
2 results
Rheology of a concentrated suspension of spherical squirmers: monolayer in simple shear flow
-
- Journal:
- Journal of Fluid Mechanics / Volume 914 / 10 May 2021
- Published online by Cambridge University Press:
- 05 March 2021, A26
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
A concentrated, vertical monolayer of identical spherical squirmers, which may be bottom heavy, and which are subjected to a linear shear flow, is modelled computationally by two different methods: Stokesian dynamics, and a lubrication-theory-based method. Inertia is negligible. The aim is to compute the effective shear viscosity and, where possible, the normal stress differences as functions of the areal fraction of spheres
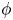 $\phi$, the squirming parameter
$\phi$, the squirming parameter 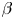 $\beta$ (proportional to the ratio of a squirmer's active stresslet to its swimming speed), the ratio
$\beta$ (proportional to the ratio of a squirmer's active stresslet to its swimming speed), the ratio 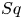 $Sq$ of swimming speed to a typical speed of the shear flow, the bottom-heaviness parameter
$Sq$ of swimming speed to a typical speed of the shear flow, the bottom-heaviness parameter 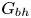 $G_{bh}$, the angle
$G_{bh}$, the angle 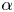 $\alpha$ that the shear flow makes with the horizontal and two parameters that define the repulsive force that is required computationally to prevent the squirmers from overlapping when their distance apart is less than a critical value. The Stokesian dynamics method allows the rheological quantities to be computed for values of
$\alpha$ that the shear flow makes with the horizontal and two parameters that define the repulsive force that is required computationally to prevent the squirmers from overlapping when their distance apart is less than a critical value. The Stokesian dynamics method allows the rheological quantities to be computed for values of 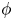 $\phi$ up to
$\phi$ up to 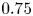 $0.75$; the lubrication-theory method can be used for
$0.75$; the lubrication-theory method can be used for 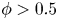 $\phi > 0.5$. For non-bottom-heavy squirmers, which are unaffected by gravity, the effective shear viscosity is found to increase more rapidly with
$\phi > 0.5$. For non-bottom-heavy squirmers, which are unaffected by gravity, the effective shear viscosity is found to increase more rapidly with 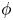 $\phi$ than for inert spheres, whether the squirmers are pullers (
$\phi$ than for inert spheres, whether the squirmers are pullers (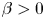 $\beta > 0$) or pushers (
$\beta > 0$) or pushers (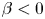 $\beta < 0$); it also varies with
$\beta < 0$); it also varies with 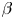 $\beta$, although not by very much. However, for bottom-heavy squirmers the behaviour for pullers and pushers as
$\beta$, although not by very much. However, for bottom-heavy squirmers the behaviour for pullers and pushers as 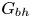 $G_{bh}$ and
$G_{bh}$ and 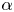 $\alpha$ are varied is very different, since the viscosity can fall even below that of the suspending fluid for pushers at high
$\alpha$ are varied is very different, since the viscosity can fall even below that of the suspending fluid for pushers at high 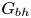 $G_{bh}$. The normal stress differences, which are small for inert spheres, can become very large for bottom-heavy squirmers, increasing with
$G_{bh}$. The normal stress differences, which are small for inert spheres, can become very large for bottom-heavy squirmers, increasing with 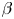 $\beta$, and varying dramatically as the orientation
$\beta$, and varying dramatically as the orientation 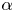 $\alpha$ of the flow is varied from 0 to
$\alpha$ of the flow is varied from 0 to 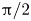 ${\rm \pi} /2$. A major finding is that, despite very different assumptions, the two methods of computation give overlapping results for viscosity as a function of
${\rm \pi} /2$. A major finding is that, despite very different assumptions, the two methods of computation give overlapping results for viscosity as a function of 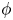 $\phi$ in the range
$\phi$ in the range 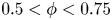 $0.5 < \phi < 0.75$. This suggests that lubrication theory, based on near-field interactions alone, contains most of the relevant physics, and that taking account of interactions with more distant particles than the nearest is not essential to describe the dominant physics.
$0.5 < \phi < 0.75$. This suggests that lubrication theory, based on near-field interactions alone, contains most of the relevant physics, and that taking account of interactions with more distant particles than the nearest is not essential to describe the dominant physics.
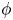
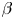
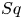
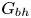
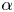
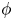
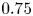
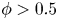
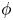
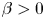
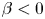
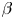
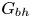
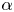
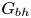
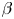
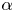
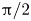
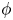
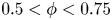
1 - Functional patterns of swimming bacteria
-
-
- Book:
- The Mechanics and Physiology of Animal Swimming
- Published online:
- 05 March 2012
- Print publication:
- 15 September 1994, pp 3-12
-
- Chapter
- Export citation
