


Introduction
Plesiosaurs are a group of Mesozoic tetrapods secondarily adapted to an aquatic lifestyle. Two plesiosaur taxa survived into the Maastrichtian: the Elasmosauridae and Polycotylidae. These animals preyed on fish and squid (Zammit et al., Reference Zammit, Daniels and Kear2008; Noè et al., Reference Noè, Taylor and Gómez-Pérez2017), but likely with different hunting techniques given their morphology. Most of the elasmosaurs had a characteristically long neck, in extreme cases consisting of up to 72 cervical vertebrae (O’Keefe & Hiller, Reference O’Keefe and Hiller2006), whereas Polycotylidae had much shorter necks (Benson & Druckenmiller, Reference Benson and Druckenmiller2014).
The fossil record of plesiosaurs in the Maastrichtian type area (southeast Netherlands and northeast Belgium; Fig. 1) is limited to extremely rare occurrences of elasmosaurs, in striking contrast to the much richer mosasaur fossil record (e.g. Mulder et al., Reference Mulder, Bardet, Godefroit and Jagt2000). In part, this may reflect habitat preferences (e.g. Schulp et al., Reference Schulp, Janssen, Van Baal, Jagt, Mulder and Vonhof2016), but also when considered over a wider geographical area, the rather meagre record of latest Cretaceous elasmosaurs across the entire North Atlantic realm is striking.
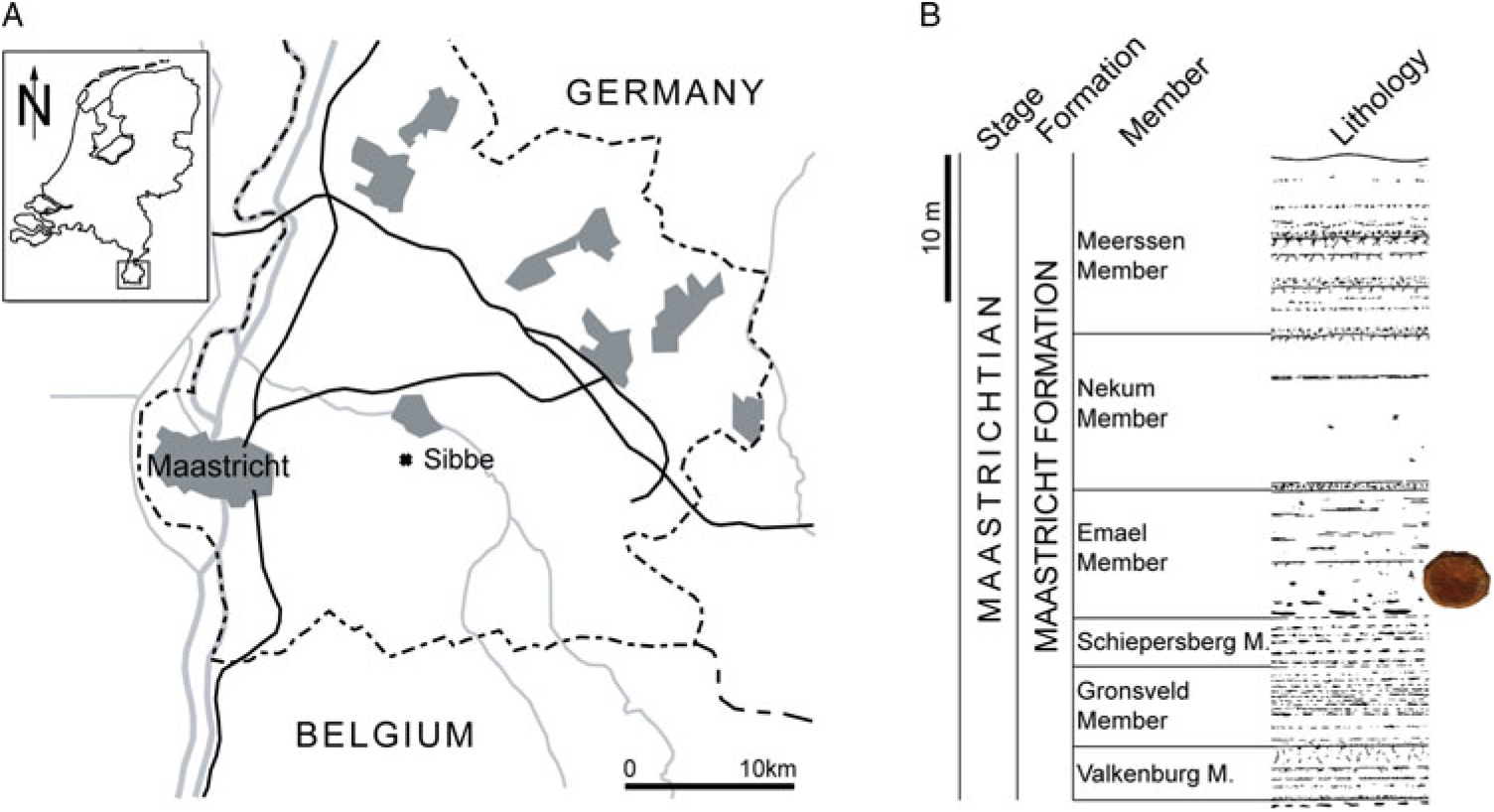
Figure 1. Map of southern Limburg (the Netherlands) and contiguous areas of Belgium and Germany, with indication of the provenance area (Sibbe, municipality of Valkenburg aan de Geul) (A); stratigraphic provenance of NHMM 2014 027 (B).
To date, just seven isolated elasmosaur vertebrae have been recorded from the Maastrichtian type area, in addition to a handful of teeth. This is an extremely low number, considering the more than 250 years of intensive collection efforts in the region (Mulder et al., Reference Mulder, Bardet, Godefroit and Jagt2000). These specimens comprise the generically and specifically indeterminate dorsal and caudal vertebrae that are currently registered as NHMM 1985 141 and NHMM K 1904, as well as three anterior cervical vertebrae (NHMM/MND KB 50.20.01, NHMM/MND K 20.01.801a and NHMM/MND K 20.01.801b) and the dorsal and cervical centra IRScNB 204 and IRScNB 205.
All of these specimens can only be identified to the family level, as Elasmosauridae gen. et sp. indet. Additional material from the Maastricht area, referred to in the literature as Plesiosaurus, was destroyed during World War II (Mulder et al., Reference Mulder, Bardet, Godefroit and Jagt2000). In view of the paucity of elasmosaurs in the fossil record of the Maastrichtian type area, a short note on a well-preserved, newly collected caudal vertebra appears warranted.
Material and methods
The single specimen of a vertebral centrum of a plesiosaur was recovered at Sibbe (municipality of Valkenburg aan de Geul, the Netherlands), by a staff member of Mergelbouwsteen Kleijnen, a building stone company that works a subterranean quarry at Sibbe (Fig. 1). The specimen was kindly donated by this firm to be preserved in the collections of the Natuurhistorisch Museum Maastricht where it now bears registration number NHMM 2014 027.
Institutional abbreviations – IRScNB: Institut royal des Sciences naturelles de Belgique, Brussels, Belgium; NHMM: Natuurhistorisch Museum Maastricht, Maastricht, the Netherlands; NHMM/MND: Museum Natura Docet material, now housed at Natuurhistorisch Museum Maastricht.
Anatomical abbreviations – np, notochord pit; nf, nutritive foramina; pa, parapophyseal facet; pf, pedicular facet.
Geological setting and stratigraphy – The specimen originates from upper Maastrichtian strata (Jagt & Jagt-Yazykova, Reference Jagt, Jagt-Yazykova, Jagt, Donovan and Jagt-Yazykova2012), assigned to the Maastricht Formation, and, in all likelihood, from the Emael Member, as quarrying at Sibbe occurs mostly in that particular unit (Fig. 1). The biocalcarenitic strata of the Maastricht Formation were deposited in a shallow-marine shelf environment (Jagt & Jagt-Yazykova, Reference Jagt, Jagt-Yazykova, Jagt, Donovan and Jagt-Yazykova2012). The Maastricht Formation as exposed at the nearby ENCI-HeidelbergCement Group quarry has been shown to comprise a number of sea-level cycles and a gradually shallowing upward trend, with the exception of the highest metres of the Meerssen Member (Schiøler et al., Reference Schiøler, Brinkhuis, Roncaglia and Wilson1997).
Systematic palaeontology –
SAUROPTERYGIA Owen, Reference Owen1860
PLESIOSAURIA de Blainville, Reference de Blainville1835
PLESIOSAUROIDEA Welles, Reference Welles1943
XENOPSARIA Benson and Druckenmiller, Reference Benson and Druckenmiller2014
Xenopsaria gen. et sp. indet.
Description
The specimen described here, NHMM 2014 027, is an isolated, platycoelous vertebral centrum, measuring 38 mm in dorsoventral height, 28 mm in antero-posterior length and 45 mm in width. Rugose ridges on both articular surfaces radiate outwards to the margins of the centrum. On both surfaces, the notochord pit is well visible as an indentation (Fig. 2) and a bulge overhangs the notochord pits (np) on either side of the centrum. On the dorsal side, two pedicular facets (pf) are seen; these are the joints for the neural spine (O’Gorman et al., Reference O’Gorman, Gasparini and Salgado2014). Dorsally there are three large nutritive foramina. Such have recently been reinterpreted as blood vessels that went through the centrum (Wintrich et al., Reference Wintrich, Scaal and Sander2017). Two of these are connected at a deeper-lying level within the vertebral centrum, but are divided by a shaft at the surface. The other foramen is circular in shape (Fig. 2). Two other foramina, also of large size, are located ventrally (vf), which is a characteristic feature of elasmosaur centra (Wintrich et al., Reference Wintrich, Scaal and Sander2017). These foramina are often referred to as ‘foramina subcentralia’ (e.g. Wintrich et al., Reference Wintrich, Scaal and Sander2017). They are so close to each other that their margins nearly coalesce (Fig. 2). The ventral side lacks the distinctive bell-shaped outline that is seen in cervical centra of some elasmosaurs such as Hydrotherosaurus and Terminonatator and Aristonectinae (Vincent et al., Reference Vincent, Bardet, Houssaye, Amaghzaz and Meslouh2013). Smaller foramina are present on the lateral surfaces, but there is no symmetry in the placement of these foramina on either side (Fig. 2). The lateral surfaces show two small facets on either side; these are quite symmetrical, being central-medial in position.

Figure 2. Xenopsaria gen. et sp. indet. (NHMM 2014 027), in articular (A and B), ventral (C), dorsal (D) and lateral (E and F) views. Abbreviations: np: notochord pit, nf: nutritive foramina, pa: parapophyse facet, pf: pedicular facet.
Discussion
Taxonomic attribution – Plesiosaur vertebral centra are relatively undiagnostic. In the Maastrichtian two different families of plesiosaurs were present: the Elasmosauridae and the Polycotylidae (e.g. Benson & Druckenmiller, Reference Benson and Druckenmiller2014). Both caudal and cervical vertebrae are often scored similarly between the two taxa (e.g. Benson and Druckenmiller, Reference Benson and Druckenmiller2014; Madzia et al., Reference Madzia, Sachs and Lindgren2018). Pliosaurid origin can be ruled out both based on the extensive ghost lineage it would require and the absence of dorsal foramina in the centra in many species of the clade (Benson & Druckenmiller, Reference Benson and Druckenmiller2014; Wintrich et al., Reference Wintrich, Scaal and Sander2017). Furthermore, the ontogenetic stage or ossification state of the specimen limits proper diagnosis.
Position within the axial skeleton – Given the limited number of polycotylid vertebrae described and discussed (see e.g. Novas et al. Reference Novas, D’Angelo, O’Gorman, Agnolín, Lirio and Isasi2015) and no reported polycotylids from the Maastricht formation so far, our discussion on the attribution of position in the axial skeleton has been focused on what has been discussed for the Elasmosauridae. In vertebral centra of elasmosaurs, those features that allow determination of the position within the axial skeleton are subject to both intraspecific and ontogenetic variations (O’Keefe & Hiller, Reference O’Keefe and Hiller2006, pp. 213–216), thus requiring caution in trying to determine the original positions of isolated elements. However, we can certainly rule out a thoracic position of the present vertebra, because it lacks any large facets for rib attachment on the dorsal part of the centrum. Furthermore, the surfaces for neural arch attachment are not as large in NHMM 2014 027 as those seen in most thoracic vertebrae of elasmosaurs, for example in the near-complete Albertonectes vanderveldei (see Kubo et al., Reference Kubo, Mitchell and Henderson2012). Similarities amongst cervical and caudal vertebral centra also preclude easy distinction between the two. However, two key differences are largely valid within the family Elasmosauridae. The parapophyses of cervical centra are placed ventrolaterally, whereas they generally occupy a more central-lateral position in caudal centra (e.g. Otero et al., Reference Otero, Soto-Acuña, Vargas and Rubilar-Rogers2014, p. 321; Sachs & Kear, Reference Sachs and Kear2017). In addition, anterior cervical centra have a clearer dumbbell-shaped outline on the ventral side of the articular surface, although this does not apply to all elasmosaurid cervicals (Vincent et al., Reference Vincent, Bardet, Houssaye, Amaghzaz and Meslouh2013). However, this is never the case in caudal vertebrae; overall, these are more octagonal in cross section (Ketchum & Benson, Reference Ketchum and Benson2010). The present specimen shows two small surfaces in a more central-lateral position, and there is no noticeable hint at a dumbbell shape. Elasmosaurid cervical centra usually have paired foramina that are present both ventrally and dorsally (Wintrich et al., Reference Wintrich, Scaal and Sander2017) and variation in the number of foramina occurs along the axial column of any individual (e.g. O’Gorman et al., Reference O’Gorman, Coria, Reguero, Santillana, Mörs and Cárdenas2018). NHMM 2014 027 has three foramina dorsally and two ventrally; based on these observations, a former position in the caudal series appears the most plausible interpretation. One issue with this interpretation is the lack of chevron facets on the specimen. We attribute this to the fact that the lateral edges of the articular surfaces are unossified.
An additional method of determining the position within the axial skeleton of a vertebral centrum is the Vertebral Length Index (VLI), a unit-less figure which quantifies the ratios in dimensions of a centrum using the following formula:

in which L stands for the length of the centrum (measured antero-posteriorly on the ventral side) and H and W the height (dorsoventral axis) and width (dextrosinistral) measured on the same articular surface (usually posteriorly), respectively (O’Keefe & Hiller, Reference O’Keefe and Hiller2006). The VLI increases over cervical positions in a posterior direction, prior to decreasing towards the posteriormost cervical centra and decreasing further along the dorsal and caudal centra. To date, VLI measurements have largely focused on cervical centra and only limited data are available on VLI values in plesiosaur caudal centra as a whole. The single elasmosaur for which VLI data of the entire vertebral column have been published is Vegasaurus molyi from the Snow Hill Island Formation in Vega Island, Antarctica (O’Gorman et al., Reference O’Gorman, Salgado, Olivero and Marenssi2015). VLI values of all five caudal vertebrae described were roughly the same, between 65 and 75. NHMM 2014 027 has a VLI of 67.5 (100 * (28/[(38 + 45) * 0.5)]). Based on the values given for Vegasaurus this would correspond to an anterior caudal position.
Ontogenetic stage – There is no indication of fusion of a neural arch to the centrum, nor does the articular surface show any sign of post-mortem taphonomic loss (fracturing). In this respect, there is correspondence with the relatively large dorsal foramina and the porosity surrounding these. The presence of a notochord pit may also be considered a juvenile feature, because the notochord gradually disappears during ontogeny. The plesiosaur axial skeleton can be subject to paedomorphism, whereby a likely sexually adult individual, based on size, is osteologically still immature (Araújo et al. Reference Araújo, Polcyn, Lindgren, Jacobs, Schulp, Mateus, Olímpio Gonçalves and Morais2015). Likewise the fusion of the neural arches to caudal centra in plesiosaurs likely occurs later than in more anterior vertebrae (Vincent, Reference Vincent2010). We therefore consider this specimen osteologically immature and possibly juvenile (Brown, Reference Brown1981; Mulder et al., Reference Mulder, Bardet, Godefroit and Jagt2000; O’Gorman et al., Reference O’Gorman, Gasparini and Salgado2014, p. 44; Otero et al., Reference Otero, Soto-Acuña, Vargas and Rubilar-Rogers2014, pp. 323–325).
Taphonomy and faunal assemblages – Plesiosaur material from the type area of the Maastrichtian Stage is exceedingly rare and so far limited to the family of the Elasmosauridae. The absence of plesiosaurs is in striking contrast to the large numbers of mosasaur and other vertebrate fossils recovered from the same strata (Schulp et al., Reference Schulp, Janssen, Van Baal, Jagt, Mulder and Vonhof2016). However, elsewhere, in particular at several localities in the Southern Hemisphere, elasmosaurs are much more abundant than mosasaurs. Attempts have already been made to explain the near-absence of Elasmosauridae in the type Maastrichtian. The δ13C ratio in marine vertebrates has been used as a proxy for trophic level, general habitat and diving behaviour, with the latter two aspects usually overshadowing the first (Schulp et al., Reference Schulp, Janssen, Van Baal, Jagt, Mulder and Vonhof2016). In that paper, the authors noted that the δ13C ratio of their elasmosaur samples was similar to the range of δ13C values of the mosasaur samples they had studied, which is why they concluded that the near-absence of elasmosaurs in the Maastrichtian type area did not primarily reflect differences in trophic level or diving behaviour between the two groups. However, at other localities where Maastrichtian rocks crop out, mosasaurs and elasmosaurs have been shown to have coexisted. Maastrichtian-aged assemblages in Angola, Morocco and California (USA) have all yielded mosasaur material in abundance, as well as a fair share of elasmosaurs (Strganac et al., Reference Strganac, Jacobs, Polcyn, Mateus, Myers, Salminen, May, Araújo, Ferguson, Olímpio Gonçalves, Morais, Schulp and da Silva Tavares2014). In faunal assemblages of the Maastrichtian Weddellian Province, elasmosaurs even predominate. These regions have all been linked to oceanic upwelling (e.g. Mulder et al. Reference Mulder, Bardet, Godefroit and Jagt2000; Vincent et al., Reference Vincent, Bardet, Houssaye, Amaghzaz and Meslouh2013; Otero et al., Reference Otero, Soto-Acuña, Salazar and Oyarzún2015). Abundant prey, connected to high levels of primary productivity, would appear to be a prerequisite for the abundance of both elasmosaurs and mosasaurs in the same faunal assemblage. Localities without a link to oceanic upwelling, such as the North Atlantic Ocean, inclusive of the Maastrichtian type area, and the Western Interior Seaway in particular, have few elasmosaurs, whereas mosasaurs constitute the apex predators in these assemblages. The limited material on record from the type Maastrichtian (with the present vertebra added, amounting to merely eight specimens after more than 250 years of collecting) attests to a more offshore or migratory mode of life of plesiosaurs in these regions. As such, the present discovery provides a rare glimpse into the distribution and diversity of a poorly represented group of marine reptiles.
Acknowledgements
Many thanks are extended to the Mergelbouwsteen Kleijnen company for donating the specimen to the Natuurhistorisch Museum Maastricht. We want to thank an anonymous reviewer as well as reviewer Sven Sachs, whose helpful remarks greatly improved the manuscript. We also thank Dr P.G. Miedema (father of the corresponding author) for his help with collection visits in the Netherlands. Finally we would like to thank Dylan Bastiaans for his help in the preparation of the specimen.



 Open access
Open access