

Introduction
Many raptor species around the world are declining, and avian scavengers like vultures are among the most threatened raptor species (Thiollay Reference Thiollay2006, Virani et al. Reference Virani, Kendall, Njoroge and Thomsett2011, Chaudhary et al. Reference Chaudhary, Subedi, Giri, Baral, Bidari, Subedi, Chaudhary, Chaudhary, Paudel and Cuthbert2012, Ogada et al. Reference Ogada, Keesing and Virani2012). While for several Asian vulture species declines have been caused by the veterinary drug Diclofenac (Green et al. Reference Green, Newton, Shultz, Cunningham, Gilbert, Pain and Prakash2004, Oaks et al. Reference Oaks, Gilbert, Virani, Watson, Meteyer, Rideout, Shivaprasad, Ahmed, Chaudhry and Arshad2004, Galligan et al. Reference Galligan, Amano, Prakash, Kulkarni, Shringarphure, Prakash, Ranade, Green and Cuthbert2014), the population declines of European vulture populations are frequently the consequence of multiple threats (Ogada et al. Reference Ogada, Keesing and Virani2012, Andevski Reference Andevski2013, Margalida et al. Reference Margalida, Bogliani, Bowden, Donázar, Genero, Gilbert, Karesh, Kock, Lubroth, Manteca, Naidoo, Neimanis, Sánchez-Zapata, Taggart, Vaarten, Yon, Kuiken and Green2014a). After the historic loss of wild large mammal populations (Stuart Reference Stuart1991), a recent prominent threat to European vultures has been sanitary regulations that banned the disposal of livestock carcasses and thus reduced reliable food sources that had been ubiquitously available for centuries (Donázar et al. Reference Donázar, Margalida, Carrete and Sanchez-Zapata2009, Reference Donázar, Cortés-Avizanda and Carrete2010, Margalida et al. Reference Margalida, Donázar, Carrete and Sánchez-Zapata2010). These changes in food abundance have been associated with changes in vulture behaviour (Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010, Margalida et al. Reference Margalida, Campion and Donazar2011a), and demographic parameters (Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012, 2014b), and models have predicted vulture declines from food shortages following sanitary regulations in Spain (Margalida et al. 2011b, Margalida and Colomer Reference Margalida and Colomer2012). However, to what extent and by what mechanism diet changes are responsible for vulture population declines is still poorly understood.
Among the four vulture species that breed in Europe, the Egyptian Vulture Neophron percnopterus is the smallest species with the most precarious conservation status. Egyptian Vulture populations have been declining for decades, and the reasons for population declines are likely a combination of several known threats such as poisoning, electrocution, landscape changes and direct persecution (Carrete et al. Reference Carrete, Grande, Tella, Sánchez-Zapata, Donázar, Díaz-Delgado and Romo2007, Mateo-Tomás and Olea Reference Mateo-Tomás and Olea2010, Velevski et al. Reference Velevski, Nikolov, Hallmann, Dobrev, Sidiropoulos, Saravia, Tsiakiris, Arkumarev, Galanaki, Kominos, Stara, Kret, Grubač, Lisičanec, Kastritis, Vavylis, Topi, Hoxha and Oppel2015). While a few populations in western Europe are currently stable, the Egyptian Vulture population in Eastern Europe has been declining at a rate of ∼ 7% per year for several decades (Velevski et al. Reference Velevski, Nikolov, Hallmann, Dobrev, Sidiropoulos, Saravia, Tsiakiris, Arkumarev, Galanaki, Kominos, Stara, Kret, Grubač, Lisičanec, Kastritis, Vavylis, Topi, Hoxha and Oppel2015). Given the changes in food availability across Europe, a better understanding of the relationships between diet and demographic parameters may provide useful information on the underlying causes of these population declines.
The Egyptian Vulture is an opportunistic scavenger with a wide diet spectrum that includes carcasses and faeces of vertebrates, human refuse, eggs, invertebrates and some slow-moving small land animals that are captured alive (Negro et al. Reference Negro, Grande, Tella, Garrido, Hornero, Donazar, Sanchez-Zapata, Benitez and Barcell2002, Hidalgo et al. Reference Hidalgo, Zabala, Zuberogoitia, Azkona and Castillo2005, Milchev et al. Reference Milchev, Spassov and Popov2012, Sanchis Serra et al. Reference Sanchis Serra, Real Margalef, Morales Pérez, Pérez Ripoll, Tormo Cuñat, Carrión Marco, Pérez Jordá, Ribera Gómez, Bolufer Marqués and Villaverde Bonilla2014). Changes in diet availability and diversity have been associated with territory abandonment and poorer reproductive performance of Egyptian Vultures in Spain (Carrete et al. Reference Carrete, Grande, Tella, Sánchez-Zapata, Donázar, Díaz-Delgado and Romo2007, Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012), but there is little information on the diet composition and the relationship between diet and territory abandonment and productivity in Eastern Europe (Milchev et al. Reference Milchev, Spassov and Popov2012).
Here we examine whether Egyptian Vulture population declines on the Balkan Peninsula may have been associated with diet choices that affected breeding productivity and territory occupancy. We first examined whether diet diversity and composition affected demographic factors contributing to population declines by relating measures of diet diversity to productivity and territory occupancy rates. Because diet changes may have been a consequence of agricultural policies and associated farming practices, we then compared whether Egyptian Vulture diet diversity and composition differed between two countries (Bulgaria and Greece) with a different history of EU regulations: while Greece has been an EU member since 1981, Bulgaria joined the EU only in 2007, and many agricultural policy related changes have happened more recently in Bulgaria than in Greece. In addition to comparing the diet between these two countries, we therefore also examined whether diet diversity and composition changed over time in Bulgaria since the country joined the EU. This work provides the first exhaustive assessment of Egyptian Vulture diet on the Balkan Peninsula and is critical to better understand the underlying causes of population declines and guide future conservation efforts for the species.
Methods
Nest monitoring
We monitored Egyptian Vulture nests in Bulgaria every year between 2006 and 2013 and in Greece between 2010 and 2013. We visited each breeding territory multiple times per breeding season to confirm if the territory was occupied, and to count the number of raised fledglings. A territory was considered occupied if a pair or a single bird was observed with territorial or nest building behaviour at the beginning of the breeding season (March/April). All territories were visited again in May to confirm which of the pairs were incubating, in June and July to register the number of hatched chicks, and in August to confirm the number of fledged juveniles. For the purpose of this analysis we considered only those territories where birds initiated a breeding attempt and where food remains were collected, and we considered productivity as the number of fledglings produced by a pair in a given year. Because pairs can use different nest sites within the same territory in different years, we considered the territory as the unit of replication in all analyses, rather than individual nest sites, to avoid pseudo-replication.
Collection of food remains
We collected nest content samples from 51 different territories (40 in Bulgaria and 11 in Greece) representing 143 breeding attempts between 2006 and 2013. Food remains were collected from nests with recorded breeding events after the end of the breeding season to avoid disturbance of breeding pairs and their chicks. We accessed all nests using climbing equipment and collected all food remains from the nest cavity, which represented the cumulative food remains from a given pair over the preceding breeding season.
Taxonomic analysis of food remains
We used collections of the National Museum of Natural History in Bulgaria to determine the identity of all food remains (bones, scales, feathers, hides or tissue, carcass parts and pellets) at three taxonomic levels: species (n = 1,936 individuals), order (n = 2,008 individuals) or class (n = 2,020 individuals). For each taxonomic group we determined the minimum number of individuals based on skeletal elements following standard guidelines (Bokonyi Reference Bokonyi1970).
Statistical analysis
We first calculated a diet diversity index for each breeding attempt based on the Shannon-Wiener diversity index which has been used in other studies of vulture diet (Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010, Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012, Milchev et al. Reference Milchev, Spassov and Popov2012), and is defined as:
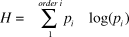 $$H = - \mathop \sum \limits_1^{orderi} p_{i} \times {\rm{log}}(p_{i} )$$
$$H = - \mathop \sum \limits_1^{orderi} p_{i} \times {\rm{log}}(p_{i} )$$
where p i is the proportion that taxonomic order i contributed to all the individual remains in a nest (based on the minimum number of individuals). This index was calculated using the function ‘diversity’ in the R package ‘vegan’. Diet composition was calculated as the proportion of individuals belonging to a taxonomic order divided by the minimum number of individuals from all remains collected in a nest. In addition to classifying animals to a taxonomic order, we also classified the source of prey items into domestic animals (e.g. domestic cats, dogs, livestock etc.) and wildlife (e.g. tortoises, wild cats, wolves, foxes, hedgehogs, etc.) following Hidalgo et al. (Reference Hidalgo, Zabala, Zuberogoitia, Azkona and Castillo2005).
Relationship between diet and productivity
To examine whether diet diversity or composition explained variability in Egyptian Vulture productivity, we used a generalised linear mixed model (GLMM) with the number of fledglings as a response variable and a Poisson error distribution (Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012). Because we collected food remains from some territories in several years, and productivity of these breeding attempts may not have been independent, we accounted for serial correlation within territories by including the identity of the territory as random intercept in the model (Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009). For each analysis we constructed two models: one containing diet diversity, or the proportional contribution of a given diet category as fixed effect, and a corresponding model without the respective effect, but with the same random intercept. We then compared the two corresponding models using a likelihood-ratio test (Lewis et al. Reference Lewis, Butler and Gilbert2011), and concluded that an effect was significant if P < 0.05. We fitted all models using the Laplace approximation in R 3.1.1 (R Development Core Team 2014) with the package ‘lme4’ using the following generic formula: glmer(productivity ∼ diet + (1| territory), family = poisson).
Relationship between diet and territory occupancy
Food availability may not only affect productivity, but also adult condition, survival, and territory retention (Carrete et al. Reference Carrete, Grande, Tella, Sánchez-Zapata, Donázar, Díaz-Delgado and Romo2007, Grande et al. Reference Grande, Serrano, Tavecchia, Carrete, Ceballos, Tella and Donázar2009), and diet composition may therefore affect how regularly a territory is occupied. We therefore examined whether the average diet diversity or composition recorded in a territory could explain the proportion of years during which a territory was occupied, termed the territory occupancy rate (Sergio and Newton Reference Sergio and Newton2003). For this analysis, we only used territories that had been monitored for at least five years, and estimated the effect of diet diversity or composition in a binomial GLM with the number of monitored years as the number of binomial trials in which a territory could be occupied using the following generic formula: glm(cbind(years_occupied,years_unoccupied) ∼ diet, family=binomial). We used an information-theoretic approach for inference by using AIC c to evaluate which of nine biologically plausible candidate models received the most support from our data (Burnham and Anderson Reference Burnham and Anderson2002). The 10 candidate models tested whether occupancy rate increased (1) with the proportion of livestock (cows, sheep, goats, pigs) in the diet; (2) with diet diversity; (3) with the proportion of all of the four most important taxonomic orders in the diet (livestock, chickens, tortoises and carnivores); (4) at intermediate diet diversity (e.g. a quadratic effect of diversity); (5–8) at intermediate diet diversity and with the proportion of one of the four most important taxonomic orders in the diet; and (9) at intermediate diet diversity and with the proportion of all wild animals in the diet. We present the mean parameter estimates with standard errors of the most parsimonious models (ΔAIC c < 10).
Geographic differences in diet
Because Bulgaria and Greece joined the EU at different times, we hypothesised that diet diversity and composition may vary between these two countries. In addition, most Egyptian Vulture nests in Bulgaria and Greece can be grouped into discrete population clusters, of which we sampled one in northern Bulgaria, and one in the eastern Rhodope mountains of southern Bulgaria and eastern Greece (Velevski et al. Reference Velevski, Nikolov, Hallmann, Dobrev, Sidiropoulos, Saravia, Tsiakiris, Arkumarev, Galanaki, Kominos, Stara, Kret, Grubač, Lisičanec, Kastritis, Vavylis, Topi, Hoxha and Oppel2015). Habitat and livestock husbandry practices vary between these regions, and we therefore examined whether diet diversity and composition varied at the national or, alternatively, at the regional level. We used the same GLMM as described above for productivity with two separate two-level categorical variables to test for significant differences in diet diversity between countries or regions, respectively. To avoid this geographic comparison being confounded by variation in sampling intensity over time, we used only nest remains collected after 2009 for this analysis, as no nest remains were collected in Greece before 2010.
Diet changes over time
EU agricultural policies have transformed landscapes in Europe with widespread consequences for bird populations (Donald et al. Reference Donald, Sanderson, Burfield and van Bommel2006, Sanderson et al. Reference Sanderson, Kloch, Sachanowicz and Donald2009). We therefore examined whether Egyptian Vulture diet diversity and composition in Bulgaria changed over time since the country joined the EU in 2007. We tested for a continuous year effect in GLMMs with diet diversity index or the proportional contribution of important prey taxa as response variables in an identical framework as described above for productivity.
Results
We collected a total of 3,237 food remains that represented at least 1,938 individuals of 137 species from 32 orders and 8 classes (Table 1). For both countries combined, mammals represented the highest proportion in the diet (48.4% of individuals), followed by reptiles (28.7%) and birds (21.5%). Although mammals as a group comprised the largest proportion of the diet, the most common species in the Egyptian Vulture’s diet were the Greek tortoise Testudo graeca (13.2%), Herman‘s tortoise Eurotestudo hermanni (11.4%), and the domestic chicken Gallus gallus domestica (7.1%). Wild animals contributed approximately 70% to the diet of Egyptian Vultures in Bulgaria and 71% in Greece.
Table 1. Relative frequency (in %, ± SD) of the most important prey categories in the Egyptian Vulture diet assessed from collecting food remains in 143 breeding attempts from Bulgaria and Greece between 2006 and 2013.

Relationship between diet and productivity
There was no significant relationship between diet diversity and productivity (n = 135 breeding attempts; χ2 = 0.40, P = 0.52; Figure 1). Although territories with a higher diet diversity appeared to raise more fledglings, the uncertainty around this effect (b = 0.12, 95% confidence interval -0.24 – 0.47) was very large (Figure 1). We also found no relationship between the number of fledglings produced by Egyptian Vultures and the overall proportion of wild animals (χ2 = 0.28, P = 0.59), or the proportion of livestock (χ2 = 1.32, P = 0.25), chicken (χ2 = 0.26, P = 0.60), carnivores (χ2 = 0.18, P = 0.67), or tortoises (χ2 = 0.76, P = 0.38) in the diet. A multivariate analysis testing the combined effects of various taxonomic orders did not indicate that diet composition explained any variability in productivity.

Figure 1. Mean (± SD) productivity in 45 Egyptian Vulture territories in Bulgaria and Greece in relation to the average diet diversity calculated from food remains collected at nests in these territories between 2006 and 2013. Lines are mean fitted response (solid) and 95% confidence interval (dashed) of a GLMM.
Relationship between diet and territory occupancy
Of our 10 multivariate candidate models used to explain the occupancy rate of 47 Egyptian Vulture territories with diet variables, those models that assumed that occupancy rate would be highest at intermediate diet diversity received overwhelming support from the data (cumulative ωAIC c = 1; Table 2, Figure 2). The most parsimonious model indicated that in addition to the quadratic effect of diet diversity, occupancy rate was lower with a higher proportion of wild animals in the diet (b = - 3.60 ± 0.97; P < 0.001). A similar model that assumed that occupancy rate increased with a higher proportion of livestock in the diet also received some support from the data (ωAIC c = 0.14; b = 4.35 ± 1.37; P = 0.002).
Table 2. Model selection summary of nine candidate models explaining the occupancy rate of Egyptian Vulture territories in Bulgaria and Greece between 2006 and 2013 with measurements of diet diversity and composition.


Figure 2. Occupancy rate of 47 territories in Bulgaria and Greece in relation to the average diet diversity measured in those territories between 2006 and 2013. Lines are mean fitted response (solid) and 95% confidence interval (dashed) of the most parsimonious candidate model (Table 2), using the mean proportion of wild animals in the diet (73%).
Geographic differences in diet
To compare diet diversity and composition between countries or regions, we only used data collected after 2009 to avoid results being confounded by temporal changes in Bulgaria. Diet diversity did not differ between countries (n = 73; χ2 = 2.42, P = 0.12), but was significantly higher in northern Bulgaria (H = 1.68, n = 10) than in southern Bulgaria and Greece (H = 1.34, n = 63; χ2 = 4.92, P = 0.03). This regional difference was due to a much lower proportion of tortoises in the diet in northern Bulgaria (12.6%) than in southern Bulgaria and Greece (29.8%; χ2 = 4.73, P = 0.03), which appeared to be offset by a marginally higher proportion of carnivores and chickens in northern Bulgaria (Table 1). The proportion of livestock was variable and did not differ between countries or regions (χ2 = 2.39, P = 0.30; Table 1).
Diet changes over time
The diversity of Egyptian Vulture diet did not change significantly in Bulgaria between 2006 and 2013 (b = -0.032 ± 0.017; χ2 = 3.69, P = 0.06, n = 123 breeding attempts). However, we found a small decrease of 1.6% per year in the proportion of tortoises (χ2 = 4.52, P = 0.03, Figure 3), and an increase of 1.3% per year in the proportion of livestock (χ2 = 5.38, P = 0.02, Figure 3). There was no distinct trend in the proportion of carnivores (χ2 = 2.34, P = 0.13), the proportion of chicken (χ2 = 3.01, P = 0.08, Figure 3), or the proportion of all wild animals in the diet over time (χ2 = 1.08, P = 0.30).

Figure 3. Proportion of the four most common taxonomic orders in the diet of Egyptian Vulture nests in Bulgaria calculated from nest remains collected between 2006 and 2013 (n = 123 nests). Lines are mean fitted response (solid) and 95% confidence interval (dashed) of a GLMM with year as a continuous predictor variable.
Discussion
This study provides the first quantitative assessment of Egyptian Vulture diet on the Balkan Peninsula, and demonstrates no direct correspondence between diet diversity and productivity. This result is in contrast to previous assessments of vultures, which generally found higher productivity with a more diverse diet (Margalida et al. Reference Margalida, Bertran and Heredia2009, Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012). Two hypotheses exist that link individual breeding performance with diet breadth (Whitfield et al. Reference Whitfield, Reid, Haworth, Madders, Marquiss, Tingay and Fielding2009): the diet specificity hypothesis predicts that species whose preferred diet decreases in availability will be forced to broaden their diet and suffer lower productivity (Katzner et al. Reference Katzner, Bragin, Knick and Smith2005); conversely, the prey availability hypothesis suggests that a broader diet reflects a higher availability of prey and thus leads to an increase in productivity (Steenhof and Kochert Reference Steenhof and Kochert1988, Arroyo and Garcia Reference Arroyo and Garcia2006, Whitfield et al. Reference Whitfield, Reid, Haworth, Madders, Marquiss, Tingay and Fielding2009). Our study indicates that the productivity of Egyptian Vultures in Bulgaria and Greece is not closely linked to the overall diversity or the proportion of certain prey taxa in the diet. Because we found no effect of diet on productivity, and productivity had a relatively small effect on population viability in theoretical models for the species (García-Ripollés and López-López Reference García-Ripollés and López-López2011, Velevski et al. Reference Velevski, Grubac and Tomovic2014), we conclude that it is unlikely that diet limitations on reproductive output are a critical threat to Egyptian Vultures on the Balkan Peninsula.
In long-lived species like Egyptian Vultures, population dynamics are more sensitive to adult survival than to productivity (Saether and Bakke Reference Saether and Bakke2000, Oro et al. Reference Oro, Margalida, Carrete, Heredia and Donázar2008, Grande et al. Reference Grande, Serrano, Tavecchia, Carrete, Ceballos, Tella and Donázar2009), and understanding the effects of diet limitations on adult survival is therefore critical (Oro et al. Reference Oro, Margalida, Carrete, Heredia and Donázar2008, García-Ripollés and López-López Reference García-Ripollés and López-López2011, Margalida et al. 2014b). In contrast to productivity, we found a hump-shaped relationship between diet diversity and territory occupancy rate. For Egyptian Vultures a lower occupancy rate may indicate low adult survival either due to a poor diet or the risk to consume poisoned carcasses (Carrete et al. Reference Carrete, Grande, Tella, Sánchez-Zapata, Donázar, Díaz-Delgado and Romo2007, Grande et al. Reference Grande, Serrano, Tavecchia, Carrete, Ceballos, Tella and Donázar2009, Hernández and Margalida Reference Hernández and Margalida2009). The hump-shaped relationship between occupancy rate and diet diversity, plus the additive effects of either the proportion of wild animals or livestock in the diet therefore appear to conform with elements of both the diet specificity and prey availability hypotheses at the extreme ends of the diet diversity spectrum, and are in contrast with the apparent positive effect of a more diverse diet found in Spain (Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012). Although our study is correlational and therefore unable to resolve whether diet diversity is a cause or a consequence of lower territory occupancy, a potential mechanism of diet-mediated territory abandonment is that lower food availability may force individuals to consume either a wider variety of animals or just a very narrow, locally easily available range of animals. Such diet restrictions could increase the risk of consuming poisoned bait or carnivore carcasses if found in the landscape, leading to mortality from the accidental ingestion of poison. Carcasses of typical livestock predators such as foxes, wolves, bears, or dogs are frequently the consequence of the illegal but ubiquitous use of poison by many livestock owners and hunters in Greece to protect their livestock, cultivations, or game animals (Mazaris et al. Reference Mazaris, Mamakis, Kalpakis, Poulopoulos and Matsinos2008, Kalpakis et al. Reference Kalpakis, Mazaris, Mamakis and Poulopoulos2009, Skartsi et al. Reference Skartsi, Dobrev, Oppel, Kafetzis, Kret, Karampatsa, Saravia, Bounas, Vavylis, Sidiropoulos, Arkumarev, Dyulgerova and Nikolov2014). Poisoning is known as a significant cause of mortality in Spain (Cortés-Avizanda et al. Reference Cortés-Avizanda, Ceballos and Donázar2009, Hernández and Margalida Reference Hernández and Margalida2009, Mateo-Tomás et al. Reference Mateo-Tomás, Olea and Fombellida2010), and 13 presumably poisoned Egyptian Vultures were found in Greece between 2002 and 2013 (HOS unpubl. data). The risk to consume a poisoned carcass may be especially high in areas where Egyptian Vultures consume a large proportion of wild animals or where relatively few livestock are available, which may explain the relationships we found between occupancy rate and the proportion of these two taxonomic groups in the diet.
Providing safe food for breeding vultures in regions with high risk of poisoning incidents may increase survival of adult and juvenile birds by reducing the risk of ingesting poisoned carcasses (Oro et al. Reference Oro, Margalida, Carrete, Heredia and Donázar2008). If provision of unlimited safe diet was considered to reduce mortality of juvenile and adult birds, this management had a large positive effect on Egyptian Vulture populations in theoretical models (García-Ripollés and López-López Reference García-Ripollés and López-López2011). Territories in which we found a very low or very high diet diversity may be priority candidates for supplementary feeding to avoid poisoning incidents. Although supplementary feeding may reduce mortality risk and slow population declines, this approach can risk conserving qualitatively inferior territories and individuals (García-Heras et al. Reference García-Heras, Cortés-Avizanda and Donázar2013, Rooney et al. Reference Rooney, Reid and Montgomery2015), or reducing productivity of the general population by increasing the interference with floaters attracted to feeding stations (Carrete et al. Reference Carrete, Donazar and Margalida2006). A more thorough evaluation of the long-term consequences of this management technique on productivity and adult survival would be useful.
Despite ongoing population declines (Velevski et al. Reference Velevski, Nikolov, Hallmann, Dobrev, Sidiropoulos, Saravia, Tsiakiris, Arkumarev, Galanaki, Kominos, Stara, Kret, Grubač, Lisičanec, Kastritis, Vavylis, Topi, Hoxha and Oppel2015), we found no temporal trends in Bulgaria that suggested that population declines were associated with a decrease in diet diversity. There was a slight decline in the proportion of tortoises in the Egyptian Vulture diet in Bulgaria, which may be related to population declines of tortoises (Petrov Reference Petrov, Fet and Popov2007, Zivkov et al. Reference Zivkov, Ivanchev, Raikova-Petrova and Trichkova2007). In contrast, the proportion of livestock in the Egyptian Vulture diet increased since Bulgaria joined the EU, although agricultural intensification was expected to lead to a decrease in small livestock farms and the overall availability of livestock carcasses. The apparent increase in livestock consumption may have been a consequence of drastic declines in the number of livestock herds after the collapse of socialist cooperative farming structures in the 1990s (Robinson and Milner-Gulland Reference Robinson and Milner-Gulland2003, Kamp et al. Reference Kamp, Urazaliev, Donald and Hölzel2011). Following this reduction in livestock, EU subsidies to livestock farmers likely resulted in an increase of livestock in several rural areas of Bulgaria in recent years, including some of the areas inhabited by Egyptian Vultures (e.g. eastern Rhodopes).
Mammals were the most common group of vertebrates in the Egyptian Vulture’s diet in our and in previous studies (Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010, Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012, Milchev et al. Reference Milchev, Spassov and Popov2012). However, we found a much higher proportion of reptiles than other studies, mostly as a consequence of the dominance of land tortoises. Tortoises were most common in the diet in southern Bulgaria and eastern Greece, which is likely a consequence of regional differences in availability (Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012), because tortoises are abundant in the dry and warm hills of that region, but considerably scarcer in northern Bulgaria (Petrov Reference Petrov, Fet and Popov2007). These geographic patterns therefore provide further support that Egyptian Vultures are opportunistic foragers whose diet is closely linked to local availability of various prey animals (Hidalgo et al. Reference Hidalgo, Zabala, Zuberogoitia, Azkona and Castillo2005, Milchev et al. Reference Milchev, Spassov and Popov2012).
The overall prominence of tortoises in the Egyptian Vulture diet must be interpreted with caution, because we used prey remains in nests as an indication of the diet consumed by Egyptian Vultures. Indices based on prey remains may be biased because soft tissues that can be consumed and digested without leaving any remains will not be detected by this approach (Simmons et al. Reference Simmons, Avery and Avery1991, Redpath et al. Reference Redpath, Clarke, Madders and Thirgood2001, Margalida et al. Reference Margalida, Bertran and Boudet2005). Thus, the approach that we adopted may overestimate animals with large hard body parts (e.g. tortoises), and underestimate the contribution of animals which may not leave any remains in the nest. In addition, our approach relies on the number of individuals to estimate diet diversity, but arguably the contribution of individuals from species of different size is not equal. Despite these limitations, our approach allows valid comparisons across space and time, and between successful and unsuccessful breeding attempts (Margalida et al. Reference Margalida, Benitez, Sanchez-Zapata, Ávila, Arenas and Donázar2012). The majority of our conclusions are therefore not affected by uncertainties about the precise contribution of various animal groups to the diet of Egyptian Vultures.
In summary, Egyptian Vultures have an extremely variable diet, and our work did not provide any evidence that diet diversity or the proportional contribution of a certain taxonomic order affects reproductive output. However, further research is urgently needed to better understand the links between diet diversity, food availability and territory occupancy, which will require studies relating food availability and consumption to the susceptibility to poisoning and ultimately to survival probabilities of adult and juvenile birds.
Acknowledgements
We acknowledge the support of field assistants helping to monitor and access nests and collect food remains, in particular Ivaylo Angelov, Tsvetomira Angelova, Nikolai Terziev, Sanie Mumun, Vanya Angelova, Hristo Hristov, Marin Kurtev, Dimitar Ragyov, Yanina Klimentova-Nikolova, Ivayla Klimentova, Sylvia Dyulgerova, Emil Yordanov, Ivaylo Ivanov, Angelos Evangelidis, Giannis Chondros, Giorgos Koukis, Vasilis Sideris, Antonios Venetakis and Archontis Exakoidis. Thanks to Georgi Popgeorgiev, Nikolay Tzankov and Andrey Stoyanov for the identification of some of the reptilian remains. This paper was initiated by the LIFE+ project “The Return of the Neophron” (LIFE10 NAT/BG/000152, www.LifeNeophron.eu) funded by the European Union and co-funded by the A. G. Leventis Foundation. Theodora Skartsi, Antoni Margalida and two anonymous reviewers provided constructive comments on an earlier draft of this paper.





