
104 results
Australian square kilometre array pathfinder: I. system description
- Part of
-
- Journal:
- Publications of the Astronomical Society of Australia / Volume 38 / 2021
- Published online by Cambridge University Press:
- 05 March 2021, e009
-
- Article
-
- You have access
- HTML
- Export citation
-
In this paper, we describe the system design and capabilities of the Australian Square Kilometre Array Pathfinder (ASKAP) radio telescope at the conclusion of its construction project and commencement of science operations. ASKAP is one of the first radio telescopes to deploy phased array feed (PAF) technology on a large scale, giving it an instantaneous field of view that covers
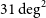 $31\,\textrm{deg}^{2}$ at
$31\,\textrm{deg}^{2}$ at 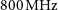 $800\,\textrm{MHz}$. As a two-dimensional array of 36
$800\,\textrm{MHz}$. As a two-dimensional array of 36 $\times$12 m antennas, with baselines ranging from 22 m to 6 km, ASKAP also has excellent snapshot imaging capability and 10 arcsec resolution. This, combined with 288 MHz of instantaneous bandwidth and a unique third axis of rotation on each antenna, gives ASKAP the capability to create high dynamic range images of large sky areas very quickly. It is an excellent telescope for surveys between 700 and
$\times$12 m antennas, with baselines ranging from 22 m to 6 km, ASKAP also has excellent snapshot imaging capability and 10 arcsec resolution. This, combined with 288 MHz of instantaneous bandwidth and a unique third axis of rotation on each antenna, gives ASKAP the capability to create high dynamic range images of large sky areas very quickly. It is an excellent telescope for surveys between 700 and 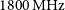 $1800\,\textrm{MHz}$ and is expected to facilitate great advances in our understanding of galaxy formation, cosmology, and radio transients while opening new parameter space for discovery of the unknown.
$1800\,\textrm{MHz}$ and is expected to facilitate great advances in our understanding of galaxy formation, cosmology, and radio transients while opening new parameter space for discovery of the unknown.
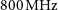

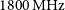
The evidence for cognitive behavioural therapy in any condition, population or context: a meta-review of systematic reviews and panoramic meta-analysis
-
- Journal:
- Psychological Medicine / Volume 51 / Issue 1 / January 2021
- Published online by Cambridge University Press:
- 18 January 2021, pp. 21-29
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
7 - Dinophyta
- from Part IV - Evolution of One Membrane of Chloroplast Endoplasmic Reticulum
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 250-297
-
- Chapter
- Export citation
Dedication
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp v-vi
-
- Chapter
- Export citation
Preface to the First Edition
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp xiii-xiv
-
- Chapter
- Export citation
12 - Heterokontophyta, Eustigmatophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 339-342
-
- Chapter
- Export citation
23 - Algae and the Environment
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 492-509
-
- Chapter
- Export citation
Part III - Evolution of the Chloroplast
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 77-79
-
- Chapter
- Export citation
13 - Heterokontophyta, Pinguiophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 343-344
-
- Chapter
- Export citation
Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 303-308
-
- Chapter
- Export citation
11 - Heterokontophyta, Synurophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 334-338
-
- Chapter
- Export citation
Index
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 526-535
-
- Chapter
- Export citation

Phycology
-
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018
-
- Textbook
- Export citation
10 - Heterokontophyta, Chrysophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 318-333
-
- Chapter
- Export citation
18 - Heterokontophyta, Raphidophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 397-400
-
- Chapter
- Export citation
15 - Heterokontophyta, Pelagophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 351-353
-
- Chapter
- Export citation
2 - Cyanobacteria
- from Part II - The Prokaryotic Algae
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 32-76
-
- Chapter
- Export citation
22 - Prymnesiophyta
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 470-491
-
- Chapter
- Export citation
21 - Heterokontophyta, Phaeophyceae
- from Part V - Evolution of Two Membranes of Chloroplast Endoplasmic Reticulum and the Chlorarachniophyta
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 415-469
-
- Chapter
- Export citation
Part IV - Evolution of One Membrane of Chloroplast Endoplasmic Reticulum
-
- Book:
- Phycology
- Published online:
- 06 August 2018
- Print publication:
- 01 March 2018, pp 231-234
-
- Chapter
- Export citation
