The “Mystery of mysteries” is easy to solve for organisms with cultural transmission
8 August 2023
Last update: 08/08/23 12:01
In support of the article How Subcultures Emerge by Petr Tureček, Michal Kozák and Jakub Slavík.
We all know and love discrete models of cultural evolution. We like to discuss the frequencies of hunting strategies, count words in literature across centuries, categorize pottery patterns, and analyze events in fairy tales… and we construct models to fit that kind of data.
It wasn’t always like this. Cavalli-Sforza and Feldman’s foundational book, “Cultural Transmission and Evolution: A Quantitative Approach,” which remains as frequently read as it is cited, contains suggestions for models of both discontinuous and continuous cultural traits.
We find there, among other things, the Galton-Pearson inheritance model. This concept, inspired by the pre-Mendelian biometric theories, proposes that each parent is characterized by a trait value — a position on a single dimension, be it skin color or body height. The distribution of offspring is characterized by the parental mean and a constant standard deviation, eta. The parental variability on the given trait doesn’t matter; a diverse pair of parents is expected to have the same distribution of offspring as a homogeneous pair, provided their mean values are equal.
So, what happened to biometric theories? The ingenious mind of Ronald Fisher envisioned a system that bridged genetic inheritance with the inheritance of continuous traits. Accepting a few assumptions, such as polygenic inheritance and incomplete dominance, we can break up the uninformative constant of the Galton-Pearson model into a few other constant parameters. Eta can be calculated from the overall population variance after selection and mutation frequency.
Cavalli-Sforza and Feldman rightly dismissed Fisher’s formula. There’s no reason to assume that cultural determinants are stored in diploid sets that undergo recombination. However, they didn’t go far enough. Francis Galton, famous empiricist, had measured his way to this conclusion, but he didn’t work with socially transmitted traits. He examined sweet pea seed size and human height. As we’ve argued in a previous paper [1], Cavalli-Sforza and Feldman should have discarded the assumption of constant offspring variance entirely.
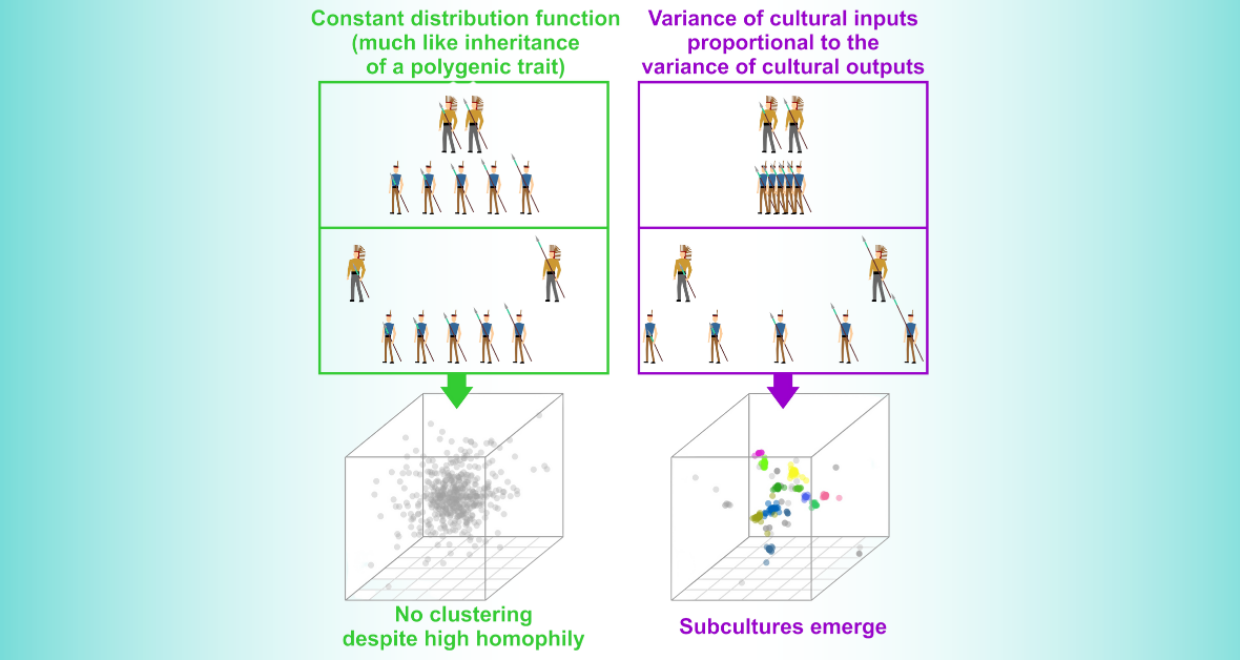
The model they revived assumes that apprentices of two master archers crafting arrows of identical length will be as variable as apprentices of two role models using very different arrows. Isn’t that a bit absurd? If non-sentient replicators like alleles manage inheritance for us, this assumption makes sense, but in social learning, we use all the information we can get; the average input should influence the outcome as much as the input range. Hence, we formulated a comparatively simple yet slightly more fitting algorithm of cultural transmission: the Parental Variability-Dependent Inheritance (or PVDI for short). It postulates the proportionality of the standard deviation of cultural “parents” and “offspring”. In this model, homogeneous inputs lead to homogeneous outputs, and heterogeneous inputs to heterogeneous outputs.
When we dig deep into the historical literature, we find that Francis Galton probably considered similar model himself. He just realized that it does not apply in the inheritance of human body height, so he abandoned it.
In a recent paper published in Evolutionary Human Sciences, we’ve merged these two models. This amalgamation enabled us to generate a wide range of transitive cases that we then applied to simulated cultural exchange in a 10-dimensional culture space. At each time step, every simulated agent chose an inspirational source — another agent — based on a homophily function, which determined preference for self-similar or dissimilar cultural parents.
Our research demonstrated that an increased emphasis on variability-dependence leads to the formation of distinct clusters in the culture space, separated by gaps. We believe these groups — characterized by large between-group variance and small within-group variance — equate to urban subcultures that often emerge in sympatry. Any other ingredient that was necessary for the formation of subcultures was positive assortment.
Previous works reaching similar conclusions posited that the distance between agents in the model influenced imitation probability. If two agents were close — below a certain threshold — they were more likely to harmonize their opinions, and vice versa. This assumption is unnecessary in our model. Once an agent is selected, the inspiration process remains the same, regardless of the distance between them.
This is intriguing because positive assortment seems almost ubiquitous in nature, and it’s even difficult to avoid. All that’s required is something ensuring variability dependence — perhaps culture can serve as a viable candidate.
A study by Nettle and colleagues[2] suggests that eager cultural fragmentation is a baseline tendency counterbalanced by the need to coordinate actions in larger groups. If the growth season is long and stable, there are many distinct languages. But if the season is unstable or short, fostering friendly relationships with more individuals is beneficial, so spontaneously emerging cliques are tirelessly glued together. Without this need, the automatic fragmentation facilitated by PVDI and positive cultural assortment can run wild.
The much-debated “origin of species by means of sympatric speciation” was no mystery or paradox for Darwin. For him, and many of his followers, a species is simply a variety plus time. Thus, the problem of species’ origin is merely the emergence of structure in the continuum of life forms. And culture can help with that. For example, Killer Whales give rise to many different ecologically specialized clans. Songbirds, known for their dialects, continue speciating due to these cultural rather than geographical boundaries – or at least scientists keep writing about this – and songbirds account for half of all bird species diversity.
Another taxon known for its capacity for culture is hominins. Studies show that our relatives and ancestors speciate more than would be expected for a mammal of this size. Perhaps our consistent tendency to form and maintain cohesive in-groups is a natural result of Variability-dependent information transmission and the inevitable inclination to interact with self-similar individuals.

[1] Tureček, P., Slavík, J., Kozák, M., & Havlíček, J. (2019). Non-particulate inheritance revisited: evolution in systems with Parental Variability-Dependent Inheritance. Biological Journal of the Linnean Society, 127(2), 518-533.
[2] Nettle, D. (1998). Explaining global patterns of language diversity. Journal of anthropological archaeology, 17(4), 354-374.
Related Posts

Where Physical and Digital Worlds Collide
In this blog for Data-Centric Engineering, Paul Clarke (Chief Technology Officer at Ocado) documents Ocado’s journey with building synthetic models of its business, its platforms and its underlying technologies, including the use of simulations, emulations, visualisations and digital twins. He explores the potential benefits of digital twins, including the opportunities for creating digital twins at […]

International Women’s Day 2020: Influential women in STEM
International Women’s Day 2020 falls on Sunday, 8th March this year. In the run up to this date, each week day we’ll be highlighting one woman whose accomplishments in science, technology, engineering and/or mathematics not only elevated their fields but also took us one step closer to a gender-equal world. We hope you’ll join us […]

Elephants, Kings, and Comparison, a Conversation with Tom Trautmann
We invited Trautmann to tell us why he writes about Indian war elephants, and how. He touched on several topics in the process: the value of analytical accidents, the recovery of lost knowledge systems, and the power and pluralism of human logics.

Wisconsin’s Response to the Coronavirus Pandemic – Now updated
From time to time, until the crisis has passed, the HEPL blog series authors will be given the opportunity to provide short updates on their country/region’s continuing response to this worldwide catastrophe and their further reflections on those responses. Each update will be labelled accordingly with the original response at the bottom of each post. […]


