
45 results
The Ethiopian wolf can act as a flagship and umbrella species to protect the Afroalpine ecosystem and foster sustainable development
-
- Journal:
- Environmental Conservation / Volume 51 / Issue 1 / March 2024
- Published online by Cambridge University Press:
- 13 November 2023, pp. 45-54
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Contents
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp iii-iii
-
- Chapter
- Export citation
Introduction to the series
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp viii-x
-
- Chapter
- Export citation
Frontmatter
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp i-ii
-
- Chapter
- Export citation
Notes on sources: The sources - introduction
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp 2-25
-
- Chapter
- Export citation
Symbols Used In Transcription
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp xiii-xiii
-
- Chapter
- Export citation
The Churches
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp 41-318
-
- Chapter
- Export citation
General Introduction
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp 1-1
-
- Chapter
- Export citation
Bedfordshire churches 1550-1914 - a general survey
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp xl-40
-
- Chapter
- Export citation
Miscellaneous Frontmatter
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp iv-v
-
- Chapter
- Export citation
Preface and acknowledgments
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp x-xi
-
- Chapter
- Export citation
List of Illustrations
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp vi-vii
-
- Chapter
- Export citation

Bedfordshire Churches in the Nineteenth Century
-
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023
Bibliography
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp xiv-xv
-
- Chapter
- Export citation
Abbreviations and symbols used in transcription
-
- Book:
- Bedfordshire Churches in the Nineteenth Century
- Published by:
- Boydell & Brewer
- Published online:
- 05 August 2023, pp xii-xiii
-
- Chapter
- Export citation
19 - Colobine Population Ecology
-
-
- Book:
- The Colobines
- Published online:
- 08 February 2022
- Print publication:
- 24 February 2022, pp 312-323
-
- Chapter
- Export citation
Does the presence of elephant dung create hotspots of growth for existing seedlings?
-
- Journal:
- Journal of Tropical Ecology / Volume 35 / Issue 3 / May 2019
- Published online by Cambridge University Press:
- 20 March 2019, pp. 132-139
-
- Article
- Export citation
Spatial patterns of primary seed dispersal and adult tree distributions: Genipa americana dispersed by Cebus capucinus – CORRIGENDUM
-
- Journal:
- Journal of Tropical Ecology / Volume 32 / Issue 1 / January 2016
- Published online by Cambridge University Press:
- 27 October 2015, p. 88
-
- Article
-
- You have access
- HTML
- Export citation
-
Within the second paragraph of page 494 incorrect language was used to characterize the summary characteristics used. Sentences 3–11 of this paragraph should have read:
Second, we calculated three univariate summary characteristics: the nearest neighbour distribution function D(r), the pair-correlation function g(r) and the K-function K(r). The use of multiple summary characteristics holds increased power to characterize variation in spatial patterns (Wiegand et al. 2013). The univariate nearest neighbour distribution function D(r) can be interpreted as the probability that the typical adult tree has its nearest neighbouring adult tree within radius r (or alternatively, the probability that the typical defecation has its nearest neighbouring defecation within radius r). The univariate pair-correlation function g(r) is a non-cumulative normalized neighbourhood density function that gives the expected number of points within rings of radius r and width w centred on a typical point, divided by the mean density of points λ in the study region (Wiegand et al. 2009). We applied g(r) to trees and defecation point patterns separately, using a ring width of 10 m. The K-function K(r) provides a cumulative counterpart to the non-cumulative pair-correlation function g(r) by analysing dispersion and aggregation up to distance r rather than at distance r (Weigand & Moloney 2004). The K-function can be defined as the number of expected points (i.e. either trees or defecations) within circles of radius r extending from a typical point, divided by the mean density of points λ within the study region. Here, we apply the square root transformation L(r) to the K-function to remove scale dependence and stabilize the variance:
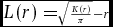 $L( r ) = \scriptstyle\sqrt {\frac{{K( r )}}{\pi }} - r$ (Besag 1977, Wiegand & Moloney 2014).
$L( r ) = \scriptstyle\sqrt {\frac{{K( r )}}{\pi }} - r$ (Besag 1977, Wiegand & Moloney 2014).
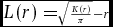
Spatial patterns of primary seed dispersal and adult tree distributions: Genipa americana dispersed by Cebus capucinus
-
- Journal:
- Journal of Tropical Ecology / Volume 31 / Issue 6 / November 2015
- Published online by Cambridge University Press:
- 28 August 2015, pp. 491-498
-
- Article
- Export citation
