, 5; text-fig. 13.
- Reference Boucot, Rong, Blodgett and Kaesler2002
Virgiana (Platymerella) manniensis (Foerste); Boucot et al., p. 963, fig. 643-2a–j.

Figure 3. Platymerella manniensis Foerste, Reference Foerste1909 specimens from Rhuddanian strata of Illinois. (1–5) FMNH PE 93304: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of subrhomboidal shell, Sexton Creek Formation (= Bowling Green Dolomite), Monterey School Quarry. (6–10) FMNH PE 93305: (6) dorsal, (7) ventral, (8) lateral, (9) posterior, and (10) anterior views of ovoidal shell, same locality. (11, 12) FMNH PE 93307: (11) cherty internal mold and (12) silicon rubble cast of shell posterior, showing thickened spondylium walls, short and low median septum, and hinge plates; specimen from Elwood Formation, National Quarry, northeastern Illinois.

Figure 5. Platymerella manniensis Foerste, Reference Foerste1909, specimens from Rhuddanian strata of Georgia and Tennessee. (1–5) TL2022.7.1: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of mostly abraded and exfoliated shell, Taylor Ridge Member, lower Red Mountain (lower Rockwood) Formation, Trenton, Georgia. (6–10) TL2022.7.3: (6) dorsal, (7) ventral, (8) lateral, (9) posterior, and (10) anterior views of relatively small, suboval shell, same locality in Trenton. (11–15) TL2022.7.7: (11) dorsal, (12) ventral, (13) lateral, (14) posterior, and (15) anterior views of partly abraded, dorsoventrally compressed (preservational) shell, Taylor Ridge Member, lower Red Mountain (lower Rockwood) Formation, Tiftonia, Tennessee.

Figure 6. Platymerella manniensis Foerste, Reference Foerste1909, serial sections of specimen TL2022.7.8, Taylor Ridge Member, lower Red Mountain (lower Rockwood) Formation, Tiftonia, Tennessee. Note thickened median septum, spondylium, and hinge plates, and posteriorly developed pseudocruralium. Number under each image represents linear distance (mm) from shell apex.
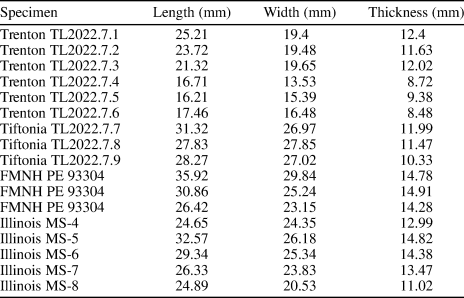
Table 1. Measurements of shell dimensions of Platymerella manniensis from Georgia, Tennessee, and Illinois.
Types
Foerste (Reference Foerste1909, p. 71, pl. 1, fig. 1A–D) illustrated four specimens, all from “northwest of Riverside, Tennessee, Clinton bed.” Among these, the complete shell with an elongate, rhomboidal outline (pl. 1, fig. 1A) matches the specimen figured by Amsden and Biernat (Reference Amsden, Biernat and Moore1965, fig. 414-6c) and Boucot et al. (Reference Boucot, Rong, Blodgett and Kaesler2002, fig. 643-2c) in both editions of the Treatise. This specimen should be regarded as the lectotype (not traced in this study).
In a subsequent description of P. manniensis, Foerste (Reference Foerste1920, p. 223) specified that the species was “originally described from the Brassfield of western Tennessee, and later found by Savage (Reference Savage1916) in the basal part of the Sexton Creek equivalent of the Brassfield in northeastern Missouri and adjacent Illinois.” The Platymerella-bearing strata in Tennessee and Illinois were assigned a “middle Llandovery” age in earlier studies (e.g., Amsden and Biernat, Reference Amsden, Biernat and Moore1965; Berry and Boucot, Reference Berry and Boucot1970), which seems to have persisted into many subsequent studies. In modern stratigraphy, these beds belong to the Taylor Ridge member of the lower Red Mountain (= lower Rockwood) Formation in the border areas of Georgia and eastern Tennessee. In western Tennessee, Ohio, Illinois, and adjacent areas, the Platymerella beds have been used as a stratigraphic marker of the basal Sexton Creek Formation (= lower Brassfield Formation, or lower Kankakee Formation) that overlies the Edgewood Group (Savage, Reference Savage1916; Shaver et al., Reference Shaver, Ault, Burger and Carr1986). The type stratum of Platymerella manniensis, therefore, should be middle–late Rhuddanian in age.
Description (based mainly on new collections of this study)
Shell small to medium sized for virgianids (Table 1), elongated rhomboidal to suboval in outline, equibiconvex with nearly equal-sized, shallow to moderately deep ventral and dorsal valves (Figs. 3–5). Hinge line short, attaining less than one-third of shell width. Ventral and dorsal umbones low, rarely extending more than 2 mm above hinge line, with small, incurved beak, and ventral umbo slightly higher than dorsal, especially in larger shells. Fold and sulcus absent, but weak trilobation may develop in some shells (for example, see Fig. 4.1, 4.2, 4.5, 4.10, 4.11) or gentle medial depression present in anterior part of dorsal valve in other shells. Costae low, rounded, usually faint in posterior part of shell, becoming better defined anteriorly, increasing by bifurcation toward anterior margin of some larger shells (e.g., Fig. 3.6, 3.7). Growth lamellae irregularly developed.
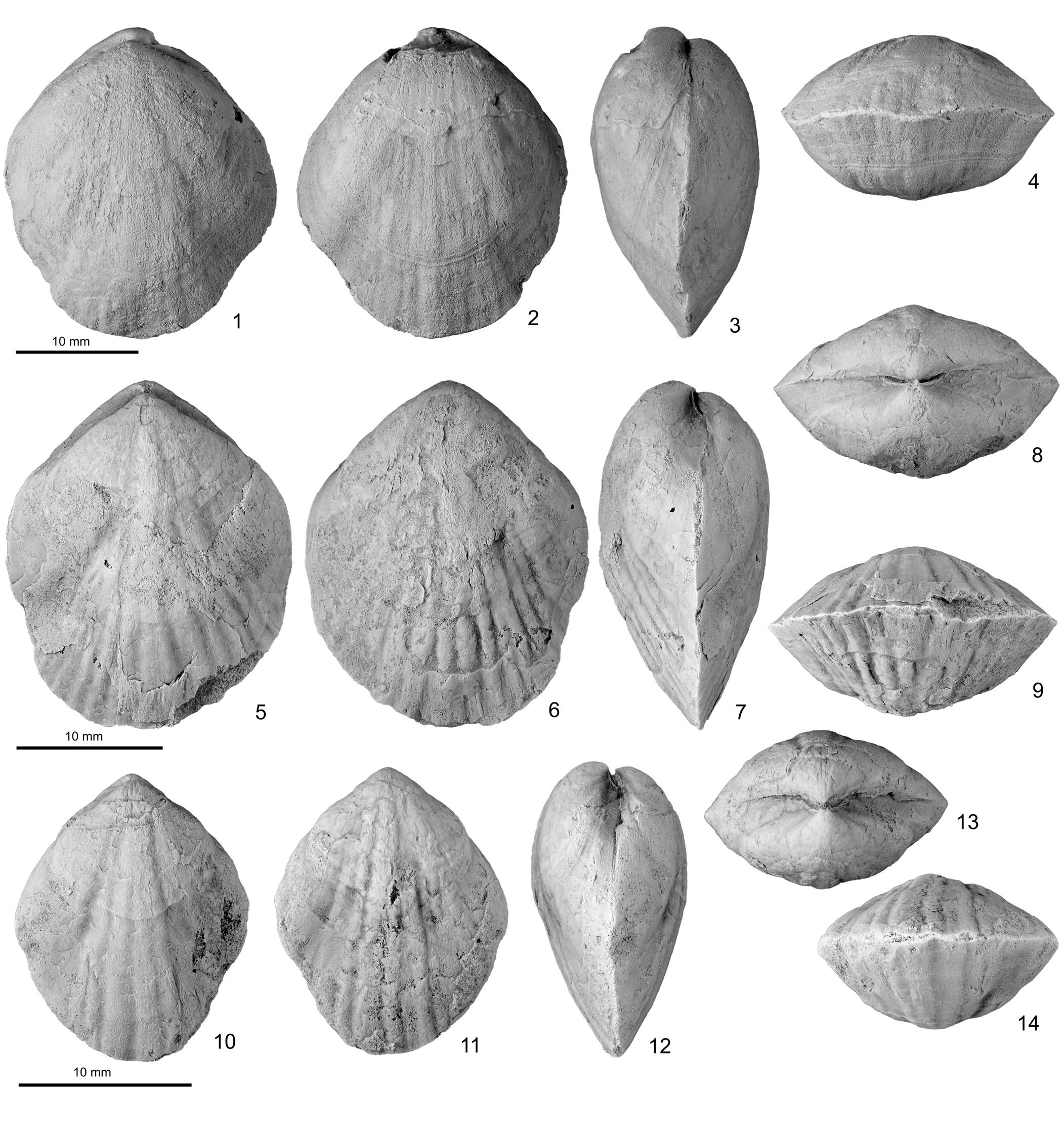
Figure 4. Platymerella manniensis Foerste, Reference Foerste1909, specimens from Rhuddanian strata of Illinois and Georgia. (1–4) FMNH PE 93306: (1) dorsal, (2) ventral, (3) lateral, and (4) anterior views of rhomboidal shell with slightly damaged ventral umbo, Sexton Creek Formation (= Bowling Green Dolomite), Monterey School Quarry. (5–9) TL2022.7.2: (5) dorsal, (6) ventral, (7) lateral, (8) posterior, and (9) anterior views of posteriorly abraded shell, Taylor Ridge Member, lower Red Mountain (lower Rockwood) Formation, Trenton, Georgia. (10–14) TL2022.7.4: (10) dorsal, (11) ventral, (12) lateral, (13) posterior, and (14) anterior views of relatively small, rhomboidal shell, same locality in Trenton.
Spondylium short, broad V-shaped in cross section, extending 2–3 mm anterior of hinge line, with lateral walls thickened by extra prismatic layer (Figs. 3.11, 3.12, 6), bearing striated spondylial floor (Fig. 6; 3.3 mm from apex). Supporting median septum slightly shorter than spondylium, also thickened prominently by extra prismatic layer, particularly in its posterior portion. Tooth knobby, rather small for size of shell (Fig. 6; 1.9 mm and 2.1 mm from apex). Dorsal valve with weakly developed hinge sockets. Hinge plates and crural bases fused smoothly without forming any distinct flanges. Inner hinge plates low, shorter than spondylium, mostly buried in continuous thickening from valve floor to hinge plates, dorsomedially inclined but with discreet lamellar layers; extra thickening of prismatic layer on inner sides of inner hinge plates forming V-shaped pseudocruralium posteriorly (Fig. 6; 1.0–1.5 mm from apex). Outer hinge plates slightly longer and higher than, but similarly thickened as, inner hinge plates. Crural bases elongate oval in cross section, becoming free, laterally compressed rod-like crura anteriorly (Fig. 6; 2.9 mm from apex).
Materials
A total of 19 free shells and numerous others embedded in small blocks (for details, see the Materials and methods section). All from mid–upper Rhuddanian strata, Georgia, Illinois, and Tennessee.
Remarks
The rhomboidal shell outline exhibited in the lectotype does not seem to be a stable character. This form is just a variant among the four specimens originally figured by Foerste (Reference Foerste1909), which also included elongate ovoidal shells. A similar range of variation in shell outline is observed in this study, among collections from the Georgia–Tennessee border area and from Illinois (Figs. 3–5). Secondary thickening, which may fill at least part of the spondylium as revealed in the serial sections of this study (Fig. 6; 1.5–2.9 mm from apex), does not seem to be a consistent feature either. The formation of a pseudocruralium by posterial thickening of the hinge plates was also noted by Foerste (Reference Foerste1920, p. 224, pl. 23, fig. 5E). Schuchert and Cooper (Reference Schuchert and Cooper1932) pondered the taxonomic significance of the pseudocruralium by proposing two distinct forms of Platymerella, one with a pseudocruralium and the other with relatively long, parallel inner hinge plates (called “septal plates” by these authors). Serial sections in this study revealed that the pseudocruralium was formed by thickened prismatic layers of the otherwise discrete inner hinge plates (as indicated by their lamellar layers). In specimens with poorly developed (or poorly preserved) prismatic thickening, the inner hinge plates would appear discrete by default.
Acknowledgments
The specimens from Tennessee and Georgia were collected by J. Santamaria. This study was funded by the Natural Sciences and Engineering Research Council of Canada (J.J.), Tellus Science Museum (J.S.), and the Illinois Geological Survey (D.M.). The constructive comments of C.M.Ø. Rasmussen and an anonymous reviewer greatly helped improve the presentation and clarity of our discussion in this paper.








 Open access
Open access