



Introduction
We here describe lizard assemblages, which recently have been collected from the middle Miocene Palasava and the late Miocene Tapar localities of Kutch, Gujarat. Although this region is well known for its fossil mammals and other vertebrate fauna (Wynne, Reference Wynne1872; Lydekker, Reference Lydekker1876, Reference Lydekker1880; Prasad, Reference Prasad1962, Reference Prasad1964, Reference Prasad1967; Sahni and Mishra, Reference Sahni and Mishra1975; Thewissen and Bajpai, Reference Thewissen and Bajpai2009; Bajpai et al., Reference Bajpai, Domning, Das, Véléz-Juarbe and Mishra2010; Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010, Reference Bhandari, Pickford and Tiwari2015, Reference Bhandari, Kay, Williams, Tiwari, Bajpai and Hieronymus2018; Patnaik et al., Reference Patnaik, Sharma, Mohan, Williams, Kay and Chatrath2014; Singh et al., Reference Singh, Sharma, Patnaik, Singh and Singh2019, Reference Singh, Jukar, Patnaik, Sharma, Singh and Singh2020; Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2021; Sharma et al., Reference Sharma, Singha, Patnaik, Tiwaria, Singh, Singha, Choudhari and Lalotra2021), very little is known about the fossil squamates of the region—only several fossil snakes have been reported previously (Head et al., Reference Head, Mohabey and Wilson2007; Rage et al., Reference Rage, Folie, Rana, Singh, Rose and Smith2008; Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2021). However, India has an immense diversity of herpetofauna today, with 801 reptile species contributing to its recognition as a mega-diverse country and one of the global biodiversity hotspots (e.g., Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000; Palot, Reference Palot2015; Uetz et al., Reference Uetz, Freed and Hošek2021). Major lizard clades in the Indian region (see Mausfeld et al., Reference Mausfeld, Schmitz, Böhme, Misof, Vrcibradic and Rocha2002; Uetz et al., Reference Uetz, Freed and Hošek2021) include geckos (e.g., Hemidactylus, Cyrtodactylus, Dravidogecko), agamids (e.g., Calotes, Saara, Laudakia, Sitana), chameleons (Chamaeleo zeylanicus Laurenti, Reference Laurenti1768), skinks (Eutropis, Ablepharus, Lygosoma), and varanids (Varanus). But despite this high diversity, pre-Quaternary fossil records of lizards have been documented only occasionally from this subcontinent. This leads to a poor understanding of squamate history in this biogeographically interesting area during the Miocene, a time period that strongly influenced the characteristics of India's modern ecosystems.
One of the lizard clades with a better-documented history on this subcontinent is the acrodontan lizards. Agamids have been described from the lower Eocene of the Vastan Lignite Mine (Prasad and Bajpai, Reference Prasad and Bajpai2008), where a high diversity of acrodontan lizards was observed (Rana et al., Reference Rana, Augé, Folie, Rose, Kumar, Singh, Sahni and Smith2013), including Tinosaurus indicus Prasad and Bajpai, Reference Prasad and Bajpai2008. Later, the dentary of this taxon was also described from the early Eocene Tadkeshwar locality (Smith et al., Reference Smith, Kumar, Ranac, Folie, Solé, Noiret, Steeman, Sahni and Rose2016). However, there is a significant gap in regards to this clade in the Neogene—subsequent fossil herpetofaunas from India come from the Plio-Pleistocene and are represented by Uromastyx and Calotes from the Upper Siwaliks (Raghavan, Reference Raghavan1991; Patnaik and Schleich, Reference Patnaik and Schleich1998). The solitary report of an agamid from the central Narmada valley indicates the continuity of this clade in the Quaternary. The material was described by Joshi and Kotlia (Reference Joshi and Kotlia2010) as a new species, Agama schleichi (note, however, that the type material of this species is represented only by jaw fragments with general rather than unique tooth morphology; the problem is that tricuspid teeth of a similar form are present in many species of Agamidae; e.g., Smith et al., Reference Smith, Schaal, Sun and Li2011). Note that Tikiguania estesi Datta and Ray, Reference Datta and Ray2006, from the Tiki Formation, originally described as a Triassic lizard, is considered now as a Quaternary or late Tertiary agamid rather than Triassic in age (Hutchinson et al., Reference Hutchinson, Skinner and Lee2012). With regard to chamaeleonids, cranial remains potentially belonging to these unusual lizards have been reported from the late Miocene Haritalyangar locality (Sankhyan and Čerňanský, Reference Sankhyan and Čerňanský2016).
As for varanids, Varanus sivalensis Falconer, Reference Falconer and Murchison1868, has been described from Pliocene to Early Pleistocene deposits of Siwalik (Falconer, Reference Falconer and Murchison1868; Lydekker, Reference Lydekker1888). The Miocene record of this clade in India is represented by the material of Varanus sp. from the late Miocene of Raun, Jammu (Rage et al., Reference Rage, Gupta and Prasad2001) and Varanidae indet. from the middle part of the early Miocene lower Murree Group in northwest Himalaya (Kumar and Kad, Reference Kumar and Kad2003).
The new lizard assemblages described in the present paper provide the opportunity to document additional crucial data about the herpetofaunal diversity in India during the Miocene, which should prove useful for future analyses of past squamate diversity and faunal biogeography. The distribution of these previously unknown lineages in the fossil record from this area are discussed here as well.
Geological setting
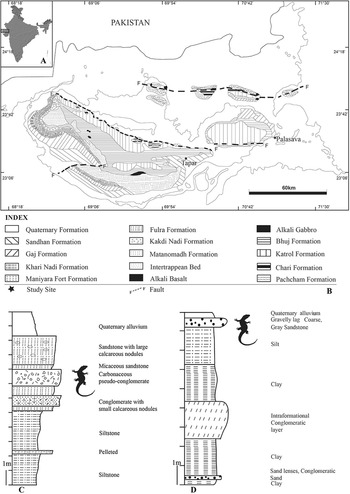
The Kutch basin of India has yielded vertebrate and invertebrate fossils ranging from Mesozoic to Pleistocene in age (Biswas, Reference Biswas1992, and reference therein). In this basin, Cenozoic sediments are deposited over the Deccan traps, which overlie older Mesozoic sediments. The Neogene deposits of Kutch are divided into the Khari Nadi, Chhasra/Gaj, and Sandhan formations (Biswas, Reference Biswas1992; Geological Survey of India, 2012). The Khari Nadi Formation, dated as early Miocene (23.03–20.43 ± 0.05 Ma) (Gradstein et al., Reference Gradstein, Ogg, Smith, Bleeker and Lourens2004; Patnaik et al., Reference Patnaik, Sharma, Mohan, Williams, Kay and Chatrath2014), comprises laminated to very thin bedded siltstone, fine-grained sandstone, and gypseous claystone, and is considered to have a conformable and gradational contact with the younger Gaj/Chhasra Formation and a weak erosional unconformity with the underlying Oligocene Maniyara Fort Formation (Biswas, Reference Biswas1992; Catuneanu and Dave, Reference Catuneanu and Dave2017; Sharma et al., Reference Sharma, Singha, Patnaik, Tiwaria, Singh, Singha, Choudhari and Lalotra2021). The lithologies of the Chhasra/Gaj Formation, which are dominated by olive green shale, gypseous shale, and claystone alternating with thin argillaceous limestone beds, has a disconformable contact with the younger Sandhan Formation (?Miocene–Pliocene; Fig. 1; Biswas, Reference Biswas1992; Catuneanu and Dave, Reference Catuneanu and Dave2017). Biswas (Reference Biswas1992) assigned an early Miocene age (Burdigalian) for the rocks of Gaj/Chhasra Formation, however the Geological Survey of India (2012) preferred to assign an age ranging from early to middle Miocene. The Sandhan Formation comprises well-sorted, medium- to coarse-grained, massive, micaceous sandstones, laminated siltstones, and claystone, with lenticular bodies of conglomerates, calcareous grits, and grayish mottled silty sandstone with calcareous nodules (Biswas, Reference Biswas1992).

Figure 1. Location of the studied material. (1) Map of India showing the study locality (represented by rectangular box); (2) geological map of Kutch (Geological Society of India, 2012); (3) stratigraphic column of the Tapar locality; (4) stratigraphic column of the Palasava locality. Shaded tetrapod figures indicate stratigraphic levels from which specimens were collected.
The present lizard assemblages were collected from the late Miocene conglomeratic beds of the Tapar site belonging to Sandhan Formation, and from the middle Miocene Palasava locality of the Gaj/Chhasra Formation (Fig. 1). The Tapar section comprises massive sandstone, siltstone, and clay beds alternating with intermittent conglomerate beds composed of calcareous nodules, agate pebbles, very coarse-grained sand, and mud clasts (Fig. 1). This conglomerate bed is richly fossiliferous and yields a variety of vertebrates, including the hominoid Sivapithecus, fragmented teeth of proboscideans (Deinotherium, Gomphotherium), rhinos, giraffids, equids, bovid, suids, rodents, turtles, crocodiles, teleosts, sharks, and batoids (Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010, Reference Bhandari, Pickford and Tiwari2015, Reference Bhandari, Bajpai, Flynn, Tiwari and Mandal2021; Singh et al., Reference Singh, Sharma, Patnaik, Singh and Singh2019, Reference Singh, Jukar, Patnaik, Sharma, Singh and Singh2020; Sharma et al., Reference Sharma, Singha, Patnaik, Tiwaria, Singh, Singha, Choudhari and Lalotra2021). The Tapar section has been dated as basal late Miocene (ca. 11–10 Ma) age based on the First Appearance Datum (FAD) of Hipparion and associated mammalian faunas (Bhandari et al., Reference Bhandari, Pickford and Tiwari2015). The lithosection of the Palasava locality consists predominantly of clay, silt, and sandstone and conglomerate alternating with successive deposits of coarse-grained sandstone and gravelly lag deposits at the top (see Fig. 1). The Palasava section, dated as middle Miocene (Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2021), has yielded a diverse mammalian fauna, including Sanitherium, Sivameryx, Brachypotherium, Zygolophodon, Gomphotherium, and Deinotherium, along with teleosts, sharks, batoids, turtles, crocodiles, snakes, and birds.
Materials and methods
The present fossil lizard remains were recovered from the late Miocene of Tapar and middle Miocene Palasava localities during the field season from 2017–2019. Medium-sized jaws were recovered from surface collection during the field work, and small-sized lizards were obtained from calcareous ferruginous conglomerate by screen-washing. The fossils were photographed with a Leica Stereozoom Microscope housed at the Department of Geology, Panjab University, Chandigarh, India. All described materials from the two localities are housed in the Department of Geology, Panjab University, Chandigarh and Department of Geology, Central University of Punjab under the collection codes: VPL/PU/KT, VPL/PU/KPS, BIOPS/CUP/KT, and BIOPS/CUP/KP.
The standard anatomical orientation system is used throughout this paper, and terminology describing individual bone structures is based on Rage and Augé (Reference Rage and Augé2010) and Čerňanský (Reference Čerňanský2010, for acrodont lizards). As for taxonomy of skinks, we use the term Scincidae here in its preferred family level usage for these lizards (see Uetz et al., Reference Uetz, Freed and Hošek2021), in contrast to the classification proposed by Hedges, Reference Hedges2014.
Repositories and institutional abbreviations
VPL/PU/KT, Vertebrate Palaeontology Laboratory/Panjab University/Kutch Tapar; VPL/PU/KPS, Vertebrate Palaeontology Laboratory/Panjab University/Kutch Palasava; BIOPS/CUP/KT, Biostratigraphy, Palaeontology and Sedimentology Laboratory/Central University of Punjab/Kutch Tapar; BIOPS/CUP/KP, Biostratigraphy, Palaeontology and Sedimentology Laboratory/Central University of Punjab/Kutch Palasava.
Systematic paleontology
Squamata Oppel, Reference Oppel1811
Iguania Cope, Reference Cope1864
Acrodonta Cope, Reference Cope1864
Agamidae Spix, Reference Spix1825
Uromastycinae Theobald, Reference Theobald1868
Genus Uromastyx Merrem, Reference Merrem1820
Type species
Stellio spinipes Daudin, Reference Daudin1802 (=Uromastyx aegyptia [Forskål, Reference Forskål1775]) is the type species of the genus Uromastyx Merrem, Reference Merrem1820.
cf. Uromastyx s.l. sp.
Figure 2
Occurrence
Palasava, middle Miocene.

Figure 2. Right dentary of cf. Uromastyx s.l. sp. from the middle Miocene Palasava locality; VPL/PU/KPS-11 in: (1) lateral; (2) medial; and (3) dorsal views.
Description
A right dentary is partially preserved (Fig. 2), lacking anterior and posterior terminations. The dentary is relatively short and robust, being deep ventrally; however, the posterior half of the ventral margin is strongly damaged. The Meckelian canal is fully open, being widely open posteriorly. It is roofed by a robust supra-alveolar shelf. The anteriormost portion of the dentary is broken off, thus the symphysis is not preserved; however, it can be estimated, based on the anterior preserved portion of the shelf, that the symphyseal region was strong because the anterior region of the shelf widens relative to the mid-region of the shelf. The posteroventral region of the shelf is broken off. A longitudinal groove appears to be present. It can be very likely interpreted as the dental groove, although this region is highly weathered (it is not clear whether it is a real structure or the result of a linear fracture; see Remarks). It seems to be somewhat narrow, forming a shallow furrow ventral to the subdental shelf. The subdental shelf bears teeth (14 tooth positions are still present). The lateral surface of the dentary is smooth. One elliptical labial foramen is located at the level of the seventh tooth position (counted from posterior). Others cannot be adequately identified; note, however, that the surface is covered by a calcite crust and sedimentary rock in several places. Posteriorly, the basal portion of the coronoid process is preserved, starting to rise dorsally at the end of the tooth row. This region is strongly built, implying a well-developed coronoid process. Unfortunately, the whole posterior portion of the dentary is broken off.
Dentition
The tooth implantation is acrodont (in terms of the location of implantation, where teeth are attached the crest of the jaw bone; see Jenkins et al., Reference Jenkins, Jones, Zikmund, Boyde and Daza2017). Teeth have blunt apices, so their appearance is rather rounded and they lack accessory cusps. The teeth increase in size posteriorly. In dorsal view, they are closely spaced. The tooth bases are more or less cylindrical, appear to touch one another, and, in at least some of the posterior teeth, overlap slightly, with the anterior edge of one tooth offset lingually relative to the posterior edge of the adjacent tooth (the imbricate alignment, sensu Čerňanský, Reference Čerňanský2011). No evidence of tooth replacement was found. Most of the crowns are highly abraded, especially those in the anterior region. The facets have a ventrolabial/dorsolingual orientation due to the dentary tooth row lying lingual to the maxillary tooth row (see Cooper and Poole, Reference Cooper and Poole1973).
Material
Right dentary VPL/PU/KPS-11.
Remarks
The teeth are abraded, as frequently occurs in old agamid individuals. The dentary described here resembles that of Uromastyx s.l. in the following combination of features (e.g., Averianov and Danilov, Reference Averianov and Danilov1996): (1) dentary heavily built; (2) the strong coronoid process (estimated based on the preserved basal portion); (3) blunt teeth with cylindrical bases, lacking accessory cusps; and (4) closely spaced teeth. Extant Uromastyx species are mainly herbivorous, which is correlated with shorter and deeper jaws than those of insectivorous forms (Herrel et al., Reference Herrel, Aerts, Fret and De Vree1999). Uromastyx is distributed in Africa and the Arabian Peninsula (Uetz et al., Reference Uetz, Freed and Hošek2021), but a member of the genus Saara (S. hardwickii [Gay, Reference Gray1827]), previously placed in Uromastyx, occurs in India today as well (Wilms et al., Reference Wilms, Böhme, Wagner, Lutzmann and Schmitz2009; Uetz et al., Reference Uetz, Freed and Hošek2021). For this reason, we use the term Uromastyx s.l. here. However, it should be noted that the coronoid process of the fossil dentary starts to rise at an unusual level—right behind the tooth row. In extant Uromastyx benti (Anderson, Reference Anderson1894) (Holmes et al., Reference Holmes, Murray, Chatrath, Attia and Simons2010a), extinct U. europaeus De Stefano, Reference De Stefano1903, (e.g., Rage and Augé, Reference Rage and Augé2015), and extant Indian S. hardwickii (see http://DigiMorph.org), there is a clearly visible gap between the end of the tooth row and the process. Although the gap is present in U. aegyptia as well (see http://DigiMorph.org), the bone starts to gradually rise dorsally more or less posterior to the tooth row in that species. However, some individual variations in the tooth row relating to this character cannot be fully excluded here. This can be seen in S. hardwickii, in which extension of the tooth row by additions posteriorly, as the jaw grows, was documented by Cooper and Poole (Reference Cooper and Poole1973). All these factors can be resolved only by further studies, where various ontogenetical series of males and females of this clade will be CT-scanned and compared. In any case, the feature in the fossil dentary from the Palasava locality does not contradict the allocation presented here. The potential presence of a dental groove deserves a comment here. A similar groove, as is present in the Palasava specimen, can be seen in Uromastyx from the Oligocene of Egypt (Holmes et al., Reference Holmes, Murray, Chatrath, Attia and Simons2010a, fig. 4a) and in some extant members (although usually weakly developed; see http://DigiMorph.org).
?Agamidae indet.
Figure 3
Occurrence
Tapar, late Miocene.

Figure 3. ?Agamidae indet. from the late Miocene Tapar locality; BIOPS/CUP/KT-304 (1, 2) and BIOPS/CUP/KT-302 (3, 4) in: (1, 3) lateral; and (2, 4) medial views.
Description
The material is poorly preserved; two small jaw fragments are available in the material (Fig. 3), both of which have two teeth attached to the bone. The otherwise smooth lateral portion of specimen BIOPS/CUP/KT-302 is pierced by a series of three small foramina (probably labial foramina).
Dentition
Tooth implantation is acrodont. Teeth are mediolaterally compressed, triangular, with more or less pointed apices. The central cusp is dominant, whereas the mesial and distal accessory cusps are weak, almost unseparated from the central one. They form sharp, only slightly bulged, mesial and distal edges. The teeth are closely packed, so the interdental gaps are small.
Materials
Two fragments of jaws BIOPS/CUP/KT-302 and BIOPS/CUP/KT-304.
Remarks
The dentition type of the material from Tapar appears different from that of the material described above as cf. Uromastyx s.l. from the older Palasava locality—the teeth are more pointed and the incipient accessory cusp is present, although only weakly developed. This might indicate the presence of two agamid types. However, this interpretation needs to be viewed with caution. The anterior portion is not preserved, and thus the crucial diagnostic features are missing here. In contrast with chamaeleonids, agamids retain a trace of the primitive pleurodont condition (often in caniniform anterior teeth) in the anterior region (Moody, Reference Moody1980). Moreover, the lingual walls of the teeth appear to be only very slightly taller than the labial walls. Thus, a subacrodont (subpleurodont, sensu Averianov and Danilov, Reference Averianov and Danilov1996) condition present in agamids can be observed only weakly here. The allocation of such fragmentary material is therefore questionable, and chamaeleonids cannot be fully excluded as well (for fossil chamaeleonids, see e.g., Čerňanský, Reference Čerňanský2010, Čerňanský et al., Reference Čerňanský, Herrel, Kibii, Anderson, Boistel and Lehmann2020a).
Scinciformata Vidal and Hedges, Reference Vidal and Hedges2005
Scincoidea Oppel, Reference Oppel1811
Scincidae Gray, Reference Gray1825
Scincidae indet.
Figure 4
Occurrence
Tapar, late Miocene.
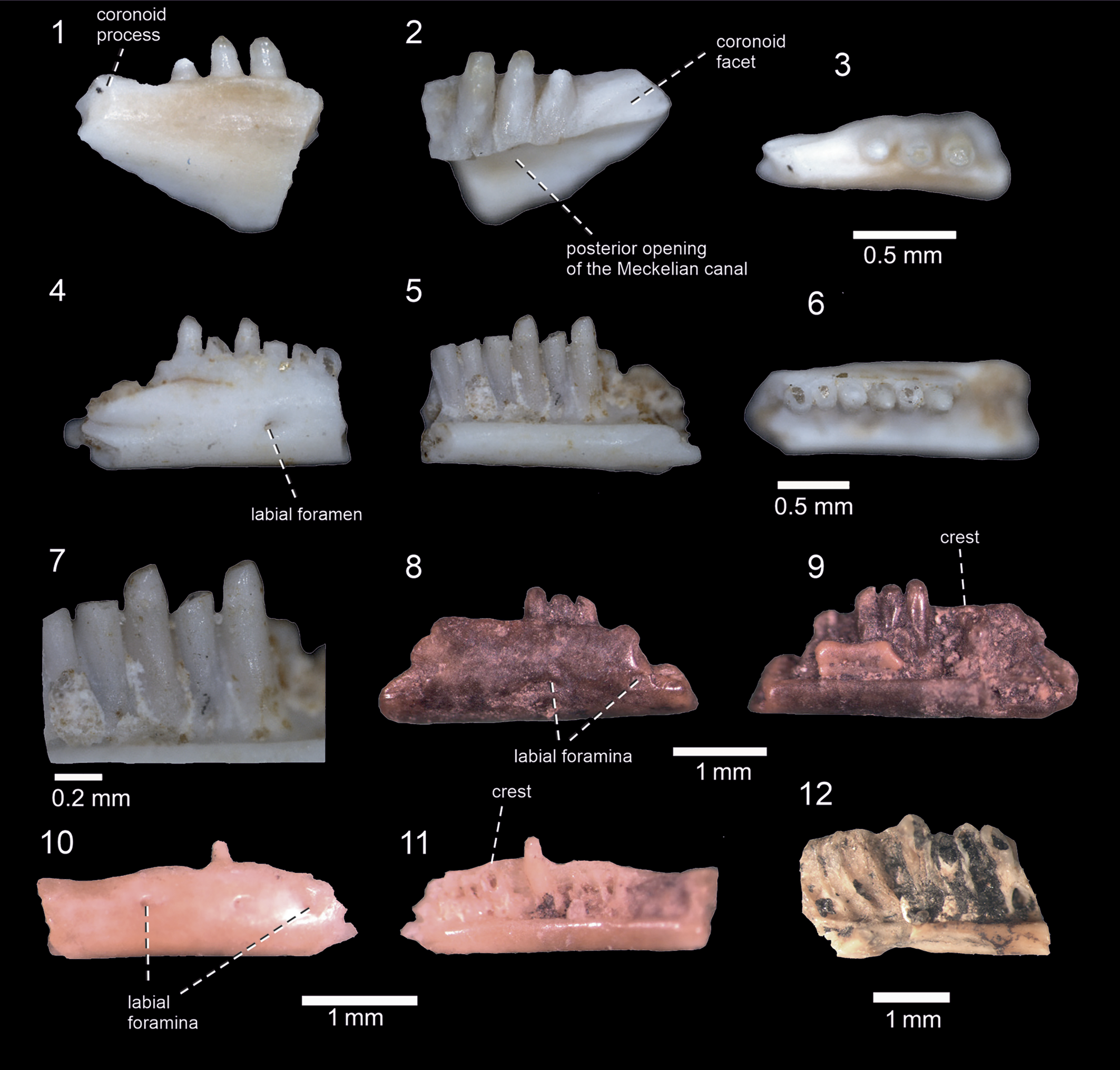
Figure 4. Right dentaries of Scincidae indet. from the late Miocene Tapar locality; BIOPS/CUP/KT-300 (1–3), BIOPS/CUP/KT-301 (4–7), VPL/PU/KT-717 (8, 9), VPL/PU/KT-718 (10, 11) and VPL/PU/KT-780 (12) in: (1, 4, 8, 10) lateral; (2, 5, 9, 11, 12) medial with (7) tooth detail; and (3, 6) dorsal views.
Description
The description is based on several dentaries, but all are only fragmentarily preserved (Fig. 4). The dentary is an anteroposteriorly elongated element with a narrow ventral section under the dental portion. In contrast, the alveolar crest, which supports teeth laterally, is high. The Meckelian canal is fully closed in the mid-region, but apparently open medially in the posterior section of the dentary. Although specimen VPL/PU/KT-718 represents the anterior portion of the dentary (note, however, that the anteriormost termination of this dentary is broken off and missing), no ventral opening of the Meckelian canal can be recognized here. Therefore, it can be estimated (if all specimens are conspecific; see Remarks) that any such opening was more likely small and located only in the anteriormost end of the dentary, rather than forming a longitudinal groove. The coronoid process is partly preserved—it slightly starts to rise dorsally, posterior to the tooth row. The process has a facet for the coronoid on its medial side. The smooth lateral surface of the bone is pierced by several labial foramina (at least three are present in VPL/PU/KT-718).
Dentition
Tooth implantation is pleurodont. The teeth are slender and high. Dorsally, they extend above the alveolar crest by one-third of their length. Interdental gaps are present (i.e., teeth are not in contact). The tooth crowns have rounded apices, bearing labial and lingual cusps. The lingual portion of the tooth crowns between the culmines lateres bears fine apicobasal ridges. The tooth crowns are slightly mesiodistally wider than the tooth bases, which are more or less swollen lingually.
Materials
Five right dentaries: BIOPS/CUP/KT-300, BIOPS/CUP/KT-301, VPL/PU/KT-717, VPL/PU/KT-718, and VPL/PU/KT-780.
Remarks
The most distinctive character of the Tapar skink dentaries is the partially closed Meckelian canal. It should be noted that all small skinks that develop the enclosed Meckel's groove tend to converge on a similar morphology. So very limited features, which are preserved in fragments from the Tapar locality, make them difficult to interpret. Moreover, even other clades that are present in India, such as geckos, have a closed Meckelian canal (e.g., Daza et al., Reference Daza, Bauer and Snively2014). The materials described here are also allocated to skinks based on their dentition (e.g., tooth crowns mesiodistally wide with apicobasal ridges and rounded apices, labial and lingual cusps present, interdental gaps present; see Kosma, Reference Kosma2004). However, an exception is VPL/PU/KT-718 (see Fig. 4.10, 4.11), in which only one tooth is preserved and, unfortunately, weathered. So in this particular specimen, its affinity to skinks cannot be strongly supported because we cannot exclude geckos here as well. Moreover, VPL/PU/KT-780 is completely without teeth, making allocation of this specimen difficult.
Among skinks, the closed Meckelian canal in the dentary can be observed in several lineages (Greer, Reference Greer1970, Reference Greer1974; Estes et al., Reference Estes, de Queiroz, Gauthier, Estes and Pregill1988; Shea, Reference Shea1999; Caputo, Reference Caputo2004; Hutchinson and Scanlon, Reference Hutchinson and Scanlon2009; Bolet et al., Reference Bolet, Daza, Augé and Bauer2015; Čerňanský et al., Reference Čerňanský, Syromyatnikova, Kovalenko, Podurets and Kaloyan2020b; Čerňanský and Syromyatnikova, Reference Čerňanský and Syromyatnikova2021): Mabuyinae, Egerniinae, Eugongylinae, Sphenomorphinae, Lygosominae, but also in Acontinae, whereas Scincinae and Ateuchosaurus have a fully open canal. The preserved morphology of the Tapar dentaries most resembles that of members of the clade Mabuyinae in the combination of the following features (Čerňanský et al., Reference Čerňanský, Syromyatnikova, Kovalenko, Podurets and Kaloyan2020b; Čerňanský and Syromyatnikova, Reference Čerňanský and Syromyatnikova2021): (1) the closed Meckelian canal; (2) narrow ventral section under the dental portion relative to high alveolar crest; (3) close tooth spacing, including the level of the crowns; and (5) tooth crown morphology. One member of the mabuyines, Eutropis, occurs in the Territory of India (Mausfeld et al., Reference Mausfeld, Schmitz, Böhme, Misof, Vrcibradic and Rocha2002), but note that one taxon from Indochina, Toenayar novemcarinata (Anderson, Reference Anderson1871), has been separated from Eutropis and placed in the monotypic genus Toenayar (Karin et al., Reference Karin, Metallinou, Weinell, Jackman and Bauer2016). The fossil dentaries described here share some features with Eutropis (e.g., the assumed restriction of the anterior opening of the Meckelian canal—only based on the preserved morphology available to us), which is in contrast to the prolonged anterior opening present in the Middle Eastern lineage, Heremites (Čerňanský and Syromyatnikova, Reference Čerňanský and Syromyatnikova2021). However, more complete material of this skink from Tapar is needed to support or exclude this claim. In any case, the fragmentary character of the currently available dentaries does not allow a precise allocation.
Anguimorpha Fürbringer, Reference Fürbringer1900
Varanidae Gray, Reference Gray1827 (sensu Estes et al., Reference Estes, de Queiroz, Gauthier, Estes and Pregill1988)
Genus Varanus Merrem, Reference Merrem1820
Type species
Lacerta varia Shaw in White, Reference White1790 (=Varanus varius) is the type species of the genus Varanus Merrem, Reference Merrem1820.
Varanus sp.
Figure 5
Occurrence
Palasava, middle Miocene.
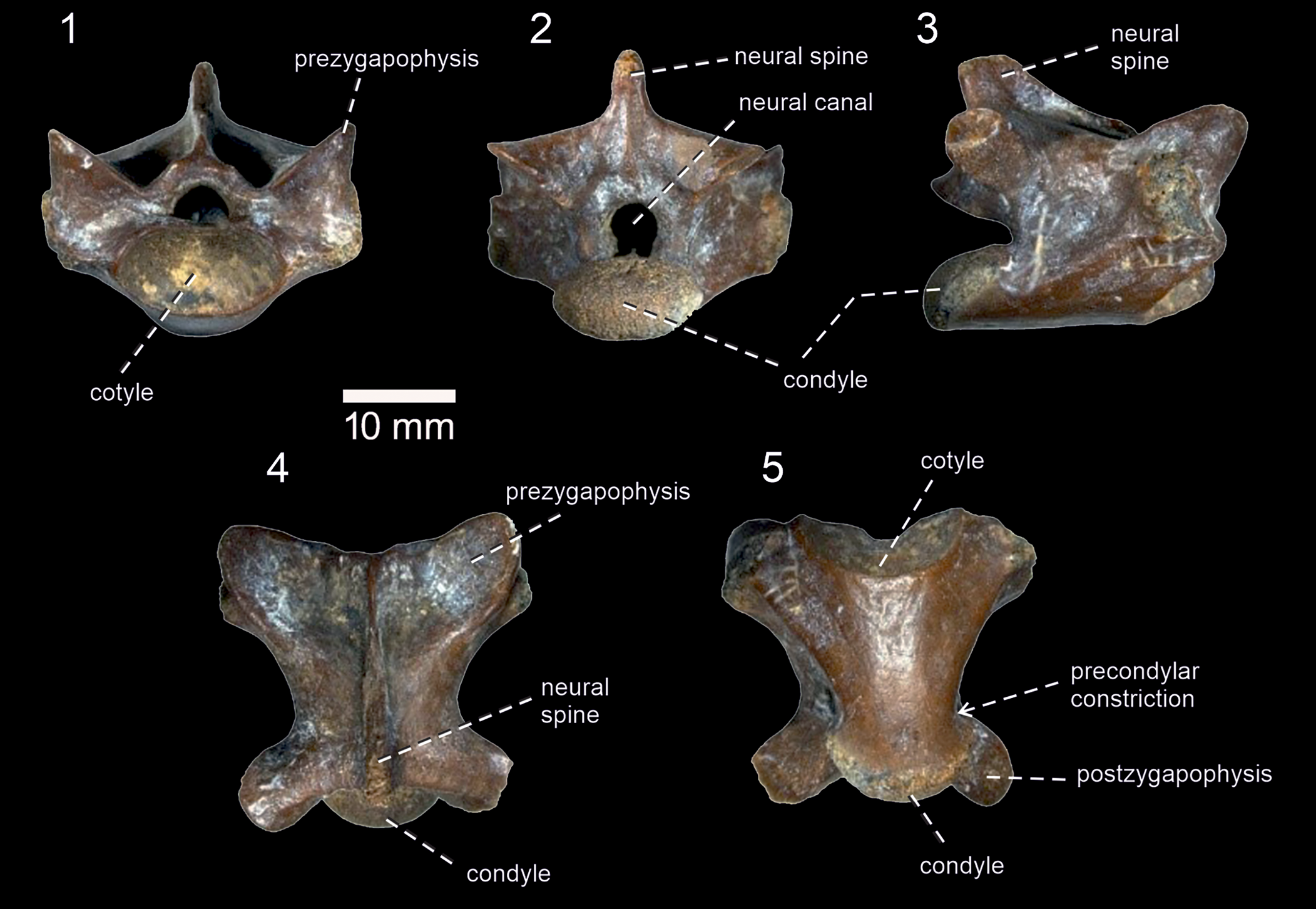
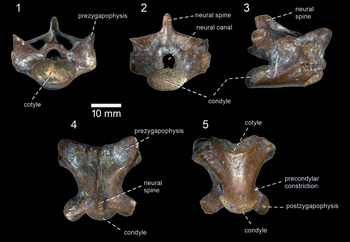
Figure 5. Varanus sp. from the middle Miocene Palasava locality; BIOPS/CUP/KP-240 in: (1) anterior; (2) posterior; (3) right lateral; (4) dorsal; and (5) ventral views.
Description
Only one isolated, but more or less complete, vertebra is preserved (Fig. 5). It represents a dorsal vertebra. Its maximum anteroposterior length is slightly higher than the maximum dorsoventral height. In lateral view, the neural spine is moderately high, trapezoidal in shape (it gradually narrows posterodorsally). It is slightly posteriorly inclined, but does not exceed the condyle posteriorly. In dorsal view, the neural spine has a narrower appearance—it somewhat widens only in the posterior elevated section. Anteriorly, it continues to the low medial ridge, which runs to the end of the neural arch. The pre- and postzygapophyses are well expanded anterolaterally and posterolaterally. A deep interzygapophyseal constriction is present, located directly in front of the postzygapophyses. The articular facets on the pre-and postzygapophysis are ovoid, although those on the prezygapophyses appear to be larger. They are markedly inclined dorsally. The synapophysis is well developed, laterally expanded, and located anteriorly to the midpoint of the vertebra. It is almost a vertical structure, which extends to the entire height of the centrum. The neural canal is a tunnel-like structure (roughly pentagonal), noticeably smaller in height and width than the cotyle. The cotyle and condyle are markedly depressed. The articular surface of the condyle is exposed mainly dorsally, whereas the cotyle is exposed ventrally. In ventral view, the centrum is roughly triangular in shape. The lateral margins of the centrum (=subcentral ridges) gradually become closer posteriorly, with more or less straight courses along almost the entire section, but they become concave in the posteriormost region. Here, the distinct precondylar constriction is present. As a result, the condyle, with its slightly laterally expanded lateral margins, exceeds the width of the posterior portion of the centrum. The centrum is slightly convex in anterior view. In lateral view, the ventral margin of the centrum is straight.
Material
One dorsal vertebra BIOPS/CUP/KP-240.
Remarks
The vertebra can be referred to Varanus on the basis of the markedly depressed cotyle and condyle, presence of a precondylar constriction, the articular surface of the condyle exposed dorsally with the cotyle exposed ventrally, and the presence of a well-demarcated anterior part (pars tectiformis) on the neural arch (Camp, Reference Camp1923; Bailon and Rage, Reference Bailon, Rage, Senut and Pickford1994; Venczel, Reference Venczel2006; Holmes et al., Reference Holmes, Murray, Yousry, Attia, Simons and Chatrath2010b; Georgalis et al., Reference Georgalis, Rage, de Bonis and Koufos2018).
Discussion
In the following, we discuss the composition of lizard fauna at both localities separately.
The middle Miocene Palasava locality
Two lizard clades can be recognized in the middle Miocene Palasava locality. In regards to paleoecology, the presence of Uromastyx s.l. is particularly interesting. Uromastyx and Leiolepis are remnants of one of the most ancient clades within Acrodonta (Moody, Reference Moody1980; Gauthier et al., Reference Gauthier, Kearney, Maisano, Rieppel and Behlke2012). Although different species of Uromastyx occupy a variety of habitats, members of this genus are generally considered to be adapted to rather arid environments (Estes, Reference Estes1983). Based on the occurrence of freshwater, terrestrial, estuarine, and marine taxa, however, Kapur et al. (Reference Kapur, Pickford, Chauhan and Thakkar2021) considered the Palasava locality to have been an environment with warm humid climatic conditions during the Miocene. Kapur et al. (Reference Kapur, Pickford, Chauhan and Thakkar2021) suggested a prevalence of woodland or forest, indicated by mammalian taxa such as a sanitheriid Sanitherium schlagintweiti von Meyer, Reference von Meyer1866, an anthracothere Sivameryx palaeindicus Lydekker, Reference Lydekker1877, rhinocerotid Brachypotherium perimense (Falconer and Cautley, Reference Falconer and Cautley1847), and the proboscideans Zygolophodon, Gomphotherium, and Deinotherium living close to freshwater and marine environments (represented by cyprinids, turtles, freshwater crocodiles, the shark Carcharinus sp., the myliobatiform Myliobatis sp., and the pristid Pristis sp.). It is plausible that the uromastycine record from Palasava represents only an occasional appearance of the group in this region. But allochthony also can be assumed (i.e., the material could be just washed into the area from regions upstream, where micro-habitats such as elevations and dry plateau could be present). These factors play an important role in the vertical distribution of species (Čerňanský, Reference Čerňanský2016). In any case, it is rather unlikely that these lizards, members of which are arid-adapted today, lived permanently in the area of the Palasava locality. Note, however, that the Palasava specimen presumably represents an extinct uromastycine—its preferred habitat is therefore unknown. Interestingly, this situation is not unique. A similar ecological contradiction in regards of fossil uromastycine lizards is also observed at the Oligocene Fayum locality in Egypt, which has been reconstructed as a swampy riverine system (Holmes et al., Reference Holmes, Murray, Chatrath, Attia and Simons2010a).
The other lizard group here is represented by a monitor lizard assigned to Varanus. The fossil record of varanids is rather sparse in Asia (Lydekker, Reference Lydekker1888; Rage et al., Reference Rage, Gupta and Prasad2001; Malakhov, Reference Malakhov2005; Suraprasit et al., Reference Suraprasit, Jaeger, Chaimanee, Chavasseau, Yamee, Tian and Panha2016; Broin et al., Reference Broin, Bailon, Augé and Rage2020; for an up-to-date table with all early and middle Miocene occurrences of Varanus worldwide, see Georgalis et al., Reference Georgalis, Abdel Gawad, Hassan, El-Barkooky and Hamdan2020a). The earliest confirmed Asian record of Varanus is reported from the early Miocene of Kazakhstan (Malakhov, Reference Malakhov2005). Fossils referable to Varanus also have been reported from the early Miocene of the Murree Group in northwest Himalaya (Kumar and Kad, Reference Kumar and Kad2003), late Miocene of Afghanistan (Broin et al., Reference Broin, Bailon, Augé and Rage2020), late Miocene of Raun, Jammu (Rage et al., Reference Rage, Gupta and Prasad2001), and middle Pleistocene of Thailand (Suraprasit et al., Reference Suraprasit, Jaeger, Chaimanee, Chavasseau, Yamee, Tian and Panha2016). Varanus pronini has been reported from the middle Miocene of western Kazakhstan (Zerova and Chkhikvadze, Reference Zerova, Chkhikvadze and Roček1986). In any case, the presence of Varanus in the middle Miocene of Palasava represents the oldest record of this taxon in the region of India south of the Himalayas. Varanus includes a clade of lizards that diversified into an exceptional range of body sizes, from the Komodo dragon (V. komodoensis Ouwens, Reference Ouwens1912) to the pygmy monitors (V. brevicauda Boulenger, Reference Boulenger1898, and V. primordius Mertens, Reference Mertens1942; see Collar et al., Reference Collar, Schulte and Losos2011). They represent mostly carnivorous (some frugivorous) forms that are widely distributed in tropical and subtropical regions (Pianka et al., Reference Pianka, King and King2004). The occurrence of this thermophilic reptile taxon in Palasava suggests a mean annual temperature that was not less than ~15°C (Böhme, Reference Böhme2003).
Aside from the agamid and varanid described here, the squamate assemblage includes snakes: Acrochordus and Python (see Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2021; Singh et al., Reference Singh, Patnaik, Čerňanský, Sharma, Singh, Choudhary and Sehgalin press) that also support the environmental conditions of the locality (i.e., the members of Acrochordus are fully aquatic piscivores) (Shine, Reference Shine1986) and Python is one of the most thermophilic squamates known (Shine, Reference Shine1986; Slip and Shine, Reference Slip and Shine1988; Snow et al., Reference Snow, Wolf, Greeves, Cherkiss, Hill and Mazzotti2010; Ivanov and Böhme, Reference Ivanov and Böhme2011; Georgalis et al., Reference Georgalis, Mayda, Alpagut, Şarbak and Güler2020b).
The late Miocene Tapar locality
Materials from the younger Tapar locality consist of acrodontan lizard jaw fragments, here questionably placed in agamids, and possibly a mabuyine skink. The potential presence of a mabuyine skink in the locality, as far as the fragmentary material can be interpreted, represents the first, but putative, evidence of this group in the Miocene of Asia. Although skinks are very successful and broadly distributed animals, comprising the largest, most diverse clade of lizards (~25% of lizards in the world) (Whiting et al., Reference Whiting, Bauer and Sites2003; Uetz et al., Reference Uetz, Freed and Hošek2021), their fossil record is extremely poor. Based on molecular data, the Mabuyinae group has been suggested to have its origin in Southeast Asia (Greer, Reference Greer1977; Honda et al., Reference Honda, Ota, Köhler, Ineich, Chirio, Chen and Hikida2003; Mausfeld and Schmitz, Reference Mausfeld and Schmitz2003; Karin et al., Reference Karin, Metallinou, Weinell, Jackman and Bauer2016). However, the complete absence of a pre-Quaternary Asian fossil record causes a problem. Thus, although relatively younger than the suggested Paleogene origin of the group (Karin et al., Reference Karin, Metallinou, Weinell, Jackman and Bauer2016), the fossils from India would be consistent with the previous paleobiogeographic scenario based on molecular data. The Tapar material might show the potential occurrence of this clade in Asia at least as early as the Miocene (for other Miocene records, see Čerňanský and Syromyatnikova, Reference Čerňanský and Syromyatnikova2021). Unfortunately, the material, as stated previously, is very fragmentary, so our interpretation is tentative.
Conclusions
Three major clades can be identified in the materials: Agamidae, Scincidae, and Varanidae. Few elements of lizards are identifiable at the generic level. Although fragmentarily preserved, the materials here described provide a rare opportunity to observe, at least partly, the past lizard diversity in Asia. It sheds important light on the composition and paleobiogeographic distribution of squamates in the Indian subcontinent during the Miocene.
Acknowledgments
For English correction and advice, we are indebted to M. Hutchinson (South Australian Museum). For critical reading of the manuscript, we thank M. Augé (Muséum national d'Histoire naturelle, Paris) and G. Georgalis (Paläontologisches Institut und Museum, University of Zürich). We also thank H.D. Sues (editor). This work was supported by SERB-DST (HRR/2018/000063) and Central University of Punjab Bathinda for the Research Seed Money Grant (Ref. no: CUPB/CC/16/00/13). AČ acknowledges financial support from the Scientific Grant Agency of Ministry of Education of Slovak Republic and Slovak Academy of Sciences, Grant Nr. 1/0191/21. NPS and RKS also thank the Director, Wadia Institute of Himalayan Geology, Dehradun, for the research and facilities.



 Open access
Open access