

Introduction
Behavioral genetics (BG) is an area of study that has, in a relatively short period of time, evolved from a promising but unproven perspective to one that is now forming the foundation for many studies of human behavior. We are now at a point where BG evidence can be summarized into “10 replicated findings” (Plomin, DeFries, Knopik, & Neiderhiser, Reference Plomin, DeFries, Knopik and Neiderhiser2016), where one of the largest meta-analyses ever conducted summarizes BG estimates (Polderman et al., Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015), and where there are not only “laws” of BG (Turkheimer, Reference Turkheimer2000), but there is even talk of expanding those laws (Chabris, Lee, Cesarini, Benjamin, & Laibson, Reference Chabris, Lee, Cesarini, Benjamin and Laibson2015). This is all to say the BG perspective has grown at a rapid clip and is now recognized as one of the most instructive ways to study human development.
Against this backdrop, the current chapter has three primary objectives. First, we will offer a brief overview of the BG perspective as it applies to the study of human aggression and violent behavior. As we see it, BG research can be divided into two broad approaches: (1) methods that seek to estimate the degree to which genetic and environmental influences affect the development of a phenotype; and (2) methods that seek to identify which genes/environments influence the phenotype. This leads to our second objective, which is to provide an overview of both of the above-mentioned areas of BG research. Finally, our third objective is to explain how these two areas of BG research have been used to develop a more holistic understanding of the etiology of human aggression and violent behavior.
The Foundation of Behavioral Genetics
Behavioral genetics (BG) developed out of the quantitative genetics paradigm. Space provisions do not allow a full review of either of these perspectives. But, briefly, one can think of quantitative genetics as the area of study that integrates molecular principles learned from biology and genomics to the study of phenotypic scores and variation (see, generally, Falconer & Mackay, Reference Falconer and Mackay1996). Quantitative geneticists can be credited with a simple, yet revelatory, proposal about the sources influence on a phenotype (note that phenotype is the term applied to any trait or outcome that varies in a population): a phenotype is made up of genetic and environmental components, such that:

where P is the phenotypic score for an individual, G is that person’s genotypic score, and E is that person’s environmental score plus noise/error.
This deceptively simple equation conceals many important philosophical and mathematical points. First, note that the G is listed before the E. Although the left-hand side of the equation is identical regardless of whether G or E is listed first, there is a great philosophical divide that can be identified here. Specifically, BG researchers and quantitative geneticists are trained to see the world such that G affects P and any influence of E can be thought of as noise. E, in this framework, becomes a nuisance parameter; something that must be accounted for but is not necessarily of primary interest.
Sociologically oriented researchers are often trained to see the world such that P = E + G. Here, the environment E is the primary influence of phenotypic scores and genetic influences G are the noise that must be controlled/accounted for (see, generally, Rafter, Posick, & Rocque, Reference Rafter, Posick and Rocque2016).
This is, of course, a gross oversimplification. But it is important to differentiate these perspectives because most phenotypes are not defined in a simple linear way, meaning it will not be possible to view one element of the equation as more important than the other. Instead, most phenotypes – including those related to aggression and violent behavior – arise due to a complicated causal process that involves G, E, their covariance (rGE), and their interaction (G × E). In other words, our simple equation must be expanded:

Because genetic and environmental influences are likely to covary and interact with one another, the researcher cannot ignore one to focus on the other. As Burt (Reference Burt2016, p. 114) recently noted, “Indeed, one important, and often overlooked, consideration in studies of environmental influences is that the ‘environment’ may not be genetically independent of the outcome variable…” This quote serves to remind us that rGE and G × E make it so that we cannot accurately estimate the impact of E on P unless we have made an effort to account for the influence of G.
The remaining sections of this chapter will, therefore, be built around a central theme: research into the etiology of human aggression and violence is complicated due to the covariance and interaction of G and E. But this is not to say that research endeavors are doomed due to irreducible complexity. On the contrary, advances have been and will continue to be made as we (scientists) incrementally work our way to a more holistic understanding of what makes humans harm one another. Below, we consider the two main streams of research that have both (1) recognized the complexity discussed above and (2) attempted to further our understanding of the causes of violence and aggression by studying G and E.
Two Modern Approaches
Generally speaking, there are two types of BG analyses. The first is what we will call variance decomposition research. This approach seeks to estimate the degree to which variance in a phenotype P can be attributed to genetic influences G and environmental influences E. The second type of BG analysis is what we will call gene finding research. Gene finding studies often begin by referencing the results from variance decomposition research. For example, imagine a variance decomposition analysis revealed some phenotype Pwas largely influenced by genetic influences G. But, as we will explain momentarily, variance decomposition only tells us how much G matters, not which genes matter. The latter issue is what the gene finding study would seek to address.
Variance Decomposition Methods
Variance decomposition models are a broad class of analytic strategies that primarily – although not exclusively – seek to estimate the impact of genetic G and environmental E influences on variance in the phenotype P. Obviously, the emphasis here is on variance in P, which redirects our focus away from any specific predicted value of P and instead forces us to think about what might cause P to exhibit variation in a population/sample. With these points in mind, variance decomposition methods deal with a slightly different version of the equations outlined above. Specifically, the focus now is on whether and how much variance in G and E contribute to variance in P:

where V stands for variance and the other subscripts are consistent with their previous definitions.
In addition to redirecting our focus to variance in P (i.e., VP), variance decomposition models can also be credited with highlighting an important point about the environmental component that has been expressed, to this point, as E in our mathematical models. Specifically, when variance decomposition models were being developed, it became apparent that more than one environmental parameter was needed to account for all of the observed variance in P. Thus, scholars who were at the forefront of model development proposed that the E component be separated into two parts: one for the “common” environment C that could account for between-family variation and one for the within-family variation.
The reason C stands for the common environment is worth attention. Early model development was carried out by studying twins and the sources of variance that caused two twins from the same family to develop similarly. Of course, it is obvious that any environment shared between two twins should make them more similar to one another. This influence, therefore, is captured by the common environment C. But observation tells us that twins do not always develop identically (Bouchard et al., Reference Bouchard, Lykken, McGue, Segal and Tellegen1990). Even though this point was obvious, it was not obvious how this could happen if the environment had a single effect captured by C. Thus, as these models were being developed, the source of variance that led to differences between twins was captured by an extra environmental component (it may have even been considered “error”) and was labeled E. It was not until sometime later that this component was coined the “nonshared” environment (though the E label stuck).
Because twins were – and often remain – the focus, it was also obvious that variance decomposition models would need to account for heritable variation. Indeed, this was the focus for much of the early research that used these techniques. But the variance in P that is due to genetic factors – like its environmental counterpart(s) – comes in more than one version. Broadly, we can speak about the heritable variation in P, which is akin to calculating:

The calculation above reveals that if one were to estimate a simple ratio of the variance in P that were due to variance in G, one would actually be calculating what has come to be known as broad-sense heritability (H2). This estimate is referred to as broad-sense because it is now well understood that G actually captures the influence of at least three forms of genetic influence (ignoring, for the time being, G × E and rGE):

where G still refers to the genetic influence, A identifies the additive genetic influence, D refers to the dominance genetic influence, and I captures epistatic genetic influence. Page limitations do not allow us to outline the differences between these various types of genetic influence so interested readers are directed to Falconer and Mackay (Reference Falconer and Mackay1996) and Plomin, DeFries, Knopik, and Neiderhiser (Reference Plomin, DeFries, Knopik and Neiderhiser2013). For a host of various reasons (Hill, Goddard, & Visscher, Reference Hill, Goddard and Visscher2008; Zuk, Hechter, Sunyaev, & Lander, Reference Zuk, Hechter, Sunyaev and Lander2012), much of the variance decomposition focus has been given to the genetic influences captured by A, such that estimates of h2 in most variance decomposition models captures what is known as narrow-sense h2:

Relevant Findings
So what has more than half-a-century of research using variance decomposition models shown us? This was precisely the question that a recent meta-analysis conducted by Polderman and colleagues (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) sought to address. As you might imagine, this was a massive undertaking. More than 2,700 studies were included in the review, leading to a grand total of 17,804 phenotypes being analyzed among more than 14.5 million twin pairs. Across all these traits, the mean h2 estimate was 0.49, leaving 0.51 attributable to the environment. Given the above discussion about the distinction between broad-sense H2 and narrow-sense h2, it is important to note that Polderman and colleagues (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) reported that a majority of the studies included in the meta-analysis found evidence to support the simpler, additive version of genetic influence (i.e., narrow-sense h2).
A mean h2 estimate of roughly 0.50, as it turns out, was not all that surprising. Indeed, Turkheimer’s (Reference Turkheimer2000) three laws of BG had anticipated this finding 15 years earlier. Polderman and colleagues’ (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) study, thus, did researchers a huge service by putting this prediction to the test. Also, the authors did not simply describe the mean h2 across all those 17,000+ phenotypes; rather, they provide h2 estimates for different domains of phenotypes. Some of the major domains are discussed in their paper, but others – especially those that were too nuanced to single out in the publication – are posted to the MaTCH website that was created to accompany the study (see http://match.ctglab.nl/#/home). Exploration of the MaTCH webtool reveals that conduct disorders was the phenotype classification that most closely resembled the traits of focus here (i.e., aggression and violence). Indeed, “aggressive behavior” was one of the most frequently used terms in the papers that were classified as having analyzed conduct disorders (see the word cloud function at http://match.ctglab.nl/#/specific/cloud). Estimates gleaned from nearly 300,000 pairs of twins revealed that this phenotype (i.e., conduct disorder) has a mean h2 estimate of approximately 0.486.
A h2 estimate of 0.486 is right in line with previous meta-analyses and systematic reviews (Mason & Frick, Reference Mason and Frick1994; Rhee & Waldman, Reference Rhee and Waldman2002). For example, the meta-analysis of aggression performed by Burt (Reference Burt2009) suggested h2 for aggressive forms of rule-breaking was 0.65, while the systematic review performed by Raine (Reference Raine1993) concluded the h2 estimate for various forms of delinquency and criminal behavior was probably in the small (e.g., perhaps around 0.20) to moderate (e.g., around 0.50) range. In all, research using variance decomposition methods to study the elements that contribute to variation in aggression and violent behavior has converged on a key conclusion: about 50% of the variation in these behaviors is due to heritable factors and the remaining portion is due to environmental factors.
Limitations
As can be deduced from the discussion above, there has been a lot of empirical attention paid to the h2 of aggressive, violent, and antisocial behavior (cf., Barnes, Boutwell, & Beaver, Reference Barnes, Boutwell, Beaver and Piquero2016; Veroude et al., Reference Veroude, Zhang-James, Fernàndez-Castillo, Bakker, Cormand and Faraone2016; Waltes, Chiocchetti, & Freitag, Reference Waltes, Chiocchetti and Freitag2016). But a fair amount criticism has be leveled against these studies (Wright et al., Reference Wright, Barnes, Boutwell, Schwartz, Connolly, Nedelec and Beaver2015). Although critics have invoked a number of arguments and highlighted several limitations, two concerns are commonly raised. The first is that variance decomposition methods often rely on twin pairs, leaving open the possibility that the results from these studies do not generalize to the singleton (i.e., non-twin) population. This concern was assessed by Barnes and Boutwell (Reference Barnes and Boutwell2013), who tested whether twins systematically differed from non-twin subjects on a range of phenotypes that are typically of interest to scholars studying aggressive and violent behavior. Specifically, Barnes and Boutwell (Reference Barnes and Boutwell2013) analyzed whether twins differed from their non-twin counterparts on 27 phenotypes, which included an index of delinquent behavior, victimization experiences, levels of self-control, drug-using behaviors, and the respondent’s level of involvement with delinquent peers. The findings from this analysis, although nuanced in places, are easy to summarize: twins and non-twins do not differ on most traits. Thus, there is little reason to worry that results from, say, the Polderman et al. (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) study will fail to generalize to the broader population of singletons.
The second concern involves something known as the equal environments assumption (EEA). Briefly, the EEA states that identical twins (i.e., monozygotic [MZ]) share the same amount of their environment as nonidentical (i.e., dizygotic [DZ]) twins. Put differently, the degree to which twins share their environment is assumed to be uncorrelated with their level of genetic overlap (i.e., whether they are MZ or DZ). Critics argue that the assumption is only rarely met, drawing into question the inferences that can be taken from variance decomposition research. Luckily, there are now dozes of studies that have analyzed the degree to which the EEA is violated in behavioral genetic research. The results of these studies were summarized by Barnes and colleagues (Reference Barnes, Wright, Boutwell, Schwartz, Connolly, Nedelec and Beaver2014) and the conclusions were clear: the EEA is a robust assumption that, even if it is violated, is unlikely to lead to large biases in BG research.
Beyond the two criticisms covered above, there are other concerns with variance decomposition models that – while not reaching the level of a flaw or even a limitation per se – do restrict their utility. Two broad points stand most prominent. First, variance decomposition models “lump together” the genetic influences such that one does not know which, or even how many, genes contribute to the h2 estimate (and the same goes for the environmental estimates). Thus, the h2 estimate can be thought of as a global average estimate that sums over the influence of an unknown number of genes with unknown effect sizes. This raises one’s attention to the importance of the BG strategies discussed in the next section; those that seek to find the genes that influence variation in P. The second concern to be aware of is that variance decomposition models often are unable to model the effects of rGE and G × E. Although various modeling strategies are available to tease apart these influences (Purcell, Reference Purcell2002; Turkheimer & Harden, Reference Turkheimer, Harden, Reis and Jude2014), most variance decomposition studies do not utilize them. Thus, a large portion of the evidence base assumes that narrow-sense h2 is not only an appropriate proxy for broad-sense h2 (see above), but also that it is not confounded with rGE and G × E. To the extent that these latter assumptions are violated, estimates gleaned from variance decomposition models will be affected in predictable ways. Readers interested in learning more about the impact of omitting rGE and G × E are encouraged to see Purcell (Reference Purcell2002) and the methodological discussion offered by Turkheimer and Harden (Reference Turkheimer, Harden, Reis and Jude2014).
Gene Finding Methods
Although variance decomposition methods are extremely useful, they do not allow one to identify which genes play a role in the etiology of the phenotype P. This, therefore, has become the focus of what we will call gene finding techniques. In an effort to provide a meaningful, yet tractable, discussion of this approach to BG research, we have chosen to highlight two broad classes of gene finding strategies: (1) candidate gene studies and (2) genome-wide association (GWA) studies.
Both of these approaches rest on the same foundation of principles and aims. Specifically, they both seek to identify the specific genes that make up the G component of our P = G + E equation. Placed in the context of the variance decomposition models from above, recall that h2 is an estimate of the degree to which the variation in P is attributable to variation in (additive) G. Gene finding studies can be thought of as a tool that can help the researcher parse out the various genes that go into that h2 estimate.
So how do we go about finding the genes that are cloaked by the h2 estimate? First, we have to recast the P = G + E equation. Let us express the variance of P as a function of the variance in the genes plus variance in the environment, assuming no rGE and no G × E:
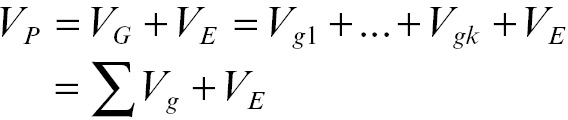
which simply shows that the variance in P that is attributable to G can be expressed as the summed influence of all the individual genes g that make up G.
If we assume the genetic influence captured by any specific gene is not affected by rGE or G × E, then we can seek to pull apart the h2 estimate one gene at a time:

where Vr captures the collective influence of the omitted genes and the environmental influences E.
So how does one go about searching for the individual genetic components that might play a role in the etiology of P? One might address the issue by drawing on previous literature to develop a hypothesis about the potential relationship between a specific genetic variant and P. We call this hypothesis-driven research and will, broadly speaking, align it with the candidate gene literature. A second strategy is to take a hypothesis-free approach and simply search the entire genome for any associations between all known genetic variants and P. We call this hypothesis-free research and align our discussion here with the development of GWA methods that have recently become the go-to approach for scholars studying the etiology of human complex traits like aggression (Pappa et al., Reference Pappa, St Pourcain, Benke, Cavadino, Hakulinen, Nivard and Evans2016) and antisocial behavior (Tielbeek et al., Reference Tielbeek, Johansson, Polderman, Rautiainen, Jansen, Taylor and Posthuma2017).
Hypothesis-Driven: Candidate Genes
Some of the first gene finding studies were conducted on animals. As such, gene finding research in humans is often guided by animal models, thus providing researchers studying human behavior the opportunity to develop a priori hypotheses about which genes might paly a role in the etiology of specific behaviors.
The molecular underpinnings of aggression have been explored among a range of subjects such as flies, zebra fish, rodents, monkeys, and other nonhuman primates (Olivier & Young, Reference Olivier and Young2002). Studies in Drosophila, using a forward genetic approach by selecting aggressive lines of flies, have indicated molecular targets such as octopamine, the insect equivalent of norepinephrine, and serotonin to be associated with fly aggression (Dierick & Greenspan, Reference Dierick and Greenspan2007, Reference Dierick and Greenspan2006; Zhou et al., Reference Zhou, Rao and Rao2008). Rodent studies have implicated brain circuits relevant to the etiology of aggression. For instance, a study utilizing an optogenetic design demonstrated that the stimulation of neurons in the mouse hypothalamus led to an increase of offensive aggression in male mice (Lin et al., Reference Lin, Boyle, Dollar, Lee, Perona, Lein and Anderson2011). Studies in rats have implicated the role of serotonin and dopamine in the regulation of aggression: with increased dopamine levels in the nucleus accumbens and decreased serotonin in the medial prefrontal cortex after the triggering of aggressive acts (van Erp & Miczek, Reference van Erp and Miczek2000). Early studies in monkeys reported lower levels of serotonin metabolite in the cerebrospinal fluid of high-ranked aggressive monkeys, compared to low-ranked monkeys (Higley et al., Reference Higley, Mehlman, Taub, Higley, Suomi, Linnoila and Vickers1992).
Although the use of such experimental designs in animal studies have a clear benefit, these controlled settings also eliminate potential relevant environmental factors that could have had crucial effects in a natural setting. Moreover, widely studied animal paradigms such as territorial or maternal aggression assess the more reactive type of aggression in which most individuals would fight, whereas pathological aggression can occur in situations where almost no one would fight (Nelson & Trainor, Reference Nelson and Trainor2007). The ecological validity is an important limitation hindering the translation of animal models to human models, which is not necessarily straightforward. For instance, the anatomical distribution of dopamine input and the expression of different dopamine receptor subtypes differ between rodents and primates (Berger, Gasper, & Verney, Reference Berger, Gaspar and Verney1991). Moreover, there has been a long debate within the field of translational neuroscience regarding the existence of the rodent prefrontal cortex and several scholars have questioned the homology between specific prefrontal regions in the rodent and human brain (Preuss, Reference Preuss1995; Uylings, Groenewegen, & Kolb, Reference Uylings, Groenewegen and Kolb2003; Wise, Reference Wise2008). Despite its limitations, animal studies have been extremely informative by indicating plausible biological pathways and candidate genes, thereby fueling the search for molecular targets in human aggression.
Relevant Findings
There has been an explosion of research activity surrounding candidate genes over the past 20 years or so (Dick et al., Reference Dick, Agrawal, Keller, Adkins, Aliev, Monroe and Sher2015). Research into the etiology of aggression and violence is no different. Building on the animal model results discussed above, as well as some of the earliest gene finding studies (see Brunner, Nelen, Breakefield, Ropers, & van Oost, Reference Brunner, Nelen, Breakefield, Ropers and van Oost1993), one of the first reports of a link between a candidate gene and antisocial behavior was published by Caspi and colleagues (Reference Caspi, McClay, Moffitt, Mill, Martin, Craig and Poulton2002). These researchers, drawing on a sample of New Zealanders, found evidence of a link between a specific gene on the X chromosome, the MAOA gene, and antisocial behavior. Certain variants of this gene were shown to have an association with antisocial outcomes for respondents who had reported being maltreated as a child. In other words, the “risk” alleles of the MAOA gene had an impact on aggression and violence if environmental risk was also present. Not only was this some of the first evidence to show a link between a candidate gene and human antisocial behavior, it was one of the first studies to report evidence of a G × E. Since Caspi and colleagues’ (Reference Caspi, McClay, Moffitt, Mill, Martin, Craig and Poulton2002) landmark study was published, others have sought to replicate the results. Some studies have were successful, while others have failed to replicate (Tilihonen et al., Reference Tilihonen, Rautiainen, Ollila, Repo-Tilihonen, Virkkunen, Palotie and Paunio2015; Vassos, Collier, & Fazel, Reference Vassos, Collier and Fazel2014).
Other candidate genes, too, have been studied. For example, Beaver and colleagues (Reference Beaver, Wright, DeLisi and Vaughn2008) analyzed the association between DRD2, DRD4, 5HTT, DAT1, MAOA, and desistance from delinquent/criminal behavior. Their results revealed that all but the 5HTT gene had a unique influence on desistance. Similar findings have been reported in various other studies, suggesting that there is robust evidence that these genes have important influences on the etiology of human antisocial behavior. But such research findings have, in recent years, raised at least as many question as have they answered (Charney, Reference Charney2012; Charney & English, Reference Charney and English2012). We consider a few of the most important questions generated by candidate gene research in the next subsection.
Limitations
There is now a relatively large body of evidence pointing to a handful of candidate genes and their relationship(s) with various forms of human antisocial behavior (see, generally, Charney & English, Reference Charney and English2012; Dick et al., Reference Dick, Agrawal, Keller, Adkins, Aliev, Monroe and Sher2015; Tielbeek et al., Reference Tielbeek, Linnér, Beers, Posthuma, Popma and Polderman2016). But, as with many new advancements, the proliferation of findings has outpaced methodological discussion about the limitations of this line of work. Dick and her colleagues (Reference Dick, Agrawal, Keller, Adkins, Aliev, Monroe and Sher2015), therefore, recently set out to rectify this concern by outlining some of the most important limitations of the candidate gene literature. At the same time, they set forth a number of recommendations for how candidate gene research can continue to develop in a way that will promote confidence in the literature base.
Confidence in a literautre base is important to consider because some areas of work have recently fallen into a crisis of confidence. The crisis began when Ioannidis (Reference Ioannidis2005) noted there is a very real chance the majority of published research findings are false-positives. This possibility is, at the very least, a realistic concern for the candidate gene literature. Some have even argued this possibility is heightened in the candidate gene literature due to the way in which scholars in the area have conducted these studies (Duncan & Keller, Reference Duncan and Keller2011). Some of the concerns surrounding candidate gene research can be traced to the very issues highlighted by Ioannidis (Reference Ioannidis2005). Others are linked to the modern practice of statistical analysis, one that may be biased by multiple testing effects or, at a minimum, is biased by what Gelman and Loken (Reference Gelman and Loken2014) called the “garden of forking paths” that emerge during the course of any statistical analysis. A key concern is that scholars may have approached many candidate gene studies as an exploratory exercise, overlooking the importance of a priori hypothesis development.
Recognizing these points, Dick and colleagues (Reference Dick, Agrawal, Keller, Adkins, Aliev, Monroe and Sher2015) set forth a “recipe” for candidate gene researchers to follow, especially those interested in studying G × Es. Rather than restating or briefly summarizing their points, we strongly encourage readers to consider the recommendations made by Dick et al. (Reference Dick, Agrawal, Keller, Adkins, Aliev, Monroe and Sher2015) for themselves.
Hypothesis-free: Genome-wide Association (GWA)
The newest wave of gene finding research is known as genome-wide association (GWA). Although the technical details of a GWA are beyond the scope of this chapter, it is important that we give some indication of how GWA studies are conducted. Plomin and colleagues (Reference Plomin, DeFries, Knopik and Neiderhiser2013, p. 143) define a GWA as “An association study that assesses DNA variation throughout the genome.” In other words, GWA searches the entire genome for any association between known genetic variants and the phenotype of focus. The process is carried out by running hundreds of thousands (sometimes even millions) of association tests and by correcting P-values for the multiple testing bias that is expected. In the end, corrected P-values are referenced to try and identify which genetic variants may have an influence on phenotypic development.
This task of GWA is easier to envision if one imagines a researcher has genetic information on a sample of respondents. Imagine also that the researcher has phenotypic data on those same respondents. For example, perhaps the researcher collected information about whether the participant had ever been diagnosed with antisocial personality disorder (ASPD). Given the abovementioned concerns stemming from candidate gene research, this researcher decides a hypothesis-free design is most appropriate for his/her study that seeks to identify whether any common genetic variants are linked to ASPD. At this point, the researcher has several ways s/he could move forward. Perhaps s/he could simply analyze the association between ASPD diagnosis and every single nucleotide polymorphism (SNP) that was tagged. This would remove the need to hypothesize about the influence of any one gene (or SNP). But a key issue would emerge: specifically, the biasing effects that would accompany that many analyses. To be sure, modern genetic technologies often identify upwards of 1,000,000 individual SNPs. If the researcher were to run 1,000,000 logistic regression models with a standard α-level of 0.05, then s/he could expect to receive back roughly 50,000 statistically significant results even if none of the SNPs had an effect on the phenotype. Thus, one of the key problems facing gene finding research is that the risk for Type I error is too high and must be controlled. And this is exactly what GWA does. To be specific, GWA results must reach genome-wide significance values of P < 5 × 10−8 (or P < 0.00000005).
Risch and Merikangas (Reference Risch and Merikangas1996) are sometimes credited as being the first to reveal the power of GWA for modern BG research (Jorgenson & Witte, Reference Jorgenson and Witte2006). Risch and Merikangas (Reference Risch and Merikangas1996) showed that association studies have more statistical power than linkage analyses, the latter of which were a popular research approach for early BG scholars interested in gene finding techniques. But modern GWA has also had to face issues surrounding statistical power. As we will discuss in the Limitations section below, statistical power is often quite low for any given GWA, which has led researchers to the realization that sample sizes of a magnitude never seen in the social sciences are necessary to reliably detect the signals in the genome. It has also led to other unique challenges that are currently facing the big data era (see, for a discussion, Liu & Guo, Reference Liu and Guo2016).
Relevant Findings
GWA is still, relatively speaking, a new technology that is just beginning to come of age. Only in the past ten years or so has GWA become available to researchers interested in studying human behavioral phenotypes. The vast majority of early GWAs were conducted in the medical sciences by researchers interested in understanding disease traits. As prices for the technology necessary to conduct a GWA came down, more and more studies were carried out on a range of human behavioral outcomes. There are now seven GWA studies on human aggressive behavior (or some closely related phenotype such as violence or antisocial behavior) (Brevik et al., Reference Brevik, van Donkelaar, Weber, Sánchez-Mora, Jacobs, Rivero and Cormand2016; Dick et al., Reference Dick, Krueger, Edwards, Agrawal, Lynskey, Lin and Almasy2011; Pappa et al., Reference Pappa, St Pourcain, Benke, Cavadino, Hakulinen, Nivard and Evans2016; Rautiainen et al., Reference Rautiainen, Paunio, Repo-Tiihonen, Virkkunen, Ollila, Sulkava and Tiihonen2016; Salvatore et al., Reference Salvatore, Edwards, McClintick, Bigdeli, Adkins, Aliev and Nurnberger2015; Tielbeek et al., Reference TiChabelbeek, Medland, Benyamin, Byrne, Heath, Madden and Verweij2012, Reference Tielbeek, Johansson, Polderman, Rautiainen, Jansen, Taylor and Posthuma2017). The first ever GWA conducted on a form of antisocial behavior was reported by Dick and colleagues (Reference Dick, Krueger, Edwards, Agrawal, Lynskey, Lin and Almasy2011). Their GWA study was performed on a sample of children (n = 3,963) and the phenotypic outcome was conduct disorder. Four genetic loci reached genome-wide significance and were pointed to as potential markers for future research to study in a candidate gene approach.
The first GWA study performed on adult antisocial behavior was reported by Tielbeek and colleagues (Reference TiChabelbeek, Medland, Benyamin, Byrne, Heath, Madden and Verweij2012). Analyzing data from nearly 5,000 respondents did not produce any genome-wide statistically significant loci. But there were a few loci that were strongly associated and at least suggestive of an association. One of the most recent GWA studies was conducted by Rautiainen et al. (Reference Rautiainen, Paunio, Repo-Tiihonen, Virkkunen, Ollila, Sulkava and Tiihonen2016) and the phenotypic outcome was antisocial personality disorder. Results identified several loci that might be associated with the phenotype.
A recent review summarizing previous candidate gene and GWA studies on aggression and closely related phenotypes found 156 genes to be nominally (P < 0.05) associated (Fernàndez-Castillo & Cormand, Reference Fernàndez-Castillo and Cormand2016). These genetic loci are predominantly involved in dopaminergic and serotonergic neurotransmission and in hormone regulation. Moreover, that same study found certain biological pathways were overrepresented across the six GWA studies, revealing significant enrichment in axon guidance, estrogen receptor signaling, neurodevelopmental processes, and synaptic plasticity pathways. Nonetheless, the authors stress that well-powered replication designs utilizing standardized measurements and more homogenous subtypes of aggression are needed to reliably identify pathways affecting aggressive behavior.
Limitations
Although GWA studies help to address some of the well-known concerns with candidate gene research, they are not without their own limitations. A number of studies have raised awareness of perhaps the most important issue surrounding GWA research: the influence of underpowered study designs in biomedical and psychological research (see, generally, Button et al., Reference Button, Ioannidis, Mokrysz, Nosek, Flint, Robinson and Munafò2013; Ioannidis, Reference Ioannidis2005; Sham & Purcell, Reference Sham and Purcell2014; Simmons, Nelson, & Simonsohn, Reference Simmons, Nelson and Simonsohn2011).
Three key problems can arise when a study suffers from low statistical power: (1) type II error increases to unacceptable levels (i.e., there are too many false-negative findings); (2) false-positive rates increase to an unacceptable level; and (3) reported effect sizes are likely to be inflated due to the “winner’s curse.” There is reason to believe that these issues have affected genetic association studies of all sorts, even those published in journals with a high impact factor (Munafo, Stothart, & Flint, Reference Munafo, Stothart and Flint2009).
Genetic epidemiological studies often lack sufficient statistical power, which then leads to low replication rates (Ioannidis, Reference Ioannidis2003; Sullivan, Reference Sullivan2007). As discussed earlier, candidate gene research proposed a number of genes or genetic variants as predictors of variation in aggression and violence. These candidate gene studies were meta-analyzed by Vassos and colleagues (Reference Vassos, Collier and Fazel2014) who found no evidence of a significant association, despite prior replication efforts. These findings indicate that either previous candidate gene results were false positives or that the previous effect size estimates were highly exaggerated (i.e., the winner’s curse). In light of these findings, it has become increasingly clear that antisocial behavior is a highly complex trait, with a large number of genetic influences, each of which only contributes a small amount of variance to the phenotype. Thus, scholars should seek to perform statistical power calculations prior to beginning a novel GWA study.
Such power calculations, however, require prior knowledge of the expected effect size of the tested genetic association. Power analyses could, therefore, be aided by effect size estimates of genetic variants derived from studies on other complex traits yielding a comparable heritability. Well-powered study designs of consortia, such as the Social Science Genetic Association Consortium (SSGAC), can anchor power analyses in other studies by taking the effect sizes of their most significant associations as an upper limit for the effect sizes in genetically similar traits. The SSGAC GWA on educational attainment reported an estimated R2 of 0.02% for all their genome-wide significant genetic variants (Okbay et al., Reference Okbay, Beauchamp, Fontana, Lee, Pers, Rietveld and Oskarsson2016). These estimates are uncorrected for inflation due to the winner’s curse and therefore might still overestimate the magnitude of their effects (Pereira & Ioannidis, Reference Pereira and Ioannidis2011). Thus, in GWA, the effect sizes are likely to be very small.
Power analyses can be performed via the user-friendly “pwr” package in the freely available statistical software R. This package contains functions for basic power calculations using effect sizes and notations from Cohen (Reference Cohen1988):pwr.r.test(r=<correlation coefficient>,
n=<sample size>, power=<0.8>,
alternative=“two.sided”, sig.
level=<EWAS=1E-7, GWAS=5E-8>)If we assume that each of the genetic variants has an effect size roughly equal to r = 0.02, we can calculate that in order to achieve 80% power to identify genetic variants associated with antisocial traits, a sample size of about 100,000 individuals is required. These power calculations reveal that, in hindsight, early GWA studies on antisocial behavior were underpowered. Figure 5.1 displays a statistical power curve for various sample sizes assuming an effect size of r = 0.02.
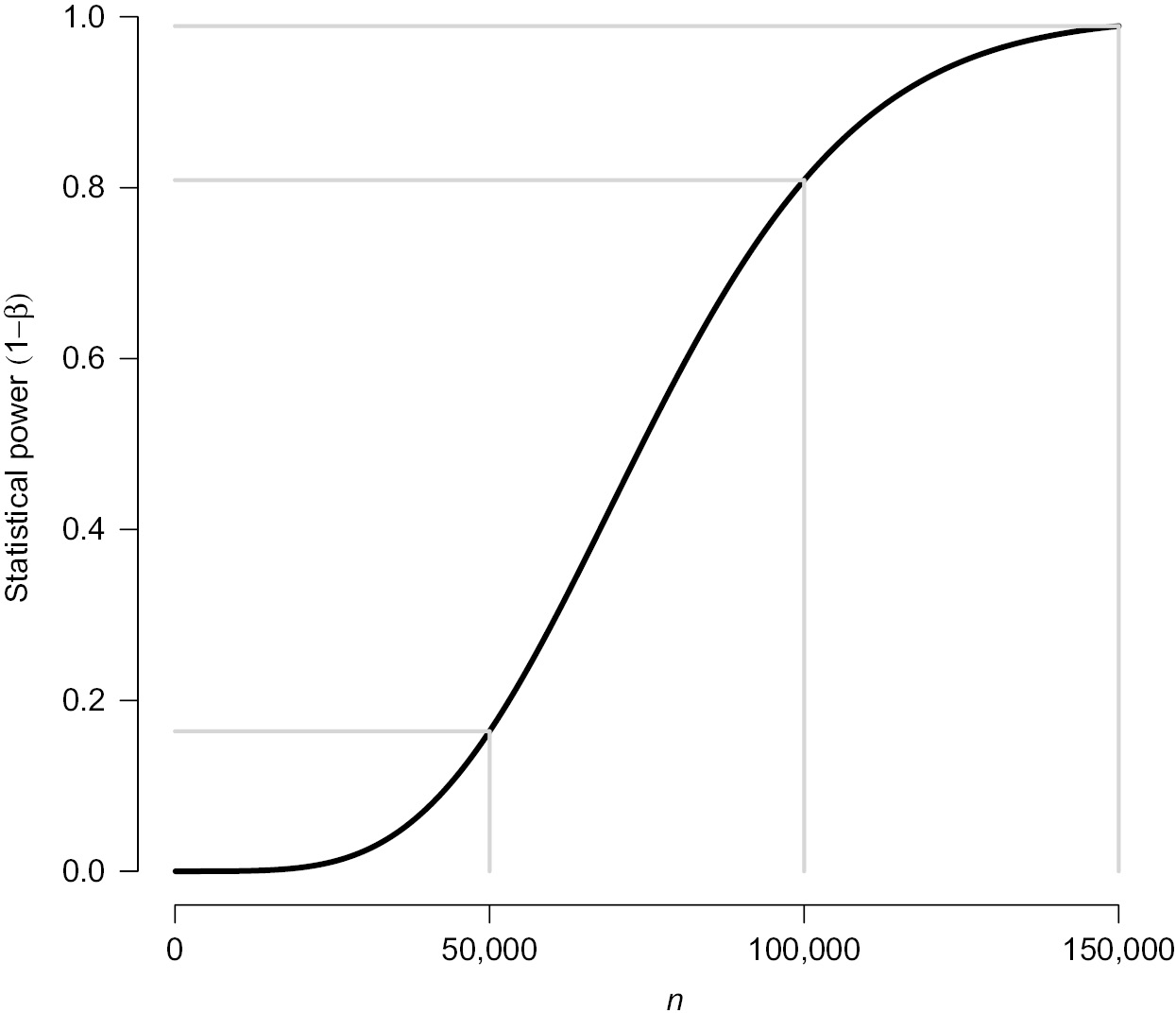
Figure 5.1 Statistical power at various sample sizes when r = 0.02.
Although GWA studies have been successful in detecting disease loci for outcomes such as Crohn’s disease, type 2 diabetes, and schizophrenia (see Figure 5.2; Franke et al., Reference Franke, McGovern, Barrett, Wang, Radford-Smith, Ahmad and Anderson2010; Schizophrenia Working Group of the Psychiatric Genomics Consortium, 2014; Scott et al., Reference Scott, Mohlke, Bonnycastle, Willer, Li, Duren and Prokunina-Olsson2007), the limitation of conventional single-marker association analyses focusing on complex outcomes, such as aggression, is that they have often been underpowered. The general conclusion that can be drawn from large-scale GWA studies is that aggression, violence, and antisocial behavior is likely to be influenced by many common causal genetic variants each conferring only a small effect (Chabris et al., Reference Chabris, Lee, Cesarini, Benjamin and Laibson2015; Tielbeek et al., Reference TiChabelbeek, Medland, Benyamin, Byrne, Heath, Madden and Verweij2012; Tielbeek et al., Reference Tielbeek, Johansson, Polderman, Rautiainen, Jansen, Taylor and Posthuma2017). Moreover, given the substantial polygenic component of antisocial behavior, it is likely that the genetic variants involved are not distributed randomly across the genome, but are distributed in ways that allow them to share a common biological function or pathway (Khatri, Sirota, & Butte, Reference Khatri, Sirota and Butte2012; Lips et al., Reference Lips, Cornelisse, Toonen, Min, Hultman and Posthuma2012). To partly overcome this issue, gene-based and pathways-based association analyses must begin to utilize prior biological knowledge on genes and pathways, yielding a more powerful analysis of GWA data (Wang, Li, & Hakonarson, Reference Wang, Li and Hakonarson2010).

Figure 5.2 Genome-wide association results as of 2013 (Hindorff et al., n.d.).
Dealing with Low Statistical Power: Gene-Set Analyses
Gene-based association analyses, rather than single-marker analyses, take into account all markers within a gene, thus examining the combined effect of all SNPs on the phenotype of interest. The advantage of a gene-based test is that it reduces the burden of multiple testing (to ≈ 21,000 tests, compared to millions of tests in GWA) and facilitates a more immediate gene-level interpretation. Moreover, because it aggregates multiple weak associations while accounting for linkage disequilibrium between the SNPs, it increases power and accounts better for heterogeneity in a gene (Liu, Wang, & Wong, Reference Liu, Wang and Wong2010).
Gene-set analyses (GSA), which are adopted from gene expression methods (Mootha et al., Reference Mootha, Lindgren, Eriksson, Subramanian, Sihag, Lehar and Houstis2003), are now commonly used in GWA studies. Strikingly, de Leeuw, Neale, Heskes, and Posthuma (Reference De Leeuw, Neale, Heskes and Posthuma2016), reviewing the statistical structure of GSA, found that as the heritability of a phenotype increased, the statistical power decreased. Moreover, larger sample sizes only modestly increased statistical power, with even less improvement for more strongly heritable phenotypes (de Leeuw et al., Reference De Leeuw, Neale, Heskes and Posthuma2016). Also, most of the current GSA tools do not sufficiently address bias due to gene size and linkage disequilibrium as well as the major limitation of confounding among gene sets, which could lead to false conclusions if not taken into account. In sum, gene-based and gene-set analyses offer a powerful tool in analyzing and interpreting genetic data, but some issues are still present, meaning further development of novel techniques should remain a priority.
Dealing with Low Statistical Power: Collaboration/Consortia
As we have shown above, issues surrounding low statistical power have recently occupied the minds of GWA analysts. Although there are several ways one can increase the statistical power of any given study (see above and, generally, Cohen, Reference Cohen1988), the most common way to induce higher power in GWA is to increase sample sizes. But the increases we are talking about are well beyond the level that could be expected by any one researcher or even a research team. For example, as was shown in Figure 5.1, a sample size of 100,000 is necessary to identify an effect size of r = 0.02; an effect size that is realistic and expected for a complex trait like aggression or violent behavior. As a result, GWA analysts have begun to form consortia so that they can combine their data in an effort to increase statistical power.
BG researchers’ willingness to form consortia represents a great example of a scientific community remaining critical of its own research and taking action to address known limitations. Of course, one’s scientific career is often judged by his/her unique contribution to the literature (see, generally, Merton, Reference Merton1957). Consortia are decidedly collaborative projects that often force a scientist to forgo his/her own unique interests in order to better the group’s collective efforts. While this may not be the best way forward for all research questions/areas, we see it as an extremely positive and commendable approach to addressing the problems facing the GWA community of scholars. With that said, one of us (JT) has established a consortium, known as the Broad Antisocial Behavior Consortium (BroadABC), for studying human antisocial behavior. The BroadABC is a collaborative research initiative to conduct genetic analyses at a large scale. Information about the BroadABC can be found here: http://broadabc.ctglab.nl.
The Future of BG Research into Human Aggression and Violence
In this final section of our chapter, we offer a brief discussion of five approaches that are sure to represent the next generation of BG research. Of course, a few sentences on any of these analytic techniques or broader perspectives does not even come close to doing justice to the true complexity that it entails. Thus, readers should not take these summaries as anything more than a cursory overview of what we believe will be the next wave of BG research.
Genome-Wide Complex Trait Analysis (GTCA)
Aggression and violent behaviors can be considered complex traits and, despite their considerable heritability (h2 ≈ 0.50), associated SNPs identified through GWA studies typically explain a fraction of estimated familial clustering. This considerable gap between twin-based heritability estimates and the tiny proportion of phenotypic variation explained by GWA has been generally referred to as the “missing heritability” problem (Eichler et al., Reference Eichler, Flint, Gibson, Kong, Leal, Moore and Nadeau2010; Manolio et al., Reference Manolio, Collins, Cox, Goldstein, Hindorff, Hunter and Chakravarti2009). Mixed-model techniques such as the genome-wide complex trait analysis (GCTA) software were introduced to partly overcome the missing heritability problem by testing the aggregate effect of all SNPs available on the GWA chips simultaneously on trait variance (Yang, Lee, Goddard, & Visscher, Reference Yang, Lee, Goddard and Visscher2011). The mixed-model technique and various extensions of it have been successful in characterizing the missing heritability of many complex traits (Lee, Wray, Goddard, & Visscher, Reference Lee, Wray, Goddard and Visscher2011; Yang et al., Reference Yang, Bakshi, Zhu, Hemani, Vinkhuyzen, Lee and van Vliet-Ostaptchouk2015, Reference Yang, Benyamin, McEvoy, Gordon, Henders, Nyholt and Visscher2010). When applied to children’s aggressive behavior, the proportion of the heritability estimated through GCTA software ranged from 0.10 to 0.54 in the different cohorts included in the EAGLE consortium (Pappa et al., Reference Pappa, St Pourcain, Benke, Cavadino, Hakulinen, Nivard and Evans2016). Since GCTA requires raw genotype data, its use remains mainly restricted to individual cohorts (unless raw data is being merged), thus yielding a limited sample size and estimates with wide confidence intervals. For example, when applied to adult antisocial behavior in an Australian study (Tielbeek et al., Reference TiChabelbeek, Medland, Benyamin, Byrne, Heath, Madden and Verweij2012), the proportion of the variance explained by common SNPs was 0.55; however, the standard error was high (0.41) and the estimate nonsignificant (P = 0.07).
Linkage Disequilibrium Score Regression
Recently, the linkage disequilibrium (LD) score regression method was introduced to decompose the inflated test statistics of GWA into true polygenic signal and confounding bias due to population stratification. The advantage of this ground-breaking tool is that it can use summary statistics, such as large meta-analyzed GWA datasets, to estimate SNP heritability in a fast and efficient manner (Bulik-Sullivan et al., Reference Bulik-Sullivan, Loh, Finucane, Ripke, Yang and Patterson2015). This means that LD regression can be performed by scholars who may not otherwise have access to genome-wide analytic resources. Indeed, we can confirm that LD regression can be estimated on many (if not most) modern personal computers. Also, the lab responsible for developing and maintaining the program has a very useful and information-rich webpage that indexes GWA results and LD regression output for hundreds of human phenotypes (http://ldsc.broadinstitute.org).
Polygenic Risk Scoring
SNP heritability estimates, if unbiased, can be helpful in determining the upper boundaries of genetic risk-prediction models, yet its clinical use is limited as it cannot serve as a guide for risk stratification (Chatterjee, Shi, & García-Closas, Reference Chatterjee, Shi and García-Closas2016). To construct risk-prediction models, a selection of markers highly associated with a trait are combined into a single polygenic risk score (PRS). First, a GWA is conducted in an initial, large training sample. Second, the SNPs are ranked based on their strength of association. Finally, polygenic scores are computed based on the weighted sum of the alleles associated with the trait in an independent replication sample (Dudbridge, Reference Dudbridge2013).
An example of such a training sample for antisocial behavior is the BroadABC, which was introduced above. The BroadABC recently performed such PRS analysis in an at-risk Finnish forensic replication sample by testing whether an antisocial genetic risk score – based on a broad conceptualization of ASB – could significantly discriminate between prisoners and matched controls (Tielbeek et al., Reference Tielbeek, Johansson, Polderman, Rautiainen, Jansen, Taylor and Posthuma2017). The software package PRSice (Euesden, Lewis, & O’Reilly, Reference Euesden, Lewis and O’Reilly2016) was employed to estimate the best-fit PRS at a broad range of P-value thresholds. Their analyses showed that case-control status of antisocial personality disorder in the Finnish prisoners sample was significantly predicted by antisocial genetic risk scores of the BroadABC, although the effect size was very small. The small effect size indicates the limited prediction accuracy and clinical utility for the current GWA outcomes. Nevertheless, this study demonstrated that population-based genetic association studies can be informative for samples that are at-risk. Therefore, large, high-quality epidemiological studies should inform and help build risk prediction models for aggression and violent behavior that incorporate both polygenic risk scores and environmental risk factors, along with their interaction (see Wertz et al., Reference Wertz, Caspi, Belsky, Beckley, Arseneault, Barnes, Corcoran, Hogan, Houts, Morgan, Odgers, Prinz, Sugden, Williams, Poulton and Moffitt2017).
Analyzing Endophenotypes with Neuroimaging
Clinical populations are likely to contain an admixture of etiologically different disorders (Bearden & Freimer, Reference Bearden and Freimer2006). Therefore, intermediate phenotypes, also referred to as endophenotypes, have been proposed as a less heterogeneous outcome that may help overcome some of the difficulties that arise when GWA is conducted on a complex phenotype. Endophenotypes are defined as “measurable components unseen by the unaided eye along the pathway between disease and distal genotype” and have become an important concept in psychiatry (Gottesman & Gould, Reference Gottesman and Gould2003, p. 636). In other words, the use of endophenotypes in genetic epidemiology seems promising since they are thought to be more proximal to the primary biological defects and are therefore hypothesized to be more sensitive to genetic signals than broad and heterogenic clinical phenotypes (Kendler & Neale, Reference Kendler and Neale2010).
In this respect, brain imaging offers a promising endophenotype-based approach by parsing the complexity of phenotypes into objectively measurable components. Converging evidence has emerged regarding promising brain endophenotypes for a range of antisocial phenotypes. For example, a meta-analysis conducted by Yang and Raine (Reference Yang and Raine2009) found significantly reduced prefrontal grey matter in antisocial individuals. Likewise, a study in psychopathic individuals reported significant bilateral volume reductions in the amygdala in these individuals compared with controls (Yang, Raine, Narr, Colletti, & Toga, Reference Yang, Raine, Narr, Colletti and Toga2009). Moreover, Motzkin and colleagues (Reference Motzkin, Newman, Kiehl and Koenigs2011) showed that diminished neural connectivity in certain regions of the brain is a robust neural correlate of psychopathy. Another study found reduced amygdala–orbitofrontal connectivity during moral judgments in youth with disruptive behavior disorders and psychopathic traits (Marsh et al., Reference Marsh, Finger, Fowler, Jurkowitz, Schechter, Yu and Blair2011). As such, these studies demonstrate that structural volume, structural connectivity and functional connectivity (e.g., during resting state) measures can potentially serve as putative brain endophenotypes of antisocial behavior.
Along these lines, Buckholtz and colleagues (Reference Buckholtz, Callicott, Kolachana, Hariri, Goldberg, Genderson and Meyer-Lindenberg2008) showed that genetic variation in the MAOA gene modulates ventromedial prefrontal circuitries, which in turn contribute to individual differences in human personality. Another study examining the impact of MAOA on brain structure and function found that the low-expression allele (MAOA-L) was linked to limbic volume reductions, amygdala hyper-responsiveness, and blunted reactivity of the prefrontal cortex (Meyer-Lindenberg et al., Reference Meyer-Lindenberg, Buckholtz, Kolachana, Hariri, Pezawas, Blasi and Egan2006).
Last, it is important to point out that neuroimaging studies – like their GWA counterparts – have begun to recognize and appreciate the importance of consortia in the creation of new knowledge. One of the – if not the – largest such consortium is the ENIGMA Consortium. ENIGMA is a large-scale collaborative analysis of neuroimaging and genetic data with the stated mission to explore the shared genetic etiology between various traits and phenotypes. Interested readers can find more information on ENIGMA here: http://enigma.ini.usc.edu.
Epigenetics
We would be remiss if we did not mention the vast potential surrounding epigenetics research. Around the time the human genome was mapped in the early 2000s, researchers began to confront the startling reality that there simply were not enough genes to go around. In other words, the human genome does not contain enough unique information to code for all the complexity and variation that we observe among human outcomes. Thus, it must be that genes interact with other genes, with environments, and perhaps even with some other “higher-order” property in the cell. That higher-order property has come to be known as the epigenome and scholars are now beginning to understand that mechanism. Briefly, the epigenome is responsible for turning genes “on” and turning them “off.” As you might imagine, the biological processes can be complex, but the short of it is this: the epigenome controls which genes are expressed, thereby allowing them to have an influence (or not) on protein production.
But these points are not sufficient to explain why epigenetics has become, as some have put it, “the currently fashionable response to any question to which you do not know the answer.” (Smith, Reference Smith2011, p. 539). Social scientists’ enthusiasm for epigenetics seems to be grounded in the possibility that it could reinvigorate research into environmental influences. But note that the environment captures everything that is not the genome. This might include biological factors such as prenatal environments and larger, social environments like neighborhood conditions. Because of the vast complexity involved, we encourage caution when conducting and interpreting epigenetics results.
Conclusion
The development of BG modeling has been a boon to the study of human variation across a wide range of phenotypes. Studies of human aggression and violence are no different. There is now a large body of evidence revealing that genetic and environmental influences play a role in the etiology of these phenotypes. Thus, the goal of this chapter was to introduce the reader to the two major strands of research that have, to date, delineated this body of evidence. As can be seen, the two approaches to BG modeling differ in their goals: variance decomposition models typically focus on estimating the degree to which variation in P is attributable to G and E, while gene finding models seek to identify the specific genetic variants that may underlie that variation in P.
But the BG literature is not nearly as bifurcated as our review may seem to indicate. Indeed, we presented the material in this chapter in a way that we thought would make it more tractable. This should not be taken as an indication that BG researchers who estimate variance decomposition models are at odds with those who estimate gene finding models. On the contrary, BG scholars are notable for their versatility and for their interests in finding the best way to estimate the impact of the various contributing factors to variance in P. The challenges to doing so, however, are formidable. Variance decomposition models are not without their limitations. Gene findings strategies, too, have come under fire for a host of substantive and statistical shortcomings. We have highlighted a few of the most important limitations and areas in need of improvement for both approaches. There is still much work to be done.
Introduction
Imaging techniques have consistently been used to demonstrate abnormalities of the brain in antisocial, aggressive, and violent individuals. These irregularities occur in several regions of the brain. Deficiencies begin in the prefrontal cortices, traverse back to the cingulate cortex, and to the temporal lobe, including the limbic system (i.e., amygdala and hippocampus). Not only do these abnormalities span various structures, but they manifest in various respects, including structural abnormalities (e.g., decreased tissue volumes), functional abnormalities (e.g., decreased neuronal activations), and even neurotransmitter performance (e.g., decreased serotonin functionality).
Researchers use different imaging techniques for measuring structural and functional characteristics of the brain. Structural studies most commonly use magnetic resonance imaging (MRI). As the name suggests, this technique uses a large magnet and the electrical current of the spinning protons in the brain to generate an anatomical image of regional or whole-brain tissue volume. Functional studies that measure brain activity most often use functional magnetic resonance imaging (fMRI), positron-emission tomography (PET), or single-photon emission computerized tomography (SPECT). fMRI scans utilize a large magnet and can produce an image of activity in the brain by measuring the changes in blood oxygen levels. Both PET and SPECT scans use radioactive tracers that are injected into the blood stream in material such as glucose. When the glucose is metabolized by active brain tissue, the scanner detects the nuclear material and creates an image of active and inactive brain regions. Functional imaging techniques are also used to measure neurotransmitter activity by looking at activity at the receptor sites and the changes in activity following administration of neurotransmitter manipulating drugs.
Researchers use these various brain imaging techniques to study participants who are at rest, performing a particular task, or even responding to a particular stimulus. Researchers conduct imaging studies on various populations as well, including not only healthy controls, but also other community samples, offender populations, and even psychiatric populations. The differences between groups presented speak to the deficits that characterize antisocial and aggressive populations.
Here, we describe the structural, functional, and neurochemical qualities of antisocial and aggressive individuals in the areas of the brain that show marked deficits: the frontal and prefrontal regions, the cingulate cortex, and the limbic system – namely, the amygdala and hippocampus. The majority of research focuses on finding significant differences in various imaging techniques between assorted violent and nonviolent groups. The results of these brain imaging studies have contributed greatly to the understanding of antisocial, aggressive, and violent behavior.
The Frontal Lobe and Prefrontal Cortex
The frontal lobe is a region of the brain consisting of a large amount of brain tissue anterior to the large brain fissure known as the central sulcus. The frontal lobe can be divided into sub regions, primarily the motor cortex, which controls movements, and the prefrontal cortex, which controls cognitive processes or what has been termed executive functioning. The prefrontal cortex can then be divided into smaller sub-regions, including the dorsolateral prefrontal cortex (DLPFC) and the orbitofrontal cortex (OFC; Kolb & Whishaw, Reference Kolb and Whishaw2009). Neuroimaging research has widely focused on the structural, functional, and neurochemical characteristics of the frontal lobes and their association to violence, aggression, and antisocial behavior.
Structural
The most common method of imaging brain structure is magnetic resonance imaging (MRI) and has been used by researchers to examine tissue volume in different areas of the brain. While some MRI studies did not find any significant differences in the frontal lobe volume of violent offenders diagnosed with a mental illness compared to controls (Barkataki, Kumari, Das, Taylor, & Sherma, Reference Barkataki, Kumari, Das, Taylor and Sharma2006; Dolan, Deakin, Roberts, & Anderson, Reference Dolan, Deakin, Roberts and Anderson2002), many studies using MRI techniques have shown volumetric differences in the frontal lobe tissue of violent offenders and antisocial, psychopathic, and aggressive individuals compared to controls – particularly regarding cortical thinning and gray matter reduction in the prefrontal cortex (Calzada-Reyes et al., Reference Calzada-Reyes, Alvarez-Amador, Galán-García, Valdés-Sosa, Melie-García, Alemán-Gómez, del Carmen Iglesias-Alonso and Fitzgerald2015; Narayan et al., Reference Narayan, Narr, Kumari, Woods, Thompson, Toga and Sharma2007; Raine, Lencz, Bihrle, La Casse, & Colletti, Reference Raine, Lencz, Bihrle, LaCasse and Colletti2000; Raine, Yang, Narr, & Toga, Reference Raine, Yang, Narr and Toga2009; Tiihonen et al., Reference Tiihonen, Rossi, Laakso, Hodgins, Testa, Perez and Frisoni2008).
A subgroup of particularly violent offenders was individuals diagnosed with psychopathy. Psychopaths are characterized by both affective and behavioral symptoms including lack of empathy or guilt and impulsivity, which may contribute to their high rates of violent recidivism (Hare, Mcpherson, Forth, & Kazdin, Reference Hare, Mcpherson, Forth and Kazdin1998; Harris, Rice, & Cormier, Reference Harris, Rice and Cormier1991; Kiehl, Kiehl, & Hoffman, Reference Kiehl, Kiehl and Hoffman1991). Yang et al. (Reference Yang, Raine, Lencz, Bihrle, LaCasse and Colletti2005) used MRI scans of participants who were rated as high in psychopathy to look at any structural differences compared with non-psychopaths. The psychopath group was divided into those that had been caught for their crimes (the unsuccessful psychopaths) and those that had not been caught (the successful psychopaths). Results indicated that those who were high in psychopathy were characterized by reduced gray matter volume in the prefrontal region. Furthermore, while unsuccessful psychopaths had even lower prefrontal gray matter volume compared to both controls and successful psychopaths, the successful psychopaths did not differ from controls. These structural imaging findings suggest that prefrontal deficits may lead to poor decision making.
In a later study, Howner et al. (Reference Howner, Eskildsen, Fischer, Dierks and Wahlund2012) used MRI to compare the cortical thickness in the frontal lobes of psychopathic offenders, offenders with antisocial personality disorder (ASPD) and nonoffender healthy controls. ASPD is characterized by a pattern of neglect for others’ rights, and often includes impulsive and aggressive behavior. While researchers did not find any significant difference in the cortical thickness of the frontal lobes between the ASPD offenders and psychopathic offenders, they did report a tendency towards a thinner frontal cortex in psychopaths when compared to healthy controls. In another study that compared both violent and nonviolent offenders to controls, Calzada-Reyes et al. (Reference Calzada-Reyes, Alvarez-Amador, Galán-García, Valdés-Sosa, Melie-García, Alemán-Gómez, del Carmen Iglesias-Alonso and Fitzgerald2015) found no differences in frontal lobe volume within the violent groups comparing psychopaths to non-psychopaths; however, when compared to controls, the violent groups showed reduced gray matter in the orbitofrontal gyrus region of the prefrontal cortex. Violent psychopaths had reduced gray matter volume in the right superior orbitofrontal gyrus and violent non-psychopaths had reduced gray matter in the left middle frontal gyrus when compared to controls. This indicates that violent behavior exhibits a specific manifestation in the frontal regions, Furthermore, psychopathy compared to ASPD presents differently in frontal lobe structures.
Neuroimaging literature has also associated antisocial and aggressive behavior to structural deficits in regions of the prefrontal cortex, including the OFC (Antonucci et al., Reference Antonucci, Gansler, Tan, Bhadelia, Patz and Fulwiler2006; Gansler et al., Reference Gansler, McLaughlin, Iguchi, Jerram, Moore, Bhadelia and Fulwiler2009; Raine, Yang, Narr, & Toga, Reference Raine, Yang, Narr and Toga2009; Tiihonen et al., Reference Tiihonen, Rossi, Laakso, Hodgins, Testa, Perez and Frisoni2008,), and DLPFC (Hirono, Mega, Dinov, Mishkin, & Cummings, Reference Hirono, Mega, Dinov, Mishkin and Cummings2000; Kola et al., Reference Kolla, Matthews, Wilson, Houle and Michael Bagby2015; Kumari et al., Reference Kumari, Aasen, Taylor, Ffytche, Das, Barkataki and Sharma2006; Tiihonen et al., Reference Tiihonen, Rossi, Laakso, Hodgins, Testa, Perez and Frisoni2008). Studies that examined the relationship between structural deficits in the OFC have produced mixed results. Antonucci et al., (Reference Antonucci, Gansler, Tan, Bhadelia, Patz and Fulwiler2006) conducted a study using MRI to detect structural differences in relation to aggression and impulsivity of non-psychotic psychiatric patients. Results demonstrated a relationship between asymmetrical volume of the OFC and aggressive behavior, such that the greater the right OFC volume compared to the left, the greater the lifetime history of aggressive behavior. Motor impulsivity, a factor in aggressive behaviors, was also found to be significantly positively correlated with bilateral OFC volume. Gansler et al., (Reference Gansler, McLaughlin, Iguchi, Jerram, Moore, Bhadelia and Fulwiler2009) in another MRI study, however, found no relationship between aggression and right OFC, but did find a significant negative relationship between left OFC volume and aggression in psychiatric patients but not healthy controls. Still another MRI study of violent offenders at a forensic hospital found violent offenders to have reduced gray matter volume bilaterally in the OFC as well as the frontopolar cortex (Tiihonen et al., Reference Tiihonen, Rossi, Laakso, Hodgins, Testa, Perez and Frisoni2008). In a longitudinal study, Trazepacz et al. (Reference Trzepacz, Yu, Bhamidipati, Willis, Forrester, Tabas and Saykin2013) was able to use MRI scanning to measure the effects of brain atrophy over time and its effect on aggression. They found that atrophy of the OFC and DLPFC overtime was associated with increases in aggression and agitation in Alzheimer’s and dementia patients with a history of aggressive behavior. These mixed results may demonstrate that structural abnormalities may be contingent on not only aggression and violence, but also mental status (psychotic versus non-psychotic populations).
Functional
In addition to structural studies, a variety of brain imaging techniques have been used by researchers to study the functionality of frontal and prefrontal cortices in violent and antisocial individuals. Studies have reported functional deficits at rest in the frontal lobes of antisocial individuals (Goyer et al., Reference Goyer, Andreason, Semple, Clayton, King, Compton-Toth and Cohen1994; Juhasz, Behen, Muzik, Chugani, & Chugani, Reference Juhász, Behen, Muzik, Chugani and Chugani2001; Soderstorm, Tulberg, Wikkelso, & Forman, Reference Soderstrom, Tullberg, Wikkelsö, Ekholm and Forsman2000). For example, Liu, Liao, Jiang, and Wang (Reference Liu, Liao, Jiang and Wang2014) conducted a resting-state fMRI comparing brain activity of patients diagnosed with ASPD compared to healthy controls. They found decreased frontal cortex activity in the ASPD group. These results support earlier findings by Volkow et al. (Reference Volkow, Tancredi, Grant, Gillespie, Valentine, Mullani and Hollister1995), who used PET scans of at-rest violent psychiatric patients with a history of intermittent explosive behavior or ASPD. The results of this study indicated significantly lower prefrontal glucose metabolism in the violent patients compared to controls.
Other functional studies have found mixed results characterizing frontal lobe deficits. Amen, Stubblefield, Carmichael, and Thisted (Reference Amen, Stubblefield, Carmichael and Thisted1996) compared aggressive psychiatric patients to nonaggressive patients using SPECT. Results showed decreased prefrontal cortex activity, but also increased activity in the anteromedial prefrontal cortex. A later PET study of aggressive dementia patients found that, at rest, aggressive patients did not differ from nonaggressive patients in OFC functioning but did show reduced bilateral dorsolateral frontal cortex functioning (Hirono, Mega, Dinov, Mishkin, & Cummings, Reference Hirono, Mega, Dinov, Mishkin and Cummings2000). Functional MRI has also been used to evaluate the connectivity between the frontal lobe and other areas of the brain. Phillipi and colleagues (Reference Philippi, Pujara, Motzkin, Newman, Kiehl and Koenigs2015) used resting-state fMRI to assess the functional connectivity of three primary cortical networks in psychopathic and non-psychopathic adult prison inmates. They found that higher psychopathy scores were associated with lower functional connectivity between the frontal and parietal lobes.
While many functional brain imaging procedures are conducted while participants are at rest, several researchers utilize various tasks during imaging to measure traits characteristics and investigate impairments in the frontal lobe of aggressive individuals (Intrator et al., Reference Intrator, Hare, Stritzke, Brichtswein, Dorfman, Harpur and Machac1997; Joyal et al., Reference Joyal, Putkonen, Mancini-Marïe, Hodgins, Kononen, Boulay, Pihlajamaki, Soininen, Stip, Tiihonen and Aronen2007; Raine et al., Reference Raine, Meloy, Bihrle, Stoddard, LaCasse and Buchsbaum1998; Schiffer et al., Reference Schiffer, Pawliczek, Müller, Forsting, Gizewski, Leygraf and Hodgins2014). Deficits in inhibitory control of impulses have been linked to impulsive violence. Reduced prefrontal activity in impulsive violent offenders has been theorized to explain poor cognitive inhibition of affective impulses (Raine et al., Reference Raine, Meloy, Bihrle, Stoddard, LaCasse and Buchsbaum1998). An externalizing mental disorder that is characterized by highly impulsive aggressive and violent behavior is intermittent explosive disorder (IED; Coccaro, Reference Coccaro2012). In order to better understand the neural mechanisms involved in the disinhibition component of IED, Moeller et al. (Reference Moeller, Froböse, Konova, Misyrlis, Parvaz, Goldstein and Alia-Klein2014) conducted an experiment using fMRI to scan the brains of individuals diagnosed with IED and healthy controls. The scans were taken during a Stroop color-naming task, which required the participant to name the color ink that a color word is printed in, with some of the color words being printed in a noncongruent color. This requires the participant to inhibit the impulse to say the word instead of the color of the ink. Results showed greater activity in the left DLPFC of the IED group compared to controls during errors on the Stroop task. Furthermore, the greater the outward expression of anger, the greater the activity in the left DLPFC during error processing. These results could indicate that left DLPFC hyperactivity could be a biological marker for impulsive aggression and related to inhibitory control. In another study that used a task to engage inhibitory processes while conducting an fMRI scan, patients diagnosed with ASPD performed a go/no-go task. This task requires the participant to engage in a behavior in response to certain stimuli and to stop that behavior when other stimuli are present. Results of this study found that healthy controls showed activation during response inhibition primarily in the right DLPFC and the left OFC, while ASPD individuals showed activation in the medial, superior, and inferior frontal gyri. These findings support neural inhibitory response pattern abnormalities in individuals with externalizing disorders (Völlum et al., Reference Völlum, Richarson, Stirling, Elliott, Dolan, Chaudhry, Del Ben, McKie, Anderson and Deakin2004).
Contrasting results were reported in Schiffer et al. (Reference Schiffer, Pawliczek, Müller, Forsting, Gizewski, Leygraf and Hodgins2014), who conducted an fMRI study with violent offenders and nonoffenders while engaging in the Stroop color-naming task. These researchers found that violent offenders displayed reduced DLPFC activation during error processing compared with controls. Similar results showed reduced activation in the left DLPFC during a go/no-go task (Völlum et al., Reference Völlum, Richardson, McKie, Reniers, Elliott, Anderson, Williams, Dolan and Deakin2010). In this study, ASPD individuals were characterized by decreased glucose metabolism during a PET scan compared with controls. While the present literature does demonstrate abnormality in prefrontal cortex functioning in relation to impulsive violence, results differ in the nature and direction of this relationship, highlighting the need for further research in this area and again indicating differences due to variable mental statuses (IED versus ASPD).
Differences in functional activity in the frontal and prefrontal regions may be closely related to emotionality and the differences in emotional regulation seen in varying mental disorders. Neuroimaging research has demonstrated impaired or dysfunctional emotional (Decety, Michalska, Aitsuki, & Lahey, Reference Decety, Michalska, Akitsuki and Lahey2009; Prehn et al., Reference Prehn, Schlagenhauf, Schulze, Berger, Vohs, Fleischer and Herpertz2013) and cognitive (Joyal et al., Reference Joyal, Putkonen, Mancini-Marïe, Hodgins, Kononen, Boulay, Pihlajamaki, Soininen, Stip, Tiihonen and Aronen2007; Schiffer et al., Reference Schiffer, Pawliczek, Müller, Forsting, Gizewski, Leygraf and Hodgins2014) processes and the combination of deficits in these areas can contribute to antisocial behavior and violence (Birbaumer, Reference Birbaumer, Veit, Lotze, Erb, Hermann, Grodd and Flor2005; Sterzer, Stadler, Krebs, Kleinschmid, & Poustka, Reference Sterzer, Stadler, Krebs, Kleinschmidt and Poustka2005). In a study to investigate the neural abnormalities in criminal psychopaths, a population characterized by affective deficits, Müller et al. (Reference Müller, Sommer, Wagner, Lange, Taschler, Röder, Schuierer, Klein and Hajak2003) conducted a study showing criminal psychopaths and healthy volunteers pictures with both positive and negative affective content while recording fMRI data. A differential pattern of activation emerged between the two groups in frontal lobe regions. During the viewing of negative emotional content, psychopaths showed increased activation of the right prefrontal areas among other limbic structures; however, when psychopaths viewed positive emotional content, reduced activation was seen in the right medial frontal gyrus when compared with controls. Neuroimaging research has also been extended to abnormalities in adolescents. Decety et al. (Reference Decety, Michalska, Akitsuki and Lahey2009) presented images to adolescents diagnosed with aggressive conduct disorder (CD). Conduct disorder is a childhood or adolescent disorder that is marked by a pattern of antisocial behavior that often includes aggression. Participants were shown people in pain or not in pain, and pain caused on purpose or by accident, during fMRI procedure. Again, a differential pattern of activation was found between the aggressive conduct disorder group and controls. When watching pain inflicted intentionally versus by accident, controls showed activation in the lateral OFC, and the superior frontal and medial prefrontal gyrus. The CD group showed only activation in the medial OFC in frontal cortex regions. Together these studies demonstrate that even beginning in adolescence, emotionality and its specific relationship to mental disorder may explain some of the irregularities in neurofunctionality for antisocial and aggressive individuals.
In addition to emotional deficits, cognitive deficits have been expressed via frontal lobe impairment in violent offenders. This was demonstrated in a series of classic early PET studies. Raine, Buchsbaum, and Lacasse (Reference Raine, Buchsbaum and LaCasse1997) conducted PET scans of offenders while they were performing a continuous performance task that had been established to activate the frontal lobes. These offenders had been tried for murder and had pleaded not guilty by reason of insanity. Violent offenders showed reduced glucose metabolism in multiple areas of the prefrontal cortex, including the right OFC, when compared to controls. In a subsequent study, Raine et al. (Reference Raine, Meloy, Bihrle, Stoddard, LaCasse and Buchsbaum1998) separated the murderers into those that had experienced a significant level of psychosocial deprivation and those that had not. They found that only murderers without psychosocial deprivation showed reduced glucose metabolism in the prefrontal cortex compared to murderers with significant psychosocial deprivation. In a third follow-up study, Raine et al. (Reference Raine, Meloy, Bihrle, Stoddard, LaCasse and Buchsbaum1998) divided the same murderers into predatory and affective murderers. These results found that affective murderers showed both reduced prefrontal glucose metabolism and increased subcortical metabolism compared with controls. Predatory murderers did not differ from controls in prefrontal glucose metabolism but showed a similar increase in subcortical metabolism as the affective murderers. The results highlight the high emotional reactivity with poor cognitive control seen in impulsive violent offenders.
Functional neuroimaging has been used to study frontal lobe activity when aggression is experimentally provoked in violent individuals diagnosed with a mental disorder. Using PET imaging during induced autobiographical memories, Spoont, Kuskowski, and Pardo (Reference Spoont, Kuskowski and Pardo2010) compared violent offenders diagnosed with a personality disorder or IED to healthy controls. In results that suggested violent individuals have deficient cognitive control of impulsive aggression, the violent group showed lower frontal activity during angry memories versus neutral memories compared to controls. The violent group also had overall lower frontal activity during both types of memory. These results are supported by an earlier study using a computer task to provoke aggression. Reported results found individuals diagnosed with bipolar disorder or IED showed decreased activation of the OFC during neutral conditions and increased OFC activity during the provoked condition compared with controls (New et al., Reference New, Hazlett, Newmark, Zhang, Triebwasser, Meyerson and Buchsbaum2009).
Neurotransmitters
Neurotransmitters are the chemicals used by the brain to function. These chemicals pass from one brain cell to another signaling a particular response. Recent studies have used functional neuroimaging to report the involvement of neurotransmitters serotonin and dopamine in aggression and violence (Kunz et al., Reference Kunz, Sikora, Krakowski, Convit and Cooper1995; Virkkunen & Linnoila, Reference Virkkunen and Linnoila1993; Schlüter et al., Reference Schlüter, Winz, Henkel, Eggermann, Mohammadkhani-Shali, Dietrich, Heinzel, Decker, Cumming, Zerres, Piel, Mottaghy and Vernaleken2016; Tiihonen et al., Reference Tiihonen, Rautiainen, Ollila, Repo-Tiihonen, Virkkunen, Palotie and Paunio2015). New et al. (Reference New, Hazlett, Buchsbaum, Goodman, Reynolds, Mitropoulou and Siever2002) investigated the role of frontal cortex serotonin in impulsive aggression by using PET scans of patients with impulsive aggression following the administration of a serotonergic stimulus or a placebo. The aggressive patients showed reduced activation in the right lateral OFC as well as the left medial OFC compared with controls when receiving the serotonergic stimulus. These results suggest that impulsively aggressive individuals may have a deficient inhibitory response process to serotonin in frontal lobe regions. In a follow-up study, New et al. (Reference New, Buchsbaum, Hazlett, Goodman, Koenigsberg, Lo and Siever2004) injected fluoxetine, which increases the amount of serotonin being used by blocking its reuptake into the cells, into impulsive aggressive patients who met the criteria for a borderline personality diagnosis. PET scans before and after the injections showed an increase of metabolic activity in the OFC, which in their previous study was hypoactive in aggressive individuals.
Additional evidence of the role of neurotransmitters comes from brain imaging studies that examine monoamine oxidase A (MAO-A), an enzyme in the brain that breaks down serotonin and dopamine, among other neurotransmitters (Bortolato et al., Reference Bortolato, Chen, Godar, Chen and Wu2011). A recent pair of studies examined the role of MAO-A in impulsive violence (Kolla et al., Reference Kolla, Matthews, Wilson, Houle and Michael Bagby2015; Kolla et al., Reference Kolla, Dunlop, Downar, Links, Bagby, Wilson and Meyer2016). Kolla et al. (Reference Kolla, Matthews, Wilson, Houle and Michael Bagby2015) administered a task that measures impulsivity to males with ASPD and a history of violence. A tracer that selectively marks MAO-A was used with PET scans to determine levels of MAO-A active in frontal cortex regions. The investigators reported that significantly lower MAO-A was found in ASPD males compared to controls in several prefrontal cortex regions including the OFC and DLPFC, as well as in the ventrolateral PFC and medial PFC.
The role of the frontal lobe structures in violence, aggression, and antisocial behavior has been widely studied using various neuroimaging techniques. MRI scans have shown structural deficits in antisocial individuals, while fMRI, PET, and SPECT studies indicate functional abnormalities and neurotransmitter deficits in violent individuals as well. Results of these studies have come to varying conclusions. While some of the results are mixed, this may be explained by emotionality and varying mental disorders. Varying mental status may be deemed antisocial, aggressive, and violent (e.g., IED, ASPD, psychopathy); however, the role that emotion plays in these disorders vary, and the disparate neuroimaging results may reflect this difference. Despite these varying results, however, the neuroimaging data compiled lays a solid ground work for future brain imaging studies of antisocial behavior and violence.
The Cingulate Cortex
Like the OFC, DLPFC, and other frontal and prefrontal regions, the cingulate cortex is a brain region often implicated in various executive functioning roles that are associated with aggressive and violent behavior. More specifically, the cingulate cortex has continuously been related to decision making, affective regulation, and impulsivity. Various studies have demonstrated structural, functional, and other abnormalities specific to the cingulate cortex as it relates to antisocial behavior, aggression, and violence. These studies have demonstrated robust findings spreading across both healthy and clinical samples, adolescents and adults, as well as males and females.
Structural
As found in the frontal and prefrontal cortices, structural abnormalities including reduced gray matter and decreased cortical thickness have been observed in the cingulate cortex in antisocial and violent populations. Similar to studies that implicate prefrontal cortices in antisocial behavior and specifically psychopathy, Yang et al. (Reference Yang, Raine, Colletti, Toga and Narr2009) performed MRIs on psychopaths and healthy controls to determine differences in gray matter. This study found reduced gray matter in the posterior cingulate gyrus among other areas in psychopaths compared to healthy controls. Using fMRI to measure, Ly et al. (Reference Ly, Motzkin, Philippi, Kirk, Newman, Kiehl and Koenigs2012) separated adult male offenders into psychopathic and non-psychopathic groups. Offenders with increased psychopathic traits had significant cortical thinning in the anterior cingulate cortex. Jiang et al. (Reference Jiang, Guo, Zhang, Gao, Wang, Situ and Huang2015) found similar results in adolescent participants aged 14 to 16. This study included both male and female participants with adolescent-onset CD. It was found that the CD group showed decreased cortical thickness in the posterior cingulate cortex. Additionally, a reduced amount of cortical folding was found in the anterior cingulate cortex. Increased cortical folding has often been associated with increased intelligence (due to the increased amount of surface area). Together, these studies implicate structural abnormalities in the cingulate cortex in antisocial behavior not only for adult males, but also for adolescents, as well as for female participants, who are often overlooked in antisocial research.
Research has also expanded literature on antisocial individuals who are comorbid with other disorders, namely borderline personality disorder (Bertsch et al., Reference Bertsch, Grothe, Prehn, Vohs, Berger, Hauenstein and Herpertz2013) and schizophrenia (Kumari et al., Reference Kumari, Uddin, Premkumar, Young, Gudjonsson, Raghuvanshi and Das2014). Bertsch et al. (Reference Bertsch, Grothe, Prehn, Vohs, Berger, Hauenstein and Herpertz2013) conducted MRIs on adult male offenders convicted for capital violent crimes. The offenders were separated into those with borderline personality disorder traits and those with psychopathic traits. Results found that increased psychopathic traits were associated with reduced gray matter in the posterior cingulate cortex. Interestingly, these significant reductions were not as marked in those with borderline personality disorder traits. Similarly, Kumari et al. (Reference Kumari, Uddin, Premkumar, Young, Gudjonsson, Raghuvanshi and Das2014) separated participants with diagnoses of ASPD or schizophrenia with serious violence, or schizophrenia without serious violence. Reduced gray matter in the anterior cingulate was found for both the ASPD disorder group and the violent schizophrenia group compared to healthy controls. This marked decrease was not found in the schizophrenia without serious violence group.
Akin to the structural neuroimaging results of prefrontal and frontal regions, disparate findings (i.e., reduced gray matter) may be contingent on violent status (violent versus nonviolent) and mental status (psychopathy versus borderline personality disorder).
Functional
In addition to structural deficiencies, functional abnormalities present in the cingulate cortex are also related to antisocial, aggressive, and violent behavior. Similar to studies regarding the frontal and prefrontal cortices, functional studies implicating the cingulate cortex demonstrate decreased neuronal activity. Like the structural deficits found in the cingulate cortex, the functional discrepancies are found in adolescents and adults, as well as clinical and healthy control samples.
Research continues to find relationships between decreased neuronal activity as it relates to psychopathy and emotionality in particular. Kiehl et al. (Reference Kiehl, Smith, Hare, Mendrek, Forster, Brink and Liddle2001) separated offenders into psychopathic and non-psychopathic groups. Psychopathic offenders showed decreased emotional-related activity in the anterior and posterior cingulate gyrus, among other areas. Furthermore, these results are not limited to adult populations. Adolescent male participants aged from 9 to 15 who met DSM criteria for CD demonstrated a marked deactivation in the anterior cingulate cortex compared to healthy controls via an fMRI after viewing negative-affective pictures (Sterzer et al., Reference Sterzer, Stadler, Krebs, Kleinschmidt and Poustka2005).
Similar to the functional connectivity deficiencies found in the frontal lobes (Philippi et al., 2016), functional deficiencies in antisocial and aggressive individuals are not limited to a single area of the brain, but also manifest as connectivity deficits between the cingulate cortex and other brain regions. Ly et al. (Reference Ly, Motzkin, Philippi, Kirk, Newman, Kiehl and Koenigs2012) found that psychopathic offenders demonstrated reduced functional connectivity between the insula and the anterior cingulate cortex relative to their non-psychopathic counterparts. Analogous results demonstrated that increased psychopathic traits were associated with decreased connectivity between the intraparietal sulcus and the anterior cingulate cortex (Philippi et al., 2016).
Studies have also indicated that it is not just psychopathy as a whole that relates to decreased functional activity in the cingulate cortex. Researchers continue to find that reduced neuronal activity is specifically related to the emotional blunting associated with psychopathy (Prehn et al., Reference Prehn, Schlagenhauf, Schulze, Berger, Vohs, Fleischer and Herpertz2013; Searo-Cardoso et al., Reference Seara-Cardoso, Viding, Lickley and Sebastian2015). Prehn et al., Reference Prehn, Schlagenhauf, Schulze, Berger, Vohs, Fleischer and Herpertz2013 separated a group of violent criminal offenders into those with emotional hyporeactivity or those with emotional hyperreactivity. While all the offenders met the DSM criteria for ASPD, only those with below-average emotional reactivity – or hyporeactivity – showed decreased activity in the anterior cingulate cortex. This same pattern is seen in nonoffender populations. Seara-Cardoso et al. (Reference Seara-Cardoso, Viding, Lickley and Sebastian2015) measured psychopathic traits in a healthy sample of adult males with no history of mental illness. MRI results demonstrated that those with increased affective-interpersonal psychopathic traits demonstrated decreased neural responses in the cingulate cortex after seeing others pain. Interestingly, those with increased lifestyle-antisocial psychopathic traits showed increased responses in the cingulate cortex. These disparate findings indicate that the emotional component of antisociality is particularly related to decreased activity in the cingulate cortex. This specificity could possibly explain studies that have shown no differences between psychopathic and nonpsychopathic individuals as related to cingulate cortex abnormalities (Dvorak-Bertsch et al., Reference Dvorak-Bertsch, Sadeh, Glass, Thornton and Newman2007).
Neurotransmitters
Like the research that has used functional neuroimaging in the frontal and prefrontal cortices to implicate neurotransmitters in matters of antisociality and aggression, studies have found similar results in the cingulate cortex. In a sample of adult male and female participants who met criteria for IED, PET scans demonstrated that those with IED showed decreased serotonin transporters in the anterior cingulate cortex compared to healthy controls (Frankle et al., Reference Frankle, Lombardo, New, Goodman, Talbot, Huang and Siever2005). Völlum et al. (Reference Völlum, Richardson, McKie, Reniers, Elliott, Anderson, Williams, Dolan and Deakin2010) found that in both healthy and ASPD patients, activation of the cingulate cortex was increased during an inhibition task after a serotonin receptor agonist was administered. Parallel to neurotransmitter behavior in the frontal and prefrontal cortex, these results indicate that deficient serotonin processing in the cingulate cortex is associated with antisocial and aggressive behavior.
Limbic System
Like the cingulate cortex, the limbic system, a medial part of the temporal lobe, is consistently implicated in antisocial, aggressive, and violent behavior, especially as it relates to emotional regulation. The amygdala and the hippocampus are two structures in the limbic system that are particularly implicated in antisocial behavior. The amygdala is often related to the processing of emotion from sensory stimuli. The amygdala is strongly and bidirectionally connected to the hippocampus, which is implicated in memory. Together, these structures are responsible for fear conditioning, threat response, and other emotionally modulated memory processes.
Akin to imaging studies of the prefrontal, frontal, and cingulate cortex, imaging studies of the limbic system also demonstrate robust findings of structural, functional, as well as neurochemical abnormalities in antisocial, aggressive, and violent individuals. Additionally, affective contributions influence how these abnormalities manifest in antisociality.
Structural
Imaging studies consistently demonstrate structural abnormalities in the limbic system for antisocial, aggressive, and violent individuals, but disparate from the prefrontal, frontal, and cingulate cortex findings, structural abnormalities of the limbic system are not limited to reduced gray matter and decreased cortical thickness. Antisociality is also characterized by aberrant morphology or shape in limbic structures, namely the hippocampus and amygdala.
Several studies have indicated that increased psychopathy is associated with abnormal gray matter in limbic structures (Boccardi et al., Reference Boccardi, Frisoni, Hare, Cavedo, Najt, Pievani and Tiihonen2011; Ermer et al., Reference Ermer, Cope, Nyalakanti, Calhoun and Kiehl2012); however, results are mixed. Parallel to volumetric decreases in other brain structures, Ermer et al. (Reference Ermer, Cope, Nyalakanti, Calhoun and Kiehl2012) used MRI to measure gray matter in adult male offenders. Results demonstrated that as psychopathy scores increased, gray matter volume and tissue density decreased in the amygdala and hippocampus. In contrast, Boccardi et al. (Reference Boccardi, Frisoni, Hare, Cavedo, Najt, Pievani and Tiihonen2011) found increased amygdala volume in psychopathic violent offenders; however, this study was cautious to remove any offenders with schizophrenia spectrum disorders, including schizotypal personality disorder. Barkataki et al. (Reference Barkataki, Kumari, Das, Taylor and Sharma2006) may shed light on these disparate findings. In this study, participants were separated into ASPD, violent schizophrenia, nonviolent schizophrenia, and healthy control groups. Participants underwent MRI procedures and results indicated that the ASPD and the violent schizophrenia group demonstrated reduced brain volume overall compared to healthy control. Furthermore, the violent schizophrenia group had decreased hippocampal volumes compared to ASPD and control groups. Together, these results indicate that schizophrenia spectrum disorders can differentially impact structural abnormalities in antisocial individuals.
Pardini et al. (Reference Pardini, Raine, Erickson and Loeber2014) used longitudinal methodologies to expand on volumetric disparities of the limbic system in antisociality, aggression, and violence. In this study, men who had been followed since childhood underwent MRI procedures as adults. Decreased amygdala volume was found in men who had demonstrated aggressive and psychopathic traits since childhood and adolescence. This study also indicated that decreased amygdala volume is a risk factor for future aggressive and violent behavior. These results show robust evidence for structural deficits in antisocial individuals even beginning in adolescence.
As mentioned, imaging studies have found not only volumetric disparities, but also morphologic abnormalities in limbic structures (Boccardi et al., Reference Boccardi, Ganzola, Rossi, Sabattoli, Laakso, Repo-Tiihonen and Tiihonen2010; Coccaro et al., Reference Coccaro, Lee, McCloskey, Csernansky and Wang2015). In one such study, violent offenders and healthy controls underwent MRI to measure volumetric differences. While volumes did not significantly differ between groups, structurally the surface shape of the hippocampus differed between violent offenders and healthy controls. Coccaro et al. (Reference Coccaro, Lee, McCloskey, Csernansky and Wang2015) found similar results, such that participants with IED demonstrated significantly different surface shape deformations in the hippocampus and the amygdala compared to their healthy counterparts.
Functional
Similar to structural imaging studies, research has continued to find functional abnormality in the limbic system. Additionally, similar to results found in the cingulate cortex, many of the anomalies are modulated by affective or emotional involvement. Hypoactivity or hyperactivity of limbic structures, mainly the amygdala, is contingent on differences in affective manifestations of various antisocial disorders.
Similar to studies that measure functional deficiencies in the frontal, prefrontal, and cingulate cortex, fMRI studies have found decreased functionality in the limbic system in those who exhibit antisocial behavior. Kiehl et al. (Reference Kiehl, Smith, Hare, Mendrek, Forster, Brink and Liddle2001) separated offenders into psychopathic and non-psychopathic groups. Psychopathic offenders showed decreased emotional-related activity in the amygdala and hippocampus. Fairchild et al. (Reference Fairchild, Hagan, Passamonti, Walsh, Goodyer and Calder2014) found similar results in adolescent females. Adolescent females with CD and healthy controls underwent similar procedures of viewing facial expressions during fMRI. CD was significantly correlated to amygdala activation, such that as CD increased, amygdala activation decreased.
While many studies demonstrate this decreased activity in the amygdala, these results are strongly contingent on the emotional characteristics related to different types of antisociality. Studies have also found increased activity in the limbic system in response to negative emotions (Hyde et al., Reference Hyde, Byrd, Votruba-Drzal, Hariri and Manuck2014; McCloskey et al., Reference McCloskey, Phan, Angstadt, Fettich, Keedy and Coccaro2016; Osumi et al., Reference Osumi, Nakao, Kasuya, Shinoda, Yamada and Ohira2012). These conflicting results demonstrate that antisocial behavior, aggression, and violence can be correlated to hypoactivity or hyperactivity dependent on the type of antisociality.
Hyde et al. (Reference Hyde, Byrd, Votruba-Drzal, Hariri and Manuck2014) demonstrated how limbic activity can increase or decrease dependent on the type of antisocial disorder (e.g., ASPD or psychopathy). Hyde et al. (Reference Hyde, Byrd, Votruba-Drzal, Hariri and Manuck2014) conducted fMRI procedure on a community sample of both male and female adults. ASPD traits were positively related to amygdala reactivity, whereas psychopathic traits were negatively related to amygdala reactivity. This study found that negative emotion is what separates the difference between ASPD and psychopathy such that the increased negative emotionality (present in ASPD, but not psychopathy) is responsible for amygdala activation. Osumi et al. (Reference Osumi, Nakao, Kasuya, Shinoda, Yamada and Ohira2012) found similar results in healthy male undergraduate students such that increased psychopathic traits correlated to decreased activity in the amygdala following a frustrating task. The authors suggest that this may indicate that psychopaths have decreased neural response to reactive aggression.
Another study that demonstrated the differences in antisociality both in behavior and in neural activity of the limbic system was conducted by McCloskey et al. (Reference McCloskey, Phan, Angstadt, Fettich, Keedy and Coccaro2016). In this study, participants were patients meeting DSM criteria for intermittent explosive disorder and healthy controls. Participants were shown human facial expressions during the fMRI procedure. Results showed that the intermittent explosive disorder group showed increased amygdala activation in response to angry faces compared to healthy controls; however, this difference did not occur after being shown happy faces. Together, these studies indicate that manifestations of antisocial behavior are different behaviorally and neurologically and that these disparities are due to how negative emotions are processed. Psychopathic characteristics demonstrate decreased emotionality and this is expressed by decreased activity in the limbic structures. In contrast, other disorders, including ASPD and intermittent explosive disorder, show increased emotionality and this corresponds to increased activity in the limbic structures.
Neurotransmitters
Similar to studies that used functional neuroimaging in other regions of the brain to associate neurotransmitters to matters of antisocial behavior, research has found similar results in the limbic system. Osumi et al. (Reference Osumi, Nakao, Kasuya, Shinoda, Yamada and Ohira2012) found that psychopathic traits were related to decreased dopaminergic functionality between the amygdala and another area of the brain – namely the striatum. Kolla et al. (Reference Kolla, Dunlop, Downar, Links, Bagby, Wilson and Meyer2016) found similar results in the hippocampus for a group of ASPD participants who had a history of violence. ASPD participants demonstrated decreased MAO-A functionality in the hippocampus, again indicating deficiencies in dopaminergic functioning.
The limbic structures, like other regions of the brain, also show marked abnormalities, and, like other areas of the brain, these abnormalities manifest in various ways dependent on mental status and emotionality. It is important for researchers to consider how different disorders process emotions in order to relate this back to the disparate findings in the structural and functional processes of various brain regions. Neuroimaging data continues to play an integral role in understanding antisociality, aggression, and violence. Neuroimaging allows researchers to better understand behavioral manifestations of aggression via the structural and functional deficiencies that are apparent throughout various regions of the brain, including frontal and prefrontal cortices, the cingulate cortex, and even the temporal lobes, namely the limbic system. This data also allows researchers to expand research to implications for the law and policy, for prevention efforts of future aggression, and for interventions for mental disorders marked by antisocial and aggressive behavior.
Policy, Prevention, and Intervention
There are significant implications for the law as well as for prevention and interventions for aggressive and violent behavior. The legal system has used imaging techniques not only as evidence during court proceedings but also in various briefing documents (Yang, Glenn, & Raine, Reference Yang, Glenn and Raine2008). Oftentimes, brain imaging results are presented as evidence for mitigating circumstances. Several court cases have pointed to structural or functional deficits as rationalizations for antisocial and aggressive behavior (e.g., John Hinckley, Jr.’s structural deficiencies). In Baird and Fugelsang (Reference Baird and Fugelsang2004), researchers use brain imaging studies to weigh in on development and mental culpability as it relates to age. While it is important to consider empirical evidence within the legal system, it is also important to remember that, as evidenced throughout the literature, there are mixed results. These mixed results are continually linked to the different manifestations of emotionality in various mental disorders. When considering policy or legal consequences, one must take into account that antisocial behavior is not a single and invariable concept. While many disorders are characterized by antisocial behavior, as well as aggressive and violent behavior, neuroimaging studies continue to show that these antisocial and aggressive disorders do not manifest in the same way, both cognitively and behaviorally. This is also important to consider in respect to prevention and intervention for antisociality and aggression.
Brain imaging research lays solid groundwork for prevention efforts and intervention methods to reduce aggression and violence. This is evident especially as related to the functional imaging studies that implicate neurotransmitters in antisociality. As indicated throughout the literature, there are various anomalies in serotonin and dopaminergic functionality. This may imply that neurochemical interventions may ameliorate aggressive and violent behavior. Research has already found that using neurochemical intervention can impact impulsively aggressive populations (New et al., Reference New, Buchsbaum, Hazlett, Goodman, Koenigsberg, Lo and Siever2004).
In light of research that indicates brain abnormalities in children and adolescents, research has also advocated for proactive and early intervention to prevent future aggressive and violent behavior (Anderson & Kiehl, Reference Anderson and Kiehl2014). These early interventions can include cognitive behavioral methods as well as enhancing internal motivations to combat treatment resistance (Bayliss, Miller, & Henderson, Reference Bayliss, Miller and Henderson2010). Early interventions are especially important when considering that, as a whole, interventions for psychopathy in adults only show modest success (Anderson & Kiehl, Reference Anderson and Kiehl2014). When considering neuroplasticity of children and adolescents, it very important to target those at risk for aggression and violence as early as possible to allow increased functionality in the brain (Anderson & Kiehl, Reference Anderson and Kiehl2014).
Affective factors, especially emotional regulation, are key components related to brain abnormalities and antisociality. Research has indicated that various intervention methods can enhance emotional regulation. In addition to cognitive behavioral therapies, integrative methods that include body-mind training (body relaxation, mental imagery, mindfulness) have shown positive results for emotional regulation (Tang et al., Reference Tang, Yang, Leve and Harold2012). Another study found that biofeedback interventions have shown positive results in antisociality (Howard, Schellhorn, & Lumsden, Reference Howard, Schellhorn and Lumsden2013). The patient in this study demonstrated a history of impulsivity, hostility, and aggression and, after several weeks of biofeedback intervention, the patient had positive results in self-regulating his emotions.
Neuroimaging provides a biological understanding of antisocial, aggressive, and violent behavior. Various imaging techniques allow researchers to consider both structural and functional manifestations of antisociality and aggression in several structures of the brain. An important implication of neuroimaging evidence is that antisocial, aggressive, and violent individuals are characterized by abnormalities in the brain. Furthermore, many of these deficiencies are linked to emotionality, and while several mental disorders are distinguished by antisocial behavior, they demonstrate differences in emotional processing, which may lead to variable manifestations of aggressive and violent behavior. Neuroimaging research also lays a basis for implications related to policy and legal standards, prevention, and intervention for antisocial individuals who are characterized by aggression and violence.
Introduction
There is growing evidence that the development of aggression is a multilevel process that cannot be entirely explained by either biological or social factors alone. Instead, biological and social factors likely contribute both independently and in interaction with one another in the development of aggression and antisocial behavior (Raine, Reference Raine2002b, Reference Raine2013). Biological domains contributing to antisocial behavior include genetics, neuropsychology, and psychophysiology. Research in these areas has demonstrated the presence of biosocial interactions, whereby biological factors interact with social factors to predict antisocial behavior (Raine, Reference Raine2002b, Reference Raine2013). Several theoretical models have been proposed to explain the pattern of biosocial interactions observed in criminological research; these include the dual-risk model, the social push perspective, the biological protective factors approach, mediation models, and the differential susceptibility theory.
Here, I review research examining the biological basis of aggression and antisocial behavior. This chapter primarily focuses on biosocial research involving psychophysiological risk factors. Psychophysiological risk factors are relatively easy and less costly to measure than other biological risk factors (Gao et al., Reference Gao, Glenn, Peskin, Rudo-Hutt, Schug, Yang, Raine, Gadd, Karstedt and Messner2012), and therefore may be of particular interest to socially oriented researchers who would like to incorporate biological measures into their research. This chapter begins with a review of research involving psychophysiological risk factors for aggression and antisocial behavior. This chapter then discusses psychophysiological research in the context of the dual-risk, social push, protective factors, differential susceptibility, and mediation models of biosocial interactions. I conclude with a discussion of how biosocial interactions could contribute in the future to improving interventions for aggression and antisocial behavior.
Psychophysiological Risk Factors for Antisocial Behavior
Psychophysiology is the study of cognition, behavior, and emotions as revealed through bodily states (Hugdahl, Reference Hugdahl2001). Psychophysiological indices are noninvasive to record and capture nearly immediate physiological changes. Psychophysiological measures are often recorded at rest, but may also be recorded in response to experimental stimuli, such as fear conditioning tasks or stress tasks. Heart rate and skin conductance are two of the best-studied psychophysiological risk factors for aggression.
Heart Rate
Heart rate is controlled by the sympathetic and parasympathetic branches of the autonomic nervous system. Low resting heart rate has been suggested as the “best-replicated biological correlate of antisocial behavior in child and adolescent populations” (Ortiz & Raine, Reference Ortiz and Raine2004, p. 159). A recent meta-analysis of 114 reports and 115 independent effect sizes yielded of random effects summary effect size of d = −0.20 (SE = 0.04, p < 0.001) for the relationship between resting heart rate and antisocial behavior (Portnoy & Farrington, Reference Portnoy and Farrington2015). Under the random effects model, the summary effect size was unmoderated by sex, type of antisocial behavior (including violence and aggression), age group, or recruitment (clinical/institutional vs. community). The effect size was also unmoderated by study design (prospective vs. concurrent). This suggests that a low heart rate could precede the onset of antisocial behavior, rather than result from antisocial behavior. Consistent with this, an analysis of 411 males participating in the Cambridge Study in Delinquent Development found that resting heart rate at age 18 predicted convictions for violence up to age 50 (Jennings, Piquero, & Farrington, Reference Jennings, Piquero and Farrington2013). Results were largely unchanged after controlling for numerous covariates, including sports participation, impulsivity, binge drinking, and body mass index. These results suggest that the relationship between resting heart rate and antisocial behavior is unlikely to be the result of confounding variables.
This raises the important question of why low resting heart rate is associated with higher levels of antisocial behavior and aggression. Although the mechanism underlying the low resting heart rate–antisocial behavior relationship is not yet fully understood, low resting heart rate is hypothesized to be associated with increased levels of antisocial behavior, because low autonomic nervous system arousal may reflect a relative lack of fear, which could facilitate antisocial behavior by impeding early fear conditioning to socializing punishments and reducing fear of the negative consequences of the antisocial act (Raine, Reference Raine2002a, Reference Raine2013). Alternatively, reduced autonomic nervous system arousal could be an unpleasant physiological state, leading those with low resting heart rates to engage in stimulating behaviors, including antisocial behaviors, in order to increase their level of arousal to a more optimal level (Quay, Reference Quay1965; Raine, Reference Raine2002a, Reference Raine2013). Recent studies have provided support for a sensation-seeking explanation of the low heart rate–antisocial behavior relationship. A study of 335 adolescent boys participating in the Pittsburgh Youth Study found that sensation-seeking but not fearlessness mediated the relationship between heart rate and aggression (Portnoy et al., Reference Portnoy, Raine, Chen, Pardini, Loeber and Jennings2014). Sijtsema et al. (Reference Sijtsema, Veenstra, Lindenberg, van Roon, Verhulst, Ormel and Riese2010) found that sensation-seeking at ages 13.5 and 16 partially mediated the relationship between heart rate at age 11 and rule-breaking at age 16 in boys. Despite these findings, which suggest that sensation-seeking may underlie the heart rate–antisocial behavior relationship, the lack of experimental evidence leaves our current understanding of this mechanism inconclusive.
In addition to heart rate at rest, researchers are also interested in heart rate reactivity to experimental conditions, including stressors tasks. A meta-analysis of child and adolescent studies reported that reduced heart rate during a stressor task was associated with increased levels of antisocial behavior (d = −0.76, p < 0.0001; Ortiz & Raine, Reference Ortiz and Raine2004). Reduced heart rate reactivity during a laboratory stressor may reflect an impairment in fear or stress processing, which as discussed above could predispose some individuals to engage in antisocial behavior.
As mentioned earlier, heart rate is controlled by the parasympathetic and sympathetic branches of the autonomic nervous system. The parasympathetic nervous system slows down heart rate, while the sympathetic nervous system increases heart rate. Some researchers have attempted to determine whether the relationship between heart rate and antisocial behavior is driven by increased parasympathetic activation or alternatively decreased sympathetic activation. Vagal tone is an index of parasympathetic regulation of the heart that is often estimated using respiratory sinus arrhythmia (RSA) or high-frequency heart rate variability (HRV). Because RSA is an index of parasympathetic nervous system activity, high RSA is associated with a low heart rate. Increased RSA at baseline is thought to index appropriate emotional regulation and engagement with the environment (Beauchaine, Reference Beauchaine2001). Consistent with this interpretation, several studies have found that low RSA serves as a risk factor for aggression (Beauchaine, Hong, & Marsh, Reference Beauchaine, Hong and Marsh2008) and antisocial behavior more generally (Mezzaccapa et al., Reference Mezzacappa, Tremblay, Kindlon, Saul, Arseneault, Seguin and Earls1997; de Wied, van Boxtel, Posthumus, Goudena, & Matthys, Reference de Wied, Boxtel, Posthumus, Goudena and Matthys2009). On the other hand, some authors noted that findings have been inconsistent (Pang & Beauchaine, Reference Pang and Beauchaine2013). In particular, a number of studies observed increased, rather than decreased, RSA in antisocial individuals (Dietrich et al., Reference Dietrich, Riese, Sondeijker, Greaves-Lord, van Roon, Ormel and Rosmalen2007; Scarpa, Fikretoglu, & Luscher, Reference Scarpa, Fikretoglu and Luscher2000; Scarpa, Haden, & Tanaka, Reference Scarpa, Haden and Tanaka2010; Slobodskaya, Roifman, & Krivoschekov, Reference Slobodskaya, Roifman and Krivoschekov1999). These inconsistencies in findings may be due to the importance of the social environment in moderating the effects of RSA on antisocial behavior, as will be discussed later in this chapter.
Skin Conductance
Unlike heart rate – which is controlled by both the sympathetic and parasympathetic branches of the autonomic nervous system – skin conductance is controlled exclusively by the sympathetic branch of the autonomic nervous system. Poor skin conductance fear conditioning (also referred to as electrodermal fear conditioning) is associated with higher levels of aggression (Gao, Raine, Venables, Dawson, & Mednick, Reference Gao, Raine, Venables, Dawson and Mednick2010a) and general antisocial behavior in children and adolescents (Fairchild, van Goozen, Stollery, & Goodyer, Reference Fairchild, Van Goozen, Stollery and Goodyer2008). Poor fear conditioning has also been found to prospectively predict future antisocial behavior. In a sample of 1,795 children from Mauritius, poor electrodermal fear conditioning at age 3 predicted criminal offending at age 23 (Gao, Raine, Venables, Dawson, & Mednick, Reference Gao, Raine, Venables, Dawson and Mednick2010b). Electrodermal fear conditioning in a laboratory setting is thought to reflect conditioning to socializing punishments in childhood. In turn, fear conditioning to socializing punishments in childhood is thought to be central to socialization and conscience development, making the failure to condition a factor that could predispose some individuals to commit aggressive and antisocial behavior (Eysenck, Reference Eysenck1977).
In general, results of psychophysiological studies suggest that antisocial and aggressive individuals display reduced levels of autonomic activity at rest and in stress conditions. This could reflect a relative lack of fear in antisocial individuals or may be driven by a desire for sensation-seeking among those with low arousal. However, some results are inconsistent. It is possible that this inconsistency is due to the failure of some studies to take into account the influence of social variables. The following section will discuss psychophysiological studies that have incorporated a biosocial perspective.
Biosocial Models of Antisocial Behavior
The Dual-Risk Model and Social Push Perspective
In a seminal work, Raine (Reference Raine2002b) reviewed biosocial research that had been conducted to date. He argued that the limited research available at the time provided support for two main models of biosocial interactions: the dual-risk model and the social push perspective. According to the dual-risk model, the presence of both a biological and social risk factor increases the likelihood of antisocial behavior. The risk of aggression is lowered in individuals with only biological or social risk. Unlike the dual-risk model – which considers antisocial behavior as the outcome variable – in the social push perspective, the biological factor is the outcome variable. The social push perspective argues that the relationship between biological factors and antisocial behavior is strongest in those from benign social backgrounds. In other words, in those from disadvantaged backgrounds, biology is less likely to play a role in the development of antisocial behavior; instead, social factors are more likely to be implicated.
A number of psychophysiological studies have provided support for the dual-risk model. Farrington (Reference Farrington, Raine, Brennan, Farrington and Mednick1997), for instance, found that the relationship between heart rate at age 18 and convictions for violence at age 18 to 40 was stronger in those with a poor relationship with a parent at age 18 or a large family size at age 10. More recently, in a study of 8–10-year-old children, Gao, Huang, and Li (Reference Gao, Huang and Li2016) found that prenatal maternal stress was only associated with dimensions of psychopathy in children who also had low resting heart rate or high RSA.
There is also support in the psychophysiological literature for the social push perspective. Raine and Venables (Reference Raine and Venables1984), for instance, found that the relationship between low heart rate and antisocial behavior was confined to those children from higher social classes. As predicted by the social push perspective, heart rate did not predict antisocial behavior in lower-class boys.
Biological Protective Factors
Since the publication of Raine (Reference Raine2002b), there has been a vast increase in the number of researchers investigating biosocial interactions. This new research has broadened our understanding of biosocial interaction and spurned the development of new models to explain these interactions. One such model is the biological protective factors model. While most researchers study risk factors – which increase the likelihood of antisocial behavior – researchers have also begun to emphasize protective factors in antisocial behavior research (Cicchetti, Reference Cicchetti2010; Rutter, Reference Rutter2012). Buffering protective factors buffer or reduce the effects of a risk factor (Lösel & Farrington, Reference Lösel and Farrington2012). In other words, even if a risk factor is present, the simultaneous presence of a protective factor reduces the probability of antisocial behavior. Protective factors are also thought to promote desistance from antisocial behavior in otherwise high-risk individuals. The biosocial protective factors model argues that biological factors may actually reduce the harmful effects of social risk factors for antisocial behavior (Farrington, Reference Farrington, Raine, Brennan, Farrington and Mednick1997; Farrington & Lösel, Reference Lösel and Farrington2012; Portnoy, Chen, & Raine, Reference Portnoy, Chen and Raine2013). Biological factors could also promote desistance from antisocial behavior and violence.
A small, but growing body of psychophysiological research is consistent with the biological protective factors model. For instance, some evidence suggests that a high heart rate may act as a protective factor for antisocial behavior. Using data from the Cambridge Study in Delinquent Development, Farrington (Reference Farrington, Raine, Brennan, Farrington and Mednick1997) found that amongst subjects with high heart rates at age 18, large family size – which was normally a risk factor for violence – was no longer associated with violent convictions. High resting heart rate has also been examined as a possible predictor of desistance. One study showed that antisocial adolescents who desisted from offending by adulthood had higher resting heart rates than antisocial adolescents who later became criminal by age 29 (Raine, Venables, & Williams, Reference Raine, Venables and Williams1995). In another study using this same sample, Raine, Venables, and Williams (Reference Raine, Venables and Williams1996) found that antisocial adolescents who desisted from adult crime had significantly better skin conductance conditioning at age 15 than persisters, who were criminal at age 29, and tended to display better conditioning than normal controls (p < 0.053). On the other hand, De Vries-Bouw et al. (Reference De Vries-Bouw, Popma, Vermeiren, Doreleijers, Van De Ven and Jansen2011) found that resting heart rate did not predict desistance from offending. Though high resting heart rate and enhanced skin conductance fear conditioning appear to be candidates as protective factors, in light of these mixed findings, more research is needed before firm conclusions can be drawn.
Differential Susceptibility Theory
The differential susceptibility theory recognizes that children raised in the same social environment may develop quite differently. This theory argues that some children may be more receptive to environmental influences – either positive or negative – than others (Belsky, Reference Belsky, Ellis and Bjorklund2005; Belsky, Bakermans-Kranenberg, & van Ijzendoorn, Reference Belsky, Bakermans-Kranenburg and van Ijzendoorn2007). In particular, high biological stress reactivity may be either adaptive or maladaptive, depending on the child’s social context (Ellis, Boyce, Belsky, Bakermans-Kranenburg, & van Ijzendoorn, Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van Ijzendoorn2011). The theory predicts that a highly reactive child who grows up in an adverse social environment will be negatively affected by that environment. On the other hand, a highly reactive child who grows up in an especially enriching environment will be positively affected by that environment. A child with low stress reactivity will be little affected by the social environment – either positive or negative. This contrasts with the dual-risk hypothesis, which makes the implicit assumption that neutral and especially enriching environments interact similarly with biological reactivity (Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van Ijzendoorn2011).
There is some evidence that lower physiological activity may be beneficial to children living in adverse social environments. High baseline RSA has been found to buffer children against antisocial behavior problems associated with higher levels of parental marital conflict (El-Sheikh, Harger, & Whitson, Reference El-Sheikh, Harger and Whitson2001), though at least one study failed to find this effect (Leary & Katz, Reference Leary and Katz2004). High RSA has also been shown to protect against the harmful effects of parental problem drinking (El-Sheikh, Reference El-Sheikh2005a) and maltreatment amongst boys (Gordis, Feres, Olezeski, Rabkin, & Trickett, Reference Gordis, Feres, Olezeski, Rabkin and Trickett2010). Research has also shown that increased reactivity could be harmful when combined with an adverse social environment; several studies found that high skin conductance was actually a risk factor for antisocial behavior amongst children at high social risk (Cummings, El-Sheikh, Kouros, & Keller, Reference Cummings, El-Sheikh, Kouros and Keller2007; El-Sheikh, Reference El-Sheikh2005b). This is in contrast to the studies mentioned earlier, which found that high skin conductance was a possible protective factor for antisocial behavior in high-risk individuals (Raine et al., Reference Raine, Venables and Williams1996).
More research is needed to reconcile the competing claims of the dual-risk and differential susceptibility theories as they relate to biological stress reactivity. Additionally, while researchers often examine children living in adverse social environments, little research has examined whether children from especially enriching environments benefit from high autonomic reactivity, as predicted by the differential susceptibility theory. More research is needed that examines children across a wide range of social contexts – both positive and negative.
Biosocial Mediation Models
Most biosocial models consider how biological and social factors interact with one another. However, there is also substantial evidence that social factors could actually affect biological and psychophysiological functioning. Chronic stress, for instance, is thought to affect the body’s biological stress response. Specifically, chronic stress is hypothesized to result in the down-regulation of the body’s stress response system as a way for individuals to cope with chronically threatening environments without constantly evoking hormonal and cardiovascular stress responses (Susman, Reference Susman2006). As discussed earlier, reduced autonomic stress responses are also associated with higher levels of antisocial behavior. In response to this, Choy, Farrington, and Raine (Reference Choy, Raine, Portnoy, Rudo-Hutt, Gao and Soyfer2015) argued that autonomic arousal may act as a mechanism linking social adversity and antisocial behavior. In other words, autonomic arousal may mediate the well-documented relationship between an adverse social environment and antisocial behavior.
Although very little empirical research has tested biosocial mediation models of antisocial behavior, there has been some preliminary research in this area. In a community sample of 454 children living in Philadelphia, early childhood social adversity – as measured by items such as parental education, parental employment, teenage pregnancy, living accommodation status, family size and structure, and parental supervision – was associated with reduced heart rate during stress (Choy et al., Reference Choy, Raine, Portnoy, Rudo-Hutt, Gao and Soyfer2015). In turn, low heart rate during stress was associated with increased levels of antisocial behavior. Consistent with the biosocial mediation model, low heart rate during stress partially mediated the observed relationship between social adversity and both general antisocial behavior and aggressive symptoms of conduct disorder. These results suggest that low heart rate during stress may partly explain why social adversity is associated with higher levels of antisocial behavior and aggression. Research in this area, however, remains very preliminary and it is not yet possible to draw firm conclusions.
Implications for Prevention
There is increasing support for the possibility that biosocial research could help to inform efforts to prevent and treat antisocial and aggressive behavior (Rocque, Welsh, & Raine, Reference Rocque, Welsh and Raine2012; Vaske, Galyean, & Cullen, Reference Vaske, Galyean and Cullen2011). Cornet, de Kogel, Nijman, Raine, and van der Laan (Reference Cornet, de Kogel, Nijman, Raine and van der Laan2014) reviewed studies that examined whether neurobiological factors – including psychophysiological factors – predicted cognitive behavioral therapy treatment outcomes in individuals with antisocial behavior. They concluded that, in general, lower levels of arousal – as indicated by psychophysiological factors, including lower skin conductance, lower resting heart rate, and higher levels of HRV, predicted more favorable outcomes in cognitive-behavioral treatment studies of antisocial behavior. This suggests that psychophysiological factors could help to explain observed heterogeneity in responses to more traditional psychological treatments; this in turn could help researchers to develop more targeted treatments for individuals with antisocial behavior.
More recent findings have not provided support for this conclusion. Cornet, van der Laan, Nijman, Tollenaar, and de Kogel (Reference Cornet, van der Laan, Nijman, Tollenaar and de Kogel2015) examined 121 convicted adult offenders detained in prisons in the Netherlands who took part in a cognitive skills training program. They found that heart rate and RSA stress reactivity did not predict treatment outcomes. On the other hand, poor concentration, a neurocognitive measure, did predict drop-out of the treatment. In general, more research is needed in order to better understand how psychophysiological measures and biological factors more generally could help to predict who benefits most from treatment for antisocial behavior. Additionally, little is known about how biological factors predict treatment outcomes in studies of aggression and violence specifically, rather than antisocial behavior more generally.
Conclusion
This chapter reviewed research into the psychophysiology of antisocial behavior and aggression. In general, studies find that reduced autonomic activity is associated with higher levels of aggression and antisocial behavior. This is evidenced by the reduced heart rate and skin conductance observed in antisocial individuals. Despite the growing acceptance of a biological basis of antisocial behavior, there is also unmistakable evidence that the development of aggression is a multilevel process that cannot be entirely explained by either biological or social factors alone. Several models have been used to explain biosocial interactions; these include the dual-risk hypothesis, social push perspective, biological mediation model, biological protective factors approach, and differential susceptibility theory.
To date, research testing the claims of the biological mediation model, biological protective factors approach, and differential susceptibility theory is limited. More research is needed to determine how well these models explain the interplay between biological and social factors in predicting antisocial behavior. This research could have important implications for the prevention of aggression and antisocial behavior. If researchers are able to identify biological factors that predict treatment outcomes, interventions could be better targeted at those who will most benefit from treatment. Additionally, a better understanding of the social contexts under which biological factors either confer risk or protection could help to improve risk assessment and prediction for aggression and antisocial behavior. Because psychophysiological factors are more easily and inexpensively measured than other biological measures (Gao et al., Reference Gao, Glenn, Peskin, Rudo-Hutt, Schug, Yang, Raine, Gadd, Karstedt and Messner2012), they may be especially beneficial in informing the future treatment and prevention of aggression and antisocial behavior.
Introduction
Neuropsychology has long sought to assess the often-subtle, yet dramatic, effects of brain lesions on information processing and behavior. Following certain brain lesions, a previously well-adapted individual can become irritable, impulsive, incapable of sustaining concentration, and neglectful of social rules. In such cases, the individual’s ability to process information in a socially adaptive way becomes severely impaired. The observation of these profound changes prompted the development of neuropsychological accounts for the deficits, seen in a broad range of behavior problems, including violent behavior.
Here, we update our original review of the neuropsychological approach to violent behavior published in an earlier edition of the handbook (Séguin, Sylvers, & Lilienfeld, Reference Séguin, Sylvers, Lilienfeld, Waldman, Flannery and Vazsonyi2007). In Section I we examine issues pertaining to the assessment of neuropsychological function and the assessment of clinical syndromes, including delinquency and criminality, associated with violence. In Section II, we review developmental issues affecting brain maturation and behavioral regulation. We integrate the aforementioned issues in Section III, where we review studies that help us understand violence from a neuropsychological perspective. As we note, because a key method in neuropsychology has been the use of lesion analyses, we examine the extent to which brain lesions contribute to violence. We then turn to the few neuropsychological studies of violence, and examine the larger body of literature on clinical syndromes associated with violence. There, we examine the extent to which neuropsychological problems have been identified in violence-prone individuals. Finally, we integrate the key observations derived from this review, address limitations in the extant research, and offer suggestions for further research on this important and still-growing area.
Section I: Assessment
Classic neuropsychological testing involves the administration of a battery of tests. These tests are designed to assess a variety of brain functions, ranging from basic perception to more complex neocortical problem-solving, and require either verbal or motor responses. The stimuli may be visual or auditory. Visual stimuli include pictures, abstract designs, and combinations of these stimuli, such as those found in various puzzles, mazes, assortments of objects, pictorial depictions of story lines, printed colors, words, and numbers. Auditory stimuli may involve spoken words, numbers, problems, or stories. Computerized batteries are usually limited to motor responses performed through the click of a mouse or by means of a touch-screen interface, although voice onset recording, eye tracking devices, electrophysiology, and functional brain imaging are increasingly used. Many of these tests qualify as neuropsychological because they were developed to test theories of brain function and were typically validated with lesion analysis studies, brain electrophysiological studies, or functional brain imaging studies. In other words, individuals with relatively well-circumscribed brain lesions performed poorly on such tests, or these tests were found to engage specific brain regions. Thus, these batteries provide a profile of strengths and weaknesses that presumably vary as a function of location and extent of lesions. Findings from these assessments can further help tailor intervention or prevention (Séguin & Pilon, Reference Séguin, Pilon, Reddy, Hale and Weissman2013).
Neuropsychological lesions can result from pregnancy or birth complications, various illnesses, aging, head injury, intracranial tumors, cerebrovascular disorders, exposure to toxic substances, or corrective surgical procedures. The extent to which these events lead to neuropsychological lesions may in turn depend on a variety of moderators, such as the developmental timing of lesions, genetic make-up, and socioeconomic factors. They can also be temporary and reversible, such as those observed under the acute effects of drugs and alcohol or of certain illnesses. Finally, the results of neuropsychological tests are often assumed to reflect the competence of the individual. Nevertheless, there may be a sizable gap between competence and performance. Thus, interpretation of test results should take into account the individual’s test motivation, affect (e.g., mood, anxiety), attention, capacity to remember the sometimes complex rules required for optimal performance, language of administration, and cultural background. We refer the reader to more specialized sources for additional information (Goldstein & McNeil, Reference Goldstein and McNeil2012; Lezak, Howieson, Bigler, & Tranel, Reference Lezak, Howieson, Bigler and Tranel2012).
Finally, and to build on the important role of motivation, the more explicit integration of cognitive and affective neuroscience in the past two decades has contributed to more systematic consideration of both “cool” and rational versus “hot” and affective neuropsychological assessment (Castellanos, Sonuga-Barke, Milham, & Tannock, Reference Castellanos, Sonuga-Barke, Milham and Tannock2006). Whereas “cool” tasks are designed to be more emotionally neutral, tasks can also vary in the degree to which they are administered in an emotionally arousing and “hot” context (Séguin, Arseneault, & Tremblay, Reference Séguin, Arseneault and Tremblay2007). Whereas the former tasks involve more “top-down” integrative regulation (e.g., dorsolateral frontal cortex), the latter tasks involve more “bottom-up” processing (e.g., ventromedial/orbital frontal cortex), such as those more generally described by Stuss (Reference Stuss2011). Although the “cool – hot” distinction has considerable face validity and seems to be taken for granted by many, it is much harder to define operationally as it may not be as categorical as the language we use to describe it implies (Welsh & Peterson, Reference Welsh and Peterson2014).
Clinical Syndromes Associated with Violence
Although physical violence is relatively easy to identify because of its overt nature, there is a paucity of studies examining the neuropsychology of violence. Much of what we know in this domain derives from research on conditions that are associated with violence rather than violence per se. Thus, to appreciate the strengths and limitations of this body of literature, we first discuss clinical syndromes most commonly associated with violence.
Violence research is conducted within two broad and overlapping nomenclatures: legal/judicial and clinical. In the legal/judicial areas, researchers have studied delinquency and criminal behavior. In the clinical arena, physical violence or aggression as a feature, and sometimes as a diagnostic criterion, can be found in a variety of disorders of the Diagnostic and Statistical Manual of Mental Disorders (DSM-5) (American Psychiatric Association, 2013) and the International Classification of Diseases – Mental and Behavioral Disorders (ICD-10CM) (World Health Organization [WHO], 1992). Accordingly, an approach linking neuropsychological function to violence as manifested in clinical disorders fits well within the objectives of the Research Domain Criteria project recently launched by the National Institute of Mental Health (www.nimh.nih.gov/research-priorities/rdoc/index.shtml). Specifically, physical aggression or violence may be found among disorders that are listed under disruptive, impulse-control, neurodevelopmental, and neurocognitive disorders, including Intermittent Explosive Disorder, other Specified Disruptive Impulse-Control and Conduct Disorder, Unspecified Disruptive Impulse-Control and Conduct Disorder, and Antisocial Personality Disorder (ASPD). Physical violence can also occur as a symptom of the Personality Change due to a General Medical Condition, Aggressive Type; Trauma or Stressor-Related Disorders, and in the newly added DSM-5 Disruptive Mood Dysregulation disorder. It is not a formal feature of, but is a potential consequence of, Delusional Disorder – persecutory type, Schizophrenia, Sleep-Wake Disorders, Bipolar Disorder, several Substance-Related and Addictive Disorders, and relational problems including Spouse or Partner Violence. Nevertheless, for none of these disorders is violence a necessary or sufficient criterion.
Psychopathy has not been listed as a separate disease category in DSM-5 or its predecessors. Nevertheless, although DSM-IV-TR regarded ASPD as essentially synonymous with psychopathy, in DSM-5, psychopathy has been added as a specifier to the ASPD category in Section III (but not to the main text) – the section of the manual devoted to newly emerging models and measures. Psychopathic traits are also listed in the Conduct Disorder (CD) category using the specifier “with limited prosocial emotions,” which captures children and adolescents with pronounced callous-unemotional (CU) traits. Indeed, research has shown that children with CD and marked CU traits are distinct from children with CD without these traits in etiology, risk for ASPD later in life (Frick, Ray, Thornton, & Kahn, Reference Frick, Ray, Thornton and Kahn2014), and neurocognitive functioning (Blair, Leibenluft, & Pine, Reference Blair, Leibenluft and Pine2014).
Physical violence has also been studied developmentally. However, in most developmental studies, aggression scales often neglect to distinguish physical from other forms of aggression. For example, the Child Behavior Checklist (CBCL, Achenbach, Edelbrock, & Howell, Reference Achenbach, Edelbrock and Howell1987) yields an aggression scale that comprises 23 items, only three of which refer explicitly to physical aggression. Physical aggression in these scales can be defined as hitting, kicking, biting, use of a weapon, and getting into fights. In some cases, this definition is broadened to include bullying and threats of violence.
Given these limitations, it is possible for neuropsychological studies in the antisocial behavior literature to include nonphysically violent forms of CD, ASPD, psychopathy, or aggression. Further, the clinical syndromes in which physical violence is present are often comorbid with other conditions characterized by impulsivity, drug and alcohol abuse, Attention Deficit Hyperactivity Disorder (ADHD), and Oppositional Defiant Disorder (ODD). Although there is merit to examining how components shared across such externalizing problems (if not across all disorders) relate to neuropsychological function (Castellanos-Ryan et al., Reference Castellanos-Ryan, Brière, O’Leary-Barrett, Banaschewski, Bokde and Bromberg2016), such an approach is also complemented by studies of specificity.
Section II: Developmental Issues
A developmental approach allows the identification of children for whom problems may be chronic as opposed to acute or transient. Accordingly, longitudinal studies in community samples have provided insight into trajectories of antisocial behaviors throughout development. A recent review of this literature indicates that antisocial behaviors tend to display different developmental trajectories, that they have shared but also unique risk factors, that the trajectories associated with violence generally emerge in the preschool years, and, although some trajectories remain high for a small proportion of children as they enter adulthood, antisocial behaviors tend to decline across adulthood – but are nonetheless accompanied by adaptational problems later in life (Séguin & Tremblay, Reference Séguin, Tremblay and Zelazo2013). The bulk of that research has been on males; so much more needs to be done to study females.
On the basis of early prospective longitudinal studies, Moffitt (Reference Moffitt1993) noted that adolescents with a form of early-onset/persistent antisocial behavior, which includes aggression and hyperactivity, showed poorer neuropsychological test performance in early adolescence than did those with a later onset. This finding has been supported by several studies (e.g., Johnson, Kemp, Heard, Lennings, & Hickie, Reference Johnson, Kemp, Heard, Lennings and Hickie2015; Fairchild et al., Reference Fairchild, van Goozen, Stollery, Aitken, Savage and Moore2009), although brain-imaging studies have not fully supported this distinction (e.g., Fairchild et al., Reference Fairchild, Passamonti, Hurford, Hagan, von dem Hagen and van Goozen2011; Jiang et al., Reference Jiang, Guo, Zhang, Gao, Wang and Situ2015). These findings underscore a key methodological point, namely, that comparing groups of adolescents or adults without knowledge of their natural developmental history can be fraught with problems. Nevertheless, the developmental sequence is often assumed to be from neuropsychological function to behavior but little research has been conducted to verify that assumption. Hence, developmental studies need to use a more fully longitudinal approach that permits the examination of reciprocal transactions between neuropsychological function and behavior (Pinsonneault, Parent, Castellanos-Ryan, & Séguin, Reference Pinsonneault, Parent, Castellanos-Ryan, Séguin, Beauchaine and Hinshaw2015).
Risk Factors that May Affect Brain Development
As developmental patterns have been studied with longitudinal designs, there has been increasing interest in conditions that can contribute both to those patterns and to poor neuropsychological function, such as a history of exposure during or after pregnancy to brain-altering psychopharmacological agents (e.g., cigarette smoke, alcohol, drugs), perinatal or birth complications, nutrition, traumatic experiences (e.g., abuse), chronic stress, or behavior problems that heighten the risk of head trauma through accidents or fights (e.g., ADHD, ODD). For example, cigarettes may exert early effects in the intrauterine environment and later in the home environment. For example, a review of 20 published articles on the association between tobacco smoke exposure in utero and cognition found robust associations for lower academic achievement and intellectual functioning after controlling for other variables (Clifford, Lang, & Chen, Reference Clifford, Lang and Chen2012). Nevertheless, genetically informed designs raise the possibility that at least some of this linkage may be genetic (e.g., D’Onofrio, Van Hulle, Goodnight, Rathouz, & Lahey, Reference D’Onofrio, Van Hulle, Goodnight, Rathouz and Lahey2012).
The mechanisms through which in-utero or early-life adverse exposures affect brain development and violence risk are unclear, but an emerging overarching hypothesis is that early-life adversity produces long-lasting epigenetic alterations that influence gene expression profiles in key biological systems, such as the hypothalamic–pituitary–adrenal axis, the immune system, as well as neurotransmitters, such as serotonin (Booij et al., Reference Booij, Tremblay, Leyton, Séguin, Vitaro and Gravel2010; Wang et al., Reference Wang, Szyf, Benkelfat, Provençal, Turecki and Caramaschi2012), which, in turn, alter the ability to learn to inhibit physical aggression. Genetic factors probably moderate these effects of early-life stress on the epigenome and influence the risk for, or resilience to, developing violent behaviors (Provençal, Booij, & Tremblay, Reference Provençal, Booij and Tremblay2015). Furthermore, there could be an assumption that neurocognitive problems lead to behavior problems. This would not be surprising given that this literature developed out of lesion research and that remedial interventions often target cognitive abilities to change behavior. However, this assumption is rarely tested as it is equally possible for transactions to change direction across developmental stages. Consequently, children who show behavior problems could eventually get more or less support and exposure to alternative problem-solving approaches through social moderators, which would enhance or diminish their repertoire of cognitive and self-regulatory skills (see Pinsonneault et al., Reference Pinsonneault, Parent, Castellanos-Ryan, Séguin, Beauchaine and Hinshaw2015). Finally, substance use during adolescence, such as smoking (Lydon, Wilson, Child, & Geier, Reference Lydon, Wilson, Child and Geier2014) or cannabis use (Castellanos-Ryan et al., Reference Castellanos-Ryan, Pingault, Parent, Vitaro, Tremblay and Séguin2017), probably also affects the developmental course of brain cognitive and motivational systems.
Section III: The Effects of Brain Lesions on the Risk for Violence
Given the background considerations reviewed thus far, we now address two complementary questions: (1) “Do some brain lesions dependably increase the risk for violent behavior?” and (2) “Do violence-prone individuals exhibit specific neuropsychological deficits?”
Interest in the cognitive and emotional regulation aspects of the brain-violence relation has centered largely on the role of the frontal lobe because of its centrality to the regulation of social behavior. Frontal patients have difficulty organizing and regulating their behavior in response to external stimuli. The more cognitive functions of the frontal lobe, often referred to as the executive functions, include working memory, which involves the online maintenance of information and the active processing of that information as an individual engages in action, interference control (attention and inhibition), and cognitive flexibility (Diamond, Reference Diamond2013). The emotional regulation function was supported by a study of individuals with lesions comprising the orbitofrontal cortex, who tend to be disinhibited, socially inappropriate, susceptible to misinterpreting others’ moods, impulsive, unconcerned with the consequences of their actions, irresponsible in everyday life, lacking in insight into the seriousness of their condition, and prone to weak initiative (Fazel, Philipson, Gardiner, Merritt, & Grann, Reference Fazel, Gulati, Linsell, Geddes and Grann2009). The main consequence of such lesions has been linked with impaired performance on a wide variety of neuropsychological tests measuring reward and punishment sensitivity, decision making, and processing of social cues (Jonker, Jonker, Scheltens, & Scherder, Reference Jonker, Jonker, Scheltens and Scherder2015). In many imaging studies, differences in brain function or structure were independent of performance deficits, and therefore require additional neuropsychological testing to be evaluated. Many of these neuropsychological deficits reflect malfunction of limbic brain regions, such as the amygdala, cingulate, and/or striatum.
The ensuing syndrome from orbitofrontal lesions was historically labeled acquired sociopathy (Damasio, Reference Damasio2000), acquired ASPD (Meyers, Berman, Scheibel, & Hayman, Reference Meyers, Berman, Scheibel and Hayman1992), or pseudopsychopathy. Because these lesions appear to lead to certain psychopathic-like behaviors, they were often thought to underlie physical violence. Nevertheless, there is limited support for this hypothesis. Studies of war veterans, for example, revealed that although veterans with prefrontal lesions exhibit more positive implicit attitudes towards violence compared with veterans without lesions, they do not differ on explicit measures of aggression. Thus, although prefrontal lesions may impact inhibition over automatic (implicit) aggressive reactions to perceived provocation (Cristofori et al., Reference Cristofori, Zhong, Mandoske, Chau, Krueger and Strenziok2016), physical violence directly resulting from lesions is rare in acquired forms of antisocial behavior. This finding may in part be due to the fact that individuals with acquired brain lesions do not necessarily possess the many pre-morbid risk factors associated with physical violence. In fact, aggression (including physical aggression) appears to be more likely to be associated with acquired frontal lesions only when there is a premorbid history of psychopathology (Fazel et al., Reference Fazel, Philipson, Gardiner, Merritt and Grann2009). Otherwise, normative behavior may be the result of interactions among these dynamically related frontal systems (Verbruggen, Reference Verbruggen2016).
Other cortical abnormalities associated with aggressive outbursts include acute episodes of temporal lobe (temporolimbic) epilepsy (also known as interictal violence), which is similar in manifestation to IED. Nevertheless, violence in epilepsy seems to be lower than in the general population (Fazel et al., Reference Fazel, Philipson, Gardiner, Merritt and Grann2009). Taken together, these findings suggest that brain lesions are rarely sufficient causes of violence. At the same time, they may lead to violence in the context of other risk factors.
Neuropsychological Studies of Physically Violent Behavior
The other main approach to the neuropsychology of violence is to study physically violent individuals. We begin with the most severe form of violence, murder, and move on to less severe forms of physical aggression. We then follow with disorders and antisocial behavior problems in which physical violence is sometimes present.
Murderers
Several studies of murderers have shown reduced activity in frontal brain regions, which is consistent with the dysfunctional frontal lobe hypothesis, a theory postulated for the first time more than two decades ago during the early years of brain imaging research. In one of the first studies, “frontal dysfunctions” were found in 65% of murderers (Blake, Pincus, & Buckner, Reference Blake, Pincus and Buckner1995). In a more recent study, among incarcerated youth, homicide offenders (80% self-reported) showed structural differences in the temporal lobes, hippocampus, and posterior insula (Cope et al., Reference Cope, Ermer, Gaudet, Steele, Eckhardt and Arbabshirani2014). Further, when these homicide offenders were compared to typically developing youth in a recent meta-analysis, they appeared to show consistently larger effect sizes across several structures (Rogers & De Brito, Reference Rogers and De Brito2016).
Few studies have examined neuropsychological performance in murderers. The neuropsychological performance in murderers seems to depend on subtype; affective/impulsive murderers tend to display poorer neuropsychological performance than predatory/instrumental murderers, especially on measures of intelligence, memory, attention, and executive functions (Hanlon, Brook, Stratton, Jensen, & Rubin, Reference Hanlon, Brook, Stratton, Jensen and Rubin2013). These differences appear to be consistent with differences in brain activation (Raine et al., Reference Raine, Meloy, Bihrle, Stoddard, LaCasse and Buchsbaum1998).
Although the frontal lobe has been implicated in these and other studies, they almost surely account for only one portion of the explanation. Indeed, in addition to frontal hypoactivation, many imaging studies found increases in the activity of subcortical areas, which is consistent with neural models of aggression, postulating that aggression results from deficits in cortical top-down control and/or facilitation of bottom-up signaling triggered from subcortical limbic circuits (Davidson, Putnam, & Larson, Reference Davidson, Putnam and Larson2000). Importantly, frontal lobe hypofunction is not specific to murderers (e.g., Siever, Reference Siever2008). The lack of specificity indicates that other factors like genetic make-up, early development, and other factors reviewed in this book may play an important role in the risk for committing homicide.
Physical Aggression
Although murder is the most severe form of violence, investigators have also examined whether neuropsychological problems are evident in milder forms of violence, such as in physical aggression as observed in community samples. There are surprisingly few neuropsychological studies of physical aggression per se.
Séguin et al. (Reference Séguin, Pihl, Harden, Tremblay and Boulerice1995) sought to develop a neuropsychological test battery on the basis of (a) the frontal lobe and memory work of Petrides and Milner (1985) and (b) reviews of the neuropsychology of delinquency and conduct disorder, such as Moffitt’s (Reference Moffitt, Tonry and Morris1990), which identified deficits in language abilities, executive function, and cerebral dominance. Using that approach, we first found that working memory, a basic ability involved in executive function, was poorest in boys from a community sample with a history of physical aggression even after controlling for nonexecutive abilities (Séguin, Pihl, Harden, Tremblay, & Boulerice, Reference Séguin, Pihl, Harden, Tremblay and Boulerice1995). In follow-up, we controlled statistically for ADHD or teacher-rated hyperactivity and still found working memory impairments even after controlling for IQ (Séguin, Boulerice, Harden, Tremblay, & Pihl, Reference Séguin, Boulerice, Harden, Tremblay and Pihl1999). In a third study, we selected young adult males not only on the basis of a physically aggressive history (teacher-rated from kindergarten to age 15), but also of hyperactivity (without a focus on inattention or impulsivity) (Séguin, Nagin, Assaad, & Tremblay, Reference Séguin, Nagin, Assaad and Tremblay2004). We found no statistical interaction, but we did find clear additive effects, even after controlling for test motivation. In other words, both physical aggression and hyperactivity exhibited significant independent and additive associations with neuropsychological function.
In sum, poor neuropsychological function is often found in adolescents and young adults from the community with a history of physical aggression. These deficits appear to be independent from other externalizing behavior problems. Developmentally, these associations and specificity to physical aggression or hyperactivity can be detected as early as the preschool years (Séguin, Parent, Tremblay, & Zelazo, Reference Séguin, Parent, Tremblay and Zelazo2009).
Neuropsychological Studies of Antisocial Disorders in Which Physical Violence May Be Present
The bulk of research on the neuropsychology of violence derives from clinically oriented studies of disorders and antisocial behavior problems that may include physical violence.
Antisocial Behaviors
Several of the behavioral problems listed here have been grouped under the global label of “antisocial behaviors.” These behaviors, among others, were examined in meta-analyses of studies of executive function by Morgan and Lilienfeld (Reference Morgan and Lilienfeld2000) and more recently updated and expanded by Ogilvie et al. (Reference Ogilvie, Stewart, Chan and Shum2011). The first meta-analysis by Morgan and Lilienfeld (Reference Morgan and Lilienfeld2000) comprised 39 studies, yielding 4,589 participants total. To be included in the meta-analysis, tests of executive functioning must have attempted to measure volition, planning, purposive action, or effective performance, and either differentiated patients with frontal lesions from other patients or preferentially activated the frontal cortex in previous studies. To investigate whether antisocial behavior was related to executive functioning deficits per se rather than neuropsychological deficits in general, three neuropsychological tests that do not rely heavily on executive functioning were analyzed as “control” measures. The antisocial behavior groups used in the meta-analysis included individuals meeting criteria for one or more of the following antisocial behavior problems: ASPD, CD, psychopathic personality disorder, criminality, or delinquency.
The results indicated that the antisocial behavior groups performed significantly worse than comparison groups, with a combined and weighted effect size (Cohen’s d [Cohen, Reference Cohen1992] of 0.62 standard deviations). Two of the three nonexecutive functioning neuropsychological tests also produced significant, albeit weaker, differences between groups, with the antisocial behavior groups performing significantly worse with effect sizes of d = 0.34 and d = 0.39 standard deviations. However, tests revealed significant heterogeneity of effect sizes, pointing to the possibility of moderators. When considering the type of antisocial grouping used in the studies, the heterogeneity of effect sizes was reduced within each group and all group effect sizes remained significantly different than zero. Moreover, criminality (d = 1.09, weighted d = 0.94) and delinquency (d = 0.86, weighted d = 0.78) were most strongly associated with executive functioning deficits. Potential moderators including age, sex, ethnicity, and IQ were not associated with the magnitude of the observed effect sizes, although scores on some of these moderators (e.g., sex, ethnicity) were not reported in all studies.
The second meta-analysis by Ogilvie, Stewart, Chan, and Shum (Reference Ogilvie, Stewart, Chan and Shum2011), built on the Morgan and Lilienfeld (Reference Morgan and Lilienfeld2000) meta-analysis, brought up the total number of studies to 126, involving 14,786 participants. This meta-analysis added more recent studies published since the original meta-analysis as well as a wider range of EF and non-EF tasks. It corroborated the results of the initial meta-analysis by showing an overall association between executive functioning and antisocial behaviors (d = 0.44), but also heterogeneity of effect sizes across definitions of antisocial behavior. This heterogeneity may in part have stemmed from the fact that studies used to estimate effect size could be considered under more than one antisocial behavior category. Nonetheless, and as in the initial meta-analysis, the largest effect size was found for criminality (d = 0.61). Other reported effect sizes were ODD/CD (d = 0.54), psychopathy (d = 0.42), delinquency (d = 0.41), physical aggression (d = 0.41), and ASPD (d = 0.19).
Although the effect sizes across meta-analyses were mostly similar, Ogilvie et al. (Reference Ogilvie, Stewart, Chan and Shum2011) suggested that some of the differences may have been due to the inclusion of a wider range of EF and non-EF tasks as well as measures and operationalization of antisocial behaviors that have become more specific and sensitive over the years. Some of our work can serve to illustrate this issue. For example, when forming groups on the basis of a history of physical aggression, the effect sizes of neuropsychological tests in a study using developmental trajectory methodology (empirically based) as was used in a later study (Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004) were much larger than those found when using arbitrary thresholds (theoretically based) in an earlier study (Séguin et al., Reference Séguin, Pihl, Harden, Tremblay and Boulerice1995).
By the time of the later meta-analysis (Ogilvie et al., Reference Ogilvie, Stewart, Chan and Shum2011), there was then also a sufficient number of studies to examine the potential additive role of ADHD. The studies that included participants with comorbid ADHD diagnoses showed the largest effect sizes. The mechanisms through which ADHD adds these other antisocial behavior problems in their association with neurocognitive functioning remain to be better studied (see also hypotheses proposed by Sonuga-Barke, Cortese, Fairchild, & Stringaris, Reference Sonuga-Barke, Cortese, Fairchild and Stringaris2016). This finding supports the need for careful selection of study participants in future studies when studying specific relationships between executive functioning and antisocial behaviors.
Both meta-analyses identified several limitations in this body of research, including small sample sizes, poor comparison group selection, and minimal control over potentially confounding factors such as psychiatric comorbidity. Furthermore, Morgan and Lilienfeld (Reference Morgan and Lilienfeld2000) pointed out that the finding that criminality and delinquency were associated with more pronounced executive functioning deficits than the other groups is difficult to interpret given the differences in comparison groups used across studies. For example, executive functioning studies frequently compared criminals or delinquents with normal or unselected samples, whereas psychopaths were frequently compared with nonpsychopathic criminals. These concerns were supported empirically (Ogilvie et al., Reference Ogilvie, Stewart, Chan and Shum2011).
Psychopathy
Psychopaths are generally more violent than other criminals (Hare, Reference Hare1999) and consequently have received considerable research attention in the past three decades. Reports of neuropsychological performance among psychopathic participants, especially executive functioning, have been mixed. The two meta-analyses examining PCL-R and non-PCL-R-defined psychopathy indicate that the average effect sizes of neuropsychological (executive functioning) deficits were small to medium (d = 0.29 in Morgan and Lilienfeld [Reference Morgan and Lilienfeld2000] and d = 0.42 in Ogilvie et al. [Reference Ogilvie, Stewart, Chan and Shum2011]).
Possible explanations for the high levels of variability across studies are differences in the operationalization of psychopathy as well as the existence of potential psychopathy subtypes. For instance, a number of studies have shown that low-anxious psychopaths show impaired functioning on neuropsychological tasks that rely primarily on frontal lobe function, such as executive functioning (Smith, Arnett, & Newman, Reference Smith, Arnett and Newman1992), cued attention (Zeier, Maxwell, & Newman, Reference Zeier, Maxwell and Newman2009), as well as economic decision making (Koenigs, Kruepke, & Newman, Reference Koenigs, Kruepke and Newman2010). In addition, successful psychopaths (defined by being nonconvicted) showed better performance on an executive functioning task than unsuccessful (convicted) psychopaths (Ishikawa, Raine, Lencz, Bihrle, & LaCasse, Reference Ishikawa, Raine, Lencz, Bihrle and LaCasse2001). It is unclear, however, whether this lattermost difference is independent of possible between-group differences in general intelligence.
Beyond studies using more classic neuropsychological tests, psychopaths appear to experience greater difficulty in shifting a dominant behavior when contingencies are changed and reversed. Newman and colleagues have proposed two pathways to account for that impairment: (1) a difficulty in regulation of affect and (2) a more fundamental information-processing deficit related to attention, i.e., a difficulty in shifting attention to peripheral but potentially meaningful information from the environment (Newman & Lorenz, Reference Newman, Lorenz, Davidson, Scherer and Goldsmith2003). This so-called response modulation theory is one of the most widely studied cognitive models of psychopathy. After its initial formulation more than three decades ago (Gorenstein & Newman, Reference Gorenstein and Newman1980), the theory has been refined over the years, taking into account new research. A recent meta-analysis (Smith & Lilienfeld, Reference Smith and Lilienfeld2015), including 94 studies involving 7340 participants, found a small to medium effect size (d = 0.41) for the association between response modulation deficits and psychopathy, which is within the same range as those found with other psychological theories of psychopathy (Lilienfeld, Smith, & Watts, Reference Lilienfeld, Smith and Watts2016). Nevertheless, the several clinical, demographic and methodological variables (e.g., anxiety levels, ethnicity, measures used) that were found to moderate the strength of the relationship, as well as the several methodological shortcomings of studies (e.g. publication bias), raise questions regarding the comprehensiveness of the theory in explaining psychopathy.
A number of other cognitive theories preceded and followed the response modulation theory. Recently, the impaired integration theory has been proposed to integrate cognitive and affective models of psychopathy with neurobiological data (Hamilton, Racer, & Newman, Reference Hamilton, Racer and Newman2015). This theory postulates that psychopathy results from an impaired ability to rapidly integrate sensory information into a unified percept. This deficit in turn leads to unelaborated mental representations and an underdevelopment of associative neural networks. Underdeveloped connectivity in specific brain networks ostensibly underlies the psychopathy profile. For instance, decreased connectivity in networks involved in affective processing underlie the callous emotional traits present in psychopathy, whereas decreased connectivity in attentional networks leads to poor integration of different types of information (e.g., cognitive-affective, perceptual-motor). Although more research is needed to support this promising theory, such an integrative approach dovetails with developments in cognitive neuroscience, modeling behavioral and cognitive processes as dynamic large-scale neural networks, rather than as isolated, static brain regions.
Criminality and Delinquency
Some of the important theoretical work of Moffitt (Reference Moffitt1993) initially centered on delinquency. In one study, early-onset/persistent delinquents performed more poorly than did “late-onset” delinquents on IQ and other neuropsychological tests (Taylor, Iacono, & McGue, Reference Taylor, Iacono and McGue2000), as predicted by Moffitt’s (Reference Moffitt1993) developmental theory of antisocial behavior. In this study, however, IQ was not used as a covariate, rendering conclusions regarding the specificity of cognitive deficits unclear. Furthermore, the use of global scales of delinquency or criminality may obscure key relations between specific behaviors and neuropsychological function. For example, one study revealed that, among juvenile delinquents, IQ was positively related to theft, but negatively related to violence (Walsh, Reference Walsh1987). Presumably, this finding reflects the requirement for planning for theft, but an impulsive problem-solving style for violence. Intrigued by this isolated report, we analyzed data from the Rutgers longitudinal study (White, Bates, & Buyske, Reference White, Bates and Buyske2001). In that study, theft and violence were initially combined within a global index of delinquency. Neuropsychological function had failed to separate persistent from adolescence-limited delinquents. Following our own work with physical aggression and hyperactivity (Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004), we identified trajectories for theft and for physical violence. Using the same tests as used by White et al. (Reference White, Bates and Buyske2001), we replicated Walsh’s (Reference Walsh1987) finding and found the poorest neuropsychological function in highly violent individuals who were low on theft (Barker et al., Reference Barker, Séguin, White, Bates, Lacourse and Carbonneau2007). In another follow-up to the Séguin et al. (Reference Séguin, Nagin, Assaad and Tremblay2004) study, we found that verbal abilities were negatively related to trajectories of physical aggression but positively associated with theft (Barker et al., Reference Barker, Tremblay, van Lier, Vitaro, Nagin and Assaad2011). In that study, lower levels of inductive reasoning were associated with increases in theft across adolescence. Interestingly, symptoms of ADHD accounted for part of the neurocognitive test links with physical aggression, but did not account for the associations with theft. Together, these studies highlight the need for a finer parsing of the relation between neuropsychological function and global indices of antisocial behavior.
One category of delinquency that is receiving increasing attention is sexual offending. A meta-analysis, involving 23 studies and a total of 1,756 participants, on neuropsychological performance in sex offenders confirmed that adult sex offenders performed worse on neuropsychological tasks than did members of the general population (d = 0.59), but also showed that sex-offending is highly heterogeneous (Joyal, Beaulieu-Plante, & de Chantérac, Reference Joyal, Beaulieu-Plante and de Chantérac2014). Subgroup analyses showed that sex offenders against children tended to perform lower on so-called higher-order executive functioning tasks compared to sex offenders against adults (albeit with small effect size, d = 0.23), whereas sex offenders against adults were similar to nonsex offenders, with lower scores in verbal fluency and inhibition. The meta-analysis highlights the need for studies in specific subgroups. Few studies have focused on adolescent sex offenders. One study examined a sample of 127 adolescent sex offenders and 56 adolescents convicted of nonsexual offenses (Morais, Joyal, Alexander, Fix, & Burkhart, Reference Morais, Joyal, Alexander, Fix and Burkhart2016). In contrast to studies conducted in adults, adolescent sex offenders who had victimized children obtained significantly higher scores on measures of complex executive functioning tasks than both adolescent offenders with peer-aged or older victims and adolescent nonsex offenders. Taken together, these findings suggest possible complex interactions involving developmental age of the offender and victim. Longitudinal studies following adolescent offenders over time are needed to understand the relevance of neuropsychological performance for recidivism and recovery.
Many of the findings related to the neuropsychology of violence may bear implications for the justice system. Although neuroscientific evidence on altered frontal-limbic brain function in relation to violence in adults appears to be increasingly being used in the courtroom (see Klaming & Koops (Reference Klaming, Koops and Spranger2012) and Steinberg (Reference Steinberg2013) for reviews of the literature and case examples), decisions for trying adolescents who committed violent crimes in adult courts hinge on research on brain development and maturation. From a brain maturation perspective, some claim that adolescent brains are not sufficiently mature to justify trying violent adolescents in adult courts. Nevertheless, the overwhelming majority of adolescents are not violent, raising the question of why legal decision making should use age per se as a criterion (Satel & Lilienfeld, Reference Satel and Lilienfeld2013). Moreover, neuropsychological function and brain maturation may account only for a relatively small amount of variance in physical violence. Thus, the sensitive issue of trying violent adolescents in adult courts should probably rest on broader grounds and evidence reviewed herein to inform the legal decision making system should be used with caution.
Intermittent Explosive Disorder
DSM-5 (American Psychiatric Association, 2013) places intermittent explosive disorder (IED) under the category of Disruptive Impulse-Control and Conduct Disorders. The primary feature of IED is the frequent experience of short and discrete episodes of aggressive behavior of rapid onset resulting in personal injuries or property damage. The course, onset, and prevalence of IED are poorly understood, although this condition appears to be more common in males than females. Not surprisingly, there is a dearth of studies on the neuropsychological correlates of IED, although its neurobiology is better understood (Coccaro, Reference Coccaro2012). In one controlled study, Best, Williams, and Coccaro (Reference Best, Williams and Coccaro2002) showed that IED participants performed poorly on the Iowa Gambling Task compared with healthy controls. Nevertheless, there were no group differences on the Self-Ordered Pointing test, a working memory test most sensitive to physical aggression (Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004). This finding is consistent with the idea that IED may be neuro-cognitively distinctive from other, more common forms of violence, suggesting an IED profile with predominantly affective and social rather than cognitive deficits (Lee et al., Reference Lee, Arfanakis, Evia, Fanning, Keedy and Coccaro2016). More research is needed to further understand whether impaired neuropsychological functioning in IED is distinct from other impulse-control disorders. Such research should be embedded in a developmental framework capturing the age of onset (Coccaro, Reference Coccaro2012).
Conduct Disorder
When Morgan and Lilienfeld (Reference Morgan and Lilienfeld2000) reviewed the CD literature they found a medium effect size of d = 0.4 (weighted d = 0.36) for poor performance on executive functioning tasks. The meta-analysis by Ogilvie et al. (Reference Ogilvie, Stewart, Chan and Shum2011), which mixed both ODD and CD but separated ADHD, reported a slightly higher effect size (d = 0.54).
Combining CD and ODD in analyses may be justified when addressing certain research questions, especially those aimed at understanding what is common among externalizing problems, if not across psychopathologies (Castellanos-Ryan et al., Reference Castellanos-Ryan, Brière, O’Leary-Barrett, Banaschewski, Bokde and Bromberg2016). At the same time, this approach is problematic when trying to examine specificity to violence, because neither ODD nor CD necessarily encompass physical aggression. We first provide an example to illustrate the potential importance of accounting for the CD symptom of physical aggression in studies of the neuropsychology of antisocial behavior. In one study that focused on physical aggression and hyperactivity, 67% of boys classified as CD and 72% of boys classified as ADHD between the ages of 14 and 16 were in the high physical aggression and high hyperactivity trajectories, respectively, on the basis of teacher ratings from age 6 to 15 (Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004). Nevertheless, across the entire sample only 5% and 6.7% of boys met criteria for CD or ADHD, respectively. This finding suggests that if one is high in physical aggression (or hyperactivity), one is more likely to meet criteria for CD (or ADHD). But that study was different from the typical ones selecting for CD and ADHD in that it selected children on the basis of physical aggression and hyperactivity instead. A selection based on CD and ADHD may not necessarily have included either physical aggression or hyperactivity, or these specific behaviors may have contributed only a low weight to the diagnoses. Thus, a focus on physical aggression and hyperactivity yielded a greater number of study participants, most of whom did not meet criteria for CD or ADHD, and those more specific behaviors were sensitive to neuropsychological function (Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004).
Second, in addition to the comorbidity of other externalizing problems with CD, we have already highlighted the problem of heterogeneity of mechanisms correlated with two of the four key symptoms of CD – physical aggression and theft. There is now a growing number of studies examining subtypes of CD symptoms that supports the replicability of the negative association between physical aggression and neurocognitive dysfunction, but that finds links between such dysfunction and theft or other nonaggressive conduct problems to be either essentially nonexistent (Barker et al., Reference Barker, Séguin, White, Bates, Lacourse and Carbonneau2007; Hancock, Tapscott, & Hoaken, Reference Hancock, Tapscott and Hoaken2010) or positive (Barker et al., Reference Barker, Tremblay, van Lier, Vitaro, Nagin and Assaad2011; Walsh, Reference Walsh1987). Thus, from a neuropsychological perspective, the conflation of symptoms within CD may mask otherwise-important heterogeneity (Burt, Reference Burt2009, Reference Burt2013).
Antisocial Personality Disorder
In DSM-5, ASPD is diagnosed only in individuals over the age of 18 with evidence of CD. As we mentioned earlier, psychopathy is often found among individuals with ASPD. We also note that PCL-R scores > 20 but < 30 (the standard PCL-R cut-off for psychopathy) may also reflect ASPD, and that many ASPD individuals will have a criminal record. Compared with other categories in which antisocial behavior plays a role, there are relatively few neuropsychological studies of ASPD. The most recent meta-analysis included 11 studies and found that ASPD’s associations with executive dysfunction are weak (effect size d = 0.19), (Ogilvie et al., Reference Ogilvie, Stewart, Chan and Shum2011). One study found poorer executive function (working memory, cognitive flexibility, and inhibitory control) in ASPD relative to nonoffenders, regardless of the presence or absence of co-occurring psychopathy (De Brito, Viding, Kumari, Blackwood, & Hodgins, Reference De Brito, Viding, Kumari, Blackwood and Hodgins2013).
Other Disorders Where Violence May Be Present
Although violence is likely to be found in the more explicitly antisocial disorders already reviewed, violence is often associated with alcohol- and substance-use disorders, schizophrenia, and bipolar disorder. Hence, we briefly review their relation to neuropsychological function.
Alcohol and Substance Use
A considerable limitation to the literature investigating neuropsychological function in violent problem behaviors and associated disorders is a potential confound with alcohol and substance abuse. In fact, most violent acts may occur under the influence of substances (Murdoch, Pihl, & Ross, Reference Murdoch, Pihl and Ross1990; Room, Babor, & Rehm, Reference Room, Babor and Rehm2005). Although the culture of illegal drugs is also associated with violence, this is not dispositive evidence for the violence potential of the drugs themselves. The other key issue to consider is developmental transactions between substance use and neuropsychological function. Although several studies note cross-sectional correlations between substance use and neuropsychological function in acute states, few have examined whether the association found before the onset of substance use was changed afterwards or if use was related to developmental change in neuropsychological function. Building on a series of longitudinal studies on the neuropsychology of physical aggression (Séguin et al., Reference Séguin, Pihl, Harden, Tremblay and Boulerice1995; Séguin, Arseneault, Boulerice, Harden, & Tremblay, Reference Séguin, Arseneault, Boulerice, Harden and Tremblay2002; Séguin et al., Reference Séguin, Nagin, Assaad and Tremblay2004), we have recently reported that early onset of cannabis use was predicted by pre-use IQ and poor short-term and working memory (Castellanos-Ryan et al., Reference Castellanos-Ryan, Pingault, Parent, Vitaro, Tremblay and Séguin2017). Nevertheless, we also showed that, by age 20, changes only in verbal IQ (accounted for by poor high-school graduation), trial-and-error learning, and reward-processing were linked to onset and frequency of use in adolescence, even after controlling for CD, ADHD, and concurrent cannabis or alcohol use. Although such studies do not demonstrate causality, the developmental transactions observed need to be taken into account in studying the neuropsychology of violence.
Our central question concerns the role of neuropsychological factors in this equation. Most research on this topic has focused on executive function. The role of executive function in alcohol-associated aggression has been illustrated well by the work of Giancola (Reference Giancola2004), who showed that aggression in reaction to provocation (using a shock paradigm) was possibly more a function of executive function than of alcohol. Nevertheless, synergistic mechanisms may be at play, as alcohol preferentially increased aggression in men with lower levels of executive function. Alcohol may also moderate the quality of executive function on a state (temporary) basis, mostly on the descending limb of the blood-alcohol curve (Pihl, Paylan, Gentes-Hawn, & Hoaken, Reference Pihl, Paylan, Gentes-Hawn and Hoaken2004). Finally, recent models have attempted to integrate cognitive theories of alcohol-induced aggression with neurobiological theories of aggression. For example, Heinz, Beck, Meyer-Lindenberg, Sterzer, and Heinz (Reference Heinz, Beck, Meyer-Lindenberg, Sterzer and Heinz2011) proposed that the presence of certain risk genotypes/alleles in combination with adverse environmental factors (e.g., early-life stress) affect the functioning of key neurotransmitter systems (e.g., serotonin, GABA). This outcome in turn would lead to an increased activation in subcortical (bottom-up) limbic circuits and impaired (top-down) prefrontal function, which may place individuals at risk for both increased alcohol intake and impulsive aggression. Acute or chronic alcohol intake, in turn, can further inhibit executive control and facilitate aggressive behavior.
Schizophrenia
Although the prevalence of violence in schizophrenia and other disorders where psychosis is present is about 10% across several studies, its relative prevalence as opposed to the general population is highly variable, with odds ratios varying from 1 to 7 for men, to 4–29 for women (Fazel, Gulati, Linsell, Geddes, & Grann, Reference Fazel, Gulati, Linsell, Geddes and Grann2009). In that meta-analytic study, the risk of violent behavior seemed to be highest when there was comorbid substance use, but the rate of violence did not differ between individuals with schizophrenia and with non-schizophrenia-related psychosis. One meta-analysis, investigating risk factors for violence in (primarily) schizophrenia, found that, among several clinical and demographic factors, substance misuse and particularly factors related to violent and nonviolent criminal history (e.g., history of assault, history of imprisonment for any offense, history of recent arrest for any offense, history of conviction for a violent offense) were the strongest predictors of risk of violent behavior (Witt, van Dorn, & Fazel, Reference Witt, van Dorn and Fazel2013).
Another meta-analysis (Schug & Raine, Reference Schug and Raine2009) compared neuropsychological performance in individuals with (a) schizophrenia and antisocial behavior, (b) schizophrenia without antisocial behaviors, and (c) antisocial behaviors without schizophrenia. The authors found that individuals with schizophrenia and antisocial behaviors showed widespread cognitive impairments (IQ, attention, executive function, and memory) relative to individuals with antisocial behaviors without schizophrenia, and exhibited reduced general intellectual functioning and memory dysfunction, relative to individuals with schizophrenia without antisocial behaviors. These associations were characterized by small effect sizes (d = 0.2–0.3 range), but may suggest that schizophrenia with antisocial behaviors may be a subcategory of schizophrenia, a finding supported by electrophysiological and imaging studies (Schug & Raine, Reference Schug and Raine2009). The authors explicitly chose liberal, though arguably valid, approaches to classifying antisociality and schizophrenia. Accordingly, they noted considerable heterogeneity of effect sizes. Because of this they also highlight the need to further clarify neuropsychological differences between violent versus nonviolent forms of antisocial problems within studies of schizophrenia. This is also important because the stigma against schizophrenia stems mostly from the minority of patients that show violence. Thus, interventions could be tailored to a better understanding of these subtypes.
Mood Disorders
Bipolar disorder is associated with several reckless behaviors characterized by impulsivity, as well as poor judgment and planning (Moeller, Barratt, Dougherty, Schmitz, & Swann, Reference Moeller, Barratt, Dougherty, Schmitz and Swann2001). Bipolar disorder may be associated with violence when it co-exists with substance abuse (OR: 6.4), but markedly less so when substance is absent (OR: 1.3) (Fazel, Lichtenstein, Grann, Goodwin, & Langstrom, Reference Fazel, Lichtenstein, Grann, Goodwin and Langstrom2010). Bipolar disorder has been characterized by overall impairments in neuropsychological functioning, including impairments in intelligence, attention, verbal learning and memory, executive functioning, response inhibition, working memory, set shifting, and processing speed. Some of these impairments, such as those in attention, processing speed, verbal learning/memory, and verbal fluency, have, albeit to a lesser extent, also been observed in healthy first-degree relatives, suggesting a potential trait marker for bipolar disorder (Cardenas, Kassem, Brotman, Leibenluft, & McMahon, Reference Cardenas, Kassem, Brotman, Leibenluft and McMahon2016). Whether violent bipolar patients are cognitively distinct from nonviolent patients is unknown.
The two core symptoms of the new DSM-5 Disruptive Mood Dysregulation Disorder are (a) recurrent severe temper outbursts that are out of proportion to the situation and inconsistent with the developmental stage and (b) high levels of irritability and anger for most of the day, nearly every day. Despite a conceptual and face-value relevance of this disorder to the study of violence (Mayes, Waxmonsky, Calhoun, & Bixler, Reference Mayes, Waxmonsky, Calhoun and Bixler2016) there have yet to be studies of its cognitive neuropsychology.
Conclusion
The neuropsychology of antisocial behavior has a rich history, dating back at least to the mid-nineteenth century. As our review shows, however, not as much can be said about the neuropsychology of physical violence. The main problem is that the specificity of neuropsychological deficits to physically violent behavior has been difficult to establish. The bulk of our knowledge derives from studies of clinical syndromes in which the presence of violence is plausible, but rarely confirmed. Global measures of antisocial, disruptive, externalizing, delinquent, or criminal behavior are also often used. This state of affairs reflects the heterogeneity of processes underlying those conditions.
Nonetheless, as this book shows, violence is a common outcome of a wide variety of heterogeneous conditions. Violence is present in many disorders for which there may be a partial neuropsychological basis, although that neuropsychological basis may not necessarily be for violence per se. To advance a research agenda in the study of the neuropsychology of violence, we recommend testing for the specificity of behavior problems, such as identifying physical aggression and isolating it from other co-occurring behavior problems. More explicit assessment of violence or physical aggression is needed with key contrasts to such disorders as ADHD, substance use, and to nonviolent forms of antisocial behavior. Although this approach should reduce the heterogeneity in the behavior of interest, it may not reduce the heterogeneity of underlying processes as much as one hopes; subtypes of physical violence will also need to be addressed. A well-documented history of behavior problems should also help to reduce heterogeneity. The study of the process of desistance from violence would be informative in this regard.
Our review shows that neuropsychological impairments, even in executive function, are not necessarily specific to physical aggression. We first observed that lesions among frontal lobe patients, despite their poor executive function, rarely lead to physical violence, though these lesions may lead to explicit forms of violence in the context of other risk factors. Further, there is considerable literature on other problems, such as ADHD, and their relation to executive function. Nevertheless, we noted that studies of ADHD have rarely controlled for co-occurring physical aggression. Further, neuropsychological variables tend to explain at most 8–10% of the variance in measures of violence. Therefore, studies need to examine potential moderators that may increase our ability to predict the risk of violence from neuropsychological dysfunction. Such factors could include a history of abuse or neglect, malnutrition, abilities to process and regulate emotions (including autonomic arousal), capacities to cope with stress and perceived provocation, perinatal factors, and genetic and epigenetic factors (e.g., Provençal et al., Reference Provençal, Booij and Tremblay2015). Although many of these variables have been widely studied, few have been examined in conjunction with neuropsychological function.
Furthermore, whereas neuroscience research up to the first decade of this century focused primarily on how cognitive deficits are linked to alterations in the function or structure of one or more single brain areas, cognitive theories have increasingly attempted to explain antisocial behavior-associated cognitive deficits as an alteration within a larger neural dynamic network. Greater application of advanced brain-imaging methodologies such as dynamic functional connectivity and multimodal imaging would help us in understanding the complex dynamic interplay between brain regions; for example, in how an alteration in one region could impact the function of others, and its possible consequences for violence. Neuropsychological function almost certainly comprises one element of an exceedingly complex model of violent behavior. As this chapter illustrates, however, it may provide one essential piece of a still-unsolved puzzle.
Introduction
After decades of rigid adherence to nature or nurture perspectives on human behavior, both conventional and antisocial, the contemporary scene is one where the interaction of nature and nurture is assumed and increasingly the object of scientific inquiry (DeLisi & Vaughn, Reference DeLisi and Vaughn2015; Rutter, Reference Rutter1997; Moffitt, Reference Moffitt2005). One consequence of this newer paradigm is the blending of constructs where the relative effects of biological and social phenomena have been shown to be dynamic, fluid, and interactive. A recent study is illustrative of this trend. Using data from 1,037 birth cohort participants in the Dunedin (New Zealand) Multidisciplinary Health and Development Study, Israel et al. (Reference Israel, Caspi, Belsky, Harrington, Hogan, Houts, Ramrakha, Sanders, Poulton and Moffitt2014) examined how human capital as measured by one’s credit score is associated with cardiovascular health, socioeconomic achievement, and self-control. Their findings were fascinating. Cognitive ability, self-control, and educational attainment were positively predictive of credit score and negatively predictive of cardiovascular disease, and these human capital factors accounted for nearly half of the correlation between credit score and cardiovascular disease. Although all cohort members were 38 years old at the time of the study, their heart age varied tremendously. The mean heart age was 38.5 years, which is roughly equivalent to their chronological age, but the variance in heart age was 22 to 85 years. In addition, a 100-point increase in credit score (which ranged from 12 to 961 and had a mean of 675.2) was associated with a 13-month difference in heart age. The differences in heart age, heart health, wealth, and self-control among participants were evident in the first years of life. Childhood human capital competencies in the first decade accounted for 22% of the link between credit score and cardiovascular disease at midlife.
In a related study using the same data, strong associations were found between childhood self-control and lifelong problem behaviors. For instance, about 45% of those with low childhood self-control had an adult criminal conviction. Among those with high childhood self-control, approximately 10% had an adult criminal conviction (Moffitt et al., Reference Moffitt, Arseneault, Belsky, Dickson, Hancox, Harrington and Caspi2011) – a nearly fivefold difference. Those with lower self-control during childhood also had worse physical health, greater depression, higher likelihood of drug dependence, lower socioeconomic status, lower income, greater likelihood of single-parenthood, worse financial planning, and more financial struggles at midlife.
Self-control is clearly implicated in a variety of biologically related outcomes (e.g., cardiovascular disease, heart health, mental health, and physical health) and environmentally related outcomes (e.g., socioeconomic achievement, credit score, family relationships, and crime) to such a degree that self-control – as a powerful engine of antisocial behavior – itself seems a blend of biological and social factors. It is. In their influential general theory of crime, Gottfredson and Hirschi (Reference Gottfredson and Hirschi1990) theorized that inadequate parenting practices (e.g., weak bonds to one’s child, low monitoring of a child’s activity, low oversight of a child’s activities and peers, inconsistent and/or low response to child deviance, inconsistent punishment, and others) failed to inculcate self-control (or, put another way, produced low self-control), which was characterized by low gratification delay, low tenacity, risk taking, self-centeredness, and a poor temper. These traits are associated with a broad swath of imprudent and antisocial behaviors including delinquency, crime, and violence. A large literature has supported the proposed theoretical links between parenting deficits, low self-control, and antisocial conduct (Botchkovar, Marshall, Rocque, & Posick, Reference Botchkovar, Marshall, Rocque and Posick2015; Finkenauer, Engels, & Baumeister, Reference Finkenauer, Engels and Baumeister2005; Vazsonyi & Huang, Reference Vazsonyi and Huang2010; Vazsonyi, Jiskrova, Ksinan, & Blatný, Reference Vazsonyi, Jiskrova, Ksinan and Blatný2016; Vazsonyi, Mikuška, & Kelley, Reference Vazsonyi, Mikuška and Kelley2017; Vazsonyi, Roberts, & Huang, Reference Vazsonyi, Roberts, Huang, DeLisi and Vaughn2015) such that better parenting correlates with better child self-control and worse parenting correlates with child behavioral problems.
However, studies using biosocial designs have also shown that socialization is far from the only source of self-control. Using data from the Early Childhood Longitudinal Study, Kindergarten Class of 1998–1999 (ECLS-K), Wright and Beaver (Reference Wright and Beaver2005) conducted two sets of analyses: first, where the genetic relatedness of respondents was not considered, and thus parental socialization was the focus, and second, those where genetic relatedness was controlled. In seven of the eight models where the outcome variable was child self-control in kindergarten or first grade, the number of significant parenting parameters declined. In seven of the models, the number of statistically significant parenting parameters decreased from three to either one or zero. Overall, their study demonstrated that parenting effects become much weaker, and even insignificant, once genetic factors were considered (Beaver, Connolly, Schwartz, Al-Ghamdi, & Kobeisy, Reference Beaver, Connolly, Schwartz, Al-Ghamdi and Kobeisy2013; Beaver, Wright, & DeLisi, Reference Beaver, Wright and DeLisi2007; Boisvert, Wright, Knopik, & Vaske, Reference Boisvert, Wright, Knopik and Vaske2012; Hay & Meldrum, Reference Hay and Meldrum2015). Similarly, Wright, Beaver, DeLisi, and Vaughn (Reference Wright, Beaver, DeLisi and Vaughn2008) found that parenting factors accounted for negligible variance in self-control – sometimes just 0–1% – and that genetic factors and nonshared environmental factors (those that make siblings different) accounted for the remaining variance.
In other words, forces of nature and nurture create deficits in self-control that facilitate criminal conduct across life. The current chapter highlights the interaction of nature and nurture in the etiology, maintenance, and reduction of antisocial behavior. Although self-control is a prominent area of biosocial inquiry, it is just one of many exciting and vibrant areas of the social sciences that reveal the sublime intricacy of criminal conduct.
Mechanisms of Nature–Nurture Interaction and Antisocial Behavior
Nature and nurture interact in a variety of ways or mechanisms to increase the likelihood of specific behavioral outcomes. In some cases, the forces of nurture are so noxious or constitute such an environmental pathogen that the biological functioning of an individual is altered to tremendous extent. The textbook example of this is Phineas Gage, who, in 1848, was injured in a railroad accident when a tamping iron blasted through his face and exited his head. Despite losing consciousness and suffering heavy bleeding, Gage miraculously not only survived the blast, but appeared to recover quickly. Unfortunately, the accident caused a dramatic personality transformation of Gage from a hard-working, responsible, intelligent, prudent, and socially well-adjusted person to an irreverent, impulsive, capricious, rowdy, irresponsible person whose life devolved into that of a drifter. As a result of the accident, Gage morphed from a conventionally behaved, upstanding citizen to deviant. Subsequent research of Gage’s skull revealed that his injuries were consistent with persons with similar injuries who display similar impairments in rational decision making, self-control, and emotional processing (Damasio, Grabowski, Frank, Galaburda, & Damasio, Reference Damasio, Grabowski, Frank, Galaburda and Damasio1994; Van Horn et al., Reference Van Horn, Irimia, Torgerson, Chambers, Kikinis and Toga2012).
Although the Gage example is dramatic, the broader effect of a traumatic brain injury on behavioral functioning reveals how environmental trauma can damage the brain and subsequently increase conduct problems. Behnken, DeLisi, Trulson, and Vaughn (Reference Behnken, DeLisi, Trulson, Vaughn, DeLisi and Vaughn2015) compared 132 serious delinquents who had lost consciousness due to a head injury to 588 delinquents who had not. The differences were striking. Those with prior head injury had significantly worse delinquent careers, had lower self-control, were more psychopathic, and had greater likelihood of ADHD. In addition, head injury was a significant predictor of career criminality and withstood controls for neurological medical condition, self-control, ADHD, psychopathy, age, sex, and race. Their findings were not unique; meta-analytic studies indicate that traumatic brain injury is several times more prevalent in antisocial or juvenile justice system-involved samples than the general population (Farrer, Frost, & Hedges, Reference Farrer, Frost and Hedges2012, Reference Farrer, Frost and Hedges2013; Farrer & Hedges, Reference Farrer and Hedges2011; Shiroma, Ferguson, & Pickelsimer, Reference Shiroma, Ferguson and Pickelsimer2010).
Why are head injuries so deleterious? One answer is that neuropsychological functioning is commonly reduced after sustaining a head trauma, particularly when there is also a loss of consciousness. Neuropsychological deficits figure prominently in general conceptual models of antisocial behavior (e.g., DeLisi & Vaughn, Reference DeLisi and Vaughn2014; Moffitt, Reference Moffitt1993) and numerous studies have shown that the most severe, pathological offenders tend to have the most neuropsychological deficits. In a study of serious offenders in the Pittsburgh Youth Study, Raine et al. (Reference Raine, Moffitt, Caspi, Loeber, Stouthamer-Loeber and Lynam2005) found that males on the life-course-persistent pathway had significantly greater neuropsychological deficits compared to behaviorally less severe comparison groups. Specifically, life-course-persistent offenders scored significantly worse on four measures of intelligence, two measures of spatial memory, and one measure of executive functioning. The life-course-persistent group also had a higher prevalence of ADHD diagnosis, higher child abuse victimization, higher child neglect victimization, more extreme family poverty, and had a greater number of head injuries that resulted in unconsciousness.
In a latent class analysis of data from the Early Childhood Longitudinal Survey-Kindergarten Class, Vaughn, DeLisi, Beaver, and Wright (Reference Vaughn, DeLisi, Beaver and Wright2009) found that 9.3% of kindergarteners comprised a severe impairment group characterized by deficits in verbal skills and attendant problems with higher impulsivity, higher externalizing behaviors, reduced self-regulation, reduced cognitive abilities, and greater classroom difficulties. Drawing on data from the Early Childhood Longitudinal Study: Birth Chort (ECLS-B), Jackson and Newsome (Reference Jackson and Newsome2016) recently found that infant neuropsychological deficits predicted antisocial behavior among males; however, the effects were only found for those who also had neonatal health risks. In short, injuries to the brain have the potential to reduce neuropsychological functioning and increase behavioral pathology.
Another source of environmental trauma occurs prenatally in the case of drugs, toxins, substances, or other teratogens that damage the developing embryo (for a review, see Graham, Glass, & Mattson, Reference Graham, Glass, Mattson, Beauchaine and Hinshaw2016). Unfortunately, these teratogens are readily present in the environment. In a landmark study, Wright et al. (Reference Wright, Dietrich, Ris, Hornung, Wessel, Lanphear and Rae2008) illustrated the effects of prenatal lead exposure by taking multiple measures of child lead concentrations among 250 persons recruited at birth between 1979 and 1984. The participants lived in impoverished neighborhoods in Cincinnati, characterized by a high concentration of older, lead-contaminated buildings. Those individuals with higher levels of lead in their blood as children were significantly more likely to be arrested and to be arrested for violent crimes later in life. Lead not only increased the incidence and severity of criminal behavior, but also psychopathic personality traits. Using data from the Cincinnati Lead Study, Wright, Boisvert, and Vaske (Reference Wright, Boisvert and Vaske2009) reported a significant association between blood lead concentrations at age 78 months or age 6.5 years and adult psychopathic personality features. These effects persisted despite controls for gender, race, mother’s IQ, child’s intellectual achievement, and quality of the home environment (an essential nurture variable).
Prenatal exposure to firsthand and secondhand cigarette smoke is another example of an environmental context that disrupts biological development and contributes to behavioral problems. In a recent study using data from the US Study of Early Child Care and Youth Development, Meldrum and Barnes (Reference Meldrum and Barnes2016) found that prenatal exposure to secondhand smoke was negatively associated with self-control from ages 54 months to 15 years. Moreover, the enduring damaging effects of this prenatal exposure withstood confounding effects including maternal self-control, maternal intelligence, maternal education, and maternal depression symptoms. Prenatal smoke exposure has also been linked to psychopathic personality traits (Beaver, DeLisi, & Vaughn, Reference Beaver, DeLisi and Vaughn2010), to neuropsychological deficits (Beaver, Vaughn, DeLisi, & Higgins, Reference Beaver, Vaughn, DeLisi and Higgins2010) to Conduct Disorder and related externalizing behaviors (Wakschlag, Pickett, Cook, Benowitz, & Leventhal, Reference Wakschlag, Pickett, Cook, Benowitz and Leventhal2002; Wakschlag, Leventhal, Pine, Pickett, & Carter, Reference Wakschlag, Leventhal, Pine, Pickett and Carter2006), and to the emergence of the criminal career (Gibson, Piquero, & Tibbetts, Reference Gibson, Piquero and Tibbetts2000).
A general rule is that individuals who have a biological (or genetic) risk for some outcome (or phenotype) are more likely than individuals without the biological risk factor to present with the particular phenotype. In similar fashion, individuals who have exposure to an environmental risk factor for a phenotype are more likely to display a phenotype than individuals who do not have the exposure to the environmental risk factor. It is also true that individuals with biological or genetic risks for a specific phenotype are more sensitive to the stressors of environmental risk factors – this is known as the diathesis-stress model.
In recent years, researchers have shown that individuals with putative biological risk factors can surmount their biological/genetic risks when exposed to positive and nurturing environments. In other words, biological factors and environmental factors exist on a continuum ranging from negative to positive and, depending on the type of interaction, differential behavioral outcomes can ensue. This more dynamic conceptualization is known as the differential susceptibility model (Belsky, Bakermans-Kranenburg, & van IJzendoorn, Reference Belsky, Bakermans-Kranenburg and Van IJzendoorn2007; Belsky & Pluess, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Pluess & Belsky, Reference Pluess and Belsky2011). In this model, biological/genetic risk factors are referred to as plasticity genes because their effects are variable depending on their environmental context.
Psychology and behavioral genetics have provided additional concepts that are critical for understanding the nature and nurture interplay that produces behavior. One is gene–environment correlation, where the influences of genes and environments are inextricably linked (Plomin, DeFries, & Loehlin, Reference Plomin, DeFries and Loehlin1977; Rutter et al., Reference Rutter, Dunn, Plomin, Simonoff, Pickles, Maughan and Eaves1997; Scarr & McCartney, Reference Scarr and McCartney1983). There are three types of gene–environment correlations, commonly abbreviated as rGE. Passive gene–environment correlations (passive rGEs) reveal that children receive both their genes (half from each parent) and their early-life environments from their parents. Since children resemble their parents in terms of their antisocial traits and behaviors, it is difficult to ascertain whether genetic risk factors explain a youth’s delinquency, whether the antisocial household environment is responsible, or some mixture of the two. The way to resolve this is to use an adoption study where the parents are not biologically related to the child and thus environmental effects can be accessed without being correlated to the child’s genes.
Evocative gene–environment correlations (evocative rGEs) indicate that people differentially elicit responses from the environment based upon their temperament and personality traits that are themselves heritable, or of a genetic etiology. Evocative gene–environment correlations explain why antisocial or aggressive children elicit or evoke negative and punitive reactions from care-givers just as obedient and good-natured children evoke positive and nurturing reactions from care-givers. In families with multiple children, the child who has the most disagreeable traits and the worst self-regulation is usually targeted by his parents for more severe discipline and other negative parenting behaviors. This likely is due to genetic factors that evoke the negative responses. Active gene–environment correlations are where individuals self-select environments, peers, and situations that are compatible with their own personality, temperament, and behaviors, all of which are heritable.
Active gene–environment correlations (active rGEs) are sometimes referred to as niche-picking because of the tendency for people to gravitate to others like themselves and to gravitate toward situations that are congruent with their interests. Consider the case of delinquent or antisocial peers. Highly prosocial youth are unlikely to have antisocial peers because their attitudes, beliefs, and behaviors conflict. What is acceptable or “cool” to a delinquent youth is anathema to what is acceptable or “cool” to conventional youth. Moreover, they seem like fundamentally different people and the awkwardness that antisocial and prosocial youth would hold toward one another is largely mutual. However, the dynamics of peer relations are not simply the outcome of environmental or social factors; genetic factors also play an important role. For instance, nearly two-thirds of the variance in delinquent peer association has been shown to be accounted for by genetic factors (Cleveland, Wiebe, & Rowe, Reference Cleveland, Wiebe and Rowe2005) and criminologists have shown that the dopamine transporter gene (DAT1) and brain-derive neurotropic factor (BDNF) are two of the genes implicated in delinquent peer association (Beaver, Wright, & DeLisi, Reference Beaver, Wright and DeLisi2008; Kretschmer, Vitaro, & Barker, Reference Kretschmer, Vitaro and Barker2014). These examples illustrate the interplay of active rGEs.
Another concept that reveals nature–nurture interaction is gene x environment (G x E, sometimes referred to as molecular genetic association, or candidate gene x environment, or cG x E) studies, where measured genes and measured environmental conditions are included in the same statistical models. Gene x environment studies are among the most scientifically exciting studies because they provide estimates of the ways that nature and nurture interact to produce or insulate from antisocial behavior. Due to space constraints, this review of gene–environment studies does not purport to be exhaustive. Readers are encouraged to read a meta-analysis of the serotonin transporter gene and antisocial conduct (Tielbeek et al., Reference Tielbeek, Karlsson Linnér, Beers, Posthuma, Popma and Polderman2016), a meta-analysis of the dopamine receptor D4 gene and externalizing behavior (Pappa, Mileva-Seitz, Bakermans-Kranenburg, Tiemeier, & van IJzendoorn, Reference Pappa, Mileva-Seitz, Bakermans-Kranenburg, Tiemeier and van IJzendoorn2015), a chapter on MAOA and antisocial conduct in males (Holland & DeLisi, Reference Holland, DeLisi, DeLisi and Vaughn2015), a systematic review of genetic association studies of aggression (Fernandez-Castillo & Cormand, Reference Fernàndez-Castillo and Cormand2016; Veroude et al., Reference Veroude, Zhang-James, Fernàndez-Castillo, Bakker, Cormand and Faraone2016), and systematic reviews of the effects of MAOA, DRD2, DRD4, DAT1, 5HTTLPR, and COMT and externalizing behaviors (Samek et al., Reference Samek, Bailey, Hill, Wilson, Lee, Keyes and McGue2016; Weeland, Overbeek, Orobio de Castro, & Matthys, Reference Weeland, Overbeek, de Castro and Matthys2015) for greater coverage. Some of these studies are reviewed next.
Gene x Environment Studies of Antisocial Behavior
Environmental Moderators Predicting Antisocial Behavior
Diathesis-stress and differential susceptibility models posit that individuals with biological or genetic risks and with environmental risks are most likely to engage in diverse forms of antisocial conduct. The seminal work by Caspi et al. (Reference Caspi, McCray, Moffitt, Mill, Martin, Craig and Poulton2002) was the first criminological G x E study to model genetic and environmental conditions in the creation of antisocial behavior. Using the Dunedin birth cohort data, Caspi and colleagues examined the moderation of childhood maltreatment by variants in the MAOA gene. Among their participants between the ages of 3 and 11, 8% had been severely maltreated, 28% experienced probable maltreatment, and 64% experienced no maltreatment. Dramatic interactive effects were found between low MAOA activity and maltreatment in the prediction of Conduct Disorder, violent convictions, violent disposition, and symptoms of Antisocial Personality Disorder. For example, 80% of youth with MAOA risk (low-activity) alleles and severe maltreatment had Conduct Disorder and 30% were convicted of a violent crime. Comparatively, among those at genetic risk with no maltreatment, 20% had Conduct Disorder and 5% were convicted of a violent crime. About 85% of males with both genetic and environmental risk factors displayed some form of antisocial behavior.
Several independent research teams using their own genetically sensitive data largely replicated the MAOA-maltreatment-crime sequela (Byrd & Manuck, Reference Byrd and Manuck2014; Choe, Shaw, Hyde, & Forbes, Reference Choe, Shaw, Hyde and Forbes2014; Taylor & Kim-Cohen, Reference Taylor and Kim-Cohen2007; Thibodeau, Cicchetti, & Rogosch, Reference Thibodeau, Cicchetti and Rogosch2015). In a longitudinal study of nearly 400 male participants from the Christchurch (New Zealand) Health and Development Study followed through age 30, Fergusson, Boden, Horwood, Miller, and Kennedy (Reference Fergusson, Boden, Horwood, Miller and Kennedy2011, Reference Fergusson, Boden, Horwood, Miller and Kennedy2012) linked MAOA and childhood exposure to sexual and physical abuse to hostility, conduct problems, property offending, and violent offending. Relying on data from the Minnesota Twin Family Study, Derringer, Krueger, Irons, and Iacono (Reference Derringer, Krueger, Irons and Iacono2010) found that persons who had experienced childhood sexual assault victimization and had low-activity alleles of MAOA displayed more antisocial behaviors and evinced more symptoms of Conduct Disorder than peers with other MAOA variants. Based on data from the Virginia Twin Study for Adolescent Behavioral Development, Foley, Eaves, Wormley, Silberg, Maes, et al. (Reference Foley, Eaves, Wormley, Silberg, Maes, Kuhn and Riley2004) demonstrated an MAOA-childhood adversity defined by interparental violence, parental neglect, and inconsistent discipline interaction in the prediction of Conduct Disorder. Among boys with low-activity MAOA alleles with the highest childhood adversity exposure, 100% had Conduct Disorder. Among those at genetic risk with the lowest level of environmental exposure, only about 5% had Conduct Disorder.
Drawing on data from the Environmental Risk (E-Risk) Longitudinal Twin Study, Kim-Cohen et al. (Reference Kim-Cohen, Caspi, Taylor, Williams, Newcombe, Craig and Moffitt2006) compared the interaction between MAOA genotype, early-life exposure to physical abuse, and various pediatric mental health and behavioral outcomes. On every measure, low-activity MAOA genotype among abused children was associated with significantly worse outcomes, and the effect sizes were large. Children had more mental health problems, more antisocial behaviors, more ADHD symptoms, and more emotional problems.
To date, a variety of environmental conditions have been shown to interact with or moderate genetic factors in producing antisocial conduct. Although the 7R allele of DRD4 is commonly the risk allele of the dopamine receptor D4 gene, other alleles have also been shown to be predictive of conduct problems. Drawing on data from the Tracking Adolescents’ Individual Lives Survey from the Netherlands, Kretschmer, Dijkstra, Ormel, Verhulst, and Veenstra (Reference Kretschmer, Dijkstra, Ormel, Verhulst and Veenstra2013) found that those with the 4R allele who had lower social well-being and greater peer victimization were more likely to engage in later delinquency compared to those with the 7R allele. Using data from the Avon Longitudinal Study of Parents and Children, Kretschmer, Vitaro, and Barker (Reference Kretschmer, Vitaro and Barker2014) found that carriers of the BDNF Met-Met variant who affiliated with aggressive peers at age 10 were more likely to be aggressively delinquent at age 15 compared to those with the BDNF val-val allele.
In addition to genetic factors predicting antisocial conduct, there is also evidence that genetic factors can significantly increase the likelihood that an individual will experience assorted forms of abuse, neglect, and victimization. Using the Add Health data, Beaver et al. (Reference Beaver, Wright, DeLisi, Daigle, Swatt and Gibson2007) reported that delinquent peers and DRD2 interacted to increase the criminal victimization among adolescents. Linkages have also been shown between DAT1 polymorphisms and childhood sexual abuse victimization among men and DAT1 and childhood emotional abuse and women (Rehan et al., Reference Rehan, Antfolk, Johansson, Aminoff, Sandnabba, Westberg and Santtila2016). In other words, environmental moderation of genetic factors can increase the likelihood of both offending and victimization, often in the same individual.
Environmental Buffers against Antisocial Behavior
A variety of molecular genetic association studies have also modeled ways that genetic variants or polymorphisms interact with environmental conditions to insulate individuals from conduct problems even when the individual has genetic vulnerability. Attachment to parents has been shown to be strongly associated with antisocial development. Kochanska, Philibert, and Barry (Reference Kochanska, Philibert and Barry2009) examined the polymorphic serotonin transporter gene (5HTTLPR) and its interaction with a child’s maternal attachment to study self-regulation among youth at ages 15 months, 25 months, 38 months, and 52 months. Children with short alleles of 5HTTLPR have higher and more unstable levels of synaptic serotonin. Among those who were insecurely attached to their mother, there were self-regulation problems. Among those who were securely attached, there were no self-regulation problems. In their study using data from a Swedish population-based cohort, Tuvblad et al. (Reference Tuvblad, Narusyte, Comasco, Andershed, Andershed, Colins and Nilsson2016) found that homozygous Val allele carriers of the COMT gene had lower levels of physical aggression when they were exposed to violence and when they experienced a positive relationship with their parents compared to carriers of the Met allele of COMT. In this case, the parental relationship trumped the noxious effects of violence exposure to reduce aggressive conduct. Using data from the National Longitudinal Study of Adolescent to Adult Health, Roettger, Boardman, Harris, and Guo (Reference Roettger, Boardman, Harris and Guo2016) reported that the 2R allele of the MAOA gene was directly associated with delinquency. However, those with the 2R allele who had high closeness to their father were less likely to be delinquent (interestingly, no buffer effect was found for those who were close to their mother).
Environmental protective factors are also helpful for understanding sex differences in antisocial conduct vis-à-vis their genetic underpinnings. Drawing on data from adolescents in Russia, Dmitrieva, Chen, Greenberger, Ogunseitan, and Ding (Reference Dmitrieva, Chen, Greenberger, Ogunseitan and Ding2011) found that males with the 7R allele of DRD4 had greater involvement in delinquency and more antisocial personality features, including short temper and thrill-seeking. Females with genetic risks did not have these behaviors, in part because they had greater parental monitoring and lower exposure to violence. Once parental monitoring and violence exposure were controlled, the sex differences in delinquency were no longer significant.
Other research reveals targets for interventions that can reduce problem behaviors in children who evince genetic risks for antisocial conduct. For instance, Jackson and Beaver (Reference Jackson and Beaver2015) found that meal deprivation was associated with verbal deficits and psychopathic personality traits and poor nutrition quality was linked to verbal deficits among youth in the National Longitudinal Study of Adolescent to Adult Health. Gene–environment interactions between MAOA and measures of food quality were detected. Their findings readily show that interventions that increase the amount and quality of nutrition that children receive can improve their verbal/cognitive ability and render their personality less antisocial. The findings put the simplicity of donating food to a food bank in a new context when one considers the downstream behavioral benefits.
Direct Genetic Effects on Antisocial Behavior
Although nature–nurture interplay is the norm in contemporary studies of antisocial conduct, it is also important to observe that genetic effects often have direct effects on antisocial phenotypes, albeit the effects usually have a small effect size – known as the fourth law of behavior genetics (Chabris, Lee, Cesarini, Benjamin, & Laibson, Reference Chabris, Lee, Cesarini, Benjamin and Laibson2015). There are several examples of these direct effects. Caspi et al. (Reference Caspi, Langley, Milne, Moffitt, O’Donovan, Owen and Williams2008) examined the association between the COMT Val158Met and antisocial behavior among three samples of children with ADHD. The samples included the Cardiff ADHD Genetic Study, which is a sample of 376 white British children selected from child psychiatry and pediatric clinics in England and Wales between 1997 and 2003. The second sample was the Environmental Risk (E-Risk) Study, which is a birth cohort of 2,232 British children drawn from the 1994 to 1995 birth registry in England and Wales, and the third sample was the Dunedin Longitudinal Study described at the outset of this chapter. Caspi and his colleagues (Reference Caspi, Langley, Milne, Moffitt, O’Donovan, Owen and Williams2008) found across the three samples that children with the Valine/Valine (Val/Val) homozygotes had more symptoms of Conduct Disorder, were more aggressive, and were more likely to be convicted of crimes than Methionine carriers (Met/Met or Val/Met). Barkley, Smith, Fischer, and Navia (Reference Barkley, Smith, Fischer and Navia2006) found that the homozygous DBH Taq1 A2 allele and the dopamine transporter gene (DAT1) were associated with greater hyperactivity during childhood and pervasive behavioral problems during adolescence among respondents from a Milwaukee longitudinal study of hyperactive and normal-activity children.
Myriad direct genetic effects have been shown for other forms of antisocial behavior. These include polymorphisms in the vasopressin 1B receptor gene (AVP 1B), aggression, and conduct problems among clinically referred children (Luppino, Moul, Hawes, Brennan, & Dadds, Reference Luppino, Moul, Hawes, Brennan and Dadds2014), the rs1465108 polymorphism in the MAOA gene, aggression, and negative urgency (Chester et al., Reference Chester, DeWall, Derefinko, Estus, Peters, Lynam and Jiang2015), low-activity alleles of MAOA and violent crime (Stetler et al., Reference Stetler, Davis, Leavitt, Schriger, Benson, Bhakta and Bortolato2014), and several serotonergic genes and aggressive behavior (Chen et al., Reference Chen, Liu, Chen, Moyzis, Chen and Dong2015). Finally, most molecular genetic association studies include aggression or other moderately serious forms of antisocial conduct as outcome variables of phenotypes. However, research has also shown direct genetic effects for more extreme forms of antisocial behavior. A study of prisoners in Finland found that low-activity variants of the MAOA gene were associated with being in the 90th percentile on a violent offending distribution, which equates to at least ten convictions for serious criminal violence. The 78 offenders who met this threshold committed 1,154 murders, attempted murders, and aggravated assaults (Tiihonen et al., Reference Tiihonen, Rautiainen, Ollila, Repo-Tiihonen, Virkkunen, Palotie and Paunio2015).
The takeaway point from molecular genetic research is that investigators have come a long way from philosophical debates about the relative merits of nature and nurture to highly sophisticated quantitative estimates of the precise effects of genetic variants, including particular alleles and the environmental contexts in which these genetic vulnerabilities are enflamed into antisocial conduct or suppressed into normative behavior.
Discussion
The nature–nurture interplay in the creation of antisocial conduct is intellectually exciting and scientifically impressive, but what is the practical utility in terms of behavioral interventions and policy? Although the nature–nurture or biosocial vernacular was not originally part of most primary, secondary, and tertiary prevention programs, most have nevertheless included targets that fit well within a nature–nurture understanding of the development of antisocial conduct. Indeed, in their biosocial reinterpretation of cognitive behavioral therapy, Vaske, Galyean, and Cullen (Reference Vaske, Galyean and Cullen2011, p. 90), observed that programs that target “social skills, coping skills, and problem-solving skills are consistently associated with activation in the medial prefrontal cortex, dorsolateral prefrontal cortex, dorsomedial prefrontal cortex, ventromedial prefrontal cortex, orbitofrontal cortex, cingulate cortex, insula, and temporo-parietal junction.” In other words, treatment and correctional interventions that attempt to reduce delinquency and conduct problems might seem like social/environmental endeavors, but they also explicitly target the brain to improve neuropsychological and, hence, behavioral functioning.
This is seen especially in programs that seek to improve executive functioning. The basic logic of targeting neuropsychological deficits relating to self-control is at the core of the Promoting Alternative THinking Strategies (PATHS) Curriculum. Designed for school entry through fifth grade, PATHS is a comprehensive educational program taught three times per week for a minimum of 20 minutes per day. Among its targets are instruction on delaying gratification, controlling impulses, self-talk, self-awareness, reading and interpreting social cues and the needs/perspective of others, verbal skills, nonverbal communication, problem solving, and decision making. Improvements in these target areas contribute to an assortment of positive program outcomes, including improved self-control, reduced conduct problems, reduced anxiety/depression symptoms, and reduced aggression, and PATHS is hailed as a model prevention program (Greenberg, Kusche, & Mihalic, Reference Greenberg, Kusche and Mihalic2006).
Relatedly, Castellanos-Ryan, Séguin, Vitaro, Parent, and Tremblay (Reference Castellanos-Ryan, Séguin, Vitaro, Parent and Tremblay2013) conducted a randomized controlled trial for behaviorally disordered kindergarten boys using data from the Montreal Longitudinal and Experimental Study. The two-year intervention targeted social and problem-solving skills among the boys and training on effective child-rearing for their parents. Eight years after the program, boys in the experimental condition had fewer drug-related delinquency problems and a main reason was due to reductions in impulsivity. Other researchers have shown a gene x intervention interaction where variants of the brain-derived neurotrophic factor (BDNF) gene interacted with an impulsivity school-based program to produce reductions in aggression among youth (Musci et al., Reference Musci, Bradshaw, Maher, Uhl, Kellam and Ialongo2014). Originally used to study antisocial conduct, genetically informed research findings are now being incorporated into prevention programs (Gajos, Fagan, & Beaver, Reference Gajos, Fagan and Beaver2016).
To conclude, the notion of nature–nurture interplay has existed for decades in psychiatry, genetics, and psychology and has more recently diffused to criminology, criminal justice, and even sociology. Environmental and biological factors are not independent silos, but instead exert independent, interactive, and reciprocal effects, particularly when the phenotype of interest is antisocial behavior. The last decade or so has witnessed a flurry of studies identifying numerous candidate genes for antisocial behavior. The next steps are replication of these early findings so that behavioral and pharmaceutical interventions can be devised to reduce the incidence and severity of crime.
Introduction
Bullying is a subtype of aggression that is characterized by intentionality, repetition, and an imbalance of power (Olweus, Reference Olweus, Smith, Morita, Junger-Tas, Olweus, Catalano and Slee1999). Bullying takes many forms including verbal, physical, relational (i.e., traditional bullying), and cyber. Most population-based studies indicate that bullying affects about 30% of children and youth worldwide (Nansel et al., Reference Nansel, Overpeck, Pilla, Ruan, Simons-Morton and Scheidt2001; National Academies of Sciences, Engineering, and Medicine, 2016; UNICEF, 2013; Vaillancourt et al., Reference Vaillancourt, Brittain, Bennett, Arnocky, McDougall, Hymel and Cunningham2010a), making it the most prevalent form of aggression youth are exposed to. Being the target of bullying is associated with significant mental health issues, which persist long after the bullying has stopped (Lereya, Copeland, Costello, & Wolke, Reference Lereya, Copeland, Costello and Wolke2015; Takizawa, Maughan, & Arseneault, Reference Takizawa, Maughan and Arseneault2014; see review by McDougall & Vaillancourt, Reference McDougall and Vaillancourt2015). There is also evidence that being the target of peer bullying in childhood has a more pronounced negative impact on adult mental health than being exposed to childhood maltreatment (Lereya et al., Reference Lereya, Copeland, Costello and Wolke2015).
Research on perpetrators of bullying suggests different outcomes than that of targets. Whereas targets of bullying tend to be marginalized, rejected, and lonely (Knack, Tsar, Vaillancourt, Hymel, & McDougall, Reference Knack, Tsar, Vaillancourt, Hymel and McDougall2012; Nansel et al., Reference Nansel, Overpeck, Pilla, Ruan, Simons-Morton and Scheidt2001), several studies have shown that perpetrators of bullying often wield considerable power and influence in their peer groups and that many are the most popular children and adolescents in their schools (Faris & Felmlee, Reference Faris and Felmlee2011; Farmer, Estell, Bishop, O’Neal, & Cairns, Reference Farmer, Estell, Bishop, O’Neal and Cairns2003; Vaillancourt, Hymel, & McDougall, Reference Vaillancourt, Hymel and McDougall2003). Longitudinal studies of perpetrators point to a problematic developmental trajectory punctuated by problems with later offending (Ttofi, Farrington, Lösel, & Loeber, Reference Ttofi, Farrington, Lösel and Loeber2011) and substance abuse (Ttofi, Farrington, Lösel, Crago, & Theodorakis, Reference Ttofi, Farrington, Lösel, Crago and Theodorakis2016). However, when controlling for family hardship and childhood psychiatric disorders, known predictors of aggression, the negative impact of bullying perpetration on adult health, wealth, crime, and social outcomes is not found (Wolke, Copeland, Angold, & Costello, Reference Wolke, Copeland, Angold and Costello2013), nor is it found for most adult psychiatric outcomes (Copeland, Wolke, Angold, & Costello, Reference Copeland, Wolke, Angold and Costello2013). Vaillancourt, Clinton, McDougall, Schmidt, and Hymel (Reference Vaillancourt, Clinton, McDougall, Schmidt, Hymel, Jimerson, Swearer and Espelage2010b) have argued that such discrepancies in outcomes should be expected because there are in fact two general types of bullies – high-status perpetrators and low-status perpetrators. High-status bullies have assets and competencies that the peer group values, such as being good athletes and being attractive (Vaillancourt et al., Reference Vaillancourt, Hymel and McDougall2003). These perpetrators engage in a mélange of prosocial and antisocial behavior (i.e., they are Machiavellian) and the aggression they direct at peers is instrumental insofar as it is used to achieve and maintain hegemony. Most perpetrators of bullying are high-status (Vaillancourt et al., Reference Vaillancourt, Hymel and McDougall2003). Low-status bullies tend to have few assets and competencies that the peer group value (Vaillancourt et al., Reference Vaillancourt, Hymel and McDougall2003), they are more reactive in their use of aggression, they tend to have problems with emotional self-regulation, and they are oftentimes life-course persistent in their use of aggression. Many low-status perpetrators are also victims of bullying (Vaillancourt et al., Reference Vaillancourt, Clinton, McDougall, Schmidt, Hymel, Jimerson, Swearer and Espelage2010b).
When targets and perpetrators are compared on different outcomes, victims invariably fare worse than bullies (Copeland et al., Reference Copeland, Wolke, Angold and Costello2013; Wolke et al., Reference Wolke, Copeland, Angold and Costello2013), and are thus the focus of this chapter. This discrepancy in outcomes is likely due to the fact that being the victim of bullying interferes with the fundamental need to belong (Baumeister & Leary, Reference Baumeister and Leary1995), whereas being the perpetrator of bullying does not. For example, Nansel et al. (Reference Nansel, Overpeck, Pilla, Ruan, Simons-Morton and Scheidt2001) reported in their large population-based study of American youth that the ability to make friends was negatively associated with being the victim of bullying, but positively associated with being the perpetrator of bullying.
In addition to the robust literature documenting the negative effects of bullying victimization on mental health, there is also a growing literature suggesting that the effects of bullying are more pernicious than previously thought. Specifically, recent studies have shown that bullying can “get under the skin” and cause biological changes that increase the risk of poorer outcomes (see Vaillancourt et al., Reference Vaillancourt, Clinton, McDougall, Schmidt, Hymel, Jimerson, Swearer and Espelage2010b; Vaillancourt, Hymel, & McDougall, Reference Vaillancourt, Hymel and McDougall2013; Vaillancourt, Sanderson, Arnold, & McDougall, Reference Vaillancourt, Sanderson, Arnold, McDougall and Bradshaw2017 for reviews). Specifically, converging evidence suggests that early-life adversity, which includes being the target of bullying, “programs physiology and behavior” (Shalev & Belsky, Reference Shalev and Belsky2016, p. 41). The focus of this chapter is on reviewing the current state of knowledge about the neurobiological effects of bullying victimization and its links to mental health.
Stress-Response System
It is well-established in animal and human studies that early adversity is associated with changes to the neuroendocrine stress-response system and that these changes are in turn linked to disease (Lupien, McEwen, Gunna, & Heim, Reference Lupien, McEwen, Gunnar and Heim2009; Miller, Chen, & Parker, Reference Miller, Chen and Parker2011). The most widely studied stress-response system is the hypothalamic–pituitary–adrenal (HPA) axis. The HPA axis helps the organism adapt to stressors, thus supporting homeostasis and promoting health (McEwen, Reference McEwen2004).
When a person is exposed to psychological or physical stressors, a biological cascade is initiated that begins with an increased production and secretion of corticotropin-releasing hormone (CRH) released from the paraventricular nucleus of the hypothalamus. CRH travels through the hypophyseal portal circulation and stimulates the anterior pituitary gland to release adrenocorticotropic hormone (ACTH). ACTH is carried through the peripheral circulation to the adrenal cortex where it triggers the production and release of cortisol, a glucocorticoid with widespread regulatory influence that helps mobilize energy needed to meet the demands of a stressor. Following activation of this system, cortisol acts on the pituitary gland, the hypothalamus, and the hippocampus as a negative-feedback inhibition, suppressing this hormonal pathway, and thus protecting the individual against the damaging effects of chronic activation of the HPA axis (Jacobson & Sapolsky, Reference Jacobson and Sapolsky1991; Sapolsky, Krey, & McEwen, Reference Sapolsky, Krey and McEwen1986).
Several psychiatric disorders have been implicated in the dysfunction of the HPA axis, and, in particular, its end product cortisol (Ehlert, Reference Ehlert2013). Specifically, researchers have focused on the dysregulation of cortisol, including its hyper-secretion or hypo-secretion, and its pattern across the day and after exposure to an acute stressor. For example, although cortisol typically follows a diurnal pattern, peaking 20–30 minutes after awakening and gradually dropping throughout the day (i.e., cortisol awakening response), in some individuals, cortisol levels are consistently high throughout the day. Moreover, in other individuals, exposure to a stressor is associated with an increase in cortisol that does not abate as would be expected. Cortisol dysregulation may result because of a failure to “(a) habituate to recurring stress, (b) inhibit allostatic processes following termination of stress, or (c) mount an adaptive response in some systems that can lead to the hyperactivation of others” (Morris, Compas, & Garber, Reference Morris, Compas and Garber2012, p. 303).
Although elevated cortisol levels are viewed as adaptive when faced with a stressor, extended activation of the HPA axis has been shown to have adverse effects, termed allostatic load (McEwen, Reference McEwen1998). The hyper-secretion of cortisol is often associated with the pathogenesis of major depression (Stetler & Miller, Reference Stetler and Miller2011) and is presumed to be due to an insensitive negative glucocorticoid feedback of the HPA axis loop (Ehlert, Reference Ehlert2013, p. 1852). Exposure to prolonged and/or severe stress may also lead to a different type of “adaptive” change, which alters the response of the HPA axis in a different direction (Tyrka, Ridout, & Parade, Reference Tyrka, Ridout and Parade2016 for review). Specifically, constant activation of the HPA axis in response to stress can cause a counter-regulatory state leading to the hypo-secretion of cortisol (Tyrka et al., Reference Tyrka, Ridout and Parade2016). The hypo-secretion of cortisol is often linked to stress-related disorders like post-traumatic stress disorder (PTSD; Morris et al., Reference Morris, Compas and Garber2012) and is presumed to be due to “a negative feedback hypersensitivity of glucocorticoids associated with an up-regulated leukocyte glucocorticoid receptor (GR) number and sensitivity” (Ehlert, Reference Ehlert2013, p. 1852). Glucocorticoids are a class of steroid hormones known as corticosteroids that bind with the glucocorticoid receptor. They are implicated in the immune system as part of a feedback mechanism that is associated with the reduction of functions like inflammation.
Although exposure to adversity can be associated with an exaggerated or blunted neuroendocrine response to stress, much of the variability in HPA axis activity has been shown to be attributable to the features of the stressor and the person. For example, results of Miller, Chen, and Zhou’s (Reference Miller, Chen and Zhou2007) meta-analysis indicated that cortisol tended to be high at stressor onset and attenuated over time. Stressors that were extreme in nature such as those involving trauma or threatening physical integrity tended to elicit a high, flat diurnal cortisol pattern. This was also true of uncontrollable stressors. Although subjective distress elicited heightened cortisol, in individuals with PTSD, cortisol levels were lower. Low cortisol has been linked to extreme traumas that are associated with the development of PTSD such as exposure to early sexual abuse (King et al., Reference King, Schwab-Stone, Flisher, Greenwald, Kramer, Goodman and Gould2001) and being a survivor of the Holocaust (Yehuda et al., Reference Yehuda, Kahana, Binder-Brynes, Southwick, Mason and Giller1995). Being the target of peer bullying is also linked to PTSD symptoms (Idsoe, Dyregrov, & Idsoe, Reference Idsoe, Dyregrov and Idsoe2012; Litman et al., Reference Litman, Costantino, Waxman, Sanabria-Velez, Rodriguez-Guzman, Lampon-Velez and Cruz2015), and consistent with the notion that bullying is a form of trauma (Mishna, Reference Mishna, Bussey and Bula Wise2007), to a blunted cortisol response in children (Carney, Hazler, Oh, Hibel, & Granger, Reference Carney, Hazler, Oh, Hibel and Granger2010; Kliewer, Reference Kliewer2006; Kliewer, Reference Kliewer2016; Ouellet-Morin et al., Reference Ouellet-Morin, Danese, Bowes, Shakoor, Ambler, Pariante and Arseneault2011a; Ouellet-Morin et al., Reference Ouellet-Morin, Odgers, Danese, Bowes, Shakoor, Papadopoulos and Arseneault2011b; Ouellet-Morin et al., Reference Ouellet-Morin, Wong, Danese, Pariante, Papadopoulos, Mill and Arseneault2013; Vaillancourt et al., Reference Vaillancourt, Duku, Decatanzaro, MacMillan, Muir and Schmidt2008) and adults (Hansen et al., Reference Hansen, Hogh, Persson, Karlson, Garde and Ørbæk2006; Hansen, Hogh, & Persson, Reference Hansen, Hogh and Persson2011). This blunted response to stress among bullied youth has been linked to the development of social and behavioral problems (Ouellet-Morin et al., Reference Ouellet-Morin, Odgers, Danese, Bowes, Shakoor, Papadopoulos and Arseneault2011b).
Bullying victimization is also associated with an atypical cortisol awakening response and an atypical response to acute stress. For example, Knack, Jensen-Campbell, and Baum (Reference Knack, Jensen-Campbell and Baum2011) found that while the diurnal pattern for nonbullied adolescents was characteristic (i.e., cortisol levels peaking 30 minutes after waking and then decreasing across the day), for bullied adolescents, an atypical pattern was found. Bullied adolescents had lower levels of cortisol 30 minutes after waking and 30 minutes before bed than their nonbullied peers. Knack et al. also found that exposure to an acute stressor (i.e., Trier Social Stress Test) in nonbullied adolescents resulted in a peak in cortisol 30 minutes after delivering the speech that was sustained 30 minutes post-speech, but in bullied adolescents, a significant drop in cortisol was found 30 minutes following the speech delivery. This fall in cortisol in bullied youth was associated with more frequent visits to the doctor. Although the drop in cortisol post-stressor among bullied youth was unexpected it was notably similar to the pattern reported by Ouellet-Morin et al. (Reference Ouellet-Morin, Odgers, Danese, Bowes, Shakoor, Papadopoulos and Arseneault2011b), who found that nonabused children showed elevated cortisol levels post-stress delivery (adapted Trier Social Stress Test), while maltreated/bullied children showed decreasing levels in cortisol post-stress delivery. In another study involving monozygotic twins discordant on bullying victimization, Ouellet-Morin et al. (Reference Ouellet-Morin, Danese, Bowes, Shakoor, Ambler, Pariante and Arseneault2011a) found that bullied twins showed a decrease in cortisol post-stress delivery, while nonbullied twins showed an increase in cortisol post-stress delivery. These differences between twins were not due to genetics, familial environment, or individual factors. Nor were they due to the perception of stress or the emotional response associated with the stressor. Rather, the results provided “support for a causal effect of adverse childhood experiences on the neuroendocrine response to stress” (p. 2011).
Atypical diurnal patterns have also been noted in bullied youth. González-Cabrera, Calvete, León-Mejía, Pérez-Sancho, and Peinado (Reference González-Cabrera, Calvete, León-Mejía, Pérez-Sancho and Peinado2017) found that cortisol concentrations across the day were flatter for adolescent victims of severe cyberbullying. Being the victim of cyberbullying was also linked with a larger area under the curve (i.e., total cortisol output over the day), which, in turn, was associated with higher perceived stress and anxiety. Brendgen, Ouellet-Morin, Lupien, Vitaro, Dionne, and Boivin (Reference Brendgen, Ouellet-Morin, Lupien, Vitaro, Dionne and Boivin2017) found that controlling for awakening levels of cortisol, twins who were more victimized by their peers than their co-twin demonstrated a steeper decline of cortisol until bedtime. The results of these studies are consistent with the idea that exposure to stressors that are extreme and/or prolonged in nature are associated with a down-regulation of the stress response system (Miller et al., Reference Miller, Chen and Zhou2007).
Finally, there is evidence that the hyper-secretion of cortisol is linked to peer victimization via depression. Depression is reliably linked to higher cortisol levels (Stetler & Miller, Reference Stetler and Miller2011). Vaillancourt et al. (Reference Vaillancourt, Duku, Becker, Schmidt, Nicol, Muir and MacMillian2011) examined adolescents every six months on four occasions and found that being the target of bullying in childhood was related to increases in depression symptoms, which, in turn, was related to increases in basal cortisol. Notably, increased cortisol was related with impaired memory associated with areas of the brain that are rich in glucocorticoid receptor sites – the hippocampus and the prefrontal cortex (Lupien et al., Reference Lupien, Fiocco, Wan, Maheu, Lord, Schramek and Tu2005). Cortisol binds to the glucocorticoid receptor and, in doing so, regulates gene transcription, which is the first step in the expression of a gene. This finding is in keeping with a well-replicated literature involving animals and humans demonstrating that high levels of cortisol are bad for the brain. Specifically, high exposure to glucocorticoids is associated with memory impairment and even atrophy of the hippocampus (e.g., Lupien et al., Reference Lupien, DeLeon, DeSanti, Convit, Tarshish, Nair and Meaney1998).
Taken together, studies examining the relation between being bullied and the stress-response system suggest a cortisol profile that is similar to what is seen among survivors of extreme trauma that often go on to receive a diagnosis of PTSD. Perhaps not surprising is the strong link between bullying victimization and PTSD (Idsoe et al., Reference Idsoe, Dyregrov and Idsoe2012; Litman et al., Reference Litman, Costantino, Waxman, Sanabria-Velez, Rodriguez-Guzman, Lampon-Velez and Cruz2015). For example, Idsoe et al. (Reference Idsoe, Dyregrov and Idsoe2012) reported that 27.6% of boys and 40.5% of girls in their large study of Norwegian adolescents scored within the clinical range on PTSD symptoms.
Inflammation
Exposure to psychosocial stressors like childhood trauma has also been shown to influence circulating inflammatory markers such as C-reactive protein, interleukin-6, and tumor necrosis factor-α (Baumeister, Akhtar, Ciufolini, Pariante, & Mondelli, Reference Baumeister, Akhtar, Ciufolini, Pariante and Mondelli2016). This relation is distinguished because inflammation has been shown to play a vital role in mental disorders (Baumeister, Russell, Pariante, & Mondelli, Reference Baumeister, Russell, Pariante and Mondelli2014). For example, peripheral inflammation as indexed by plasma C-reactive protein has been linked to the development of PTSD (Eraly, Nievergelt, & Maihofer, Reference Eraly, Nievergelt and Maihofer2014).
Considering that bullying victimization is a form of trauma (Mishna, Reference Mishna, Bussey and Bula Wise2007), it is not surprising that recent studies have shown longitudinal links between bullying in childhood and inflammation in adulthood. For example, Copeland et al. (Reference Copeland, Wolke, Lereya, Shanahan, Worthman and Costello2014) found in their longitudinal population-based study that, when compared with those not involved in bullying, being the target of bullying in childhood predicted greater C-reactive protein levels in adulthood, while being the perpetrator of bullying in childhood predicted lower C-reactive protein levels in adulthood. Moreover, cumulative experiences with bullying victimization predicted increases in C-reactive protein levels even when controlling for relevant covariates that have been shown to be associated with C-reactive protein levels and involvement with bullying. In another population-based study of individuals followed prospectively for 50 years, Takizawa, Danese, Maughan, and Arsenault (Reference Takizawa, Danese, Maughan and Arseneault2015) found that being the target of frequent bullying in childhood was associated with increased levels of C-reactive protein in mid-life. And in a recent study of adolescents, Arana et al. (in press) reported that relational peer victimization (e.g., rumour spreading, peer group exclusion) was associated with higher levels of depression, somatic complaints, and inflammation. Specifically, relational peer victimization indirectly influenced interleukin-6 through depression and C-reactive protein through depression and interleukin-6.
The results of these studies are interesting because there is evidence for a bidirectional association between the inflammatory system and other related systems concerned with the pathogenesis of mental disorders, such as the HPA axis (Baumeister et al., Reference Baumeister, Russell, Pariante and Mondelli2014; Miller, Maletic, & Raison, Reference Miller, Maletic and Raison2009). In particular, cortisol has been shown to have an anti-inflammatory effect (Straub, Buttgereit, & Cutolo, Reference Straub, Buttgereit and Cutolo2011), such that lower cortisol levels are associated with higher levels of inflammation and higher cortisol levels are associated with lower levels of inflammation. As mentioned, researchers have typically reported an association between blunted cortisol and bullying victimization (Carney et al., Reference Carney, Hazler, Oh, Hibel and Granger2010; Hansen et al., Reference Hansen, Hogh, Persson, Karlson, Garde and Ørbæk2006; Hansen et al., Reference Hansen, Hogh and Persson2011; Kliewer, Reference Kliewer2006; Kliewer, Reference Kliewer2016; Ouellet-Morin et al., Reference Ouellet-Morin, Danese, Bowes, Shakoor, Ambler, Pariante and Arseneault2011a; Ouellet-Morin et al., Reference Ouellet-Morin, Odgers, Danese, Bowes, Shakoor, Papadopoulos and Arseneault2011b; Ouellet-Morin et al., Reference Ouellet-Morin, Wong, Danese, Pariante, Papadopoulos, Mill and Arseneault2013; Vaillancourt et al., Reference Vaillancourt, Duku, Decatanzaro, MacMillan, Muir and Schmidt2008), which is consistent with studies on childhood adversity (Lin, Neylan, Epel, & O’Donovan, Reference Lin, Neylan, Epel and O’Donovan2016), including bullying (Arana et al., in press; Copeland et al., Reference Copeland, Wolke, Lereya, Shanahan, Worthman and Costello2014; Takizawa et al., Reference Takizawa, Danese, Maughan and Arseneault2015), being linked to higher levels of inflammation.
Genetic Factors
In addition to its relation to HPA dysregulation and inflammation, childhood trauma has been shown to be associated with epigenetic alterations such as DNA methylation (see Szyf & Bick, Reference Szyf and Bick2013 for review). DNA methylation is type of epigenetic mechanism that “maintains gene activity or changes gene expression by activating or silencing the gene, resulting in the development of phenotypes that are time-dependent and are not determined by the DNA sequence at that locus” (Vaillancourt et al., Reference Vaillancourt, Hymel and McDougall2013, p. 243–244). Simply stated, while experiences cannot alter the nucleotide acid sequence of DNA (i.e., do not alter the genetic code); they can alter the epigenome that tells the genome what to do. DNA methylation is thought to be the most stable type of epigenetic alteration and it is presumed to be a mechanism by which early-life adversity has long-term effects on the developing person (Tyrka et al., Reference Tyrka, Ridout and Parade2016).
Besides studies showing that early negative life experiences are associated with DNA methylation (Szyf & Bick, Reference Szyf and Bick2013), there is also one study showing that being bullied by peers was associated with epigenetic alterations. Specifically, Ouellet-Morin et al. (Reference Ouellet-Morin, Wong, Danese, Pariante, Papadopoulos, Mill and Arseneault2013) found that bullying victimization influenced DNA methylation of the serotonin transporter gene (5-HTT). Bullied twins had higher 5-HTT DNA methylation than nonbullied twins and increases in DNA methylation were associated with a blunted cortisol response to stress. These results suggest that biological modifications (i.e., DNA methylation) and HPA functioning may be functionally associated. According to Tyrka et al. (Reference Tyrka, Ridout and Parade2016) this is indeed the case. Childhood adversity is linked to altered methylation patterns, which are implicated in the stress-response system.
The 5-HTT gene has been connected in the development of many psychiatric conditions, especially in the context of environmental adversity (Uher & McGuffin, Reference Uher and McGuffin2008). In particular, a functional polymorphism (i.e., genetic variation) in the promoter region of the serotonin transporter gene (5-HTTLPR) has been linked to depression in the context of trauma. For example, Caspi et al. (Reference Caspi, Sugden, Moffitt, Taylor, Craig, Harrington and Poulton2003) found that individuals maltreated in childhood were far more likely to be depressed in adulthood if they had two copies of the short allele (SS) in the 5-HTTLPR and less likely to be depressed if they had at least one copy of the long allele (L). The short allele is considered a risk variant of the 5-HTT because it has been shown to have lower transcriptional efficiency of the promoter than the long allele (Caspi et al., Reference Caspi, Sugden, Moffitt, Taylor, Craig, Harrington and Poulton2003; Lesch et al., Reference Lesch, Bengel, Heilis, Sabol, Greenberg, Petri and Murphy1996).
Several researchers have replicated Caspi et al.’s (Reference Caspi, Sugden, Moffitt, Taylor, Craig, Harrington and Poulton2003) finding with regard to bullied youth. Sugden et al. (Reference Sugden, Arseneault, Harrington, Moffitt, Williams and Caspi2010) found that bullied children who carried the SS genotype were at greater risk for developing emotional problems than bullied children with the SL or LL genotype. Benjet, Thompson, and Gotlib (Reference Benjet, Thompson and Gotlib2010) reported that relationally bullied girls were far more likely to be elevated on symptoms of depression if they carried the SS genotype than if they were homozygous for the long allele or heterozygous for the short and long alleles. Other researchers have also shown that 5-HTTLPR moderates the relation between health problems and peer victimization (Banny, Cicchetti, Rogosch, Oshri, & Crick, Reference Banny, Cicchetti, Rogosch, Oshri and Crick2013; Iyer, Dougall, & Jensen-Campbell, Reference Iyer, Dougall and Jensen-Campbell2013). Although these results are intriguing, it is worthy to note that there is a lot of controversy associated with candidate gene by environment interactions because of their low replicability and their high rates of false discoveries (Duncan, Pollastri, & Smoller, Reference Duncan, Pollastri and Smoller2014). Still, the genetics research suggests at least two simple pathways to consider. Candidate gene by environment interaction studies support that biological risk can interact with trauma to produce mental health problems (i.e., biological risk + exposure to bullying ➔ poor mental health) and epigenetic studies support that exposure to trauma is associated with biological alterations, which, in turn, are associated with mental health problems (i.e., exposure to bullying ➔ biological changes ➔ poor mental health). What has not been considered to date is how these two pathways interact over time in the context of being bullied to confer a risk for poorer mental health outcomes.
Conclusion
The research reviewed in this chapter suggests that experience of being bullied is not inconsequential. Rather, being subjected to repeated, intentional humiliation and oppression by peers seems to become biologically embedded. Furthermore, the neurobiological changes associated with this trauma appear to presage psychobiological sequela that undermine victims’ ability to cope with subsequent psychosocial stressors and confer a risk for poorer outcomes, like the mental health difficulties linked to this prevalent problem.
Introduction
Aggression and violent behavior represent serious threats to public safety. In the United States alone, approximately 5.4 million people were the victims of violent crimes in 2014 (Truman & Langton, Reference Truman and Langton2015). To put this number in perspective, the most recent estimates from the National Crime Victimization Survey data suggest that approximately 20.1 out of every 1,000 Americans were victims of violent crime in 2014 (Truman & Langton, Reference Truman and Langton2015). In addition, the Federal Bureau of Investigation’s most recent Uniform Crime Report data indicate that more than 1,100,000 violent crimes were reported to the police in 2013 (US Department of Justice, Federal Bureau of Investigation, 2014). Consistently high rates of violent crime highlight the need for research examining the etiology of aggressive and violent behavior. In conjunction with this research, innovative policy changes are also needed to address the primary sources of violence in order to reduce the threat to public safety.
Criminological theory and research has traditionally examined environmental explanations for aggression and violent behavior. Mainstream criminological theories, for instance, generally locate the source of antisocial behavior in neighborhoods, peer associations, or parenting practices. Recently, however, there has been a shift in the field with increased interest on exploring the possibility that genetic factors may influence aggression and violent behavior.
The aim of this chapter is to provide an overview of the connection between molecular genetics and aggression and violent crime. In order to do so, we will first briefly discuss findings from behavioral genetics studies. Second, we will briefly discuss the structure of DNA and how genes influence variation in behavioral phenotypes. Third, we will present the current research on candidate genes studies and genome-wide association studies (GWAS) in relation to aggressive and violent behavior. Fourth, we will outline the extant research on gene–environment interactions. Finally, we will discuss the implications of molecular genetics studies for policies, interventions, and possible preventative treatments for aggression and violent behavior.
Behavioral Genetics
Behavioral genetic research typically involves estimating the influence of genetic and environmental influences on variation in human behavior. To do this, researchers utilize quantitative genetics methodologies that are capable of disentangling the proportion of phenotypic variance that is attributable to genetic and environmental factors. The environmental component of the variance is further divided into shared and nonshared environmental factors. Shared environments are environmental components that influence children in the same household to develop similarly to one another. In contrast, nonshared environments are environmental components that influence children who grow up in the same household to develop differently from one another. Common methodologies used by behavioral geneticists include twin studies and adoption studies (Plomin, DeFries, Knopik, & Neiderhiser, Reference Plomin, DeFries, Knopik and Neiderhiser2013).
In twin studies, researchers compare the behavior of twins living in the same household to obtain heritability estimates (Plomin et al., Reference Plomin, DeFries, Knopik and Neiderhiser2013). By incorporating twins with different levels of genetic relatedness, behavioral geneticists are able to calculate estimates of the influence of genetic and environmental factors on variance in human behavioral outcomes. For example, dizygotic (DZ) twins share approximately 50% of their genetic material, whereas monozygotic (MZ) twins share around 100% of their genetic material. As a result, researchers can use correlations between MZ and DZ twins to determine which portions of the variance of behavioral phenotypes are the result of genetic and environmental factors. According to these estimates, if the assumptions of twin-based methodologies are met – and mathematical simulations indicate that they are (Barnes et al., Reference Barnes, Wright, Boutwell, Schwartz, Connolly, Nedelec and Beaver2014) – MZ twins will be more phenotypically similar to each other than DZ twins if the traits in question are heritable.
Behavioral geneticists are also able to estimate the influence of genetic and environmental factors by examining adoptees. Adoption studies measure the influence of genetic and environmental factors on human behavior by taking into account the genes the adopted child inherited from their biological parents, as well as the environment provided by their adoptive parents (Beaver, Reference Beaver, Wright and Walsh2008). Phenotypic similarities between adoptees and their biological parents would suggest that the phenotype was more influenced by genetics than by the environment (Beaver, Reference Beaver2011). Likewise, if a child resembles their adoptive parents more so than their biological parents on several phenotypes, it would serve as evidence that those particular phenotypes were influenced more by the environment than by biology.
In general, findings from twin and adoption studies reveal that approximately 50% of the variance in behavioral phenotypes is attributable to genetic factors. This estimate has been supported by a recent meta-analysis conducted by Polderman and colleagues (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) that examined twin studies published in the last half-century. Polderman et al.’s (Reference Polderman, Benyamin, de Leeuw, Sullivan, van Bochoven, Visscher and Posthuma2015) analysis of more than 2,700 studies examining nearly 17,800 traits in more than 14,500 twin pairs, revealed that approximately 49% of the variance of human phenotypes is the result of genetic factors. Related studies have also revealed that approximately 50% of the variance in antisocial phenotypes, including aggression and violent behavior, is attributable to genetic factors (Ferguson, Reference Ferguson2010; Mason & Frick, Reference Mason and Frick1994; Miles & Carey, Reference Miles and Carey1997; Rhee & Waldman, Reference Rhee and Waldman2002). The remaining half of the variance of these phenotypes is attributed to environmental influences, with the majority of this variance being explained by environmental factors that are not shared between twins. As a whole, behavioral genetics studies suggest that aggressive and violent behavior are due, at least in part, to the influence of genetics.
Molecular Genetics
The methodologies and analytical approaches used in behavioral genetic research are valuable for identifying the proportion of variance that is due to genetic and environmental factors, but these methodologies do not provide information on the specific genes involved in creating phenotypic variance. Molecular genetics studies, however, can be used to identify specific genes that are implicated in the development of human phenotypes. Molecular genetic studies are able to pinpoint specific genes involved in phenotypic outcomes by testing for associations between possessing particular genetic markers and behavioral outcomes. Using molecular genetics methodologies, a number of genes have been linked with the development of antisocial phenotypes including aggressive and violent behavior (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002; Guo, Roettger, & Shih, Reference Guo, Roettger and Shih2007). Before discussing specific genes that are involved in the development of aggression and violent behavior, we will briefly outline the structure of DNA and explain how specific genes can influence variation in phenotypes.
Genes are housed on deoxyribonucleic acid (DNA), which resides in the nucleus of all cells (expect for red blood cells). The genetic code is written in nucleotide bases, which, along with a sugar phosphate backbone, make up the structure of the DNA molecule. There are four nucleotide bases in DNA – Adenine (A), Thymine (T), Guanine (G), and Cytosine (C). Each of these bases is able to bond with one, and only one, of the other three bases. To illustrate, A bonds with T and G bonds with C. As a result of these pair-specific bonds between nucleotide bases, the DNA strands in the double helix are complementary to one another.
Genes are composed of contiguous nucleotide base pairs that work together and contain the information required for the production and structure of proteins. Proteins are complex organic molecules that are necessary for the development, structure, and function of organisms. For example, proteins compose the structure of cells, transport nutrients throughout the body, and are responsible the transportation of information between neurons. Proteins are composed of amino acids that are linked together in a particular order. The specific configuration of amino acids in proteins is specified by sequences of nucleotide bases present in genes. In order to create proteins, the DNA code is first transcribed into ribonucleic acid (RNA) and then translated into the corresponding sequence of amino acids. The nucleotide bases in the subsequent RNA are complementary to the DNA sequence, however, in RNA molecules “U” is substituted in place of “T.” Importantly, the reading frame for translating the genetic code to amino acids is three nucleotide bases – referred to as codons. Each codon corresponds to an individual amino acid and, therefore, the order of nucleotide bases and codons in a gene is responsible for the sequence of amino acids that are joined together to form a protein. If there are errors in the order of nucleotide bases in a gene, then this can lead to changes in the structure of the protein that can result in compromised protein functionality.
Most of the genes in the human genome have only one version. However, a small subset of genes, referred to as polymorphisms, have more than one version present in the population (Mielke, Konigsberg, & Relethford, Reference Mielke, Konigsberg and Relethford2006). The different versions of genes available within the population are called alleles. The presence of different alleles within a population accounts for the variation in physical and behavioral phenotypes observed between people. For instance, hair texture may be determined by a polymorphic gene that has one allele that codes for curly hair and another allele that codes for straight hair.
Most of the polymorphisms present in the population do not appear to cause visible or functional differences in phenotypes because the differences between the available alleles do not lead to changes in protein structure. That is, many polymorphisms do not lead to functional differences because the different alleles still code for the same protein. There are, however, some polymorphisms in the population that correspond to functional differences that lead to the production of different phenotypes. Polymorphisms can lead to the development of different phenotypes when the differences between the available alleles leads to different amino acids being substituted into proteins that may then affect the protein’s structure and function. For instance, a polymorphism that codes for a neurotransmitter may have two alleles where version A functions efficiently, but version B may code for a different amino acid leading the subsequent neurotransmitter to be less efficient. As a result, individuals who have version B of the gene may exhibit a different behavioral phenotype than individuals with version A (e.g., temper, impulsivity).
In general, there are two main kinds of polymorphisms that can lead to differences in phenotypes. The first type of polymorphism involves the substitution of single nucleotides within genes, while the second type of polymorphism involves sections of repeating sequences of nucleotides – referred to as variable-number tandem repeats (VNTRs). VNTRs are further broken down into microsatellite and minisatellite polymorphisms based on the number of nucleotides involved in the repeated sections.
The first type of polymorphism, single-nucleotide polymorphisms (SNPs), are the result of the substitution of one nucleotide in a gene. Changing a single nucleotide within a codon can lead to the substitution of a different amino acid in the resulting protein, leading to structural and functional differences. To illustrate, a transcribed gene may have one copy that reads AUACUUCAUUAG, and another copy that reads AUACUUCAGUAG. In this case, substituting a G in place of the U in the second allele leads to the substitution of a glutamine amino acid in place of histidine. The substitution of a single amino acid within a protein can lead to changes in the structure and function of the protein that can lead to phenotypic changes. Notably, SNPs are the most common type of polymorphisms and are believed to be responsible for the majority of human genetic diseases (Plomin, DeFries, Craig, & McGuffin, Reference Plomin, DeFries, Craig and McGuffin2001).
VNTRs are polymorphisms that are characterized by repeating sections of nucleotides within genes. There are two main types of VNTRs, microsatellites and minisatellites, which differ according to the number of nucleotides involved in the repeating sections. In both of these types of VNTRs, the differences between alleles are due to differences in the length of the gene. That is, some alleles, with more repeated sections, are longer than others. The first type of VNTRS, microsatellites, are polymorphisms where a small set of nucleotide bases (usually less than six bases) repeat for a variable number of times. Minisatellites, on the other hand, are VNTR polymorphisms that have a larger number of repeated bases (usually greater than ten) than in microsatellites. VNTRs have been implicated in a number of human genetic diseases. For example, Huntington’s disease has been found to be the result of a microsatellite polymorphism where a three-nucleotide segment (CAG) of the huntingtin gene can repeat a variable number of times. Alleles that have between 6 and 35 CAG repeats usually result in properly functioning huntingtin proteins, whereas huntingtin genes that have 36 or more CAG repeats tend to result in a protein mutation that leads to the development of Huntington’s disease (Budworth & McMurray, Reference Budworth and McMurray2013).
The types of polymorphisms we have discussed can influence phenotypes in several ways. The most straightforward way polymorphisms can affect phenotypes is through monogenic effects. Monogenic effects refer to phenotypes that are the result of a single gene. In monogenic effects, only one gene is responsible for the development of a phenotype, and the presence or absence of an allele can be used to predict whether an individual will exhibit a specific phenotype. Several human diseases are the result of monogenic effects, including polycystic kidney disease and Tay-Sachs disease.
A second way that polymorphisms can influence phenotypes is through polygenic effects. Polygenic effects involve a process through which multiple genes work together to influence an individual’s phenotype. In polygenic effects, genes work together probabilistically. In these cases, the effect of each individual gene is small, but the genes function together to increase the likelihood of developing certain phenotypes. In other words, individuals who carry more alleles that predispose them to developing a certain phenotype will be more likely to the exhibit the phenotype in question than individuals who carry fewer of these specific alleles. Human behavioral outcomes such as aggression and violent behavior are likely the result of polygenic effects where multiple genes work together to increase the risk of developing aggressive behavioral outcomes.
A final way polymorphisms can influence phenotypes is through pleiotropic effects. Pleotropic effects occur when a single gene influences multiple phenotypes. For example, cystic fibrosis is a genetic disease that arises from a single gene mutation that affects the function of multiple organs including the lungs, pancreas, liver, and kidneys. In relation to aggression and violent behavior, a number of genes associated with neurotransmission have been shown to influence multiple behavioral outcomes associated with antisocial behavior (Beaver, DeLisi, Wright, & Vaughn, Reference Beaver, DeLisi, Wright and Vaughn2009; Beaver, Wright, & Walsh, Reference Beaver, Wright and Walsh2008; Gill, Daly, Heron, Hawi, & Fitzgerald, Reference Gill, Daly, Heron, Hawi and Fitzgerald1997; Rowe et al., Reference Rowe, Stever, Gard, Cleveland, Sanders, Abramowitz and Waldman1998).
Candidate Genes for Aggression and Violent Behavior
Molecular genetic studies are designed to pinpoint genetic polymorphisms that are involved in the development of phenotypic variance, including variation in aggressive and violent phenotypes. In general, candidate gene studies are largely “theory-driven” and are aimed at identifying specific genes associated with certain phenotypes. The targets of candidate gene studies are generally selected in accordance with neurobiological theories of behavior. Accordingly, most of the genes implicated as candidate genes for aggressive and violent phenotypes are related to neurotransmission. To date, candidate gene studies have identified several genes involved in the development of ADHD, alcoholism, aggression, and violence (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002; Dick & Faroud, Reference Dick and Foroud2003; Guo et al., Reference Guo, Roettger and Shih2007; Li, Sham, Owen, & He, Reference Li, Sham, Owen and He2006; Rujescu, Giegling, Gietl, Hartman, & Moller, Reference Rujescu, Giegling, Gietl, Hartman and Moller2003). Many of the genes identified as contributing to these antisocial phenotypes, including aggression and violence, code for proteins involved in the transportation, reception, and breakdown of neurotransmitters.
Neurotransmitters are proteins that deliver chemical messages from one cell to another in the nervous system. In general, neurotransmitters deliver a message from one cell to another by first being released by the presynaptic neuron and then traversing the synapse (space between neurons) to receptors on the postsynaptic neuron. Once the neurotransmitters have delivered the message to the postsynaptic neuron they are removed from the synapse through one of two mechanisms. In the first mechanism, referred to as reuptake, neurotransmitters are removed from the synapse and delivered back to the presynaptic neuron by transporter proteins. In the second mechanism, neurotransmitters that still remain in the synapse are metabolized (chemically broken down) by enzymes. If the transporter proteins or the enzymes responsible for metabolizing neurotransmitters have structural abnormalities, they may not function efficiently, which may cause neurotransmitter levels to fluctuate, leading to disturbances in mood and behavior.
Candidate gene studies have focused on genes related to the transportation and reception of neurotransmitters along with genes that code for enzymes that breakdown neurotransmitters in the synapse. To date, researchers have identified genes related to three classes of neurotransmitters that appear to be associated with aggression and violent behavior. Specifically, researchers have identified genes involved in the transportation and reception of dopamine, serotonin, and GABA that are associated with antisocial outcomes. In addition, researchers have identified genes coding for two enzymes (MAOA, COMT) that are involved in the breakdown of neurotransmitters that are linked with aggression and violent behavior. In the following section, we discuss the extant research pertaining to genes related to the transportation of neurotransmitters and enzymes that have been associated with aggression and violent behavior.
Genes Related to the Transportation, Reception, and Breakdown of Neurotransmitters
There are three types of neurotransmitters that have been linked with aggression and violent behavior. First, we will discuss the role of dopamine levels and dopaminergic genes in aggression and violent behavior. Second, we will discuss the role of serotonin levels and serotonergic polymorphisms in antisocial outcomes. Finally, we will discuss GABA levels and GABAergic genes associated with aggression and violent behavior.
Dopamine is an excitatory neurotransmitter that is involved in the reward system of the brain. There is a considerable amount of variation in baseline dopamine levels between different people and dopamine levels can change in response to environmental factors. High levels of dopamine have been shown to be associated with increased involvement in impulsive and violent behavior (Niehoff, Reference Niehoff1999). However, there is also evidence linking low levels of dopamine to involvement in aggressive behavior (Raine, Reference Raine1993). As a result, the relationship between dopamine levels and aggressive behavior may be nonlinear such that high and low levels of dopamine are both associated with aggressive conduct. Molecular genetic studies have revealed that many of the genes involved in the transportation of dopamine are polymorphic, including DAT1, DRD2, DRD3, DRD4, and DRD5.
A number of polymorphisms that influence the transportation of dopamine have been associated with aggressive and violent behavior. For instance, DAT1, a dopamine transporter gene, is a polymorphism with a minisatellite that repeats up to 11 times, where the 10-repeat allele has been linked with violent delinquency and violent behavior in adulthood (Guo et al., Reference Guo, Roettger and Shih2007). Empirical studies have also revealed that several dopamine receptor genes are associated with antisocial outcomes. For example, DRD2, a dopamine receptor gene that codes for D2 dopamine receptors, is a polymorphism with a SNP that has two alleles present in the population – A1 and A2. Empirical research has demonstrated that the A1 allele is associated with involvement in violent delinquency (Guo et al., Reference Guo, Roettger and Shih2007). The association between the A1 allele and aggressive behavior may be explained by inefficient dopamine reception in carriers of the A1 allele. Not surprisingly, studies have revealed that individuals who carry the A1 allele tend to have fewer D2 dopamine receptors, which may result in poorer regulation of dopamine levels (Pohjalainen et al., Reference Pohjalainen, Rinne, Någren, Lehikoinen, Anttila, Syvalahti and Hietala1998).
Serotonin is an inhibitory neurotransmitter that is involved with the regulation of behavior and impulse control. A significant body of research demonstrates that low levels of serotonin are associated with aggressive and violent behavior. The first studies examining serotonin levels and aggression, used a cerebrospinal fluid (CSF) measures of the main metabolite of serotonin, 5-hydroxyindoleactic acid (5-HIAA), and found that serotonin levels accounted for more than 60% of the variation in aggression in subjects with personality disorders (Brown et al., Reference Brown, Goodwin, Ballenger, Goyer and Major1979). This finding led to a flurry of research on the association between serotonin levels and aggression. Subsequent studies examining this association have revealed mixed results. In general, however, previous studies have found evidence of an inverse association between serotonin levels and violent offending, childhood disruptive behavior, and arson (Kruesi et al., Reference Kruesi, Rapoport, Hamburger, Hibbs, Potter, Lenane and Brown1990; Limson et al., Reference Limson, Goldman, Roy, Lamparski, Ravitz, Adinoff and Linnoila1991; Virkkunen et al., Reference Virkkunen, Rawlings, Tokola, Poland, Guidotti, Nemeroff and Linoila1994). To date, three meta-analyses have been conducted on studies that have examined the relationship between serotonin and aggressive and violent behavior and they have revealed some evidence for a significant inverse relationship between serotonin and aggressive/violent behavior (Balaban, Alper, & Kasamon, Reference Balaban, Alper and Kasamon1996; Duke, Begue, Bell, & Eisenlohr-Moul, Reference Duke, Begue, Bell and Eisenlohr-Moul2013; Moore, Scarpa, & Raine, Reference Moore, Scarpa and Raine2002; Raine, Reference Raine1993). Molecular genetics studies have revealed that several of the genes involved in the transportation and breakdown of serotonin appear to be involved in the development of aggressive and violent phenotypes.
For instance, 5HTTLPR, a serotonin transporter gene, is polymorphic with a minisatellite that results in two different alleles. One of the alleles (S) is significantly shorter than the other allele (L). The S allele of 5HTTLPR has been identified as conferring increased risk for the development of antisocial phenotypes, including ADHD (Cadoret et al., Reference Cadoret, Langebehn, Caspers, Troughton, Yucuis, Sandhu and Philibert2003), heavy drinking (Herman, Smolen, & Hewitt, Reference Herman, Philbeck, Vasilopoulos and Depetrillo2003), and violent aggression (Haberstick, Smolen, & Hewitt, Reference Haberstick, Smolen and Hewitt2006; Liao, Hong, Shih, & Tsai, Reference Liao, Hong, Shih and Tsai2004; Retz, Retz-Junginger, Supprian, Thome, & Rösler, Reference Retz, Retz-Junginger, Supprian, Thome and Rösler2004). Moreover, at least one study has demonstrated that the S allele of 5HTTLPR is significantly more prevalent in violent offenders than in nonviolent offenders (Retz et al., Reference Retz, Retz-Junginger, Supprian, Thome and Rösler2004). There is some reason to believe that the association between 5HTTLPR and antisocial behavior can be explained by the S allele coding for transporter proteins with reduced serotonin reuptake abilities (Lesch et al., Reference Lesch, Bengel, Heils, Sabol, Greenberg, Petri and Murphy1996). Several serotonin receptor genes have also been linked with the development of aggressive and violent phenotypes. For instance, HTR1B, a gene that codes for the serotonin 1B receptor, is a polymorphism with several different alleles, where some alleles code for higher or lower HTR1B gene expression. The low-expression alleles have been linked with greater levels of anger, hostility, and aggression (Conner et al., Reference Conner, Jensen, Tennen, Furneaux, Kranzler and Covault2010; Jensen et al., Reference Jensen, Covailt, Conner, Tennen, Kranzler and Furneaux2009).
GABA is another inhibitory neurotransmitter that has been associated with aggression and antisocial behavior. In general, previous research has revealed that low levels of GABA are linked with aggressive behavior in humans and nonhumans (Bjork et al., Reference Bjork, Moeller, Kramer, Kram, Suris, Rush and Petty2001). These findings, along with findings from studies that indicate that GABA levels are under the influence of genetic factors (Petty et al., Reference Petty, Fulton, Kramer, Kram, Davis and Rush1999), suggest that genes related to regulating GABA levels may be associated with aggression and violent behavior. For instance, several studies have indicated that GABRA2, a GABA A receptor gene with several SNPs, is linked with increased risk of externalizing behavior, conduct disorder, and antisocial personality disorder (Dick et al., Reference Dick, Agrawal, Schuckit, Bierut, Hinrichs, Fox and Begleiter2006a; Dick et al., Reference Dick, Beirut, Hinrichs, Fox, Bucholz, Kramer and Foroud2006b; Dick et al., Reference Dick, Latendresse, Lansford, Budde, Goate, Dodge and Bates2009). In addition, there is some evidence that GABBR2, a GABA B receptor gene, is associated with antisocial behavior (Terranova et al., Reference Terranova, Tucci, Sartore, Cavarzeran, Pietra, Barzon and Ferrara2013).
Along with genes involved in the transportation and reception of neurotransmitters, genes that code for enzymes that breakdown neurotransmitters have also been associated with aggression and violent behavior. Specifically, genes related to two enzymes (MAOA, COMT) have been implicated in developing aggressive behavior. The MAOA gene codes for the production of the monoamine oxidase A (MAOA) enzyme, which breaks down monoamine neurotransmitters such as serotonin and dopamine. The MAOA gene contains a minisatellite that repeats between two and five times. The alleles for MAOA are generally considered to consist of two groups, one group of high-activity MAOA alleles (4R, 3.5R) and a group of low-activity MAOA alleles (2R, 3R). The low-activity MAOA alleles are less efficient at breaking down neurotransmitters and, as a result, carriers of the low-activity MAOA alleles may have fluctuations in their neurotransmitter levels. Low-activity MAOA alleles have been associated with involvement in aggressive and antisocial behavior in males (Ficks & Waldman, Reference Ficks and Waldman2014). The empirical research surrounding this association, however, is inconclusive, with some studies suggesting a positive association between the low-activity MAOA alleles and aggression, and some studies suggesting a negative association (Manuck, Flory, Ferrell, Mann, & Muldoon, Reference Manuck, Flory, Ferrell, Mann and Muldoon2000), or no direct association at all (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002; Verhoeven et al., Reference Verhoeven, Booij, Kruijt, Cerit, Antypa and Does2012). A recent meta-analysis examining the main effect of MAOA on behavioral outcomes revealed a significant positive relationship between the low–activity MAOA alleles and increased involvement in aggressive and antisocial behavior (Ficks & Waldman, Reference Ficks and Waldman2014). The inconsistent findings regarding the relationship between MAOA and antisocial behavior are likely explained by the effects of environmental factors moderating the influence of MAOA on behavioral outcomes. For instance, several studies have documented that the low-functioning alleles of MAOA interact with childhood maltreatment to predict antisocial behavior (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002) and conduct disorder (Foley et al., Reference Foley, Eaves, Wormley, Silberg, Maes, Kuhn and Riley2004).
COMT is a gene that codes for the production of the catechol-O-methyltransferase (COMT) enzyme, which breaks down catecholamines such as dopamine and epinephrine. The COMT gene contains a SNP with two alleles. One of the alleles codes for the amino acid Methionine (Met) while the other codes for the amino acid Valine (Val). Individuals who carry the Met allele have been shown to have reduced COMT activity, which is associated with increased levels of catecholamines. The Met allele has also been identified as conferring increased risk for the development of antisocial phenotypes. Specifically, the Met allele has been shown to be associated with involvement in aggressive and violent behavior (Hirata, Zai, Nowrouzi, Beitchman, & Kennedy, Reference Hirata, Zai, Nowrouzi, Beitchman and Kennedy2013; Jones et al., Reference Jones, Zammit, Norton, Hamshere, Jones, Milham and Owen2001; Volavka, Bilder, & Nolan, Reference Volavka, Bilder and Nolan2004).
Genome-Wide Association Studies (GWAS)
Genome-wide association studies (GWAS) test for associations between common genetic variants and variation in phenotypes. Unlike candidate gene studies, GWAS are data-driven and examine the entire genome of participants and test to see if there is an association between the frequency of common allelic variants and the development of certain phenotypes. As a result, GWAS are able to identify novel genes associated with particular phenotypes that might not be predicted by any existing theory of the etiology of particular phenotypes. GWAS generally focus on associations between SNPs and diseases or behavioral phenotypes. Previous GWAS that have investigated behavioral phenotypes have revealed several polymorphisms that are associated with conduct disorder (Dick et al., Reference Dick, Aliev, Krueger, Edwards, Agrawal, Lynskey and Bierut2011), anger-proneness (Mick et al., Reference Mick, McGough, Deutsch, Frazier, Kennedy and Goldberg2014), and hostility (Merjonen et al., Reference Merjonen, Keltikangas-Järvinen, Jokela, Seppälä, Lyytikäinen, Pulkki- Råback and Lehtimäki2011).
GWAS have only recently been applied to investigating aggression and violent behavior. One of the first GWAS to examine the etiology of antisocial behavior did not find any genetic polymorphisms that reached the level of genome-wide significance; however, the authors did identify one gene (DYRK1A) that appeared to be nearing significance (Tielbeek et al., Reference Tielbeek, Medland, Benyamin, Byrne, Heath, Madden and Verweij2012). A second GWAS by Salvatore and colleagues (Reference Salvatore, Edwards, McClintick, Bigdeli, Adkins, Aliev and Dick2015) was also not able to identify any polymorphisms that reached genome-wide significance; however, they were able to identify several SNPs that were nearing significance for adult antisocial behavior. A third study by Pappa and colleagues (Reference Pappa, St Pourcain, Benke, Cavadino, Hakulinen, Nivard and Tiemeier2015) was able to identify one gene on chromosome 2 that neared genome-wide significance for predicting childhood aggression. In addition, they were able to identify several other polymorphisms that may be related to childhood aggression (Pappa et al., Reference Pappa, St Pourcain, Benke, Cavadino, Hakulinen, Nivard and Tiemeier2015). While these studies suggest several genes that may be involved in the development of aggressive and violent behavior, the inability of these studies to detect genes that reach the level of statistical significance highlights several concerns with GWAS.
One major concern for GWAS is that genes associated with phenotypes may only explain a very small portion (< 1%) of the variance in the particular trait. As a result, large sample sizes are needed in order to detect these associations. Further exacerbating the problem, the analytic strategy employed in GWAS necessitates a very conservative estimate of statistical significance (p < 5.0 x 10–8), making it difficult to find polymorphisms that reach the level of genome-wide significance.
The findings of GWAS also raise concerns about what researchers have termed as “missing heritability.” Missing heritability refers to the gap between the heritability estimates of twin studies and heritability as estimated by associations identified in GWAS. Unfortunately, SNPs (as detected by GWAS) only appear to account for approximately 10% of phenotypic outcomes, and 5% or less of the variance in behavioral phenotypic outcomes (Plomin, Reference Plomin2013). Therefore, these findings contrast research from behavioral genetic studies that imply that approximately 50% of the variance of human phenotypes is attributable to genetic factors. One explanation for missing heritability is that GWAS are only able to identify the additive genetic effects of common SNPs, and, therefore, they miss heritability that is due to rare gene variants and genes that may be involved in gene–environment interactions (Manolio et al., Reference Manolio, Collins, Cox, Goldstein, Hindorff, Hunter and Visscher2009; Manuck & McCaffery, Reference Manuck and McCaffery2014). That is, while associated polymorphisms may only have a small effect on their own, polymorphisms may interact with specific environments to have larger effects on particular phenotypes. As a result, GWAS may miss associated genes that do not appear to significantly influence aggression on their own, but may be involved in gene–environment interactions for the prediction of aggression and violent behavior.
Gene–Environment Interactions
The previous sections have discussed individual genes that have been identified as influencing the development of aggressive and violent behavior, however, genetic factors do not work independently to influence the formation of antisocial phenotypes. Frequently, instead, genetic factors work interactively with the environment to produce behavioral phenotypes. To illustrate, alleles that confer increased risk for developing antisocial behavior may have a more pronounced influence on the development of aggressive and violent behavior when paired with adverse environments compared to more favorable environments. In this way, the influence of an allele may be dependent on the environment. This dependent relationship between genotype and environmental factors is referred to as a gene–environment interaction.
Gene–environment interactions capture the complex relationships between genotypes and environmental factors. The logic of gene–environment interactions is based on individuals each having unique genotypes that confer different genetic predispositions and different susceptibilities to the influence of environmental factors. As a result of these genetic differences, some individuals may be more affected by certain environmental factors than others. Therefore, gene–environment interactions are able to explain why different people may react to the same environment in different ways. To illustrate, individuals with a genetic propensity toward aggressive behavior may be more susceptible to respond to environmental factors, such as maltreatment, with aggressive behavior than individuals without the genetic predisposition toward aggression. In this way, behavioral phenotypes are the product of an interaction of both genetic propensities and environmental factors.
For example, one of the earliest studies to explore gene–environment interactions revealed that carriers of low-functioning MAOA alleles were more likely to develop antisocial behavior in response to childhood maltreatment than carriers of the high-functioning alleles (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002). To illustrate, only 12% of the study’s participants were both carriers of the low-functioning MAOA alleles and exposed to maltreatment, yet they comprised 44% of the serious convictions of all the participants in the study. Importantly, individuals who were exposed to only one of the risk factors (genetic or environmental) were significantly less likely to develop antisocial phenotypes than individuals exposed to both risk factors. The gene–environment interaction between low-functioning MAOA alleles and childhood maltreatment has also been demonstrated to predict conduct disorder (Foley et al., Reference Foley, Eaves, Wormley, Silberg, Maes, Kuhn and Riley2004) and mental health problems (Kim-Cohen et al., Reference Kim-Cohen, Caspi, Taylor, Williams, Newcombe, Craig and Moffitt2006). In addition, a recent meta-analysis by Byrd and Manuck (Reference Byrd and Manuck2014) revealed consistent support for the interaction between the low-functioning MAOA alleles and childhood maltreatment for the prediction of developing an antisocial phenotype.
Several other gene–environment interactions have also been implicated in the development of aggressive and violent phenotypes. For instance, several genes related to dopamine function have been documented to interact with environmental risk factors. For example, empirical studies have indicated that DRD2 interacts with having a criminal father to influence involvement in violent delinquency (DeLisi, Beaver, Vaughn, & Wright, Reference DeLisi, Beaver, Vaughn and Wright2009) and DAT1 interacts with delinquent peers to predict violent behavior (Vaughn, DeLisi, Beaver, & Wright, Reference Vaughn, DeLisi, Beaver and Wright2009). DRD2 and DRD4 have also been shown to interact with neighborhood characteristics to predict involvement in violent delinquency (Beaver, Gibson, DeLisi, Vaughn, & Wright, Reference Beaver, Gibson, DeLisi, Vaughn and Wright2012). Similarly, dopamine risk, as conferred by possessing the risk alleles of DRD2, DRD4, and DAT1, has been shown to interact with neighborhood characteristics to predict increased involvement in violent behavior (Barnes & Jacobs, Reference Barnes and Jacobs2013). Furthermore, there is some evidence that the serotonin transporter gene 5HTTLPR interacts with socioeconomic status to predict psychopathic traits (Sadeh et al., Reference Sadeh, Javdani, Jackson, Reynolds, Potenza, Gelernter and Verona2010) and childhood maltreatment to predict the development of antisocial personality disorder (Douglas et al., Reference Douglas, Chan, Gelernter, Arias, Anton, Poling and Kranzler2011).
Further complicating the relationship between genes and the environment, gene–environment interactions appear to have a number of different forms. The most common form of gene–environment interaction is where risk alleles interact with environmental risk factors to produce antisocial behavior. In this case, the environmental risk factors can be seen as “triggering” the effects of an individual’s genetic propensity towards antisocial behavior. This form of gene–environment interaction is referred to as the “diathesis-stress” model. The diathesis-stress model of gene–environment interactions has received the most empirical attention and support (Manuck & McCaffery, Reference Manuck and McCaffery2014). For example, the diathesis stress model is indicated in the MAOA-childhood maltreatment interaction mentioned above.
While gene–environment interaction research frequently indicates that risk alleles will have their greatest effects on behavioral outcomes when paired with adverse environments, this is not always the case. For instance, DRD2 has been shown to interact with delinquent peers to predict victimization for individuals who appear to have a low level of environmental risk. To illustrate, in a study by Beaver and colleagues (Reference Beaver, Wright, DeLisi, Daigle, Swatt and Gibson2007), the A1 risk allele appeared to have a more pronounced effect on victimization for males with a low number of delinquent peers compared to males with a high number of delinquent peers. This kind relationship can be explained by the “social push hypothesis,” where individuals will be most susceptible to their genetic predispositions in good environments, whereas, in adverse environments, environmental factors may have more of an influence over the development of behavioral phenotypes that genetic predispositions.
A third model for gene–environment interactions, referred to as the “differential susceptibility model,” has been posed by Belsky and colleagues (Belsky & Pluess, Reference Belsky and Pluess2009). According to this model, alleles should be considered to be “plasticity alleles.” Plasticity alleles determine how malleable an individual is to environmental influences. In this case, someone who has a high number of plasticity alleles would be more influenced by environmental factors for the development of behavioral phenotypes. For instance, an individual with a high degree of plasticity would be more likely to develop aggressive and violent behavior in a criminogenic environment. Conversely, this same individual would also be more likely to be affected by the influences of a prosocial environment. Empirical studies that have examined the differential-susceptibility hypothesis have revealed some support for this model (Manuck & McCaffery, Reference Manuck and McCaffery2014). For example, the S allele of 5HTTLPR appears to operate as a plasticity allele that interacts with parenting practices to predict the presentation of positive affect or depressive symptoms, when compared to behavioral outcomes of individuals with the L allele (Taylor et al., Reference Taylor, Way, Welch, Hilmert, Lehman and Eisenberger2006).
Policy Implications, Prevention Efforts, and Interventions
New advancements in the study of the genetic contributors to crime open opportunities for new policies that can attempt to prevent and intervene in the development of aggression and violent behavior. Previous research demonstrates that individuals’ responses to treatment, intervention, and preventive programs are affected by individual characteristics. For instance, males and high-rate offenders may be more responsive to particular treatment programs than females and low-rate offenders. The new knowledge obtained through molecular genetic research may be able to be used to tailor preventative and intervention programs to individuals’ genotypes. For example, Gajos, Fagan, and Beaver (Reference Gajos, Fagan and Beaver2016) recently outlined how genetic research can be used to inform policies pertaining to prevention programs. Specifically, Gajos and colleagues suggest that gene–environment interactions (especially as related to the differential susceptibility model) are a key area of research for genetically informed preventative research and policies.
Similarly, our overview of the existing molecular genetic research suggests that gene–environment interactions may be the most promising area of research for exploring both preventative and intervening practices. Of particular importance, gene–environment interactions can be used to explain why two different individuals respond to the same environment in different ways. As a result, findings from this area of research may be able to distinguish between certain genotypes that may make individuals more or less amenable to preventative practices and intervention policies. Therefore, findings from this area of research may be able to be used to design targeted treatment strategies for individuals who have different genotypes.
Recent studies have attempted to examine whether genotype can be incorporated into creating targeted intervention strategies. For instance, a study examining the influence of parental training on parental discipline practices and childhood externalizing behaviors found that changes in parental discipline practices were more effective in reducing childhood externalizing behavior in children who possessed the seven-repeat allele of DRD4 (Bakermans-Kranenburg, van Ijzendoorn, Pijlman, Mesman, & Juffer, Reference Bakermans-Kranenburg, Van Ijzendoorn, Pijlman, Mesman and Juffer2008). In addition, the ten-repeat allele of DAT1 has been associated with increased responsivity to family-based intervention strategies designed to decrease the presentation of ADHD symptoms and behavioral problems (van den Hoofdakker et al., Reference Van den Hoofdakker, Nauta, Dijck-Brouwer, van der Veen-Mulders, Sytema and Emmelkamp2012). While these studies do not directly address aggression and violent behavior it is reasonable to suggest that findings implicating gene-moderated responses to intervention programs may be generalizable to other behavioral phenotypes.
Findings from molecular genetics studies can also be used to inform preventative strategies by helping to identify which individuals may be the most at risk for developing aggressive or violent behavior in response to certain environmental stresses. For instance, knowledge of an individual’s genotype can be used to identify if the individual is at “high risk” for developing aggressive or violent behavior in response to adverse environments. As a result, this knowledge could be used to target preventative programs to those individuals deemed the highest risk for developing aggression. Two studies that have been conducted by Brody and colleagues using this line of thinking have found that genetically informed preventative programs can be used to decrease involvement in risky behavior in teenagers. In the first study, Brody, Beach, Philibert, Chen, and Murry (Reference Brody, Beach, Philibert, Chen and Murry2009) found that parenting interventions affected the association between 5HTTLPR genotype and engagement in risky behavior in adolescents. In the second study, Brody and colleagues (Reference Brody, Chen, Beach, Kogan, Yu, DiClemente and Philibert2014) found that parenting interventions moderated the relationship between DRD4 genotype and substance use in male adolescents. While these findings are promising, to date, very little research has attempted to alter environmental factors in such a way to ameliorate individual’s genetic risk of developing aggressive or violent behavior. Therefore, preventative strategies based on the logic of gene–environment interactions for preventing aggression and violence are currently situated in the theoretical realm. Further research will have to be conducted in order to determine if the moderating effects of prevention efforts on genetic risk for substance use and risky behavior carry over to aggressive and violent behavior.
Conclusion
Molecular genetics research is rapidly increasing knowledge pertaining to the development of aggression and violent behavior. Over the past two-and-a-half decades molecular genetics research has identified several candidate genes for aggression and violent behavior in humans. In addition, research exploring gene–environment interactions moves the field closer to being able to understand why some individuals may develop aggressive and/or violent behavior in response to certain environments while others do not develop these behaviors. More recently, GWAS have been employed to attempt to identify genes involved in the etiology of violence and aggression; however, this line of research is only in its infancy. Together these new avenues of research are useful in both the exploration of causes of aggressive and violent behavior and introducing possible new directions for preventative and intervention programs.
Introduction
Throughout history, the intersection between the ingestion of psychoactive intoxicants and aberrant and violent behavior has garnered significant and inglorious attention. Nations have struggled with how best to effectively and humanely handle drug experimentation and abuse and its links to violence. This may be particularly difficult with respect to young people, where, at least in Western nations, experimentation with drug use begins. Our approach to these vexing issues is to enact policies based upon scientific findings. We realize there is often a substantial lag time between the accumulation of research and its dissemination and absorption by the general population. There are signs, however, that this does indeed eventually occur.
We consider the co-occurrence of drug abuse and violence in adolescence to be a generalized neuro-dysregulation syndrome of altered behavior that is genetically and developmentally sensitive and socially and culturally contingent over time. With respect to addiction, we focus more on the concept of drug abuse because children and adolescents are less likely to experience the full bloom of the addiction career and also to demarcate from behavioral addictions such as gambling, compulsive sex, internet use, or exercise dependence.
Here, we provide readers a grasp of the inherent multilevel dynamism of drug abuse and violent delinquency across the biosocial spectrum. Specifically, we address the genetic and neurobiological landscape linking the research currents to those found in the corpus of literature on the life-course. We then draw connections to policy, prevention, and treatment implications of our approach.
The Genetic Architecture of Drug Abuse and Addiction
It appears that some youth are more prone to drug abuse than other youth. We direct attention now to examining the genetic architecture of the addiction-crime phenomenon. Interestingly, violence researchers are only beginning to recognize the value of genetics as related to aggression, but addiction scientists have been busy studying the genetic underpinnings of substance-use disorders for decades. Both violence and addiction researchers largely accept that drug abuse and offending runs in families; however, they often disagree about whether biology or environment is the driver. We, like many others, recognize that both biological factors and environmental drivers are significant causes of violence and addiction.
Much like violence, addiction is not a single gene or Mendelian disorder. There is no single addiction gene. Addictive behaviors are polygenic (i.e., caused by many genes). Addiction, like antisocial behavior, is a complex multifactorial behavioral phenotype. The level of complexity researchers are confronting is aptly stated by Volkow and Muenke (Reference Volkow and Muenke2012, p. 773):
In the case of substance use disorders, the powerful modulatory role played by complex environmental factors on brain processes which further muddle the picture, is particularly relevant. This is because, in the absence of drug exposure, itself an environmental factor, the specific addiction phenotype would remain hidden, even in the presence of an overwhelming genetic load. On the other hand, brain development and architecture, which are partly determined by genetic factors, can be affected by exposure to drugs. These two way interactions highlight the importance of genes involved in human brain development and function in the subsequent emergence of personality styles and emotional behavior reactivities.
Despite the challenges of uncovering the genetic architecture of addiction liability, there has been an increase in knowledge on this topic.
Similar to many, if not most, traits and behavioral phenotypes, investigations into the heritability of addiction using twin samples indicate that approximately 50% of the variance is due to genetic factors (Demers, Bogdan, & Agrawal, Reference Demers, Bogdan and Agrawal2014). One might wonder whether the substance of abuse matters with regard to these heritability estimates. The answer is not really. Regardless of whether the substance is alcohol, nicotine, marijuana, or cocaine, the results are relatively invariant. Results do change, however, depending on the developmental period at hand. Studies on the developmental sensitivity of heritability estimates suggest that genetic factors actually increase from early adolescence to adulthood (Kendler, Schmitt, Aggen, & Prescott, Reference Kendler, Schmitt, Aggen and Prescott2008). While heritability studies do indeed possess utility when beginning to study a phenomenon, it is now more important to study the many genes that place children and adolescents vulnerable to drug abuse and dependence at risk, and to study these genes in conjunction with modifiable environment risk factors.
Although there is no specific gene that is responsible for addiction vulnerability, the assumption underlying the investigation into specific genes is that there are important genes that underlie key neural systems in the brain that are in turn linked to both addiction and violence-proneness. As such, they serve as biomarkers. Many of these genes are found in key neurotransmitter systems such as the dopaminergic, serotonergic, or many other such systems. It is important to point out that these genes are polymorphic, meaning that they occur in more than one form. A number of genes have been implicated in drug abuse and addiction. Importantly, many of these genes overlap with externalizing and antisocial behavior as well, suggesting their role in both violence and drug abuse liability. GABRA2 is one such gene. This particular gene, expressed more strongly in men, codes for proteins in a major inhibitory neurotransmitter region, and is associated with alcohol dependence (Edenberg et al., Reference Edenberg, Dick, Xuei, Tian, Almasy, Bauer and Begleiter2004) and general externalizing behavior (Dick et al., Reference Dick, Latendresse, Lansford, Budde, Goate, Dodge and Bates2009). Low-activity alleles of the monoamine oxidase A (MAOA) have been found to be associated with a wide swath of antisocial behaviors, including addiction (Guo, Wilhelmsen, & Hamilton, Reference Guo, Wilhelmsen and Hamilton2007), and occur at a significantly higher level among persons who suffer from alcohol dependency or antisocial personality (Samochowiec et al., Reference Samochowiec, Lesch, Rottman, Smolka, Syagailo, Okladnova and Sander1999). Using the Add Health data, Beaver and colleagues (Reference Beaver, DeLisi, Vaughn and Barnes2010) found that adolescents possessing the low-activity MAOA alleles were more likely to join a gang and, while in the gang, more likely to fight and use weapons. Other genes involved in neurotransmission (chemicals that transmit signals that allow for communication between neurons in the brain) include those in the dopaminergic and serotonergic systems.
Dopamine in normal amounts is released when we eat, drink, and engage in sex. In contrast, super-normal amounts of dopamine are released with the use of amphetamines, cocaine, opiates, nicotine, alcohol, and cannabis – greatly stimulating the reward pathway. Dopamine receptor genes (i.e., DRD2) have been associated with heightened risk for addiction (Noble, Reference Noble2000). Another dopamine gene, DRD4, has been explored due to its linkage with this system and its association with novelty seeking. Research on serotonin, an important regulator of mood, has found that decreases in serotonin are associated with alcohol abuse and aggression (Nelson & Chiavegatto, Reference Nelson and Chiavegatto2001). One genetic polymorphism in the serotonergic region that has garnered heightened interest is the low-activity short allele (5-HTTLPR). A bevy of studies has produced interaction effects with this gene, suggesting it may be particularly sensitive to environmental stressors such as childhood maltreatment or witnessing or experiencing other traumatic events.
Researchers have been keen to study how genes interact with environments because these types of studies illuminate ways in which a modifiable environmental factor moderates genetic risk. Hicks and colleagues (Reference Hicks, South, DiRago, Iacono and McGue2009) found that several environmental risk factors (i.e., academic achievement, peer affiliations, familial relationship problems and other stressful life events) each interacted with genetic vulnerability to produce combined antisocial behavior and substance use. However, the literature on gene–environment interaction studies is notably inconsistent and findings cannot always be replicated in different samples. To illustrate the magnitude of this problem, Weeland and colleagues (Reference Weeland, Overbeek, Orobio de Castro and Matthys2015) conducted a systematic review of 53 published gene–environment interaction studies of externalizing behavior that focused on major candidate genes such as MAOA, DRD2, DRD4, DAT1, 5-HTTLR, and COMT and identified mixed results across samples. The environmental factor assessed in most of these interaction studies was some form of family conflict or adversity. So much about the intertwined nature of genes and environment is unknown. There are likely numerous environmental factors that are understudied and gene–environment interactions can occur throughout the life-course even in gestation and infancy.
Let’s now turn to what is perhaps the most interesting and important concept involving genes and environment: the concept of gene–environment correlation. Traditional thinking on drug abuse and violence among juveniles tends to regard the environment as actively bathed over us as individuals who are passive recipients. From the standpoint of gene–environment correlation, however, things are quite different; indeed, it is posited that an individual’s genetic makeup largely activates or influences the surrounding environment. The three types of gene–environment correlation interactions include active, evocative, and passive typologies. All three are likely important with regard to the etiology of drug abuse and violence. Active gene–environment correlation interactions occur when an individual seeks out environmental niches that reinforce their genetic proclivities. So, adolescents seeking out risky environments or friends who are more likely to experiment with substances is one avenue by which active gene–environment correlation works. In the evocative form of gene–environment correlation, the genetic-based attributes set in motion environmental responses. For instance, challenging adolescents who possess difficult temperaments or personalities provoke behavioral responses from others (harsh parenting or rejection) in the environment that might lead to further rebellious or self-medicating behavior. Passive gene–environment correlations occur when the environment is selected for by parents who share the same genes with the child. If the parent has had a history of drug abuse, they may expose the child to social environments that facilitate experimentation or regular use. As you might conclude, genes alone are just one part of the hypothetical recipe. Now, we turn to upstream neurobiological factors and take a look at what is going on “under the hood,” so to speak.
Neurobiology: What’s Going on Under the Hood?
It seems at times that we are awash in neuroscientific findings. Fields outside of neuroscience are linking their theories to what is being learned about the brain. While some may lament these developments as neurocentrism, a more positive spin on this trend is that the brain, as the seat of thought and behavior, is so fundamental to so many areas that it is becoming hard to avoid and this knowledge can be harnessed for the greater good. A growing number of violence researchers have recognized these developments and begun to incorporate neuroscience theories and research into their work. The problem of addiction and violence needs robust research from neuroscience to shed new light not only on the causal origins of problem behavior but also to inform prevention and policy.
The human brain is not only comparatively larger than that of other mammals but it also features substantial frontal regions (neocortex) that facilitate executive functioning and higher-level cognitive tasks. For adolescents, the basic functions of reward, motivation, emotion, and behavior inhibition are still developing, but substantial variation can also be observed. For example, some adolescents are more prone to anger and negative emotionality than others. These executive functions are important for a wide swath of human behavior including, of course, addiction and antisocial behavior (Berkman, Falk, & Lieberman, Reference Berkman, Falk and Lieberman2011). An abundance of research has shown that frontal regions are impaired in antisocial and risky behavior phenotypes (Brower & Price, Reference Brower and Price2001). Because there are an enormous amount of new connections being rapidly formed early in the life-course (and even through adulthood), plasticity to environmental stimuli is the rule rather than the exception. Several well-designed studies have shown that early problems (as early as age 3) in executive function are tied to a host of problem behaviors later in life, including addiction and offending (Fergusson, Horwood, & Ridder, Reference Fergusson, Horwood and Ridder2007; Moffitt et al., Reference Moffitt, Arseneault, Belsky, Dickson, Hancox, Harrington and Caspi2011; Tarter, Kirisci, Habeych, Reynolds, & Vanyukov, Reference Tarter, Kirisci, Habeych, Reynolds and Vanyukov2003). However, there is ample room for change and these links are not inevitable but probabilistic.
It is generally well accepted in addiction science that the key to understanding addictive processes lies in what is known as the brain’s reward pathway, or, more technically, the mesolimbic reward pathway. The mesolimbic reward pathway is made up of the ventral tegmental area (VTA), the amygdala, and the nucleus accumbens (NAc). The VTA is the front door of the reward pathway and is located beneath a substantial number of the brain’s opiate receptors. When stimulated, the VTA releases a flood of dopamine that streams to the amygdala. Being the command center of the fight/flight response that is crucial for detecting and evaluating threat, the amygdala passes information on to executive centers of the brain for processing. The NAc is the final destination for dopamine levels and levels of this pleasurable chemical are recorded, thereby representing a drug-using memory. The reward pathway, also known as the mesolimbic reward system, is essential for survival. This system evolved to provide the positive reinforcement for eating, drinking, sex, and other life-preserving functions. As a consequence, feel-good chemicals in the form of dopamine are released when we engage in eating, drinking, and sex. When we ingest psychoactive drugs, supernormal amounts of dopamine are released, resulting in what addiction researchers have commonly termed “hijacking” of this key neural circuit. For those who are especially vulnerable or susceptible, compulsive drug-seeking follows, representing one pathway to crime.
This drive for repeated reinforcement of rewarding behavior is moderated by one’s ability to self-regulate. Childress (Reference Childress, Miller and Carroll2006) has suggested that this “stop” (behavior inhibition) and “go” (drive) analogy provides a simple way for understanding the interchange between the reward pathway and executive governance. Although much more complex, adolescents who are high on “go” and low on “stop” are obviously the most at risk. Some drugs of abuse such as heroin or amphetamines may overwhelm the ability to exercise self-control. This is complicated by the phenomenon known as tolerance, where stronger or larger doses of the drug are needed to achieve the desired effect. Even more deleterious, larger amounts of the drug are sometimes needed to just feel normal. This is due to the NAc being hyper-stimulated. Moreover, developmental periods such as adolescence, where episodic use of drugs or minor engagement in delinquent acts is often considered normative (especially for males) (Moffitt, Reference Moffitt1993), are also time periods when executive functions are not fully developed. Steinberg (Reference Steinberg2007) regards this basic relationship between reward and behavior inhibition a central feature of adolescent risk.
So what are the avenues by which addiction and violence among adolescents are closely linked? One influential framework for approaching this question is Goldstein’s (Reference Goldstein1985) tripartite explanation of the drugs-crime nexus in which he articulated three forms of drug-related violence: systemic, psychopharmacological, and economic-compulsive. However, it is clear that this conceptualization is biosocial in nature. Although one could readily subscribe to the view that systemic violence is largely social, that view would only be partially correct as there is likely a mix of genes, brain, and environmental reasons as to why those in the drug trade are more violent than others.
Outside of violence stemming from turf disputes and interpersonal conflict associated with the drug trade, there are two major pathways to violence for adolescents that are directly linked to neural substrates of addiction. One of these pathways involves violence that is a direct cause of the drug use; the second is violence as a consequence of compulsive drug-seeking. In the former case, the causal effect is difficult to measure as it can be confounded by the individuals’ previous tendency toward using violence. For example, a meta-analysis of 13 imaging studies involving youth (age 12 to 17) with conduct problems found gray matter deficits in frontal regions and emotional processing areas prior to being classified as drug-dependent (Rogers & De Brito, Reference Rogers and De Brito2016). However, one could see this as an enhancement effect where drugs amplify the pre-existing tendency toward violence or via the drug’s ability to lower neural inhibitions and thereby removing any barriers to the use of violence. We know some drugs of abuse are better at this than others. For example, alcohol is likely to have a greater enhancement effect than cannabis. Compulsive drug-seeking occurs when adolescents steal or rob others to obtain money to buy drugs; this behavior is closely coupled with drug-selling. Although we often think of systemic violence around the drug trade, many adolescents sell drugs within a network or keep the drugs for themselves. For instance, in a study of adolescent offenders who sold drugs, approximately 70% kept more than half of the drugs for themselves (Shook, Vaughn, Goodkind, & Johnson, Reference Shook, Vaughn, Goodkind and Johnson2011). These low-level dealings, however, can escalate into physical altercations or bad feelings that erupt into aggressive acts in future interpersonal encounters.
Findings on the neural substrates of addiction and of violence are common. Although the reward pathway is considered key, other areas of the brain are involved. According to Volkow (Reference Volkow2003, p. 3): “We are beginning to understand that drugs exert persistent neurobiological effects that extend beyond the midbrain centers of pleasure and reward to disrupt the brain’s frontal cortex – the thinking region of the brain, where risks and benefits are weighed and decisions made.” Given that the adolescent (and even young adult) brain is still undergoing tremendous development, poor decisions and behavior dysregulation are unsurprising among adolescents who abuse drugs. This observation not only has broad implications for prevention and policy but also highlights the importance of unraveling the developmental origins of addiction and violence.
Childhood Precursors
It is critical that a biosocial approach to understanding drug abuse and violence be situated within the broader framework of a developmental, life-course perspective. This is due to the fact that, invariably, salient biological and socioenvironmental factors change and evolve as we age and develop across the spectrum of childhood, adolescence, and beyond. One clear example of this is the human brain. We have made the case that neuroscience plays an essential role in understanding both drug abuse and violence; however, it is now very well established that the brain is not hardwired or static across the life-course, but rather it looks and functions differently as we grow and are shaped by our experiences and the social environment. Along the same lines, our childhood, adolescent, and adult experiences are not independent of one another, but rather what we experience as children is profoundly related to our growth and development during later life stages. Indeed, drawing from the work of Glen Elder (Reference Elder1974) and others (Hser, Longshore, & Anglin, Reference Hser, Longshore and Anglin2007; Piquero, Jennings, & Barnes, Reference Piquero, Jennings and Barnes2012), we can see that a life-course perspective provides a critical foundation for understanding, preventing, and potentially interrupting long-term trajectories of drug abuse and violence.
One important developmental, life-course insight is that problem behavior can be observed across nearly the full spectrum of childhood. How early can we meaningfully identify problem behaviors among children? Arguably, quite early. Researchers have noted that there is substantial variation in the degree to which one-year-old infants exhibit noncompliance, throw temper tantrums, and exercise physical aggression (van Zeijl et al., Reference Van Zeijl, Mesman, Stolk, Alink, Van Ijzendoorn, Bakermans-Kranenburg and Koot2006; Carter, Briggs-Gowan, Jones, & Little, Reference Carter, Briggs-Gowan, Jones and Little2003). We would argue that caution should be exercised in making too much of infant “externalizing,” but such behavior becomes more and more meaningful as we get further into the toddlerhood stage. Indeed, substantial research has accrued examining the trajectories of behavior problems across toddlerhood (age 1 to 3) and the preschool years (age 3 to 5; Campbell, Reference Campbell2006). The bulk of these studies point to two core insights. First, problem behavior, including physical aggression, is quite common during early childhood. Indeed, it has been observed that the pinnacle in the prevalence physical aggression among humans is between the ages of 2 and 4 (Tremblay, Reference Tremblay, Morizot and Kazemian2015). Second, while the vast majority of children exhibit some degree of aggression during toddlerhood and the preschool years, these behaviors tend to dissipate as children move further and further away from their third and fourth birthdays. By the time children reach middle childhood (age 6 to 12), most have developed the cognitive, emotional, and social skills needed to deal with conflict such that physical aggression and other impulse-related outbursts are simply less common than among younger children.
What relevance does child problem behavior have when it comes to a broader, biosocial understanding of drug abuse and antisocial behavior across the life-course? Regretfully, child externalizing seems to very often set the stage for later behavior problems. Indeed, multiple longitudinal studies have found that young people who consistently exhibit elevated levels of aggression and/or other forms of problem behavior across early and middle childhood are far more likely than their peers to persist in this behavior into adolescence and, frequently, well into adulthood (Aguilar et al., Reference Aguilar, Sroufe, Egeland and Carlson2000; Broidy et al., Reference Broidy, Nagin, Tremblay, Bates, Brame, Dodge and Lynam2003; Campbell et al., Reference Campbell, Spieker, Burchinal and Poe2006; Farrington, Lambert, & West, Reference Farrington, Lambert and West1998; Moffitt, Reference Moffitt1993; Piquero, Jennings, & Barnes, Reference Piquero, Jennings and Barnes2012; Roisman et al., Reference Roisman, Monahan, Campbell, Steinberg and Cauffman2010; Sampson & Laub, Reference Sampson and Laub2003). Along the same lines, a number of studies have also found that early and persistent behavior problems during childhood often portent of issues with alcohol and drug abuse at later developmental stages (Englund et al., Reference Englund, Egeland, Oliva and Collins2008; Moffitt et al., Reference Moffitt, Caspi, Harrington and Milne2002). Of course, we should be very clear in stating that not all children who exhibit problem behaviors during childhood persist in such behavior in later development stages. However, evidence clearly indicates that an important subset of individuals (roughly 3–6%) exhibit serious behavior problems early on in life and, regretfully, often continue to struggle for decades.
What predicts early and persistent behavior problems among children? A review of the extant literature suggests that the answer is – perhaps unsurprisingly – both biological and social. One biologically based factor that is often examined is temperament. Temperament is a largely innate and stable characteristic that is moderately heritable (Tuvblad et al., Reference Tuvblad, Isen, Baker, Raine, Lozano and Jacobson2010) and can be measured in children as young as 3 to 6 months of age (Gartstein & Rothbart, Reference Gartstein and Rothbart2003). Fundamentally, temperament refers to the ways in which individuals experience and, in turn, regulate their responses to the world around them (Rothbart, Reference Rothbart2011). Evidence indicates that children with “difficult” temperaments (i.e., those who are highly active, difficult to soothe, and easily distressed) at 6 months of age are far more likely to exhibit physical aggression as toddlers and during the early preschool years (Naerde et al., Reference Nærde, Ogden, Janson and Zachrisson2014). Moreover, other studies suggest that a temperamental “lack of control” at age 3 to 5 is robustly predictive of externalizing behavior further down the road (Althoff, Verhulst, Rettew, Hudziak, & Vander Ende, Reference Althoff, Verhulst, Rettew, Hudziak and van der Ende2010; Caspi et al., Reference Caspi, Henry, McGee, Moffitt and Silva1995; Fearon et al., Reference Fearon, Reiss, Leve, Shaw, Scaramella and Ganiban2014). Simply put, temperament is a salient example of how our biological makeup relates to the risk of behavior problems during childhood as well as drug abuse and violence later on in life.
Of course, social and contextual factors also play a very important role in the etiology of behavior problems during childhood and beyond. Our assessment is that biological factors, such as temperament, are clearly part of the puzzle, but by no means the sole driver of behavior problems. Indeed, substantial research points to the importance of social factors such as parenting practices, family stress, home environment, and community violence exposure as central in understanding behavior problems across the life-course. In fact, findings from a variety of prospective studies suggest that a “difficult temperament” may be most troublesome when combined with exposure to environmental adversities such as harsh parenting and family stress. In other words, the etiology of child behavior problems is likely more a function of bio × social interactions, rather than independent biological and/or social risk factors.
Adolescent Expressions
As noted above, a full account of the etiology of drug abuse and antisocial behavior is not complete without giving careful consideration to the childhood years. However, adolescence – typically understood as beginning around the time of puberty and stretching into the early 20s – is a critically important time in which the full flowering of problem behaviors often begins to take place. Moreover, in keeping with a life-course perspective, adolescent trajectories related to drug abuse and antisocial behavior are most fully understood when situated within a broader understanding of the intrapersonal, contextual, and behavioral factors that precede – and, indeed, follow – such behavior. Below, we delve into two issues that are situated in the framework and logic of a developmental, life-course perspective and of direct relevance to adolescent drug abuse and violence.
One important issue relates to “child-persistent” versus “adolescent-onset” behavior problems. Above, we noted that a minority of individuals begin to exhibit serious behavior problems early in life and that, for some, such behaviors persist into the adolescent years and beyond. This is certainly the case, but many of us know from our own experience that there is a degree to which taking part in risky behavior – including alcohol and drug use and delinquency – is somewhat normative. As argued by Terrie Moffitt (Reference Moffitt1993), a bit of teenage experimentation with substance use and relatively minor property crimes, status offenses, and skirmishes is certainly disconcerting but not unexpected. Moffitt theorized that adolescent-onset problem behaviors were primarily rooted in a gap between biological and social maturity such that young people act out and imitate antisocial peers in an effort to be viewed as full-fledged adults. Whereas child-persistent behavior problems are understood to be rooted primarily in biosocial risk (e.g., genetic susceptibility, neurological impairment, elevated social adversity), adolescent-onset problem behavior is viewed more in normative developmental terms. We are of the mind that a bit of (short-lived) risky behavior is, indeed, normative. That being said, let’s be unequivocal in stating that growing evidence from a variety of studies indicates quite clearly that it is not developmentally normative for youth to begin to frequently take part in serious antisocial behaviors during adolescence. More precisely, whether it begins during childhood or around the onset of puberty, research has repeatedly shown that the lives of individuals who exhibit serious behavior problems during adolescence are often marked by early childhood issues (e.g., difficult temperament, lower cognitive functioning) and long-term difficulties (e.g., mental health and substance abuse problems) that are anything but the norm.
In reference to children, we discussed early-onset behavior problems, such as physical aggression, and their implications for long-term development; however, up to this point, we have not discussed early- versus later-onset substance use. This is partially the case because, by and large, substance use among children prior to puberty is relatively uncommon. However, by the time early adolescence rolls around, we begin to see an important minority of young people taking part in alcohol and other drug use. Research suggests that the age at which young people begin initiating substance use seems to matter. Indeed, studies have repeatedly found alcohol and drug use during the first few years of adolescence – typically operationalized as either age 11/12 to 14 or prior to age 15 – to be linked with academic and mental health issues, later substance use and delinquency, and the diagnosis of substance use disorders during adulthood (Anthony & Petronis, Reference Anthony and Petronis1995; Chen, Storr, & Anthony, Reference Chen, Storr and Anthony2009; DeWit, Aldaf, Offord, & Ogborne, Reference DeWit, Adlaf, Offord and Ogborne2000; Ellickson, Tucker, & Klein, Reference Ellickson, Tucker and Klein2003; Meier et al., Reference Meier, Hall, Caspi, Belsky, Cerdá, Harrington and Moffitt2016; Odgers et al., Reference Odgers, Caspi, Nagin, Piquero, Slutske, Milne and Moffitt2008). Importantly, while results are not entirely uniform, it should be noted that even studies that have accounted for genetic factors (i.e., the same genes that predict early alcohol/drug use may predict later substance abuse and behavioral problems) suggest that early substance use initiation seems to have long-term developmental implications.
Overall, we can say with confidence that biosocial factors are related to both the onset of antisocial behavior and drug use during adolescence. Whereas early theorizing was rooted in the understanding that adolescent-onset antisocial behavior was best understood as primarily a socially influenced phenomenon, a mounting body of evidence indicates that there is, in part, a genetic basis to the emergence of serious conduct problems during the teenage years. Similarly, evidence certainly seems to indicate that the prevention of alcohol and drug use initiation during adolescence is important with respect to the development of later behavior problems, including adult drug abuse, addiction, and criminal offending. Importantly, however, research also seems to indicate that alcohol and other drug use at an early age may be best understood not as a cause of addiction in adulthood, but rather as a reflection of underlying genetic risk related to both early-onset use and addiction. Similarly, evidence also suggests that drug abuse and other high-risk and delinquent behaviors may be related to overlapping genetic and neurological vulnerabilities related to risk-taking and externalizing in general.
Prevention and Treatment Efforts
As discussed in this chapter, violent behavior and its genetic and neurobiological underpinnings are highly dependent and must be viewed through a social, economic, and environmental lens. Targeting a single dimension, such as a school environment, is commonly less effective than those programs that target multiple dimensions (such as the school environment and students within the school) in preventing the onset of violence or problem behavior. As a result, some of the most effective prevention programs target multiple layers of influence to maximally optimize protective factors and reduce (or mitigate) risk. This framework upon which many intervention and prevention programs today are built is known as the social ecological model (also referred to more generally as ecological models). The individual (or micro) level is the most common prevention target, including demographic factors, personality and attitudes. This level would include modifiable (e.g., amenable to prevention or intervention) biological targets. Among adolescents, life skills training and educational programs targeting individual-level risk and protective factors are common.
Few interventions or prevention programs inherently target gene–environment interactions for various reasons. First, an array of ethical issues present when considering genetic testing for violence or problem behavior predisposition among youth. Second, these genetic influences are less modifiable than the environment and are therefore less amenable to intervention. However, interventions might be most cost-effective if targeted towards youth predisposed (with biological and environmental susceptibility) to violent behavior. Because the research on many of these gene–environment interactions remains inconclusive, it is important to consider the mechanisms of action to move this field of research beyond exploration and towards causational modeling. For example, are deviant peers directly related to antisocial behavior? Or, is this relationship, which is particularly strong among males, driven by testosterone? These are important considerations for future research to continue considering the effect of biological, in addition to psychosocial, effects on addiction that are useful implications for prevention science.
Noticeably absent from this chapter is a discussion of how biosocial theory fits into current prevention (and intervention) programs. A long-lasting criticism of biosocial theories of crime relates to its implications, as if “bad apple” youth would be identified and assigned a scarlet letter to follow them through life. This premonition could not be further from the intent of biosocial theories, which are derived from sound clinical research and applied to social science in an effort to improve, not detract from, population health. The role of biosocial theory in prevention remains unclear, and the undeniable biological underpinnings of deviant behavior have not yet been optimally incorporated into our evidence-based programs. However, there is little doubt that innovators in prevention science will integrate fundamental biologic elements, including susceptibility, into indicated (or targeted) substance use prevention programs.
Let’s consider this analogy in the domain of medical screening (e.g., the genetic testing for Huntington’s, which has no cure). This test was highly controversial given its implications – the patient testing positive will have a high likelihood of developing a debilitating chronic disease, which will result in premature mortality and severe morbidity. Considering these circumstances, why would someone get this test? Although the prognosis is poor, Huntington’s may be prevented through use of creatine (Science Daily, 2014), healthy behaviors, exercise, and diligently following a restrictive diet. Or, a person might wish to cognitively prepare for the development of symptoms. If a susceptibility test for addiction or criminality were created, results might be sought by diligent parents who wish to gain training to ensure all protective measures are optimized in their home. A positive test does not indicate that a child will become addicted; instead, it is a call to maximize all possible protective factors and minimize risks. The conversation must be shifted away from labeling youth and towards expanding our capacity for prevention, acknowledging the strong evidence in support of biological roots for problem behavior.
Policy Implications
Violent behavior is just beginning to be viewed as a public health problem. This movement shifts the attention away from the criminal justice system (e.g., tertiary prevention and treatment, which is costly) and toward health behavior, risk reduction, and policy (e.g., primary and secondary prevention, which is less costly). To this end, a recent APHA (2013) policy statement, entitled “Defining and Implementing a Public Health Response to Drug Use and Misuse,” identified the movement to shift away from the criminalization of drug possession and use as a core component of a public health approach. Specifically, the APHA statement highlights how the mass incarceration of drug users in the criminal justice system has made treatment more difficult, created other public health problems, and – not inconsequentially – contributed to the problem of mass incarceration in the United States. Because the majority of persons housed in United States jails and prisons are incarcerated for drug-related offenses (Carson & Golinelli, Reference Carson and Golinelli2012), even a small dent in the rate of substance use at the population level will result in great cost savings in criminal justice.
Understanding violence and antisocial behavior and how it fits within the framework of public health allows us to leverage the strengths of epidemiology and public health practice and policy to address the challenges associated with these problem behaviors. Because violence and antisocial behavior are so closely coupled with mental health problems and substance use disorders, healthcare resources must be dedicated to address the root of these problems rather than a behavioral manifestation (e.g., violence that is due to untreated mental health conditions may be resolved through medication management). In this way, application of a public health approach to violence opens up exciting possibilities with respect to large-scale health-promotion efforts designed to prevent violence (and its known causes) before they begin, increase treatment access for sufficient causes, and reduce drug-related health consequences among those in the criminal justice system.
Conclusion
The struggle to develop effective policies and treatments for drug abuse and violence among young people in a free society is a consistent theme. However, there is growing awareness that a science of addiction and human violence have much to offer. As previously mentioned, there is typically a substantial time lag between the accumulation of research findings and their adoption by practitioners and policy-makers. In this chapter we considered the underlying genetic, neuro-scientific, and socio-developmental foundations of the co-occurrence of drug abuse and violence that have a direct bearing on policy and treatment. Consistent with our holistic definition of a generalized neuro-dysregulation syndrome of altered behavior that is genetically and developmentally sensitive and socially and culturally contingent over time, each domain along the biosocial continuum is a necessary component. We encourage research and prevention that embraces this inherent dynamism.
Introduction
Typically defined as the intentional harming of an individual who is motivated to avoid such treatment (Anderson & Bushman, Reference Anderson and Bushman2002), aggression is a construct of substantial interest to public health broadly. In its most serious instantiations, aggression in the form of violent crime (e.g., murder, aggravated assault) can constitute a direct threat to human life, while other forms of aggression may function in more subversive, yet potent, ways (e.g., domestic violence, bullying). Aggression can also manifest subtly in the form of exclusionary social practices. Noting how this heterogeneity in behavioral content is belied by homogeneity of harmful intent, personality theorists have attempted to identify a coherent set of traits that is helpful in explaining meaningful patterns of this behavior. To the extent that a trait model such as the Big Five (John & Srivastava, Reference John and Srivastava1999) or Five-Factor Model (FFM; McCrae & John, Reference McCrae and John1992) has the capacity to account for variation in individuals’ tendencies to act aggressively toward another, it must be seriously considered as an important theoretical account of aggression. For the purposes of this review, we focus our attention on how one of the most extensively researched structural models of personality, the FFM, contributes to our understanding of aggressive behavior. Additionally, we review several other well-researched personality configurations (e.g., Dark Triad) that have been linked to aggression, and conclude that key common traits underlie these various instantiations.
FFM Background
The Big Five/FFM has enjoyed a rich developmental history (for a review see Digman, Reference Digman1990). Henceforth we use “FFM” to refer to the Big Five and Five-Factor Models of personality, given their substantial theoretical and empirical overlap. This model is rooted in the lexical hypothesis, which has two key premises: (1) differences that are most socially relevant in people’s lives will become symbolically represented in language, and (2) the more important a difference, the more likely it is to be delineated from other words and expressed as an independent term (e.g., Ashton & Lee, Reference Ashton and Lee2005). Put differently, this hypothesis purports that the aspects of human personality most important to differentiate have become the most robustly encoded into language, and these differences can be quantified. Once distilled, these traits can be conceived of as enduring patterns of behavior, cognition, and emotion.
One widely used operationalization of the FFM (e.g., NEO PI-R, Costa & McCrae, Reference Costa and McCrae1992) consists of a hierarchical structure that includes Neuroticism (i.e., a tendency to experience strong negative emotions such as anger, sadness, vulnerability), Extraversion (i.e., an approach orientation, or a tendency to seek out social interaction, exciting activities, and positive emotions), Openness to experience (i.e., willingness to consider different values, ideas, experiences, and tastes), Agreeableness (i.e., a tendency to be gentle with and trusting of others, and motivation toward social harmony and cooperation), and Conscientiousness (i.e., a driven, organized approach to work and the ability to delay gratification and act nonimpulsively). This model also posits the existence of six narrower, more specific facets that comprise each domain, although other faceted models of the FFM exist as well (e.g., DeYoung, Quilty, & Peterson, Reference DeYoung, Quilty and Peterson2007). The FFM is arguably the predominant model of general and pathological personality (e.g., Samuel & Widiger, Reference Samuel and Widiger2008; APA, 2013) and, as such, benefits from a large empirical literature. It enjoys considerable convergent and divergent validity across raters and cultures (Carlson, Vazire, & Oltmanns, Reference Carlson, Vazire and Oltmanns2013; McCrae, Costa, Del Pilar, Rolland, & Parker, Reference McCrae, Costa, Del Pilar, Rolland and Parker1998), presence in childhood (John, Caspi, Robins, Moffitt, & Stouthamer-Loeber, Reference John, Caspi, Robins, Moffitt and Stouthamer-Loeber1994), temporal stability across the lifespan (Blonigen, Reference Blonigen2010), and support from behavioral genetics research (Yamagata et al., Reference Yamagata, Suzuki, Ando, One, Kijima and Yoshimura2006). More importantly for our purposes, it has been linked to a wide variety of important outcomes such as happiness, relationship satisfaction, and job performance (Ozer & Benet-Martinez, Reference Ozer and Benet-Martinez2006).
FFM and Aggression
The most recent meta-analytic review of the relations between FFM personality and aggression included aggregated data from over 30 studies published since 2000 with over 10,000 participants. As expected, the authors found evidence that Agreeableness, Conscientiousness, and Neuroticism were the most consistent correlates of aggression. For instance, Agreeableness was negatively associated with elevated levels of aggression (effect size [ES, r]: −0.33; Jones, Miller, & Lynam, Reference Jones, Miller and Lynam2011). It is notable that this moderate relationship was consistent across population (e.g., community vs. prison, student vs. community) and method of assessment (self-report vs. laboratory paradigms). Next, Conscientiousness also evinced a significant negative effect size (ES) on aggression (ES: −0.18). In contrast, Neuroticism evinced a small, but positive relationship with aggression (ES: 0.17). Though moderation analyses suggested that these effect sizes varied slightly by sample characteristics (e.g., gender), all effects were in the expected direction (Table 13.1).
Table 13.1 Effect Sizes of FFM and Aggression/ASB
| Aggression | Proactive Aggression | Reactive Aggression | ||
|---|---|---|---|---|
| Five-Factor Model | ||||
| Neuroticism | 0.17a | 0.07b | 0.42b | |
| Angry hostility | 0.22a | 0.34b | 0.59b | |
| Extraversion | −0.03a | −0.07b | −0.18b | |
| Warmth | −0.23a | −0.30b | −0.29b | |
| Openness | −0.10a | −0.13b | −0.07b | |
| Agreeableness | −0.33a | −0.49b | −0.50b | |
| Straightforwardness | −0.25a | −0.38b | −0.29b | |
| Altruism | −0.26a | −0.39b | −0.39b | |
| Compliance | −0.26a | −0.44b | −0.58b | |
| Conscientiousness | −0.18a | −0.12b | −0.15b | |
| Dark Triad | ||||
| Psychopathy | 0.44c | 0.36d | 0.35d | |
| Self-centered impulsivity | 0.42i | |||
| Fearless dominance | −0.04i | |||
| Narcissism | 0.23c | 0.11e | 0.53e | |
| Machiavellianism | 0.39c | 0.33f | 0.32f | |
| Psychopathy | Violent Offending | Violent Recidivism | ||
| Factor 1 | 0.40g | 0.16h | ||
| Interpersonal | 0.44d | 0.42d | ||
| Affective | 0.38d | 0.31d | ||
| Factor 2 | 0.57g | 0.24h | ||
| Lifestyle | 0.41d | 0.49d | ||
| Antisocial | 0.28d | 0.30d |
Note: FFM facets are reported only if their effect sizes are │0.20│.
a Jones, Miller, & Lynam (Reference Jones, Miller and Lynam2011); b Miller & Lynam (Reference Miller and Lynam2006); c Vize, Lynam, Collison, & Miller (Reference Vize, Lynam, Collison and Miller2018); d Blais. Solodukhin, & Forth (Reference Blais, Solodukhin and Forth2014); e Bettencourt, Talley, Benjamin, & Valentine (Reference Bettencourt, Talley, Benjamin and Valentine2006); f Jonason, Duineveld, & Middleton (Reference Jonason, Duineveld and Middleton2015); g Lestico, Salekin, DeCoster, & Rogers (Reference Leistico, Salekin, DeCoster and Rogers2008); h Walters (Reference Walters2003); i Miller & Lynam (Reference Miller and Lynam2012).
A second meta-analysis, consisting of studies published before 2001, corroborates these findings, indicating that low Agreeableness and Conscientiousness are the primary FFM traits associated with antisocial behavior (ASB; Miller & Lynam, Reference Miller and Lynam2001). Although ASB is not synonymous with aggression, virtually all operationalizations of ASB include aggressive behavior, and both constructs are subsumed under a latent “externalizing” factor of psychopathology (Krueger, Caspi, Moffitt, & Silva, Reference Krueger, Caspi, Moffitt and Silva1998). Additionally, numerous studies have confirmed the relations between low Agreeableness and Conscientiousness and a range of ASBs, including aggression, substance use, and risky sex (e.g., Miller, Lynam, & Jones, Reference Miller, Lynam and Jones2008). A third meta-analysis found that psychoticism (as operationalized by Eysenck’s three-factor model; e.g., Eysenck & Eysenck, Reference Eysenck and Eysenck1985), a trait that blends low Agreeableness and low Conscientiousness, was a robust correlate of ASB (Cale, Reference Cale2006; r = 0.39).
In addition to these findings at the broader domain level, Jones et al. (Reference Jones, Miller and Lynam2011) also meta-analyzed the relations between FFM and aggression at the narrower, facet level. Importantly, the pattern in the facet analyses closely mirrored the findings from the domain level. For the sake of parsimony, we report only those with effect sizes > │0.20│. Using this criterion, three facets of Agreeableness (Straightforwardness [ES: 0.25]; Altruism [ES: −0.26]; Compliance [ES: −0.26]), one facet of Neuroticism (Angry-Hostility [ES: 0.21]), and one facet of Extraversion (Warmth [ES: −0.23]) were the most notable correlates, although several other expected facets neared this arbitrary cut-off (e.g., Trust, Modesty, and Tenderminded facets of Agreeableness; Deliberation facet of Conscientiousness).
FFM and Proactive/Reactive Aggression
A classic (albeit controversial, see Bushman & Anderson, Reference Bushman and Anderson2001) distinction in the aggression literature is made between proactive (PA) and reactive (RA) forms of aggression. Describing the former, Albert Bandura (1983, p. 57) noted, “A great deal of aggression is prompted by its anticipated benefits … the instigator is the pull of expected success, rather than the push of aversive treatment.” This description emphasizes that aggression can have an instrumental function and can be used to bring about some desired end (e.g., intimidation, retribution, monetary reward, etc.). Alternately, RA can be understood in terms of the frustration-aggression model (e.g., Dollard, Miller, Doob, Mowrer, & Sears, Reference Dollard, Miller, Doob, Mowrer and Sears1939), in which aggression is understood as a response to an obstacle to goal attainment, and in light of its instigating precursors, such as an influx of anger. RA can be thought of as a “hot-headed,” angry, impulsive response to provocation, and PA is thought to be “cold-blooded,” premeditated, and deliberate, although Bushman and Anderson note that these characteristics can describe both forms of aggression. There is also research suggesting these variants are underscored by different cognitive processes (e.g., Brugman et al., Reference Brugman, Lobbestael, Arntz, Cima, Schuhmann, Dambacher and Sack2015; Dodge & Coie, Reference Dodge and Coie1987). It is worth noting, however, that self-reported RA and PA tend to be substantially correlated, suggesting that individuals who engage in one form are likely to engage in the other as well (Miller & Lynam, Reference Miller and Lynam2006).
Though the literature on personality differences that underlie PA and RA is small, a study by Miller and Lynam (Reference Miller and Lynam2006) revealed an interesting distinction consistent with theoretical characterizations. First, both constructs were significantly correlated with Agreeableness (PA, r = −0.49; RA, r = −0.50) and its underlying facets. The correlations with the Conscientiousness domain were also similar (PA, r = −0.15; RA, r = −0.12). However, while RA displayed a medium-to-large correlation with Neuroticism (r = 0.42), PA did not show a significant correlation (r = 0.07). It is also of note that while PA did evince significant relationships with the Neuroticism facets Angry Hostility (PA, r = 0.34; RA, r = 0.59) and Impulsiveness (PA, r = 0.22; RA, r = 0.36), RA was more strongly correlated to every Neuroticism facet. This is consistent with the conception of RA as a relatively more emotion (i.e., anger, frustration)-driven response, though it is notable that this data suggests that PA is also characterized to some degree by this description. In line with the previously discussed meta-analytic findings, neither PA nor RA were significantly correlated with the Extraversion domain, but both were significantly negatively correlated with the Warmth facet (PA, r = −0.30; RA, r = −0.29) and the Positive Emotions facet (PA, r = −0.23; RA, r = −0.35). Neither PA nor RA were significantly correlated with the Openness domain, and there were only minor facet-level differences.
Despite these differences, it must be emphasized that overall, PA and RA displayed relatively similar correlations with the FFM. In fact, Miller and Lynam (Reference Miller and Lynam2006) reported that PA and RA themselves displayed a large correlation (r = 0.54), and an even higher similarity index across the 30 FFM facets (rICC = 0.79), which takes into account the absolute similarity (i.e., convergence in shape and magnitude) of two sets of correlations. Thus, regardless of the distinct theoretical underpinnings of PA and RA, their trait profiles are quite similar. Although some scholars have gone as far as to suggest that it is “time to pull the life-support plug” on the distinction between PA and RA (Bushman & Anderson, Reference Bushman and Anderson2001), we are more cautious and suggest that these small, yet detectable FFM profile differences warrant further investigation. For example, extant neurobiological research suggests that men who scored highly on a measure of RA exhibit structural differences in the amygdala compared to a nonaggressive group and to a primarily PA group (Rosell & Siever, Reference Rosell and Siever2015), suggesting that different neural underpinnings may, in part, underlie these processes. Particular attention should be paid to the role of negative emotional reactivity as both theory and the reported FFM findings suggest this is critical to possible differences in these forms of aggression.
FFM and Relational Aggression
Relational aggression is a relatively new, but increasingly popular focus among aggression researchers, who have recognized that deliberate harm can also take the form of damage to another’s social status and interpersonal relationships (Archer & Coyne, Reference Archer and Coyne2005). This form of aggression may be a particularly worthwhile focus of research given the increasing opportunities for its manifestation through social media outlets. Though research on this type of aggression’s relation to personality is nascent compared to the general aggression literature, initial studies suggest that it shares many of the same correlates as other forms. For instance, a study by Tackett, Daoud, De Bolle, and Burt (Reference Tackett, Daoud, Bolle and Burt2013) on children and adolescents found that a general aggression measure (Agg) and a relational aggression (RAgg) measure were both significantly negatively correlated with trait Agreeableness (Agg r = −0.65; RAgg = −0.32), as well as Conscientiousness (Agg r = −0.37; RAgg r = −0.18). Both general (r = 0.62) and relational aggression (r = 0.33) were also positively related to Neuroticism. Relatively similar patterns of findings have also been reported in young adult samples (e.g., Miller, Zeichner, & Wilson, Reference Miller, Zeichner and Wilson2012). Thus, while further research is encouraged, current findings suggest that like other types of aggression, similar personality traits underlie this form.
Pathological Personality Traits and Aggression
Over the past 20 to 30 years, greater interest has been shown in developing comprehensive, dimensional trait models of personality pathology that capture more extreme, more impairing, and/or more distressing traits (e.g., Livesley, Schroeder, Jackson, & Jang, Reference Livesley, Schroeder, Jackson and Jang1994; Widiger, Reference Widiger1993). Research into these dimensional models of personality pathology/personality disorder (PD) came about as a result of a variety of theoretical and empirical critiques (e.g., Clark, Reference Clark2007; Widiger & Trull, Reference Widiger and Trull2007) of the categorical diagnostic models within the American Psychiatric Association’s Diagnostic and Statistical Manual of Mental Disorders (DSM). The wealth of data supporting the benefits of a dimensional, trait approach to the conceptualization, assessment, and diagnosis of PDs led to the inclusion of a trait-based “Alternative Model” in the fifth edition of the DSM (DSM-5; American Psychiatric Association, 2013). This model emphasizes that the core of personality disturbance is impairment in personality functioning (Criterion A) coupled with evidence of maladaptive elevations in dimensional traits (Criterion B; APA, 2013). The inclusion of these traits was designed to allow for elaboration of individual differences in personality psychopathology, especially given the evidence that dimensional personality traits can reproduce the nomological networks of the traditional DSM PD constructs (see Miller, Reference Miller2012 for a review).
This research culminated in the publication of the Personality Inventory for DSM-5 (PID-5; Krueger et al., Reference Krueger, Derringer, Markon, Watson and Skodol2012), a 220-item self-report measure developed to assess the five pathological traits detailed by the Alternative Model (Negative Affect, Detachment, Psychoticism, Antagonism, Disinhibition). Although similar to the FFM in structure and content, the Alternative Model was explicitly developed as a pathological model of personality psychopathology to be used in clinical and research settings. PID-5 and NEO PI-R domains consistently evidence moderate to strong convergence, with the exception of mixed findings in regard to the relation between Openness and Psychoticism (e.g., Few et al., Reference Few, Miller, Rothbaum, Meller, Maples, Terry and MacKillop2013; c.f. Gore & Widiger, Reference Gore and Widiger2013).
Most extant literature to date on the relation of the DSM-5 pathological traits to aggression involves the constructs of Antagonism and Disinhibition, which parallel the FFM domains Agreeableness and Conscientiousness (Gore & Widiger, Reference Gore and Widiger2013) – the two strongest personality correlates of aggression and antisocial behavior. Initial studies support moderate to large correlations between these DSM-5 personality domains and behavior dysfunction and aggression (Anderson et al., Reference Anderson, Sellbom, Ayearst, Quilty, Chmielewski and Bagby2015). However, empirical examinations of the DSM-5 traits in relation to behavioral or functional outcomes are sparse (Al-Dajani, Gralnick, & Bagby, Reference Al-Dajani, Gralnick and Bagby2016). Though findings have been reported supporting the role of Antagonism in aggressive driving violations (Beanland, Sellbom, & Johnson, Reference Beanland, Sellbom and Johnson2014), studies assessing a range of aggressive behavior are lacking.
Specific Personality Profiles
In addition to the examination of these general and pathological traits’ relations to aggression and antisocial behavior, there is much interest in specific, multi-trait configurations that are linked both conceptually and empirically with aggression. In the following section, we focus on a set of related constructs – psychopathy, narcissism, and Machiavellianism – that are often studied together as part of the Dark Triad (Paulhus & Williams, Reference Paulhus and Williams2002), as well as diagnostic profiles described in DSM-5.
Psychopathy
Psychopathy is a personality profile that is characterized by traits such as egocentricity, callousness, superficial charm, and recklessness/disinhibition and is associated with persistent, pervasive antisociality. Research suggests that psychopathy, like other PDs, can be conceptualized as a configuration of general personality traits (e.g., Miller, Lynam, Widiger, & Leukefeld, Reference Miller, Lyman, Widiger and Leukefeld2001; Miller & Lynam, Reference Miller and Lynam2003) with a large emphasis on Agreeableness/Antagonism and Conscientiousness/Disinhibition.
In general, psychopathy manifests a robust correlation with aggression. A recent meta-analytic review of psychopathy’s relation with aggression as part of the Dark Triad reported an effect size (r) of 0.44 (Vize, Lynam, Collison, & Miller, Reference Vize, Lynam, Collison and Miller2018). Generally, these findings have been unequivocal across forensic, community adult, youth, and psychiatric samples (e.g., Blais, Solodukhin, & Forth, Reference Blais, Solodukhin and Forth2014; Leistico, Salekin, DeCoster, & Roger, Reference Leistico, Salekin, DeCoster and Rogers2008; Miller, Rausher, Hyatt, Maples, & Zeichner, Reference Miller, Rausher, Hyatt, Maples and Zeichner2014). Of note, psychopathy has been extensively studied in forensic populations, where it is vastly overrepresented (~ 15–30% of forensic populations vs. 0.2–3.3% 12-month community prevalence rate; Cale & Lilienfeld, 2006; APA, 2013). Thus, it is unsurprising that psychopathy has been linked to violent criminal offending, as well rates of violent recidivism (Lestico, Salekin, DeCoster, & Rogers, Reference Leistico, Salekin, DeCoster and Rogers2008; Walters, Reference Walters2003).
However, there are substantial, ongoing debates about the structure of psychopathy and which components are more or less relevant and important to the construct (e.g., Lilienfeld et al., Reference Lilienfeld, Patrick, Benning, Berg, Sellbom and Edens2012; Miller & Lynam, Reference Miller and Lynam2012). These debates are especially important in this context, as these components manifest differential relations with outcomes like aggression. These debates are too substantial to delve into in detail in the current chapter, but surround the importance/centrality of a set of traits deemed fearless dominance (Lilienfeld & Widows, Reference Lilienfeld and Widows2005), boldness (Patrick, Fowles, & Krueger, Reference Patrick, Fowles and Krueger2009) and emotional stability (Few, Miller, & Lynam, Reference Few, Miller and Lynam2013). This configuration of traits involves resilience to stress and decreased susceptibility to negative emotions along with an assertive interpersonal approach. Although these traits can be found in historical accounts of psychopathy, including Cleckley’s (Crego & Widiger, Reference Crego and Widiger2016) and several existing measures (Lilienfeld, Watts, Francis Smith, Berg, & Latzman, Reference Lilienfeld, Watts, Smith, Berg and Latzman2015), concerns have been raised that these traits demonstrate an almost entirely adaptive nomological network, including mostly null correlations with externalizing-related outcomes like aggression (see Miller & Lynam, Reference Miller and Lynam2012 for a meta-analytic review) that most consider critical to psychopathy. Conversely, the more consensual traits – callousness, lack of remorse, egocentricity, impulsivity, irresponsibility – demonstrate reliably moderate to strong correlations with externalizing problems such as aggression (meta-analytic r = 0.42; Miller & Lynam, Reference Miller and Lynam2012). Although there has been some suggestion that certain aspects of psychopathy are primarily related to proactive but not reactive aggression (Reidy, Shelley-Tremblay, & Lilienfeld, Reference Reidy, Shelley-Tremblay and Lilienfeld2011), this is inconsistent with meta-analytic evidence that demonstrates a comparable link between psychopathy and both reactive and proactive aggression (Blais et al., Reference Blais, Solodukhin and Forth2014).
Narcissism
Narcissism is another personality/PD construct with a long theoretical history (e.g., Freud; Foss, Reference Foss2014) that is of substantial interest to researchers from a wide array of behavioral science domains (e.g., Campbell & Miller, Reference Campbell and Miller2011). Despite extant controversies regarding the nature of narcissism as a construct (Miller, Lynam, Hyatt, & Campbell, in press), academics, clinicians, and lay-persons alike agree that this profile is typically characterized by traits such as grandiosity, callousness, entitlement, exploitativeness, and noncompliance (Miller, Lynam, Siedor, Crowe, & Campbell, Reference Miller, Lynam, Siedor, Crowe and Campbell2016; Thomas, Wright, Lukowitsky, Donnellan, & Hopwood, Reference Thomas, Wright, Lukowitsky, Donnellan and Hopwood2012).
Vize and colleagues’ recent meta-analytic review reported a small to moderate effect size for narcissism’s relation to aggression (r = 0.23). Whether this relation is contingent upon the presence of provocation is less clear. Bettencourt, Talley, Benjamin, and Valentine (Reference Bettencourt, Talley, Benjamin and Valentine2006) conducted a meta-analysis examining the relationship between personality traits and aggressive behavior under “provoked” vs. “neutral” conditions, and results suggested that trait narcissism was only related to aggressive behavior after provocation. This is consistent with findings from the seminal study by Bushman and Baumeister (Reference Bushman and Baumeister1998; see also Twenge & Campbell, Reference Twenge and Campbell2003) that posited the “ego-threat” hypothesis, which suggests that narcissistic individuals will react particularly aggressively when faced with an insult to their status, potentially as a method of conserving their inflated self-perception.
However, several recent publications have identified links between narcissism and aggression even in the absence of an explicit ego-threat (e.g., Lobbestael, Baumeister, Fiebig, & Eckel, Reference Lobbestael, Baumeister, Fiebig and Eckel2014; Reidy, Foster, & Zeichner, Reference Reidy, Foster and Zeichner2010), which suggests that while an ego-threat may potentiate elevated levels of aggression in narcissistic individuals, it might not be necessary. Thus, it remains unclear whether aggression is rooted in an emotional response (e.g., anger) coupled with an appetite for retaliation, as suggested by the narcissistic rage hypothesis, a separate process arising from an interest in demonstrating social dominance (Kohut, Reference Kohut, Eissler, Freud, Kris and Solnit1972; Krizan & Johar, Reference Krizan and Johar2015), or is simply the manifestation of a general behavioral pattern for narcissistic individuals to aggress more often due to interests in dominance paired with low levels of empathy and compliance. From a general trait perspective, narcissism is characterized by low Agreeableness (Miller et al., in press), suggesting that narcissistic individuals may be globally prone to being aggressive.
Machiavellianism
Similar to its Dark Triad counterparts, Machiavellianism exhibits a strong association with aggression (meta-analytic r = 0.39; Vize et al., Reference Vize, Lynam, Collison and Miller2018). It is characterized by a cynical, dog-eat-dog world view, a dispositional tendency to manipulate others, as well as strategic, nonimpulsive thinking (Jones & Paulhus, Reference Jones and Paulhus2011). It is moderately to largely associated with self-reported reactive and proactive aggression (Jonason, Duineveld, & Middleton, Reference Jonason, Duineveld and Middleton2015), physical aggression, verbal aggression, anger, and hostility (Jones & Neria, Reference Jones and Neria2015; Stead & Fekken, Reference Stead and Fekken2014).
The association between Machiavellianism and aggression is difficult to interpret, however, in light of recent work that suggests that, as measured, Machiavellianism may simply be a proxy for psychopathy (McHoskey, Worzel, & Szyarto, Reference McHoskey, Worzel and Szyarto1998; Miller, Hyatt, Maples-Keller, Carter, & Lynam, in press). As measured, Machiavellianism’s trait profile is nearly indistinguishable from psychopathy (Miller et al., in press; O’Boyle et al., Reference O’Boyle, Forsyth, Banks, Story and White2015), and characterized by interpersonal antagonism, disinhibition, and anger – traits known to be substantially correlated with aggression (e.g., Jones et al., Reference Jones, Miller and Lynam2011). This similarity is problematic due to the inclusion of disinhibition (e.g., impulsivity; failure to delay gratification; low ambitiousness), which is counter to theoretical descriptions of the construct and experts’ conceptualizations (Miller, Hyatt, et al., in press). As such, the relation between Machiavellianism and aggression can be difficult to interpret, as it includes disinhibitory content that should be excluded from measures of Machiavellianism. In fact, when one removes the variance shared by measures of psychopathy and Machiavellianism, the latter no longer demonstrates substantial correlations with externalizing outcomes (Sleep, Lynam, Hyatt, & Miller, Reference Sleep, Lynam, Hyatt and Miller2016).
Borderline Personality Disorder
Borderline personality disorder (BPD) is characterized by a pervasive pattern of instability in interpersonal relationships, emotion regulation, self-image, and impulse control (Skodol et al., Reference Skodol, Gunderson, Pfohl, Widiger, Livesley and Siever2002). Despite its heterogeneous nature, aggression and emotional dysregulation are considered to be core features (Mancke, Herpertz, & Bertsch, Reference Mancke, Herpertz and Bertsch2015), and expert ratings and meta-analytic review suggest that aggression-relevant traits are central to the construct (e.g., angry/hostility, noncompliance, deceitfulness; Samuel & Widiger, Reference Samuel and Widiger2008). Indeed, empirical work has linked BPD to elevated aggression across methodology (Dougherty, Bjork, Huckabee, Moeller, & Swann, Reference Dougherty, Bjork, Huckabee, Moeller and Swann1999; Russel, Moskowitz, Zuroff, Sookman, & Paris, Reference Russel, Moskowitz, Zuroff, Sookman and Paris2007). Longitudinal findings suggest that over the course of a year, 73% of individuals with BPD engaged in violent behavior, although this relation is diminished once BPD’s shared variance with psychopathy is removed (Newhill, Eack, & Mulvey, Reference Newhill, Eack and Mulvey2009).
BPD has primarily exhibited relations to reactive forms of aggression (Berenson, Downey, Rafaeli, Coifman, & Leventhal-Paquin, Reference Berenson, Downey, Rafaeli, Coifman and Leventhal-Paquin2011; Gardner, Archer, & Jackson, Reference Gardner, Archer and Jackson2012), although links to both reactive and proactive aggression have been reported (Lobbestael, Cima, & Lemmens, Reference Lobbestael, Cima and Lemmens2015). Indirect evidence for a link between BPD symptoms and aggressive, antisocial behaviors comes from the degree to which BPD appears to be over-represented in prison settings, particularly among female offenders (Sansone & Sansone, Reference Sansone and Sansone2009). Indeed, 30% of aggression-prone populations (e.g., prison inmates) exhibit borderline features (Black et al., Reference Black, Gunter, Allen, Blum, Arndt, Wenman and Sieleni2007). Recently, Jackson, Sippel, Mota, Whalen, and Schumacher (Reference Jackson, Sippel, Mota, Whalen and Schumacher2015) reviewed BPD’s relation to interpersonal violence (IPV), and found that individuals with BPD were more likely to perpetrate IPV against a partner. Separate findings indicate that BPD is associated with verbal aggression (South, Turkheimer, & Oltmanns, Reference South, Turkheimer and Oltmanns2008), minor to severe physical violence (Whisman & Schronbrun, Reference Whisman and Schonbrun2009), and spousal homicide (e.g., 1/3 of men incarcerated for spousal homicide; Dixon, Hamilton-Giachritsis, & Browne, Reference Dixon, Hamilton-Giachritsis and Browne2008).
Personality Traits and Aggression: Conclusions and Mechanisms
In sum, a robust body of literature supports the importance of certain personality traits in the prediction of aggression. The evidence is resoundingly clear that the FFM domains low Agreeableness/Antagonism and low Conscientiousness/Disinhibitionare among the most robust individual difference correlates of aggression. Though high Neuroticism/Negative Affect has also been linked to aggression, a meta-analytic examination of the facet-level data suggests much of this relation is driven by traits related to anger/hostility. Interestingly, this trait has sometimes been considered to be “interstitial” in nature, in that it seems to be an admixture of negative emotionality and antagonism. Thus, scales of this trait tend to load on both domains in structural analyses. It is significant to note that the contributing roles of these traits are consistent across the aforementioned models, as well across populations, assessment method, and the lifespan.
Although these relations are well-established, questions as to how these traits manifest “in the moment” to lead to aggressive responding requires further investigation. In other words, the proximal mechanisms by which low trait Agreeableness (i.e., Antagonism) and Conscientiousness (i.e., Disinhibition) function to precipitate aggressive behavior are still largely under investigation. However, advances in this area have emerged. For example, Bresin and Robinson (Reference Bresin and Robinson2015) found that participants who were low in trait Agreeableness displayed a relatively exaggerated tendency to view negative images (e.g., a snarling dog) for a longer period of time than positive images, in line with previous research suggesting that similar individuals also have a difficult time disengaging from antisocial stimuli (Wilkowski, Robinson, & Meier, Reference Wilkowski, Robinson and Meier2006).
Complementary research has also found that low Agreeableness is related to a particular tendency to interpret ambiguous social behavior as hostile, and also an increased likelihood of responding to these cues with aggression (Miller, Lynam, & Jones, Reference Miller, Lynam and Jones2008). This is consistent with other findings that suggest that low Agreeableness is associated with the tendency to view power assertion as an appropriate conflict resolution strategy (Graziano, Jensen-Campbell, & Hair, Reference Graziano, Jensen-Campbell and Hair1996). Furthermore, individuals low in Agreeableness appear to be more susceptible to aggressing after exposure to violent priming, which may be due to their relative inability to activate prosocial thoughts when primed with antisocial stimuli (Meier, Robinson, & Wilkowski, Reference Meier, Robinson and Wilkowski2006).
Implications for Intervention
In contrast to the large body of research on FFM personality and aggression, very little research has been conducted into personality per se as the target of intervention. This dearth of study in this area is likely related to the impressive temporal stability of traits (test-retest coefficients from childhood to older adulthood = 0.31–0.74; Blonigen, Reference Blonigen2010), as well as the contested subject of personality change as a result of therapeutic intervention (e.g., Rogers, Reference Rogers2007; c.f. Samstag, Reference Samstag2007). However, Hopwood et al. (Reference Hopwood, Morey, Donnellan, Samuel, Grilo, McGlashan and Skodol2013) reported that symptoms of personality pathology are significantly less stable than personality traits, suggesting that pathological manifestations of traits may be more dynamic and susceptible to environmental dynamics. Thus, instead of specifically addressing trait-level change, it is likely that interventions focusing on individually relevant, practical change will proffer the most benefit. It is in these efforts that the research on mechanisms of antagonism will be able to speak most loudly.
For example, understanding that individuals low in Agreeableness are prone to misinterpreting ambiguous situations as hostile should inform intervention efforts that are aimed at addressing this distorted, deleterious cognitive pattern. In this way, micro-level modifications to individual tendencies may summate to meaningful change. This approach is represented by the Fast Track program, which has demonstrated modest but meaningful efficacy in decreasing aggression and increasing prosocial behavior, in part by teaching social skills and conflict resolution (Bierman et al., Reference Bierman, Coie, Dodge, Greenberg, Lochman, McMahon and Pinderhughes2010). On the other hand, intensive interventions efforts that involve significant environmental changes (e.g., daily in-home consultations with an interventionist), such as Multisystemic Therapy, have shown to be successful in reducing arrest rates and aggression in peer relationships (Henggeler, Melton, & Smith, Reference Henggeler, Melton and Smith1992). Thus, while few interventions have purported to directly target personality as the dependent variable in a treatment outcome study, researchers have made strides in reducing aggression by directing efforts at 1) mechanisms by which antagonistic tendencies lead to aggressive behavior and 2) the environmental contingencies that contribute to aggression. In sum, while personality is a multifactorially determined construct involving the intersection of biology and environment, there is cause for tentative optimism in treatments that aim to reduce aggressive behavior.
Conclusion
In this chapter, we have reviewed the evidence for the importance of personality in understanding aggression using a basic, general trait perspective to guide our review. A rich literature, including numerous meta-analyses, strongly supports the importance of several key traits: low Agreeableness (Antagonism) and low Conscientiousness (Disinhibition). Although anger/hostility (Neuroticism) and (lack of) warmth (Extraversion) seem to be important facets from other domains, Antagonism and Disinhibition are most central to FFM accounts of aggression.
In addition to general and pathologically oriented trait models, we also reviewed several frequently studied personality profiles that have been of interest to clinical practitioners, criminologists, and social-personality psychologists alike. Antagonism and Disinhibition appear to generally underlie the majority of the multidimensional configurations psychopathy, narcissism, and Machiavellianism (e.g., Paulhus & Williams, Reference Paulhus and Williams2002). As such, investigations into the underlying mechanisms by which antagonistic and disinhibited traits lead to aggress against others in different contexts and manner will also inform our understanding of how these broader constructs exert their effects on relevant outcomes like aggression. It is also hoped these investigations will prove useful in the development of interventions aimed at reducing aggressive behavior.
To this end, we encourage further investigation into the role of personality in aggression, and suggest collaboration across disciplines, including (but not limited to) clinical psychology, social-personality psychology, criminology, educational psychology, developmental psychopathology, neuroscience, and genetics. In conjunction, we encourage the burgeoning research on the mechanisms that underlie the relations between personality and aggression, especially since their bivariate relations are no longer in question. These types of mechanistic approaches are more likely to yield information that may be germane to the development of prevention and intervention approaches.


