Introduction
King penguins (Aptenodytes patagonicus Miller) are major consumers in the Southern Ocean (Guinet et al. Reference Guinet, Cherel, Ridoux and Jouventin1996). During the breeding season, they mostly eat myctophid fish that are caught in deep waters, mainly in the Polar Frontal Zone (South Georgia: Scheffer et al. (Reference Scheffer, Bost and Trathan2012), Marion: Pistorius et al. (Reference Pistorius, Hindell, Crawford, Makhado, Dyer and Reisinger2017), Crozet: Bost et al. (Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015)). Numbers of king penguins have increased throughout the Southern Ocean over the past 50 years (Weimerskirch et al. 1989, Reference Weimerskirch, Stahl and Jouventin1992, Sanders Reference Sanders2006, Crawford et al. Reference Crawford, Whittington, Upfold, Ryan, Petersen, Dyer and Cooper2009). This increase has been interpreted as a recovery from historical exploitation in the 19th century (Rounsewell & Copson Reference Rounsewell and Copson1982, Weimerskirch et al. Reference Weimerskirch, Stahl and Jouventin1992) or change in the functioning of trophic food webs (Guinet et al. Reference Guinet, Cherel, Ridoux and Jouventin1996). However, their numbers may fluctuate extensively in response to large-scale climatic events such as the Sub-Tropical Indian Dipole and El-Niño Southern Oscillation (ENSO; Le Bohec et al. Reference Le Bohec, Durant, Gauthier-Clerc, Stenseth, Park, Pradel, Grémillet, Gendner and LeMaho2008, Bost et al. Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015). Until now, the Morne du Tamaris Colony, Ile aux Cochons, Iles Crozet, in the southern Indian Ocean, has been known as the largest king penguin colony (Guinet et al. Reference Guinet, Jouventin and Malacamp1995). This colony, discovered and photographed by a cartographic team in 1962, was estimated from surface measurements of the colony and breeding densities to contain more than 300 000 pairs of king penguins (Bauer Reference Bauer1963) (Table I). Later estimates from satellite images indicated that it had increased in the surface area occupied, and hosted c. 500 000 breeding pairs in 1982–1988 (Guinet et al. Reference Guinet, Jouventin and Malacamp1995). The Morne du Tamaris colony accounted for a significant proportion of the total consumption by the seabird community in the Indian Ocean at this time (Guinet et al. Reference Guinet, Cherel, Ridoux and Jouventin1996) and was considered to be the second largest penguin colony in the world (Convey et al. Reference Convey, Morton and Poncet1999). The monitoring of this immense colony has become a priority for monitoring programmes in the Southern Ocean, especially since the 1997 El-Niño event negatively affected the foraging capacities, and as a result the survival and growth rates, of king penguins on nearby Ile de la Possession, 100 km to the east of Ile aux Cochons (Bost et al. Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015).
Table I Estimates of colony surface area (bare surfaces), surfaces occupied by breeding king penguins, and number of breeding pairs based on 1.6 incubating individuals per square metre (Bauer Reference Bauer1967), based on oblique photographs taken from a helicopter and satellite imagery, for the Ile aux Cochons colony of Morne du Tamaris.
This paper reports on changes in size over the past 50 years at Ile aux Cochons. Satellite imagery and recent photographs taken from helicopters were used to estimate the surface area occupied by the colony and to infer changes in the population size.
Materials and methods
Ile aux Cochons (67 km2) is one of the three larger islands of the Crozet archipelago located at 46.1°S, 50.2°E. It is a remote island that is rarely visited. The large king penguin colony is located c. 1.5 km from the eastern shore of the island on the border of an ancient volcanic cone, the Morne du Tamaris (144 m, Fig. 1).
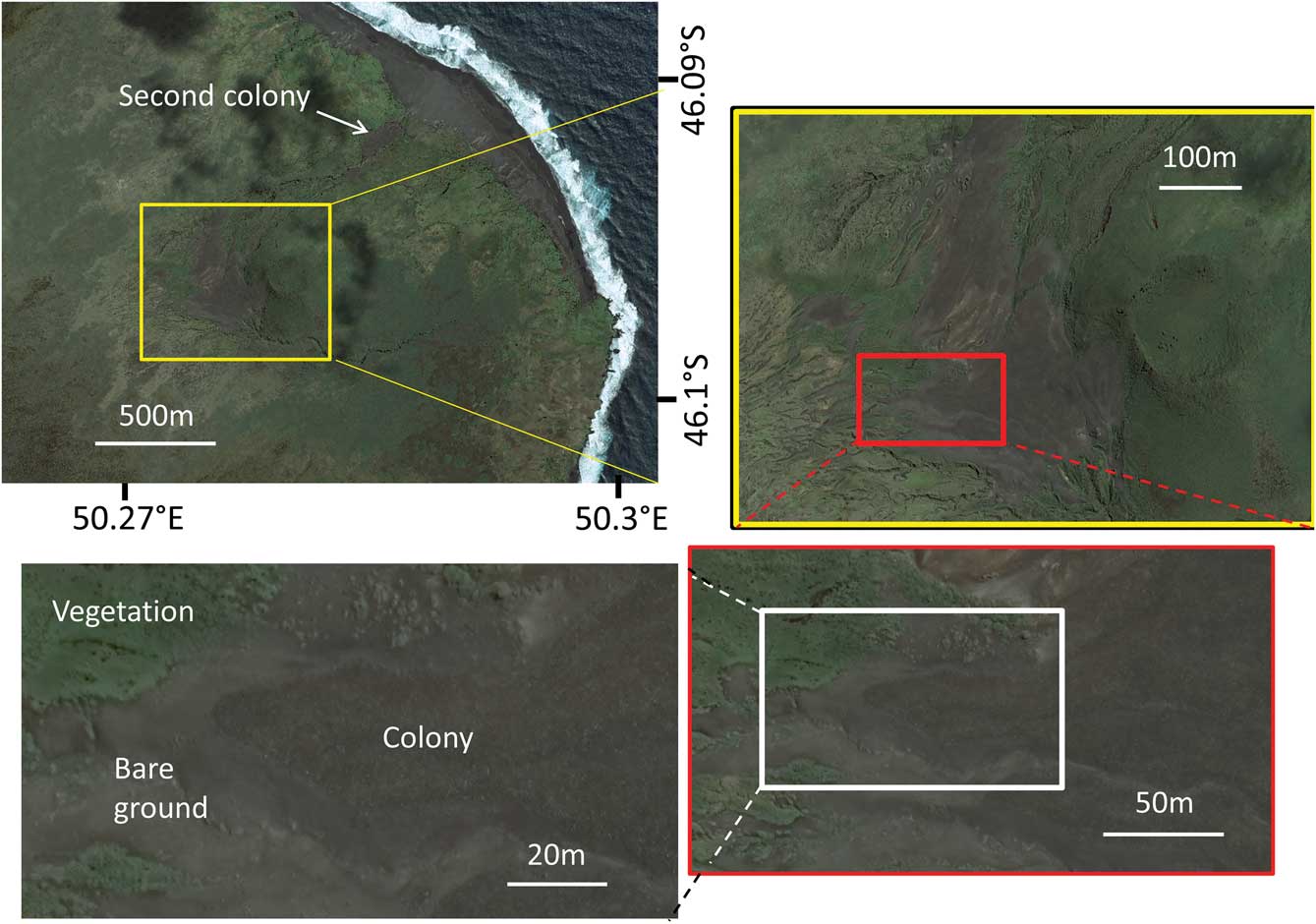
Fig. 1 Satellite images of the Morne du Tamaris king penguin colony at different scales.
King penguins breed in large colonies on flat or gently sloping, unvegetated ground, where they are dispersed regularly, at densities of 1.6–2.2 incubating birds per square metre (Bauer Reference Bauer1967, Barrat Reference Barrat1976). Densities of incubating penguins increase only slightly when the number of incubating birds in a colony increases, as king penguins do not build nests, and disperse, slightly more than one flipper length apart (Barrat Reference Barrat1976). Laying starts in mid-November, with peak numbers of breeding birds being observed in January (Weimerskirch et al. Reference Weimerskirch, Stahl and Jouventin1992, Descamp et al. Reference Descamp, Gauthier-Clerc, Gendner and Le Maho2002).
Published (Bauer Reference Bauer1967, Guinet et al. Reference Guinet, Jouventin and Malacamp1995) and more recent oblique photographs taken from helicopters, and satellite images, were compiled (Table I). In addition to helicopter flights over the Morne du Tamaris colony in 1962 and 1982, flights were carried out from the RS Marion Dufresne in December 1999 and from the RS Akademik Tryoshnikov on 30 December 2016 during the Antarctic Circumpolar Expedition (ACE; PGR, FL). In addition, high-resolution multispectral satellite images taken in January 2015 and April 2017 from the WorldView-3 VHR satellite were used, with the visible bands (2/3/5) pan-sharpened to provide 31-cm resolution colour images from Digital Globe (https://www.digitalglobe.com) (Table I).
From satellite images it can be seen that breeding ‘patches’ occur in bare areas, clearly distinguishable from surrounding vegetated areas (Fig. 1). The surface area of the bare ground can be used to estimate the surface area occupied by the colony, provided that breeding birds occupy the entire bare ground (Guinet et al. Reference Guinet, Jouventin and Malacamp1995). However, during recent years, even at peak breeding time in January, nesting birds have not occupied the entire surface area of bare ground (Fig. 1). Thus when possible, with the most recent high-resolution satellite images from 2015 and 2017, both the entire bare surface area and the surface area occupied by breeding penguins were estimated. Surfaces were estimated by importing satellite images into the Quantum GIS geographical information system and outlining the border of the surfaces. The number of breeders was estimated by using the conservative figure of 1.6 pairs per square metre (Bauer Reference Bauer1967).
Results
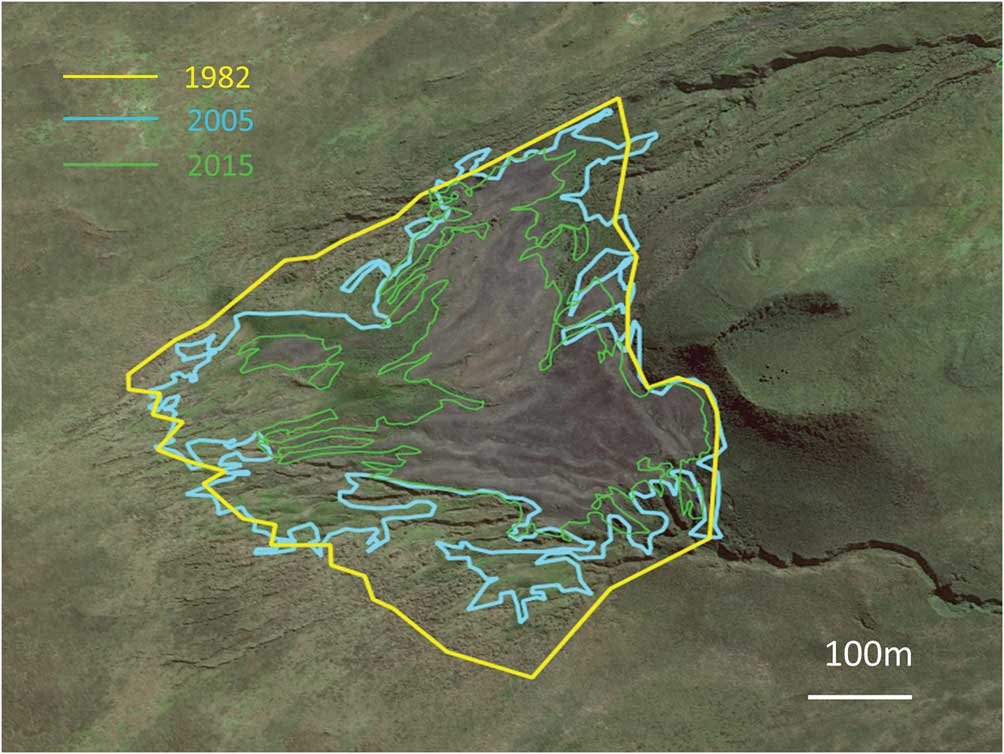
A comparison of estimates of the total surface area occupied by the colony of Morne du Tamaris between 1962 and 2017 indicates that areas of bare and occupied surfaces have shrunk by 64% from their maximum extent in 1982 to a minimum in 2017 (Table I). The total surface area has decreased progressively since 1982, as shown by surface contour lines in Fig. 2. Recent satellite images show that a large section of the former maximum surface has been re-vegetated rapidly over the last 10 years.
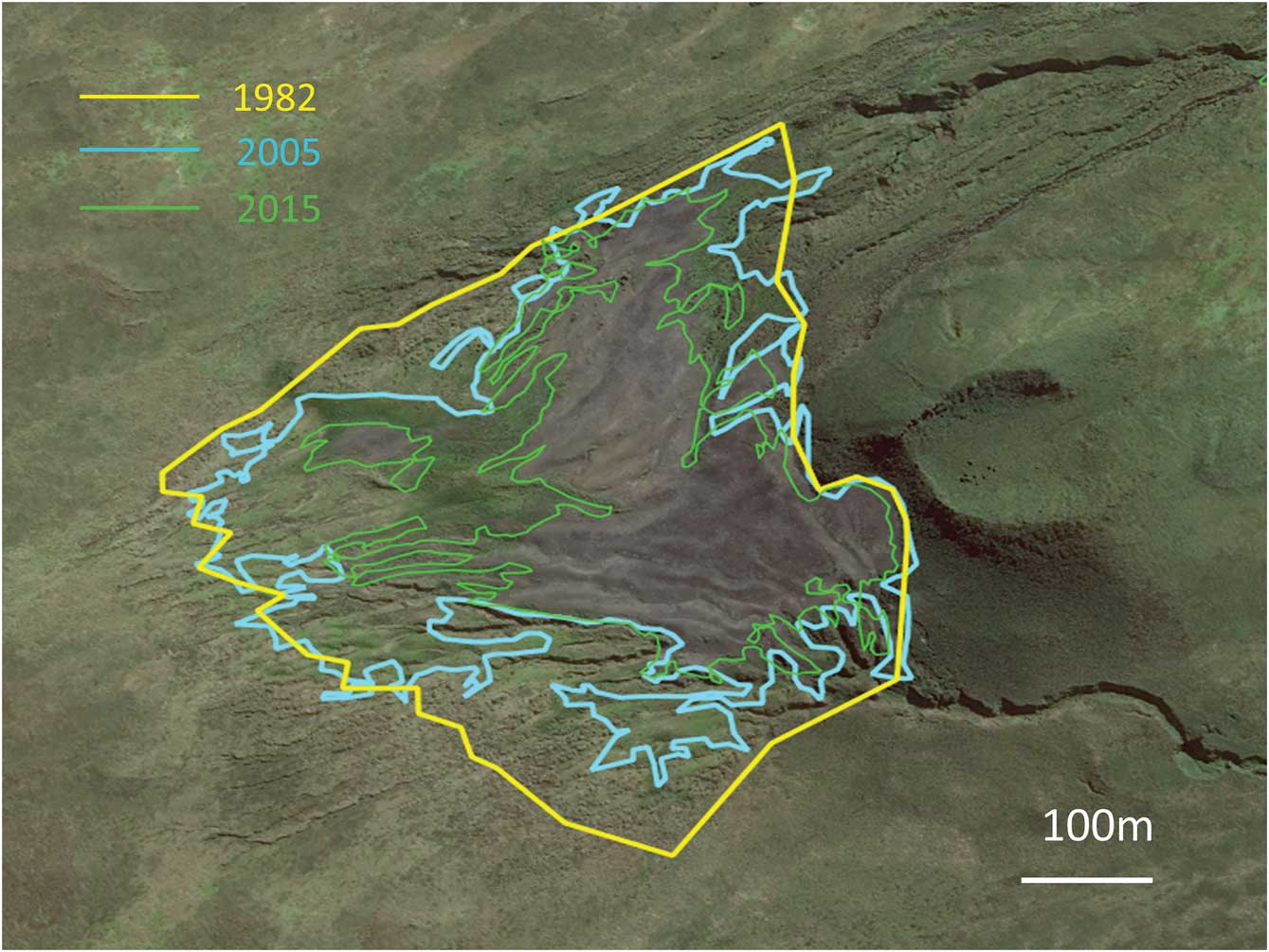
Fig. 2 Contours of the extent of the king penguin colony in 1982 (total bare ground occupied by breeders), 2005 (bare ground) and 2015 (bare ground), applied on the 2015 satellite image.
When peak numbers of incubating birds were present the colony occupied only 40% of the surface available in January 2015, and 32% in April 2017, when adult birds were brooding late chicks, and large chicks were grouped in crèches. Oblique photographs taken by helicopter confirmed that the colony was occupying a reduced area of bare ground. The 2015 data are considered for comparison with historical counts because this was the period of maximum occupancy of the colony, and the images obtained in 2015 clearly distinguish areas of bare ground from the areas occupied by incubating birds (Table I, Fig. 3). The data from April 2017 cannot be used in the comparison with historical counts, as most of the breeders at this time had left their chicks unattended in crèches (Barrat Reference Barrat1976). Thus, considering that the colony fully occupied the bare ground in 1982, as indicated by Guinet et al. (Reference Guinet, Jouventin and Malacamp1995), the population of king penguins declined by 85% between 1988 and 2015, from 494 000 to 76 000 pairs.
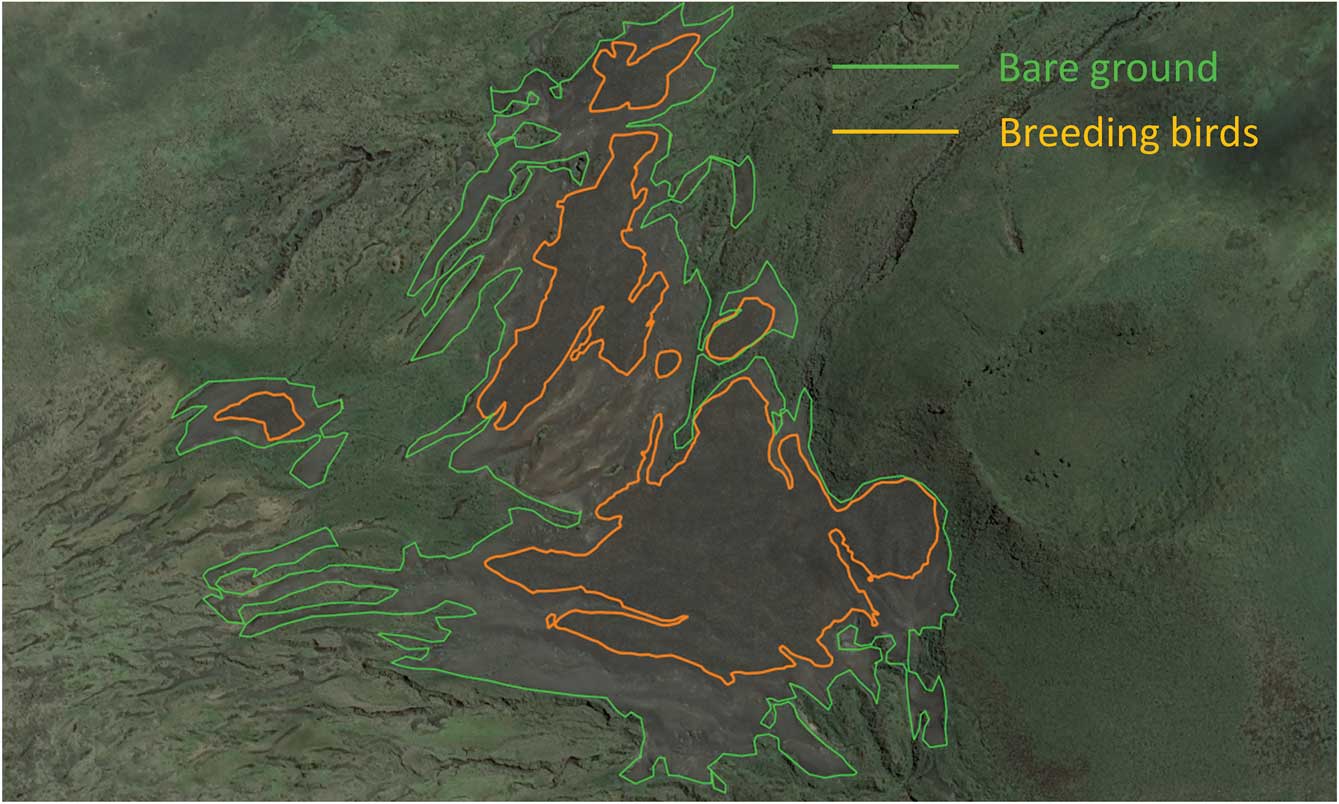
Fig. 3 Colony extent (bare ground, green) and surface area occupied by breeding king penguins (orange) on 24 January 2015.
A second, smaller, colony of king penguins is located on the path used by penguins between the beach where penguins land and the Morne du Tamaris colony (second colony in Fig. 1). This colony was not mentioned in the previous counts. This colony had an estimated 17 500 breeding birds in January 2015 (estimated from surface area occupied by breeding birds), and 14 000 breeding birds in late December 2016 (direct count from aerial photographs), c. one month before the colony attains peak numbers.
Discussion
The analysis of recent aerial photographs and satellite imagery shows that the king penguin colony of Morne du Tamaris on Ile aux Cochons has experienced a massive, unexpected decline, decreasing by 85% over the past 30 years. In the 1980s, the colony was the second largest penguin colony in the world, after the colony of chinstrap penguins (Pygoscelis antarctica Forster) at Zavodovski Island, South Sandwich Islands (Convey et al. Reference Convey, Morton and Poncet1999, Boersma & Borboroglu Reference Boersma and Borboroglu2013).
The 1962 estimate of slightly more than 317 000 pairs was made from vertical photographs taken from a helicopter, and is considered to be fairly accurate (Bauer Reference Bauer1963). The estimate from the Spot satellite image in 1988 was based on the surface area of the colony, and assumed that breeders occupied all the bare ground at the colony site, which was suggested by helicopter photographs in 1982 (Guinet et al. Reference Guinet, Jouventin and Malacamp1995). As it is not known for certain that the surface was fully occupied, the 494 000 breeders in 1988 may be an overestimate, although the bare surface had definitely increased between 1962 and 1982, suggesting an extension of the colony over this time. A low-quality lateral photograph taken in December 1998 suggests that the colony was not at its full extent at this time. The 2005 and especially the 2015 satellite images taken in January were of high quality and allowed the delineation of the bare surface area and of the area occupied by breeders. It showed that in 2005 and 2015, the full extent of the surface occupied by breeders was much smaller than the area of bare surface, associated with the retraction of the surface occupied by breeding birds.
The use of satellite images to monitor penguins is becoming increasingly popular (Guinet et al. Reference Guinet, Jouventin and Malacamp1995, Fretwell et al. Reference Fretwell, LaRue, Morin, Kooyman, Wienecke, Ratcliffe, Fox, Fleming, Porter and Trathan2012, LaRue et al. Reference LaRue, Lynch, Lyver, Barton, Ainley, Pollard, Fraser and Ballard2014) and offers the potential to assess and estimate population change in remote regions such as isolated islands in the Crozet archipelago. Whereas detection of large colonies is now relatively easy, obtaining population estimates remains challenging and requires making a series of assumptions and ground truthing (LaRue et al. Reference LaRue, Lynch, Lyver, Barton, Ainley, Pollard, Fraser and Ballard2014). In contrast to emperor penguins (Aptenodytes forsteri Gray), for which colonies can be detected from guano deposits on the ice (Fretwell & Trathan Reference Fretwell and Trathan2009) and population estimates are difficult, king penguins breed on bare flat ground, at relatively constant densities. The range of king penguin densities (1.6–2.2 incubating birds m-2) (Bauer Reference Bauer1967, Barrat Reference Barrat1976) depends on the ratio of the total number of birds with respect to available bare ground and on the stage of the breeding cycle. In this study the numbers in the period of maximum occupation of the colonies were compared, and a figure of 1.6 individuals m-2 was used for all periods. As a massive decline would be expected to result in a slight decrease in densities over time, using the same minimal densities over time results in a conservative estimate of the extent of decline.
The Morne du Tamaris colony increased between 1962 and 1982 (Guinet et al. Reference Guinet, Jouventin and Malacamp1995), like other king penguin colonies monitored in Iles Crozet (Weimerskirch et al. 1989, Reference Weimerskirch, Stahl and Jouventin1992). The extensive decline between 1982–88 and 2015–17 on Ile aux Cochons was unexpected, because the monitored colonies elsewhere in the Crozet archipelago (Ile de la Possession) remained stable overall with interannual fluctuations over this period (Bost et al. Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015). King penguin colonies at Marion Island (900 km west) and Iles Kerguelen (1200 km east–south-east) have increased since the 1960s (Crawford et al. Reference Crawford, Whittington, Upfold, Ryan, Petersen, Dyer and Cooper2009, Weimerskirch unpublished data), thus the decline of king penguins seems to be a site-specific phenomenon at Ile aux Cochons. The decline appears to be progressive, or at least the bare area occupied by the colony has decreased progressively, with a gradual recovery of the vegetation around the periphery of the colony compared to its maximum extent in 1982.
Several hypotheses might explain the massive decline in this colony. First, the decline occurred from the late 1990s, coinciding with the strong Dipole event in 1997 that affected the foraging capacities of king penguins on Ile de la Possession, the second most important island of Iles Crozet for breeding king penguins, with negative consequences for breeding performance and colony size (Bost et al. Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015). The 1997 event may have affected the population of Ile aux Cochons more severely than other colonies because of stronger density-dependent effects, and the population has not recovered, as has been observed on Ile de la Possession (Delord et al. Reference Delord, Barbraud and Weimerskirch2004, Bost et al. Reference Bost, Cotté, Terray, Barbraud, Bon, Delord, Gimenez, Handrich, Naito, Guinet and Weimerskirch2015).
Second, the decline may have resulted from the partial relocation of the colony. King penguins are relatively faithful to their birthplace and first breeding site (Saraux et al. Reference Saraux, Viblanc, Hanuise, Le Maho and Le Bohec2011) and it seems unlikely that adults and young individuals from Ile aux Cochons have a different behaviour than those from other colonies. However, observations from helicopter flights in 2016 noted a smaller colony nearer to the beach, on the penguins’ access route to the large colony. This colony was recorded for the first time on the 2005 satellite images, and was still present in the 2015, 2016 and 2017 satellite images. This colony was not present in 1962 (Bauer Reference Bauer1963) or during ground observations in 1974 and 1982 (Derenne et al. Reference Derenne, Mougin, Steinberg and Voisin1976, Voisin Reference Voisin1984). It was not mentioned in 1988, but the Spot images focused on the Morne du Tamaris colony (Guinet et al. Reference Guinet, Jouventin and Malacamp1995). However, the size of this new colony (17 000 pairs) can account for the relocation of only a small fraction of the original colony.
Third, feral cats (Felis catus L.) and house mice (Mus musculus L.) are present on Ile aux Cochons (Derenne & Mougin Reference Derenne and Mougin1976, Derenne et al. Reference Derenne, Mougin, Steinberg and Voisin1976). Cats apparently occurred in small numbers in the 1970s (Derenne et al. Reference Derenne, Mougin, Steinberg and Voisin1976) and neither species is known to be a predator of king penguin chicks. However, the behaviour of these two introduced species towards native fauna has changed during recent years on other sub-Antarctic islands, as shown by observations on Gough and Marion islands of mice attacking albatross chicks, causing the decline of at least some populations (Wanless et al. Reference Wanless, Angel, Cuthbert, Hilton and Ryan2007, Davies et al. Reference Davies, Dilley, Bond, Cuthbert and Ryan2015, Dilley et al. Reference Dilley, Schoombie, Schoombie and Ryan2016), or at Kerguelen, where cats have been observed to attack wandering albatross (Diomedea exulans L.) chicks, reducing breeding success (Weimerskirch unpublished data).
Fourth, diseases or parasites can seriously affect seabird populations, reducing breeding success, survival of adults and population growth rates (Weimerskirch Reference Weimerskirch2004, Cooper et al. Reference Cooper, Crawford, De Villiers, Dyer, Hofmeyr and Jonker2009). There are no data available about the occurrence of any such diseases on Ile aux Cochons. Tick infestations also affect seabird populations, and have been found to be a vector of Lyme disease in king penguins (Gauthier-Clerc et al. Reference Gauthier-Clerc, Jauhlac, Frenot, Bachelard, Monteil, Le Maho and Handrich1999), but again a massive tick infestation appears to be unlikely compared to the situation recorded on Ile de la Possession. The extent of the decrease of the colony of Morne du Tamaris would be unprecedented if it were a result of disease outbreak, but previous large mortalities of king penguins (although not to the same extent as in the Ile aux Cochons colony) have been reported in 1992–1993 on Marion Island (Cooper et al. Reference Cooper, Crawford, De Villiers, Dyer, Hofmeyr and Jonker2009), at the time of the rapid decline described here.
Finally, a catastrophic event seems unlikely to have occurred. The colony is well inland and therefore should not be impacted by a tsunami, and there is no evidence of a volcanic eruption on the satellite images or recent helicopter photographs. Furthermore, the progressive decline of the extent of the colony suggests a gradual, long-term decrease in this colony.
In conclusion, the cause of the massive decline of the colony remains a mystery, and needs to be resolved. Although the decline started at least 20 years ago, it appears to be ongoing, and the causes of the decline may still be active. Ile aux Cochons is rarely visited, and the use of satellite images has allowed the detection of this unexpected phenomenon. However, to be able to understand the cause of the decline, it is necessary to study the colony on land and at sea. The last visits ashore were in 1974 and 1982. It would be of considerable importance to examine the foraging ecology (including at-sea distribution) of individuals from this colony to detect potential adverse trophic conditions, and make observations on the conditions that occur on land today, such as the possible presence of diseases and parasites, and the effects of native or introduced predators.
Acknowledgements
This study was funded by the French Polar Institute (Program IPEV 109), by Réserve Nationale des Terres Australes Françaises), and by ACE, a scientific expedition carried out under the auspices of the Swiss Polar Institute, supported by funding from the ACE Foundation and Ferring Pharmaceuticals. We thank the crews and especially the helicopter pilots for their help, Joris Laborie and Adrien Chaigne for help with QGIS and two anonymous referees for comments on the manuscript.
Author contributions
HW oversaw the project, analysed satellite images, and wrote a first draft of the manuscript. PGR and FLB analysed the aerial photographs from the colony during the ACE cruise and estimated the population size from the photographs. All authors contributed to the overall editing and organization of the manuscript.