


1. Introduction
The term ‘Cambrian radiation’ (popularly known as the ‘Cambrian explosion’) is embedded in modern scientific literature and was coined for the early Cambrian geologically rapid diversification of metazoans. Most of these early Cambrian metazoans appear to represent members of the stem lineage of extant clades. The first appearances of these groups are clustered geochronologically by higher-rank phylogeny (Budd, Reference Budd2003; Landing & Westrop, Reference Landing, Westrop, Lipps and Waggoner2004; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007). Budd (Reference Budd2003) and Budd & Jensen (Reference Budd and Jensen2000, Reference Budd, Jensen, Donoghue and Smith2003) argued that this clustering reflects the true sequence of divergence of these high-ranking groups and implies rather late origins, probably near the Precambrian–Cambrian boundary, followed by their rapid evolutionary radiation in the early Cambrian. This event is marked in the fossil record by the first appearances and increase in diversity and abundance of many groups of animals, accompanied by the independent acquisition of mineralized skeletons in many lineages.
Skeletal biomineralization was likely an epiphenomenon of the general radiation of body plans and tissues (Bengtson, Reference Bengtson, Lipps and Waggoner2004). Skeletal elements are considered principal aspects of many body plans, and their origin and diversification are thought to have helped spur evolutionary radiation in the Cambrian. Skeletons certainly diversified along with the taxa that obtained them, with 80% of modern skeletal morphotypes present by the middle Cambrian (Thomas, Sherman & Stewart, Reference Thomas, Sherman and Stewart2000). The diversity of minerals employed in early skeletalized animals suggests a limit to the role of ocean geochemistry in the emergence of skeletons, although the primary acquisition of particular skeletal carbonate mineralogies was likely driven by the ocean geochemistry (Zhuravlev, Reference Zhuravlev1993; Bengtson, Reference Bengtson and Bengtson1994, Reference Bengtson, Lipps and Waggoner2004; Ushatinskaya & Zhuravlev, Reference Ushatinskaya and Zhuravlev1994; Hardie & Stanley, Reference Hardie and Stanley1997; Stanley & Hardie, Reference Stanley and Hardie1998; Porter, Reference Porter2007; Zhuravlev & Wood, Reference Zhuravlev and Wood2008; Kiessling, Aberhan & Villier, Reference Kiessling, Aberhan and Villier2008). Mineralized skeletal parts are only one of many strategies to escape predation, since skeletonized species constitute a minority in modern and ancient marine ecosystems (e.g. Conway Morris, Reference Conway Morris1986). However, it was probably the anti-predatory selective advantage of mineral skeletons that drove early evolution in many clades (Bengtson, Reference Bengtson and Bengtson1994, Reference Bengtson, Lipps and Waggoner2004). Further diversification of predators and their increased pressure on epibenthos in the Cambrian may well have triggered the early Cambrian rapid evolutionary radiation in different clades (Bengtson, Reference Bengtson and Bengtson1994, Reference Bengtson, Lipps and Waggoner2004).
In order to understand in detail the fossil record of early Cambrian skeletal biomineralization and structure of the Cambrian radiation, a better-resolved sequence of first appearances of mineralized skeletons in the early Cambrian class-to-phylum-level animal groups is presented herein. Our study incorporates a new carbon isotope chemostratigraphy of the northern Siberian Platform, where a continuous isotopic and fossil record is known from mainly carbonate sections of the Cambrian System (online Fig. S1 at http://journals.cambridge.org/geo). The Siberian data are stratigraphically correlated with those from other well-known units of the Cambrian world, such as Western Mongolia, Kazakhstan, South China, Iran and India, Australia, Avalonia, the Mediterranean region of West Gondwana, Laurentia and Baltica (Fig. 1; online Appendices 1 & 2 at http://journals.cambridge.org/geo). This record, supported by available chemostratigraphy and radiometric dating, allows us to constrain the timing of the first appearances of skeletonization in various animal groups (Fig. 2).

Figure 1. Schematic palaeogeographic map for the early Cambrian with crustal units discussed in this paper (adapted from Fatka, Kraft & Szabad, Reference Fatka, Kraft and Szabad2011 and Álvaro et al. in press).

Figure 2. Global first appearances of mineralized skeletons in animals during the Cambrian radiation. Question marks indicate uncertainty in first appearance or place within higher-ranked group; affinities of chancelloriids to the Lophotrochozoa and hyolithelminths to the Cnidaria are uncertain (see main text and online Appendix 1 at http://journals.cambridge.org/geo). Cambroclavids (with paracarinachitids included) are tentatively attributed to the Lophotrochozoa. Vertical dashed lines for sponges reflect sporadic occurrence of their presumably biomineralized spicules in the Precambrian (see main text and online Appendix 1 at http://journals.cambridge.org/geo). ECC – Ediacaran calcified cnidariomorphs; Ch – Chaetognatha; D – Deuterostomia.
2. Stratigraphical setting
The traditional three-fold subdivision of the Cambrian System into Lower, Middle and Upper Cambrian series has been abandoned recently in favour of a subdivision into four series of ten stages (Babcock et al. Reference Babcock, Peng, Geyer and Shergold2005; Babcock & Peng, Reference Babcock and Peng2007). The uppermost two series of the revised Cambrian timescale more or less correspond to the traditional Middle and Upper Cambrian series, while the former Lower Cambrian is subdivided into two series. The new Cambrian timescale begins with the Fortunian Stage of the Terreneuvian Series, the base of which is defined as the first occurrence of the trace fossil Treptichnus pedum at Fortune Head in Newfoundland (Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007). It is anticipated that the base of Cambrian Series 2 and Stage 3 will coincide with the first appearance of trilobites (Babcock & Peng, Reference Babcock and Peng2007), here interpreted to correlate with the base of the Atdabanian Stage in Siberia (Fig. 3). The remaining boundaries within Series 1–2 are less certain. In the present work, we will provisionally correlate the base of Cambrian Stage 2 of the Terreneuvian Series with the base of the Watsonella crosbyi Zone of Avalonia and South China (see Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007) and the base of Cambrian Stage 4 with the lower Botoman Stage in Siberia (Fig. 3). The base of Cambrian Series 3, Stage 5 is presently highly debated, but is here correlated with the first occurrence of the trilobite Oryctocephalus indicus, which more or less coincides with the traditional Lower–Middle Cambrian boundary in many areas. The further development of the chronostratigraphy of Cambrian Series 1 and 2 is directly related to our precision in dating and correlation of the first appearances of diverse skeletal fossils in principal Cambrian sequences.
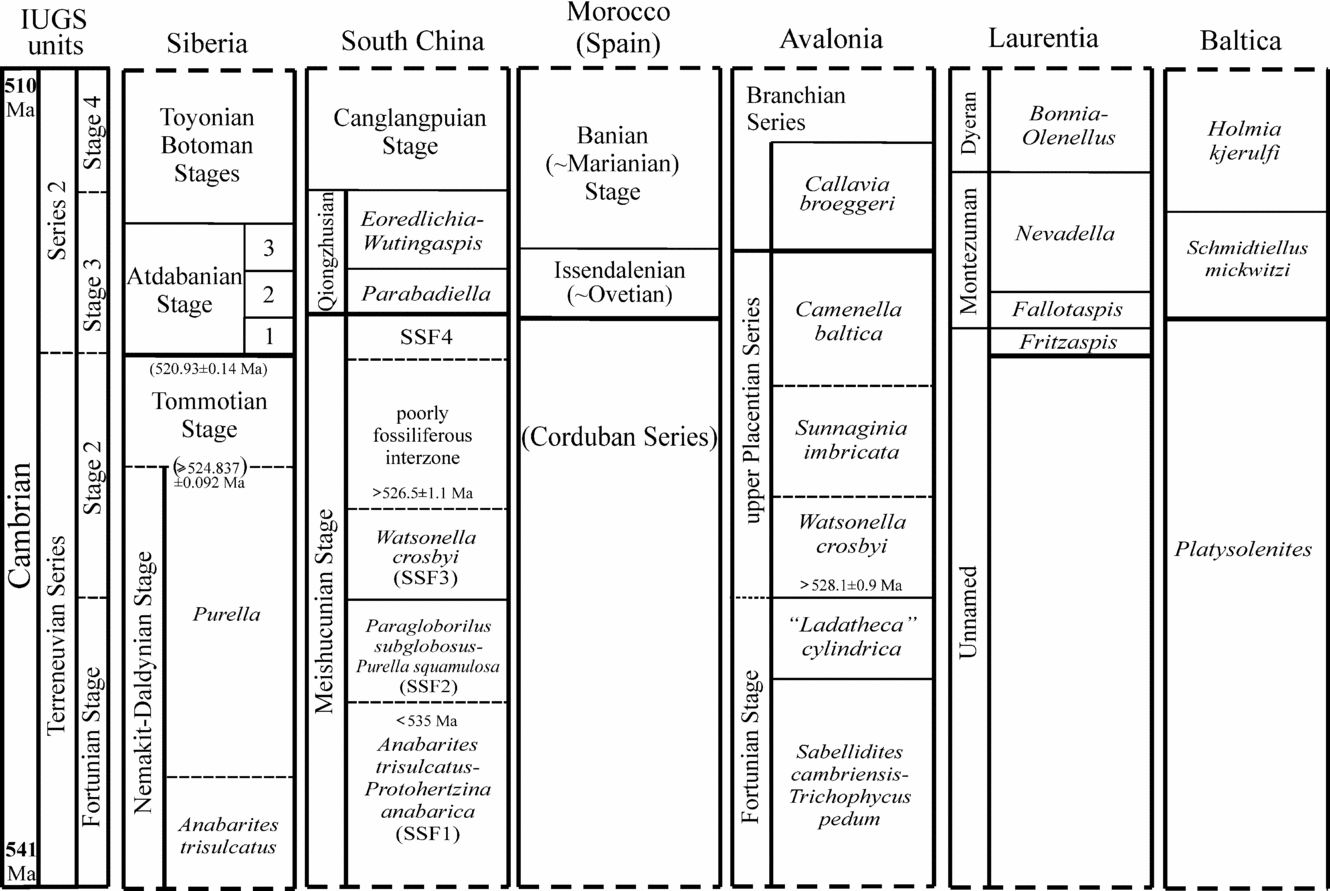
Figure 3. Correlation chart of the major crustal units discussed in this paper. It is formally accepted that the Purella Zone corresponds to the entire upper part of the Nemakit–Daldynian Stage (e.g. Rozanov et al. Reference Rozanov, Khomentovsky, Shabanov, Karlova, Varlamov, Luchinina, Pegel’, Demidenko, Parkhaev, Korovnikov and Skorlotova2008). According to the chemostratigraphic record (e.g. Kaufman et al. Reference Kaufman, Knoll, Semikhatov, Grotzinger, Jacobsen and Adams1996; see also Fig. 4 herein), a hiatus of disputed duration exists, however, at the base of the Tommotian Stage in its stratotype in the southeastern Siberian Platform. The Purella Zone is shown herein to cover its duration, since the missing record exists in the northern part of the Platform. The first occurrences of trilobites are highlighted by thick horizontal lines within the Stage 3 interval. The Atdabanian Stage is subdivided into three parts (lower, middle and upper): 1 – Profallotaspis jakutensis and Repinaella zones; 2 – Delgadella anabara Zone; 3 – Judomia Zone. SSF4 = Sinosachites flabelliformis–Tannuolina zhangwentangi Assemblage Zone. Absolute ages for the Precambrian–Cambrian boundary after Bowring et al. Reference Bowring, Grotzinger, Condon, Ramezani, Newall and Allen2007; Cambrian Series 2–Series 3 boundary after Ogg, Ogg & Gradstein, Reference Ogg, Ogg and Gradstein2008. Absolute ages of the lower and upper boundaries of the Tommotian Stage are based on data from Morocco (see main text and Maloof et al. Reference Maloof, Schrag, Crowley and Bowring2005, Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Bengtson, Conway Morris, Cooper, Jell and Runnegarb). The fossiliferous Watsonella crosbyi Zone in China is older than 526.5 ± 1.1 Ma (after Compston et al. Reference Compston, Zhang, Cooper, Ma and Jenkins2008) and its lower boundary in Avalonia is shown to be older than 530.7 ± 0.9 Ma (Isachsen et al. Reference Isachsen, Bowring, Landing and Samson1994) or 528.1 ± 0.9 Ma (Compston et al. Reference Compston, Zhang, Cooper, Ma and Jenkins2008). That boundary is, however, younger than 535.2 ± 1.7 Ma, a combined depositional age for Bed 5 (shown as < 535 Ma in figure) from the upper Anabarites trisulcatus–Protohertzina anabarica Zone (Zhu et al. Reference Zhu, Li, Hou, Pan, Wang, Deng and He2009; see main text for discussion).
The age of the Precambrian–Cambrian boundary, marked worldwide by a negative carbon isotope anomaly, is dated in the well-constrained section in Oman to 542 ± 0.3 Ma (Amthor et al. Reference Amthor, Grotzinger, Schröder, Bowring, Ramezani, Martin and Matter2003), later revised to 541 ± 0.13 Ma (Bowring et al. Reference Bowring, Grotzinger, Condon, Ramezani, Newall and Allen2007). An associated biotic crisis is inferred from the disappearance of the Ediacaran biota (e.g. Kimura & Watanabe, Reference Kimura and Watanabe2001; Narbonne, Reference Narbonne2005). As recognized herein, the Cambrian radiation event (‘Cambrian explosion’) occurred within a period of c. 25 Ma. The Cambrian radiation began with the diversification of skeletonized bilaterians following this negative anomaly and concluded with the Botoman–Toyonian biotic crisis (Zhuravlev & Wood, Reference Zhuravlev and Wood1996; Zhuravlev, Reference Zhuravlev, Zhuravlev and Riding2001; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007), which was the first mass extinction episode in the Phanerozoic (Signor, Reference Signor, Hall, Doyle-Jones and Widawski1992). The onset of the crisis on the Siberian Platform (Sinsk event), is marked by decreasing δ13C values above peak VII of the Siberian reference scale (Brasier et al. Reference Brasier, Corfield, Derry, Rozanov and Zhuravlev1994a, Reference Brasier, Rozanov, Zhuravlev, Corfield and Derryb; Zhuravlev & Wood, Reference Zhuravlev and Wood1996; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007; Fig. 4).

Figure 4. First appearances of skeletal fossil groups on the Siberian Platform (S1–S13, see Appendix 1 for details) in the key sections correlated with the δ13C chemostratigraphy (after Brasier, Khomentovsky & Corfield, Reference Brasier, Khomentovsky and Corfield1993; Brasier et al. Reference Brasier, Rozanov, Zhuravlev, Corfield and Derry1994b; Kouchinsky et al. Reference Kouchinsky, Bengtson, Missarzhevsky, Pelechaty, Torssander and Val'kov2001). Zones: Ns – Nochoroicyathus sunnaginicus; Dr – Dokidocyathus regularis; Dl – Dokidocyathus lenaicus–Tumuliolynthus primigenius; Rz – Retecoscinus zegebarti; C. pinus – Carinacyathus pinus; Nk – Nochoroicyathus kokoulini; Fl – Fansycyathus lermontovae; Pj – Profallotaspis jakutensis; R – Repinaella; D. anabara – Delgadella anabara; Bm – Bergeroniellus micmacciformis; Bg – Bergeroniellus gurarii; Ba – Bergeroniellus asiaticus. Numbers in brackets are projections of the corresponding first appearances in the Atdabanian Stage of the Anabar Uplift onto the Lena-Aldan reference scale.
The Cambrian radiation interval shows prominent and frequent oscillations of the carbon isotope ratio (e.g. Brasier et al. Reference Brasier, Corfield, Derry, Rozanov and Zhuravlev1994a, Reference Brasier, Rozanov, Zhuravlev, Corfield and Derryb; Brasier & Sukhov, Reference Brasier and Sukhov1998; Figs 4, 5). An overall rising trend in δ13C values characterizes the sedimentary sequence of the Fortunian Stage in Siberia, Mongolia, China and Western Gondwana. SHRIMP U–Pb zircon analyses from the lower part of this trend in South China provide a revised age of 539.4 ± 2.9 Ma (Compston et al. Reference Compston, Zhang, Cooper, Ma and Jenkins2008), whereas secondary ion mass spectrometry (SIMS) of the same tuffite (Bed 5 of the Meishucun section) resulted in a c. 533 Ma estimate (Brooks et al. Reference Brooks, Crowley, Bowring, Cervato and Jin2006). Nano-SIMS measurements by Sawaki et al. (Reference Sawaki, Nishizawa, Suo, Komiya, Hirata, Takahata, Sano, Han, Kon and Maruyama2008) provided an age estimate of 536.5 ± 2.5 Ma for Bed 5, whereas SIMS analyses by Zhu et al. (Reference Zhu, Li, Hou, Pan, Wang, Deng and He2009) yielded an age of 536.7 ± 3.9 Ma. A combined depositional age for Bed 5 was calculated as 535.2 ± 1.7 Ma by Zhu et al. (Reference Zhu, Li, Hou, Pan, Wang, Deng and He2009). The fauna known from below Bed 5 is considered herein to be older than 535 Ma (Figs 2, 3). The dated bed is situated in the upper part of the Anabarites trisulcatus–Protohertzina anabarica (SSF1) Assemblage Zone of the lower Meishucunian Stage.

Figure 5. First appearances of skeletal fossil groups in the Tsagan Oloom and Bayan Gol formations of Western Mongolia (M1–M7, see Appendix 2 for details) (after Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 6), in the Tsagan Gol, Bayan Gol and Salany Gol sections composite. Features of the δ13C curve, from W to C after Brasier et al. (Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 5) and from D to F after fig. 7 therein; adapted for unit thickness of the Bayan Gol Formation in Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 6a. Zones after Brasier et al. (Reference Brasier, Shields, Kuleshov and Zhegallo1996). A – correlation with Siberia, after Voronin et al. (Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982), Astashkin et al. (Reference Astashkin, Pegel’, Repina, Belyaeva, Esakova, Rozanov, Zhuravlev, Osadchaya and Pakhomov1995), Khomentovsky & Gibsher (Reference Khomentovsky and Gibsher1996) and Esakova & Zhegallo (Reference Esakova, Zhegallo and Rozanov1996); B – chemostratigraphic correlation, after Brasier et al. (Reference Brasier, Shields, Kuleshov and Zhegallo1996).
The rising trend of δ13C values continues into the Cambrian Stage 2, where several highly positive peaks preceding the Tommotian Stage in Siberia top it (Fig. 4). A single preserved prominent positive peak from the same rising trend is present in the Watsonella crosbyi (SSF3) Assemblage Zone of the Middle Meishucunian Stage in South China and is older than 526.5 ± 1.1 Ma (after Compston et al. Reference Compston, Zhang, Cooper, Ma and Jenkins2008). The lower boundary of the Cambrian Stage 2 and Watsonella crosbyi Zone has been preliminarily estimated to be close to 530 Ma (Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007). The estimate is based on the absolute age of 530.7 ± 0.9 Ma obtained from the Watsonella crosbyi Zone in Avalonia (Isachsen et al. Reference Isachsen, Bowring, Landing and Samson1994), but the date was revised to 528.1 ± 0.9 Ma by Compston et al. (Reference Compston, Zhang, Cooper, Ma and Jenkins2008, p. 417). The uppermost peak in the rising trend of δ13C values within Stage 2 is dated in Morocco to 525.4 ± 0.5 Ma (Maloof et al. Reference Maloof, Schrag, Crowley and Bowring2005), later revised to 525.343 ± 0.088 Ma (Maloof et al. Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Maloof, Ramezani, Bowring, Fike, Porter and Mazouadb). The high-resolution successions of δ13C peaks in Morocco and Siberia (Brasier et al. Reference Brasier, Rozanov, Zhuravlev, Corfield and Derry1994b; Kouchinsky et al. Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007) proved to be very similar, and their correlation resulted in a c. 525 Ma estimate of the age of the Nemakit–Daldynian–Tommotian boundary dated in Morocco to 524.837 ± 0.092 Ma (Maloof et al. Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Maloof, Ramezani, Bowring, Fike, Porter and Mazouadb). Chemostratigraphic δ13C correlation with radiometrically dated sections of Morocco allows dating of the lower boundary of the Atdabanian Stage in Siberia to a maximum age of 520.93 ± 0.14 Ma (Maloof et al. Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Maloof, Ramezani, Bowring, Fike, Porter and Mazouadb), thereby providing an estimate of c. 521 Ma for the upper boundary of Stage 2 and the Terreneuvian Series.
A recalculated age of 515.56 ± 1.16 Ma attributed to the upper Antatlasia gutta-pluviae Zone of the Moroccan Banian Stage (originally 517.0 ± 1.5 Ma by Landing et al. Reference Landing, Bowring, Davidek, Westrop, Geyer and Heldmaier1998) can be correlated with the Botoman Stage (probably, the Bergenellious asiaticus Zone) of Siberia and the Bonnia–Olenellus Zone of Laurentia (Zhuravlev, Reference Zhuravlev1995; Landing et al. Reference Landing, Bowring, Davidek, Westrop, Geyer and Heldmaier1998; Maloof et al. Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a). The conclusion of the Cambrian radiation and the onset of the Botoman–Toyonian biotic crisis are, therefore, dated herein to be c. 515 Ma (Figs 2, 3). The upper boundary of the Cambrian Series 2 is estimated to be c. 510 Ma (Ogg, Ogg & Gradstein, Reference Ogg, Ogg and Gradstein2008), because it is somewhat younger than the estimated age of 511 ± 1 Ma for the upper Branchian Series of Avalonia (Landing et al. Reference Landing, Bowring, Davidek, Westrop, Geyer and Heldmaier1998).

Figure 6. Map of the Siberian Platform with localities referred to in the main text and online Appendix 1 at http://journals.cambridge.org/geo. The localities are indicated by circles with the following numbers: 1 – Sukharikha River, middle reaches; 2 – Kotuj River (between the Sergej-Koril-Uoran rapids and mouth of the Kugda Brook) and the lowermost reaches of the Kotujkan River; 3 – Bol'shaya Kuonamka River, lower reaches; 4 – Olenyok River at the mouth of the Erkeket River and Khorbosuonka River at the mouth of the Mattajya River; 5 – Lena River, lower reaches, sections Chekurovka and at the mouth of the Ulakhan-Ald'yarkhaj Brook; 6 – Lena River, middle reaches, between sections Isit’ and Achchagyi-Kyyry-Taas; 7 – Aldan River, between sections ‘Dvortsy’ and Ulakhan-Sulugur; 8 – Uchur-Maya region, sections along the Uchur River, between the Gonam and Selinde rivers, including the Mount 1291 m, Mount Konus, Nemnekey, and Selinde localities.
2.a. Siberia
The former Lower Cambrian includes on the Siberian Platform and in the Altai-Sayan Folded Area the Tommotian, Atdabanian, Botoman and Toyonian stages, in ascending order (Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Rozanov et al. Reference Rozanov, Khomentovsky, Shabanov, Karlova, Varlamov, Luchinina, Pegel’, Demidenko, Parkhaev, Korovnikov and Skorlotova2008; Varlamov et al. Reference Varlamov, Rozanov, Khomentovsky, Shabanov, Abaimova, Demidenko, Karlova, Korovnikov, Luchinina, Malakhovskaya, Parkhaev, Pegel, Skorlotova, Sundukov, Sukhov, Fedorov and Kipriyanova2008). The lowermost Cambrian strata (approximately, Fortunian Stage equivalent) were recognized on the Platform as the Nemakit–Daldynian (or Manykayan Stage by some authors, e.g. Missarzhevsky, Reference Missarzhevsky1982, Reference Missarzhevsky1989; Val'kov, Reference Val'kov1982, Reference Val'kov1987; see discussion in Khomentovsky & Karlova, Reference Khomentovsky and Karlova2002, Reference Khomentovsky and Karlova2005). The Nemakit–Daldynian Stage is now subdivided into the Anabarites trisulcatus and Purella antiqua zones, in ascending order (Khomentovsky & Karlova, Reference Khomentovsky and Karlova1993, Reference Khomentovsky and Karlova2002). In the Lena-Aldan and Uchur-Maya regions of the southeastern Siberian Platform, the first appearances of taxa are reported from the Ust’-Yudoma Formation (Nemakit–Daldynian Stage) and from the overlying Pestrotsvet Formation (Tommotian–Atdabanian stages) (Figs 4, 6). In the northern part of the Platform, the regional first occurrences are recorded from the Manykay, Medvezh'ya and Emyaksin formations of the Anabar Uplift (Nemakit–Daldynian–Atdabanian stages), upper Turkut and Kessyuse formations of the Olenyok Uplift (Nemakit–Daldynian–lower Tommotian), and uppermost Sukharikha–lower Krasny Porog formations abutting the lower Tommotian boundary on the northwestern margin of the Platform (Figs 4, 6). Detailed biostratigraphy and carbon isotope chemostratigraphy was obtained throughout these units and provides the basis for global correlations used herein (Fig. 4).
2.b. Western Mongolia
Ediacaran and lower Cambrian beds in Mongolia are better studied in sections of the Khasagt-Khairkhan Ridge in Western Mongolia, where carbon isotope chemostratigraphy and sequence stratigraphy are also available (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Esakova & Zhegallo, Reference Esakova, Zhegallo and Rozanov1996). This region represented an isolated Zavkhan Terrane that separated from Eastern Gondwana and collided with Siberia during the Ediacaran–Early Palaeozoic interval (Ruzhentsev & Mossakovsky, Reference Ruzhentsev and Mossakovsky1995; Debrenne, Maidanskaya & Zhuravlev, Reference Debrenne, Maidanskaya and Zhuravlev1999; Kheraskova et al. Reference Kheraskova, Didenko, Bush and Volozh2003). The chemostratigraphic record from this region is fragmentary because of thick intercalations of siliciclastic sediment from which the δ13Ccarb values cannot be read (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996). A composite section through the uppermost Tsagan Oloom and Bayan Gol formations in Western Mongolia is provided herein (Fig. 5).
2.c. Kazakhstan
The region was situated in proximity to East Gondwana and embraces Kazakhstanian terranes (Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001; Popov et al. Reference Popov, Bassett, Zhevchuzhnikov, Holmer, Klishevich and Bassett2009). Among them, sections in the Lesser Karatau Range that include the Kyrshabakty and Chulaktau formations and the Shabakty Group in ascending order are the best studied and most continuous lower Cambrian sequences in Kazakhstan (Missarzhevsky & Mambetov, Reference Missarzhevsky and Mambetov1981; Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001). The Kyrshabakty and Chulaktau formations are biostratigraphically correlated, respectively, with the Nemakit–Daldynian and upper Nemakit–Daldynian–Tommotian stages of Siberia (Missarzhevsky & Mambetov, Reference Missarzhevsky and Mambetov1981; Mambetov, Reference Mambetov and Mambetov1993). First occurrences in members of the lower Shabakty Group are biostratigraphically correlated with the upper Atdabanian–Botoman stages of Siberia (Missarzhevsky & Mambetov, Reference Missarzhevsky and Mambetov1981; Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001).
2.d. South China, India and Iran
From the Yangtze Platform (South China), a set of the oldest first appearances of bilaterian taxa with mineralized skeletons is described herein mainly from the Zhujiaqing Formation of Yunnan and Maidiping Formation of Sichuan. They are correlated bio- and chemostratigraphically with the Nemakit–Daldynian Stage of Siberia (Qian et al. Reference Qian, Zhu, Li, Jiang and Van Iten2002; Li, Zhang & Zhu, Reference Li, Zhang and Zhu2001; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007, Reference Li, Ling, Jiang, Pan, Chen, Cai and Feng2009; Li & Xiao, Reference Li and Xiao2004; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007) and comprise in ascending order the assemblage zones Anabarites trisulcatus–Protohertzina anabarica (SSF1) of the lower Meishucunian Stage, Paragloborilus subglobosus–Purella squamulosa (SSF2) and Watsonella crosbyi (SSF3) of the middle Meishucunian Stage (Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007) (Fig. 3). The upper Meishucunian strata comprise the Sinosachites flabelliformis–Tannuolina zhangwentangi Assemblage Zone (SSF4) and directly underlie the trilobitic interval of the Cambrian in the shallow water realm of the Yangtze Platform. The first occurrences of skeletonized bilaterians within the Parabadiella and Eoredlichia–Wutingaspis zones of the Qiongzhusian Stage are biostratigraphically correlated (but not directly constrained by chemostratigraphy) with the middle–upper Atdabanian and upper Atdabanian–lower Botoman stages of Siberia, respectively (Qian et al. Reference Qian, Zhu, Li, Jiang and Van Iten2002; Li, Zhang & Zhu, Reference Li, Zhang and Zhu2001; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007; Li & Xiao, Reference Li and Xiao2004; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Fig. 3). This interval of earliest trilobitic zones is also more coarsely resolved by the Pelagiella subangulata Taxon Range Zone (SSF5) in the shallow water realm, roughly correlative with the Ninella tarimensis–Cambroclavus fangxianensis Assemblage Zone and the Rhombocorniculum cancellatum Taxon Range Zone at the northern margin of the Yangtze Platform. First appearances of skeletal fossils from sections of the upper Krol and lower Tal formations in the Lesser Himalayas of India and lower Soltanieh Formation in the Elburz Mountains of Iran are correlated with SSF1 of South China (Hamdi, Brasier & Jiang, Reference Hamdi, Brasier and Jiang1989; Hamdi, Reference Hamdi and Hushmandzadeh1995; Hughes et al. Reference Hughes, Peng, Bhargava, Ahluwalia, Walia, Myrow and Parcha2005). The three regions belonged to East Gondwana (Fig. 1).
2.e. Australia
The first appearances of skeletal fossils in East Gondwana are also derived from South Australia. In the Arrowie Basin, first occurrences are documented herein from the lower Wilkawillina and Ajax Limestones, Wirrapowie Limestone and lower Mernmerna Formation, and the Moorowie Formation. These first appearances span the reportedly pre-trilobitic interval and Abadiella huoi–Pararia janeae zones, correlated biostratigraphically with the Atdabanian–Botoman stages of Siberia (Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990; Gravestock et al. Reference Gravestock, Alexander, Demidenko, Esakova, Holmer, Jago, Lin, Melnikova, Parkhaev, Rozanov, Ushatinskaya, Zang, Zhegallo and Zhuravlev2001; Jago, Sun & Zang, Reference Jago, Sun and Zang2002; Jago et al. Reference Jago, Zang, Sun, Brock, Paterson and Skovsted2006; Skovsted, Brock & Paterson, Reference Skovsted, Brock and Paterson2006; Topper et al. Reference Topper, Brock, Skovsted and Paterson2010, Reference Topper, Skovsted, Brock and Paterson2011). In the Stansbury Basin, the earliest skeletal fossils are derived from the Mount Terrible and Winulta formations, broadly correlated with the Nemakit–Daldynian–Tommotian stages of Siberia (Daily, Reference Daily1976, Reference Daily1990; Gravestock & Shergold, Reference Gravestock, Shergold, Zhuravlev and Riding2001; Jago, Sun & Zang, Reference Jago, Sun and Zang2002; Jago et al. Reference Jago, Zang, Sun, Brock, Paterson and Skovsted2006). Other first occurrences from the overlying Kulpara Formation and Parara Limestone are reported from pre-trilobitic beds and Abadiella huoi–Pararia janeae zones (Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990; Gravestock et al. Reference Gravestock, Alexander, Demidenko, Esakova, Holmer, Jago, Lin, Melnikova, Parkhaev, Rozanov, Ushatinskaya, Zang, Zhegallo and Zhuravlev2001; Jago, Sun & Zang, Reference Jago, Sun and Zang2002; Jago et al. Reference Jago, Zang, Sun, Brock, Paterson and Skovsted2006).
2.f. Mediterranean region
Several blocks now in North Africa and Western Europe were situated along the western margin of Gondwana (Fig. 1). Among these, first occurrences from the Corduban Series of central Spain are discussed herein. Based on biostratigraphic correlation with Morocco and chemostratigraphy, the Corduban Series represents a sub-trilobitic part of the lower Cambrian of Western Gondwana (Geyer & Landing, Reference Geyer and Landing2004; Fig. 3). The first appearances from the lower Ovetian Stage in W Europe (Spain, France, Germany, Sardinia) are correlated biostratigraphically with those from the lower Issendalenian Stage in the Anti-Atlas Mountains of Morocco (Geyer & Landing, Reference Geyer and Landing2004). The base of the Issendalenian Stage in Morocco correlates chemostratigraphically with the Delgadella anabara Zone of the Atdabanian Stage of Siberia (Kirshvink et al. Reference Kirschvink, Magaritz, Ripperdan, Zhuravlev and Rozanov1991; Maloof et al. Reference Maloof, Schrag, Crowley and Bowring2005, Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Bengtson, Conway Morris, Cooper, Jell and Runnegarb). First appearances of skeletal fossils from the upper Ludwigsdorf limestones of the Saxothuringian microcontinent (Germany) are correlated with the Atdabanian–Botoman stages of Siberia and the upper Qiongzhusian–lower Canglangpuian stages of South China (Elicki, Reference Elicki1994; Geyer & Elicki, Reference Geyer and Elicki1995).
2.g. Avalonia
The region formed a relatively small terrane of West Gondwanan affinity. In its portion represented by SE Newfoundland, the oldest first appearances are located within the Quaco Road Member of the upper Chapel Island Formation, within the Ladatheca cylindrica Zone of the Fortunian Stage and Watsonella crosbyi Zone of the Cambrian Stage 2 (Landing, Reference Landing2004; Landing & Westrop, Reference Landing, Westrop, E. Landing and Westrop1998; Landing et al. Reference Landing, Myrow, Benus and Narbonne1989, Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007). Other important first occurrences are reported from the upper Cuslett and lower Fosters Point formations (Camenella baltica Zone, correlated with the lower–middle Atdabanian Stage), as well as from the basal part of the overlying Brigus Formation (Callavia broeggeri Zone, correlated with the upper Atdabanian–Botoman stages) (Shergold & Geyer, Reference Shergold and Geyer2003; Fig. 3). In the portion of Avalonia represented by South Britain, the first occurrences regarded herein are from the Home Farm Member, Lower Comley Sandstone and Comley Limestone (Camenella baltica and Callavia zones).
2.h. Laurentia
In the Northwest Territories of Canada the first appearances are reported from the Ingta Formation of the Wernecke Mountains, correlated with the basal Cambrian Anabarites–Protohertzina Zone (Pyle et al. Reference Pyle, Narbonne, Nowlan, Xiao and James2006) and underlain by the Precambrian–Cambrian boundary negative excursion (Narbonne, Kaufman & Knoll, Reference Narbonne, Kaufman and Knoll1994; Pyle et al. Reference Pyle, Narbonne, James, Dalrymple and Kaufman2004), and from the Sekwi Formation of the Mackenzie Mountains and basal Rosella Formation of the Cassiar Mountains, correlated with the Fallotaspis and Nevadella zones (Voronova et al. Reference Voronova, Drosdova, Esakova, Zhegallo, Zhuravlev, Rozanov, Sayutina and Ushatinskaya1987; Dillard et al. Reference Dillard, Pope, Coniglio, Hasiotis and Lieberman2007). In Greenland, important first occurrences are reported from the lower Buen Formation, biostratigraphically correlated with the Nevadella Zone and upper Atdabanian Stage (Conway Morris et al. Reference Conway Morris, Peel, Higgins, Soper and Davis1987; Conway Morris, Reference Conway Morris, Cowie and Brasier1989; Debrenne & Reitner, Reference Debrenne, Reitner, Zhuravlev and Riding2001; Conway Morris & Peel, Reference Conway Morris and Peel2008, Reference Conway Morris and Peel2010), and from the upper Bastion–Ella Island formations, correlated with the Bonnia–Olenellus Zone and Botoman Stage (Skovsted, Reference Skovsted2003, Reference Skovsted2004, Reference Skovsted2006). First occurrences in California and Nevada are from the Campito Formation, from the Fritzaspis, Fallotaspis and Nevadella zones (Durham, Reference Durham1971; Hollingsworth, Reference Hollingsworth2005, Reference Hollingsworth2007; Fig. 3).
2.i. Baltica
The earliest skeletal fauna (Mobergella fauna) on the Baltic Shield known from the Kalmarsund Sandstone and subsurface deposits in southern Sweden is correlated with the Schmidtiellus mickwitzi Zone (Bengtson, Reference Bengtson1968, Reference Bengtson1970, Reference Bengtson1977). First appearances from the Zawiszyn Beds of the upper Klimontovian Stage in Poland (Lendzion, Reference Lendzion1972, Reference Lendzion1978; Reference Lendzion, Volkova, Gnilovskaya, Lendzion, Kirjanov, Palij, Pashkyavichene, Piskun, Posti, Rozanov, Urbanek, Fedonkin and Yankauskas1983; Bengtson, Reference Bengtson1977), Lükati Formation and lower part of the Tiskre Formation in northern Estonia are also attributed to the Schmidtiellus mickwitzi Zone of the Dominopol’ Stage (Bergström, Reference Bergström1973; Mens & Pirrus, Reference Mens and Pirrus1977; Moczydłowska, Reference Moczydłowska2002). These beds are broadly correlated with the Atdabanian Stage of Siberia (Moczydłowska, Reference Moczydłowska2002). The oldest regional first occurrences are reported from the Lontova Formation in Estonia, Platysolenites antiquissimus Zone, broadly correlated with the Terreneuvian Series (Fig. 3).
3. First appearances of mineralized skeletal parts in animal groups
3.a. Sponges and spongiomorphs
The oldest fossils attributed to sponges are reported from the Cryogenian Period. One of the earliest lines of evidence for sponges comes from well-preserved molecular biomarkers of demosponges from strata of the Neoproterozoic Huqf Supergroup of the South Oman Salt Basin pre-dating the termination of the Marinoan glaciation and having a minimum age of c. 635 Ma (McCaffrey et al. Reference McCaffrey, Moldowan, Lipton, Summons, Peters, Jeganathan and Watt1994; Love et al. Reference Love, Fike, Grosjean, Stalvies, Grotzinger, Bradley, Bowring, Condon and Summons2006, Reference Love, Grosjean, Stalvies, Fike, Grotzinger, Bradley, Kelly, Bhatia, Meredith, Snape, Bowring, Condon and Summons2009). Fossils interpreted as sponge-grade metazoans are also found in the pre-Marinoan Trezona Formation of South Australia with a maximum age of 659.7 ± 5.3 Ma (Maloof et al. Reference Maloof, Rose, Beach, Samuels, Calmet, Erwin, Poirier, Yao and Simons2010c). Palaeophragmodictya reticulata Gehling & Rigby, Reference Gehling and Rigby1996, from the Ediacaran Rawnsley Quartzite in South Australia, was interpreted as a hexactinellid (Gehling & Rigby, Reference Gehling and Rigby1996; Debrenne & Reitner, Reference Debrenne, Reitner, Zhuravlev and Riding2001), or a stem-group sponge (Mehl, Reference Mehl1998), but later it was reinterpreted as an attachment disc of a problematic organism of uncertain affinity to sponges or cnidarians (Serezhnikova, Reference Serezhnikova2007). The Late Ediacaran Fedomia mikhaili Serezhnikova & Ivantsov, Reference Serezhnikova and Ivantsov2007 and Vaveliksia vana Ivantsov, Malahovskaya & Serezhnikova, Reference Ivantsov, Malahovskaya and Serezhnikova2004, from the White Sea coast, are likely sponges (Ivantsov, Malahovskaya & Serezhnikova, Reference Ivantsov, Malahovskaya and Serezhnikova2004; Serezhnikova, Reference Serezhnikova2007; Serezhnikova & Ivantsov, Reference Serezhnikova and Ivantsov2007).
3.a.1. Demosponges and hexactinellids
The earliest reported sponge spicules from c. 750 Ma strata in Nevada were attributed to demosponges (Reitner & Wörheide, Reference Reitner, Wörheide, Hooper and van Soest2002; Müller et al. Reference Müller, Li, Schröder, Qiao and Wang2007). Monaxonous thin-walled and hollow spicules of possible hexactinellids derive from Alaska (Allison & Awramik, Reference Allison and Awramik1989; Debrenne & Reitner, Reference Debrenne, Reitner, Zhuravlev and Riding2001), from beds inferred by chemostratigraphy to be Neoproterozoic, most likely pre-Varangerian (Kaufman, Knoll & Awramik, Reference Kaufman, Knoll and Awramik1992). Skeletal remains of sponges found in thin-sections from the Doushantuo Formation on the South China Platform are interpreted as demosponges, because they consist exclusively of siliceous monaxonal spicules (Li, Chen & Hua, Reference Li, Chen and Hua1998). Their maximum age is c. 580 Ma (Condon et al. Reference Condon, Zhu, Bowring, Wang, Yang and Jin2005). These latter, however, were regarded as possible pseudofossils (inorganic crystals) by Zhou, Yuan & Xue (Reference Zhou, Yuan and Xue1998). Spicule-like objects of hexactinellid habit from the Ediacaran Doushantuo and Dengying formations of Hubei Province were observed in thin-section (Tang, Zhang & Jiang, Reference Tang, Zhang and Jiang1978; Zhao et al. Reference Zhao, Xing, Ding, Liu, Zhao, Zhang, Meng, Yin, Ning and Han1988; Steiner et al. Reference Steiner, Mehl, Reitner and Erdtmann1993), but these might also be pseudofossils composed of inorganic crystals (Zhou, Yuan & Xue, Reference Zhou, Yuan and Xue1998) or might instead represent acanthomorphic acritarchs (Zhang, Yuan & Yin, Reference Zhang, Yuan and Yin1998). Spicules with demosponge affinities are known from Cloudina reefs of Namibia (Reitner & Wörheide, Reference Reitner, Wörheide, Hooper and van Soest2002), with an age of c. 550 Ma (after Wood, Grotzinger & Dickson, Reference Wood, Grotzinger and Dickson2002; Grotzinger, Adams & Schröder, Reference Grotzinger, Adams and Schröder2005). Disarticulated and clustered tetracts, pentacts, hexacts and polyactines attributed to upper Ediacaran hexactinellids are preserved in iron oxides in chert layers of the upper Tsagan Oloom Formation of Western Mongolia (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Brasier, Green & Shields, Reference Brasier, Green and Shields1997), but the stratigraphic position of strata yielding these spicules requires further age constraint. Hexactiniellid and/or demospongiid siliceous spicules were mentioned but not illustrated by Brasier & Singh (Reference Brasier and Singh1987, p. 326), Mazumdar & Banerjee (Reference Mazumdar and Banerjee1998) and Tiwari (Reference Tiwari1999). These fossils were recovered from the lowermost Cambrian basal Chert-Phosphate Member of the lower Tal Group, in the Mussoorie, Garwhal and Korgai synclines of the Lesser Himalayas and from the lower part of the Gangolihat Dolomite (Deoban Formation) of the Inner Kumaun Lesser Himalayas (inner carbonate belt) (Tiwari, Pant & Tewari, Reference Tiwari, Pant and Tewari2000), correlated with the lowermost Cambrian (lower Meishucunian) based on the occurrence of protoconodonts (Azmi & Paul, Reference Azmi and Paul2004).
Abundant and extraordinarily preserved hexactinellids and demosponges (including articulated specimens) are well documented from the early Cambrian deep basinal settings of the Yangtze Platform, from the Niutitang black shales of Hunan (Ding & Qian, Reference Ding and Qian1988; Steiner et al. Reference Steiner, Mehl, Reitner and Erdtmann1993) and Hetang black shales of Anhui (Yuan et al. Reference Yuan, Xiao, Parsley, Zhou, Chen and Hu2002). Hexactinellid sponge spicules occur in chert beds of the basal member of the Niutitang Formation of Hunan Province (Steiner et al. Reference Steiner, Mehl, Reitner and Erdtmann1993: p. 6, figs 2–4) and of equivalent strata in Zhejiang Province (Steiner et al. Reference Steiner, Li, Qian and Zhu2004a, fig. 2h), of which the ages are constrained to the lower Meishucunian Stage owing to the occurrence of Kaiyangites. Megasters are not rare in the lower Meishucunian of Shaanxi Province (Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007). By contrast, there are no remains interpreted as sponge spicules from the Meishucunian Stage in its stratotype area of eastern Yunnan (Rigby & Hou, Reference Rigby and Hou1995), where the first spicules and sponge body fossils occur in the basal Yu'anshan Formation of the Qiongzhusian Stage (Steiner et al. Reference Steiner, Zhu, Weber and Geyer2001).
In Siberia, siliceous spicules attributed to the Hexactinellida are known from the lowermost Tommotian Nochorojcyathus sunnaginicus Zone (Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Pel'man et al. Reference Pel'man, Ermak, Fedorov, Luchinina, Zhuravleva, Repina, Bondarev, Borodaevskaya and Repina1990; Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992) and traditionally referred to as Protospongia sp. (Protospongia Salter, Reference Salter1864). Similar fossils have also been reported by Khomentovsky, Val'kov & Karlova (Reference Khomentovsky, Val'kov, Karlova, Khomentovsky and Gibsher1990) and Khomentovsky & Karlova (Reference Khomentovsky and Karlova1993) (but not illustrated) from the pre-Tommotian part of the lower Cambrian, from the upper Ust’-Yudoma Formation (Fig. 4; online Appendix 1 at http://journals.cambridge.org/geo). The earliest spicules of demosponges occur in the Fansycyathus lermontovae Zone of the upper Atdabanian Stage in northern Siberia (Fedorov in Shabanov et al. Reference Shabanov, Astashkin, Pegel, Egorova, Zhuravleva, Pel'man, Sundukov, Stepanova, Sukhov, Fedorov, Shishkin, Vaganova, Ermak, Ryabukha, Yadrenkina, Abaimova, Lopushinskaya, Sychev and Moskalenko1987; online Appendix 1 at http://journals.cambridge.org/geo). The earliest articulated hexactinellids and demosponges in Siberia occur in the lower Botoman Sinsk lagerstätten (Goryansky, Reference Goryansky1977; Ivantsov et al. Reference Ivantsov, Zhuravlev, Leguta, Krasilov, Melnikova and Ushatinskaya2005; Ponomarenko, Reference Ponomarenko2005; Rozanov et al. Reference Rozanov, Parkhaev, Demidenko, Karlova, Korovnikov, Shabanov, Ivantsov, Luchinina, Malakhovskaya, Mel'nikova, Naimark, Ponomarenko, Skorlotova, Sundukov, Tokarev, Ushatinskaya and Kipriyanova2010; online Appendix 1 at http://journals.cambridge.org/geo).
3.a.2. Calcareans
The earliest calcarean sponge spicules of Dodecaactinella sp. occur in the lower Dokidocyathus regularis Zone of Siberia (Kruse, Zhuravlev & James, Reference Kruse, Zhuravlev and James1995; Fig. 2). Pentactines of Sulugurella sulugurica Fedorov in Pel'man et al. Reference Pel'man, Ermak, Fedorov, Luchinina, Zhuravleva, Repina, Bondarev, Borodaevskaya and Repina1990 were originally attributed to the Calcarea and reported from the Nochorojcyathus sunnaginicus Zone of the Tommotian (Pel'man et al. Reference Pel'man, Ermak, Fedorov, Luchinina, Zhuravleva, Repina, Bondarev, Borodaevskaya and Repina1990) and from the probably slightly older uppermost Sukharikha Formation of the Sukharikha River section (Rowland et al. Reference Rowland, Luchinina, Korovnikov, Sipin, Tarletskov and Fedoseev1998), but their hexactinellid affinity is more plausible (A. Zhuravlev, pers. comm., Reference Zhuravlev and Wood2008). Calcarean spicules are also known from the Atdabanian–Botoman-equivalent strata of Mongolia, Western Gondwana, Australia and Laurentia (online Appendix 1 at http://journals.cambridge.org/geo). The first articulated calcarean Gravestockia pharetronensis Reitner, Reference Reitner1992 occurs in Australian beds correlated with the Atdabanian Stage (Reitner Reference Reitner1992; Debrenne & Reitner, Reference Debrenne, Reitner, Zhuravlev and Riding2001; online Appendix 1 at http://journals.cambridge.org/geo). According to Zhuravlev & Wood (Reference Zhuravlev and Wood2008), spicules of first representatives of calcarean sponges are probably high-magnesium calcitic in original composition. Stem-group sponges, which may combine mineralogy and morphology of calcarean and hexactinellid spicules, are also reported from the Cambrian Series 2 (Botting & Butterfield, Reference Botting and Butterfield2005; Harvey, Reference Harvey2010; see also Eiffelia Walcott, Reference Walcott1920 in online Appendix 1 at http://journals.cambridge.org/geo).
3.a.3. Archaeocyaths and other probable aspicular sponges
First archaeocyaths are known from the lowermost Tommotian Stage of the Siberian Platform (e.g. Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969, Reference Rozanov, Repina, Apollonov, Shabanov, Zhuravlev, Pegel’, Fedorov, Astashkin, Zhuravleva, Egorova, Chugaeva, Dubinina, Ermak, Esakova, Sundukov, Sukhov and Zhemchuzhnikov1992, Reference Rozanov, Khomentovsky, Shabanov, Karlova, Varlamov, Luchinina, Pegel’, Demidenko, Parkhaev, Korovnikov and Skorlotova2008; Shabanov et al. Reference Shabanov, Korovnikov, Pereladov, Fefelov, Rozanov and Varlamov2008; Riding & Zhuravlev, Reference Riding and Zhuravlev1995; Kruse, Zhuravlev & James, Reference Kruse, Zhuravlev and James1995). Among them, an undetermined form is reported from the uppermost Ust’-Yudoma Formation (Rozanov et al. Reference Rozanov, Repina, Apollonov, Shabanov, Zhuravlev, Pegel’, Fedorov, Astashkin, Zhuravleva, Egorova, Chugaeva, Dubinina, Ermak, Esakova, Sundukov, Sukhov and Zhemchuzhnikov1992; Shabanov et al. Reference Shabanov, Korovnikov, Pereladov, Fefelov, Rozanov and Varlamov2008), while several species occur in the basal Pestrotsvet Formation of the Lena-Aldan region (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). Archaeocyaths were also reported from the basal Pestrotsvet Formation at Selinde in the southeastern Siberian Platform (Korshunov, Repina & Sysoev, Reference Korshunov, Repina and Sysoev1969; Khomentovsky & Karlova, Reference Khomentovsky and Karlova2002) and the uppermost Sukharikha Formation in the Igarka region of the northwestern margin of the Siberian Platform (Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969, but see Luchinina et al. Reference Luchinina, Korovnikov, Sipin and Fedoseev1997 and Rowland et al. Reference Rowland, Luchinina, Korovnikov, Sipin, Tarletskov and Fedoseev1998). In the latter two localities, carbonates display δ13C oscillations with high positive peaks characteristic of the lower Tommotian boundary beds, although the position of these peaks with respect to this boundary has been questioned (Kouchinsky et al. Reference Kouchinsky, Bengtson, Missarzhevsky, Pelechaty, Torssander and Val'kov2001, Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Val'kov and Young2005, Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007). The skeleton of archaeocyaths is thought to have been originally mineralized with high-magnesium calcite (Zhuravlev & Wood, Reference Zhuravlev and Wood2008). The radiocyaths and cribricyaths are thought to be different groups of aspicular sponges with, respectively, aragonitic and high-magnesium calcitic biomineralization (Zhuravlev & Wood, Reference Zhuravlev and Wood2008). The radiocyaths first appear in the upper Tommotian of the Siberian Platform (Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992), whereas the oldest cribricyaths are known from the Altai-Sayan Folded Area in the first half of the Atdabanian Stage (Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992).
3.b. Cnidariomorphs and problematic tubular forms
Fossils that can be reasonably interpreted as diploblastic-grade metazoans with biomineralized skeletons are represented by a few but locally abundant forms in the upper Ediacaran strata (Grant, Reference Grant1990; Grotzinger, Watters & Knoll, Reference Grotzinger, Watters and Knoll2000; Grotzinger, Adams & Schröder, Reference Grotzinger, Adams and Schröder2005; Wood, Grotzinger & Dixon, Reference Wood, Grotzinger and Dickson2002; Amthor et al. Reference Amthor, Grotzinger, Schröder, Bowring, Ramezani, Martin and Matter2003; online Appendix 1 at http://journals.cambridge.org/geo). There is also a variety of mineralized solitary and modular calcareous corallomorphs in the lower Cambrian (Debrenne, Lafuste & Zhuravlev, Reference Debrenne, Lafuste and Zhuravlev1990). For convenience, all of these fossils are herein collectively termed cnidariomorphs, including such accretionarily growing mineralized solitary tube-like forms as anabaritids and protoconulariids. Hyolithelminths and ‘coleolids’ have a more uncertain biological affinity, and their interpretation as bilaterian organisms (such as annelids) cannot be excluded.
3.b.1. Ediacaran calcified cnidariomorphs
The earliest biomineralizing cnidariomorph fossils are represented by such solitary calcareous (probably aragonitic or high-magnesium calcite) tubular forms as Cloudina and Namacalathus (ECC in online Appendix 2 at http://journals.cambridge.org/geo and Fig. 2). Their lowermost occurrence is documented from the Nama Group in Namibia (Germs, Reference Germs1972; Grant, Reference Grant1990) and coincides with a pronounced positive carbon excursion reaching 8‰ and dated 548.8 ± 1 Ma (Grotzinger et al. Reference Grotzinger, Bowring, Saylor and Kaufman1995; Grotzinger, Watters & Knoll, Reference Grotzinger, Watters and Knoll2000; Condon et al. Reference Condon, Zhu, Bowring, Wang, Yang and Jin2005; Grotzinger, Adams & Schröder, Reference Grotzinger, Adams and Schröder2005; Zhuravlev et al. Reference Zhuravlev, Liñán, Gámez Vintaned, Debrenne and Fedorov2011). Namapoikia rietoogensis Wood, Grotzinger & Dixon, Reference Wood, Grotzinger and Dickson2002 (a calcareous, probably aragonitic, modular form with a robust biomineralized but aspicular skeleton) is of uncertain affinity to sponges or cnidarians and occurs in the Nama Group of Namibia (Wood, Grotzinger & Dixon, Reference Wood, Grotzinger and Dickson2002) in the uppermost Ediacaran beds of the same estimated age.
3.b.2. Corallomorphs
The earliest solitary calcareous corallomorph, Cysticyathus tunicatus Zhuravleva, Reference Zhuravleva1955, appears in the lower Dokidocyathus regularis Zone of the Tommotian Stage of the Siberian Platform, whereas the oldest modular corallomorphs are the khasaktiids Vittia Sayutina and Khasaktia Sayutina (Sayutina, Reference Sayutina1980) from the upper Tommotian of Siberia (Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992; Kruse, Zhuravlev & James, Reference Kruse, Zhuravlev and James1995; online Appendices 1, 2 at http://journals.cambridge.org/geo). Khasaktiids are reconstructed as originally high-magnesium calcitic (Zhuravlev & Wood, Reference Zhuravlev and Wood2008). The oldest known korovinellid- or khasaktiid-like structures described from the uppermost Krol Formation (near the Precambrian–Cambrian boundary) of India (Flügel & Singh, Reference Flügel and Singh2003; online Appendix 1 at http://journals.cambridge.org/geo) were interpreted as sponges/stromatoporoids (Flügel & Singh, Reference Flügel and Singh2003), but their microbial origin was also suggested (Debrenne, Gangloff & Zhuravlev, Reference Debrenne, Gangloff and Zhuravlev1990).
3.b.3. Anabaritids
Anabaritids are a problematic early Cambrian group with originally calcareous, probably aragonitic, skeletons that typically had triradiate symmetry (Kouchinsky et al. Reference Kouchinsky, Bengtson, Feng, Kutygin and Val'kov2009). Anabaritids mark the basal Cambrian strata in Siberia, Mongolia, Kazakhstan, China, India, Iran and Laurentia, but may also occur in the uppermost Ediacaran of Siberia (Karlova & Vodanyuk, Reference Karlova, Vodanyuk and Khomentovsky1985; Karlova, Reference Karlova1987; Knoll et al. Reference Knoll, Grotzinger, Kaufman and Kolosov1995) and Western Mongolia (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Esakova & Zhegallo, Reference Esakova, Zhegallo and Rozanov1996; Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996; online Appendices 1, 2 at http://journals.cambridge.org/geo; Fig. 2).
3.b.4. Protoconulariids
Protoconulariids have a disputed affinity with Palaeozoic conulariids, a group most convincingly placed within the Cnidaria (Van Iten, Zhu & Li, Reference Van Iten, Zhu and Li2010). Calcium phosphatic protoconulariids first appear in the upper Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone of the lower Meishucunian Stage of China. They are represented by Arthrochites emeishanensis Chen, Reference Chen1982, Hexangulaconularia formosa He in Xing et al. Reference Xing, Ding, Luo, He and Wang1984 and probably related forms, such as Carinachites spp. and Emeiconularia trigemme Qian et al. Reference Qian, Van Iten, Cox, Zhu and Zhuo1997 (Brasier, Reference Brasier, Cowie and Brasier1989a; Qian, Reference Qian1989, Reference Qian1999; Qian & Bengtson, Reference Qian and Bengtson1989; Conway Morris & Chen, Reference Conway Morris and Chen1992; Qian, Li & Zhu, Reference Qian, Li and Zhu2001; Zhu et al. Reference Zhu, Li, Zhang, Steiner, Qian and Jiang2001; Qian et al. Reference Qian, Zhu, Li, Jiang and Van Iten2002; Steiner et al. Reference Steiner, Li, Qian and Zhu2004a). Hexangulaconularia formosa is also known from the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone of the Lesser Himalayas (Brasier & Singh, Reference Brasier and Singh1987; Hughes et al. Reference Hughes, Peng, Bhargava, Ahluwalia, Walia, Myrow and Parcha2005). A probably coeval occurrence of Carinachites sp. is described from the upper Ingta Formation in northwestern Canada (Laurentia), also assigned to the Anabarites trisulcatus–Protohertzina anabarica Zone (Pyle et al. Reference Pyle, Narbonne, Nowlan, Xiao and James2006; online Appendices 1, 2 at http://journals.cambridge.org/geo; Fig. 2).
3.b.5. Hyolithelminths
Hyolithelminths have calcium phosphatic tubes that are circular (Hyolithellus Billings, Reference Billings1871) or oval (Torellella Holm, Reference Holm1893) in cross-section. They are similar to Sphenotallus Hall, Reference Hall1847 and Byronia Matthew, Reference Matthew1899 also known from the Cambrian Series 2 and may be cnidarians as well, but do not show budding typical of the Cnidaria (Neal & Hannibal, Reference Neal and Hannibal2000; Van Iten, Zhu & Collins, Reference Van Iten, Zhu and Collins2002; Van Iten et al. Reference Van Iten, Vyhlasova, Zhu and Qian2005; Peng et al. Reference Peng, Zhao, Wu, Yuan and Tai2005). Hyolithelminths are alternatively compared to annelid tubes (Fisher, Reference Fisher and Moore1962; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Kiel & Dando, Reference Kiel and Dando2009; Johnston et al. Reference Johnston, Johnston, Collom, Powell and Pollock2009; Skovsted & Peel, Reference Skovsted and Peel2011). Hyolithelminths are well known from the lower Tommotian Stage of Siberia (Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983). Their first representatives are reported, but not illustrated, from the upper Purella Zone of the Nemakit–Daldynian Stage (Khomentovsky et al. Reference Khomentovsky, Val'kov, Karlova, Nuzhnov, Khomentovsky, Yakshin and Karlova1983; Khomentovsky, Val'kov & Karlova, Reference Khomentovsky, Val'kov, Karlova, Khomentovsky and Gibsher1990; Khomentovsky & Karlova, Reference Khomentovsky and Karlova1993; Varlamov et al. Reference Varlamov, Rozanov, Khomentovsky, Shabanov, Abaimova, Demidenko, Karlova, Korovnikov, Luchinina, Malakhovskaya, Parkhaev, Pegel, Skorlotova, Sundukov, Sukhov, Fedorov and Kipriyanova2008). Probably the earliest hyolithelminths are represented by Hyolithellus spp. from the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone of South China (Brasier, Reference Brasier, Cowie and Brasier1989a; Qian & Bengtson, Reference Qian and Bengtson1989; Qian, Reference Qian1999; Qian et al. Reference Qian, Zhu, Li, Jiang and Van Iten2002), Ingta Formation of Laurentia (Pyle et al. Reference Pyle, Narbonne, Nowlan, Xiao and James2006) and Lower Tal Formation of India (Brasier & Singh, Reference Brasier and Singh1987; online Appendices 1, 2 at http://journals.cambridge.org/geo; Fig. 2).
3.b.6. Problematic tubular forms
Other widespread tubular problematics are represented by calcareous, probably aragonitic forms Coleolella Missarzhevsky in Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969, Coleoloides Walcott, Reference Walcott1889 and, possibly, Coleolus Hall, Reference Hall1876 (to which ‘Coleolus’ trigonus Sysoev, Reference Sysoev1962 is attributed; see online Appendix 1 at http://journals.cambridge.org/geo; Fig. 2). Coleoloides trigeminatus, Coleolella billingsi (Sysoev, Reference Sysoev1962) and ‘Coleolus’ trigonus Sysoev, Reference Sysoev1962 are reported from the lowermost Tommotian N. sunnaginicus Zone (Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983) of Siberia. They also occur in probably older beds, where carbonates display high positive δ13C peaks, whose position with respect to the lower Tommotian boundary has been questioned (Kouchinsky et al. Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Val'kov and Young2005, Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007). C. typicalis is known from the Tiksitheca licis Zone of Western Mongolia and Watsonella crosbyi Zone of Avalonia (Landing et al. Reference Landing, Myrow, Benus and Narbonne1989). Coleoloides is also reported from the Anabarites trisulcatus–Protohertzina anabarica Zone of India (Brasier & Singh, Reference Brasier and Singh1987), but the material requires further revision.
3.c. Protoconodonts
The protoconodonts are a group of phosphatic simple-cone-shaped sclerites with a deep cavity, lamellar and often longitudinally fibrous wall, and with accretionary growth on the inner side and basal margin (Bengtson, Reference Bengtson1976, Reference Bengtson1977, Reference Bengtson1983). Protoconodonts are microstructurally different from para- and euconodonts (see Section 3.q); finds of animals with grouped protoconodonts in the Chengjiang fossil lagerstätte (Chen & Huang, Reference Chen and Huang2002; Vannier et al. Reference Vannier, Steiner, Renvoise, Hu and Casanova2007) and partially articulated protoconodont apparatuses of lower Meishucunian species Mongolodus longispinus (Vannier et al. Reference Vannier, Steiner, Renvoise, Hu and Casanova2007) demonstrate their affinity with chaetognathans (Szaniawski, Reference Szaniawski1982, Reference Szaniawski2002; Hamdi, Brasier & Jiang, Reference Hamdi, Brasier and Jiang1989; Azmi, Reference Azmi, Pandey, Azmi, Bhandari and Dave1996; McIlroy & Szaniawski, Reference McIlroy and Szaniawski2000; Doguzhaeva, Mapes & Mutve, Reference Doguzhaeva, Mapes and Mutvei2002; Azmi & Paul, Reference Azmi and Paul2004; Qian et al. Reference Qian, Li, Zhu, Steiner and Erdtmann2004; Pyle et al. Reference Pyle, Narbonne, Nowlan, Xiao and James2006; Vannier et al. Reference Vannier, Steiner, Renvoise, Hu and Casanova2007; see also Remarks in section ‘Protoconodonts’ in online Appendix 1 at http://journals.cambridge.org/geo). The primary nature of phosphatization in protoconodonts has been questioned, however, because microstructures rather suggest that in analogy with grasping hooks of modern chaetognaths the protoconodonts had an organic composition, i.e. of hardened chitin (Steiner & Li, Reference Steiner and Li2010).
Among these fossils, Protohertzina anabarica and P. unguliformis (probably morphotypes within the same species collectively referred to as the Protohertzina anabarica group, e.g. by Brasier, Reference Brasier, Cowie and Brasier1989b) represent the earliest skeletal remains attributed to protoconodonts and, probably, bilaterian animals in general (Qian & Bengtson, Reference Qian and Bengtson1989). The first protoconodonts of the P. anabarica group appear in the Anabarites trisulcatus Zone of the Nemakit–Daldynian Stage on the Siberian Platform and in roughly contemporaneous beds in South China, India, Iran, Laurentia and probably Western Mongolia and Kazakhstan (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). With the resolution of stratigraphic correlation available, it is not possible to warrant, however, their earlier appearance than the first calcareous sclerites and shells (see next Section).
3.d. Molluscs, hyoliths, halwaxiids and chancelloriids
Coiled, conical or cyrtoconic shells occur frequently in lower Cambrian faunas worldwide. These fossils resemble shells of undisputed molluscs in overall form and shell microstructure. It is largely accepted, therefore, that they are the earliest representatives of the Phylum Mollusca, falling more likely within its stem group (e.g. Peel, Reference Peel1991; Budd, Reference Budd2001). Univalved shells with solid walls (not composed of sclerites) presumably represent complete and adult external skeletons (but see Martí Mus, Palacios & Jensen, Reference Martí Mus, Palacios and Jensen2008) and are considered herein to be molluscs. Some other problematic shells or plates, probably parts of multiplated chiton-like exoskeletons, may also belong to this group or to the Halwaxiida (see Section 3.k; online Appendices 1 & 2 at http://journals.cambridge.org/geo).
The complete exoskeleton of hyoliths consists of a conical shell (conch), apertural lid (operculum) and may include a pair of curved retractable appendages (helens) protruding between the conch and operculum. These fossils are usually classified either within the Phylum Mollusca or Annelida, or in a separate phylum-level group (e.g. Runnegar et al. Reference Runnegar, Pojeta, Morris, Taylor, Taylor and McClung1975; Runnegar, Reference Runnegar1980; Kouchinsky, Reference Kouchinsky2000).
The Halwaxiida Conway Morris & Caron, Reference Conway Morris and Caron2007 is an apparently monophyletic group that includes biomineralizing calcareous scale-bearing siphogonuchitids and halkieriids. Their compound scleritomes (= full set of sclerites of one individual, after Bengtson, Reference Bengtson1985) could also contain conical shells (Bengtson, Reference Bengtson1992a). Determining whether specific early Cambrian mollusc-like shells represent the complete skeleton or only a part of the scleritome is not always straightforward (see Section 3.k and ‘Other fossils’ in online Appendix 1 at http://journals.cambridge.org/geo). Representatives of the Halwaxiida have been united with sedentary Chancelloriida in the problematic Coeloscleritophora, whose members share possession of hollow calcareous sclerites secreted at a fixed size by internal soft tissue and a similar skeletal microstructure (Bengtson & Missarzhevsky, Reference Bengtson, Missarzhevsky and Taylor1981; Bengtson & Conway Morris Reference Bengtson, Conway Morris, Lipps and Signor1992; Conway Morris & Peel, Reference Conway Morris and Peel1995; Bengtson, Reference Bengtson and Briggs2005; Porter, Reference Porter2008). Halwaxiids can be considered to be stem-group lophotrochozoans, while the general body morphology and affinity of chancelloriids is more problematic (Bengtson & Hou, Reference Bengtson and Hou2001; Janussen, Steiner & Zhu, Reference Janussen, Steiner and Zhu2002; Randell et al. Reference Randell, Lieberman, Hasiotis and Pope2005; Bengtson, Reference Bengtson and Briggs2005; Sperling, Pisani & Peterson, Reference Sperling, Pisani, Peterson, Vickers-Rich and Komarower2007; Porter, Reference Porter2008).
These four groups (molluscs, hyoliths, halwaxiids and chancelloriids) appeared in the geological record at about the same time. Their evolutionary radiation in the earliest Cambrian was an important early step for metazoans, which gave rise to numerous forms typical of the lower Cambrian strata worldwide and dominated most of the pre-trilobitic bilaterian fossil assemblages. With the current fidelity of stratigraphic correlation available it is not yet possible to further resolve the relative order of first appearances of molluscs, hyoliths and coeloscleritophorans (Fig. 2).
The oldest such fossils occur at the same level in the upper Anabarites trisulcatus–lower Purella zones of the Nemakit–Daldynian Stage of the southeastern Siberian Platform (Khomentovsky, Val'kov & Karlova, Reference Khomentovsky, Val'kov, Karlova, Khomentovsky and Gibsher1990) correlated with the lowermost part of the rising trend in the lower Cambrian δ13C values, around feature Z of the Siberian δ13C reference scale (Brasier, Khomentovsky & Corfield, Reference Brasier, Khomentovsky and Corfield1993; Fig. 4). The fossils are represented by mollusc-like shells, such as cyrtoconic planispiral Oelandiella Vostokova, Reference Vostokova1962 and sinistrally coiled Barskovia Golubev, Reference Golubev1976, conchs of hyoliths, siphogonuchitid sclerites and scaly shells of Purella Missarzhevsky, Reference Missarzhevsky, Zhuravleva and Rozanov1974. The earliest hyoliths are also described from the probably time-equivalent basal Purella Zone of Western Mongolia (Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996), SSF1 of South China (Qian & Bengtson, Reference Qian and Bengtson1989; Steiner et al. Reference Steiner, Li, Qian and Zhu2004a) and India (Brasier & Singh, Reference Brasier and Singh1987; Hughes et al. Reference Hughes, Peng, Bhargava, Ahluwalia, Walia, Myrow and Parcha2005). Siphogonuchitid sclerites and shells of Maikhanella Zhegallo in Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982 are also reported from SSF1 beds of South China (Qian & Bengtson, Reference Qian and Bengtson1989; Steiner et al. Reference Steiner, Li, Qian and Zhu2004a), whereas Purella defines the base of the Purella Zone in Western Mongolia (Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996; Fig. 5).
The earliest confirmed appearance of Chancelloria is represented by articulated sclerites from the Purella Zone of the Nemakit–Daldynian Stage of Siberia (Khomentovsky, Val'kov & Karlova, Reference Khomentovsky, Val'kov, Karlova, Khomentovsky and Gibsher1990) and correlated with a trough between Siberian isotopic features Z and I (Brasier, Khomentovsky & Corfield, Reference Brasier, Khomentovsky and Corfield1993; Fig. 4). Identification of fossils reported as single rays of Chancelloria sp. from the Anabarites trisulcatus and basal Purella zones of the Nemakit–Daldynian Stage of the southeastern Siberian Platform (Khomentovsky, Val'kov & Karlova, Reference Khomentovsky, Val'kov, Karlova, Khomentovsky and Gibsher1990; Khomentovsky & Karlova, Reference Khomentovsky and Karlova2005; Brasier, Khomentovsky & Corfield, Reference Brasier, Khomentovsky and Corfield1993) is questionable (Zhuravlev et al. Reference Zhuravlev, Liñán, Gámez Vintaned, Debrenne and Fedorov2011). Likewise, the occurrence of Chancelloriidae in SSF1 of South China (Qian & Bengtson, Reference Qian and Bengtson1989) has not been confirmed (M. Steiner, unpub. obs.; Online Appendix 1 at http://journals.cambridge.org/geo). Single-rayed sclerites of Cambrothyra are known, however, from SSF1. This organism is treated as closely related to, but outside the Chancelloridae sensu stricto (Steiner et al. Reference Steiner, Li, Qian and Zhu2004a, fig. 2; Moore et al. Reference Moore, Porter, Steiner and Li2010).
3.e. Cambroclavids and paracarinachitids
Cambroclaves are originally calcareous (probably aragonitic) sclerites without concentrically arranged growth increments and consist of a basal hollow shield usually bearing a spine (Qian, Reference Qian1978; Mambetov & Repina, Reference Mambetov, Repina and Zhuravleva1979; Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990; Conway Morris & Chen, Reference Conway Morris and Chen1991; Conway Morris et al. Reference Conway Morris, Crampton, Xiao and Chapman1997; Elicki & Wotte, Reference Elicki and Wotte2003). Paracarinachitids are probably related forms, but were formed by overlapping growth increments (Qian & Bengtson, Reference Qian and Bengtson1989). These fossils are regarded as protective sclerites of bilaterally symmetrical animals without reliable systematic position (Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990). They were alternatively interpreted as receptaculitids (algae or sponges) (Dzik, Reference Dzik1994), protoconodonts (Mambetov & Repina, Reference Mambetov, Repina and Zhuravleva1979), acanthocephalans (Qian & Yin, Reference Qian and Yin1984) or such ecdysozoan groups as priapulids (Conway Morris et al. Reference Conway Morris, Crampton, Xiao and Chapman1997) and lobopodians (Qian, Reference Qian1999; Liu et al. Reference Liu, Shu, Han, Zhang and Zhang2007). A fragment of an articulated body covered with cambroclavid-type sclerites was reported from the Sirius Passet fauna and has been tentatively assigned to the Ecdysozoa (Conway Morris & Peel, Reference Conway Morris and Peel2010). The fibrous ultrastructure of the wall in cambroclavids and paracarinachitids (Qian & Bengtson, Reference Qian and Bengtson1989; Conway Morris & Chen, Reference Conway Morris and Chen1991), typical of the other calcareous fossils discussed above, such as molluscs, hyoliths, halwaxiids and chancelloriids, are nevertheless compatible with a lophotrochozoan affinity.
The first sclerites of cambroclavids (Zhijinites Qian, Reference Qian1978) and paracarinachitids (Paracarinachites Qian & Jiang in Luo et al. Reference Luo, Jiang, Wu, Song and Ouyang1982) are known from the Yangtze Platform, Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (SSF2) of the middle Meishucunian Stage, marked by a rising trend in the carbon isotope record below a prominent positive peak ZHUCE (Qian, Reference Qian1999; Qian et al. Reference Qian, Zhu, Li, Jiang and Van Iten2002; Qian & Bengtson, Reference Qian and Bengtson1989; Bengtson, Reference Bengtson, Schopf and Klein1992b; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
3.f. Tommotiids
Tommotiids are represented by calcium phosphate sclerites with evidence of basal marginal accretionary growth. These fossils have been interpreted as sclerites of larger scleritomes of problematic animals (Bengtson, Reference Bengtson1970, Reference Bengtson, Lipps and Waggoner2004; Holmer et al. Reference Holmer, Skovsted, Brock, Valentine and Paterson2008; Skovsted et al. Reference Skovsted, Brock, Paterson, Holmer and Budd2008, Reference Skovsted, Holmer, Larsson, Högström, Brock, Topper, Balthasar, Petterson Stolk and Paterson2009b). The taxonomy of tommotiids was discussed by Landing (Reference Landing1984, Reference Landing1995), Bengtson (Reference Bengtson1986), Laurie (Reference Laurie1986), Bengtson et al. (Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990), Conway Morris & Chen (Reference Conway Morris and Chen1990), Esakova & Zhegallo (Reference Esakova, Zhegallo and Rozanov1996) and Skovsted et al. (Reference Skovsted, Balthasar, Brock and Paterson2009a). Based on the organophosphatic shell composition and morphological similarities, tommotiids have been regarded as closely related to brachiopods (Williams & Holmer, Reference Williams and Holmer2002). Recent discoveries of articulated tommotiids show distinct affinities to lophophorates, including a sessile habit and brachiopod-like shells within the scleritome (Skovsted et al. Reference Skovsted, Brock, Paterson, Holmer and Budd2008, Reference Skovsted, Holmer, Larsson, Högström, Brock, Topper, Balthasar, Petterson Stolk and Paterson2009b; Holmer et al. Reference Holmer, Skovsted, Brock, Valentine and Paterson2008).
Classical tommotiids, such as Camenella Missarzhevsky in Rozanov & Missarzhevsky, Reference Rozanov and Missarzhevsky1966, occur in the basal Tommotian Stage deposits of the southeastern Siberian Platform (Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969). Without description or illustration, they are also reported from apparently older beds of the northern Siberian Platform (Fedorov & Shishkin, Reference Fedorov, Shishkin and Krasnov1984; Khomentovsky & Karlova, Reference Khomentovsky and Karlova1993; Luchinina et al. Reference Luchinina, Korovnikov, Sipin and Fedoseev1997; Meshkova et al. Reference Meshkova, Nikolaeva, Kulikov, Zhuravleva, Luchinina, Musatov, Sidoras and Zhuravleva1976; Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969). These beds contain pre-Tommotian prominent positive carbon isotopic peaks (Kouchinsky et al. Reference Kouchinsky, Bengtson, Missarzhevsky, Pelechaty, Torssander and Val'kov2001, Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007; Fig. 4; online Appendices 1 & 2 at http://journals.cambridge.org/geo). Probably the earliest appearance of Camenella is that reported from Western Mongolian beds at the beginning of the rising trend towards positive peak D (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 9; Fig. 5), which may correspond to the rising trend towards Siberian peaks I or I′ (Fig. 4).
Porcauricula hypsilippis (Jiang, Reference Jiang1980) is known from lower SSF2 beds (Qian & Bengtson, Reference Qian and Bengtson1989), in the rising trend towards a prominent positive peak ZHUCE in China (Brasier et al. Reference Brasier, Magaritz, Corfield, Luo, Wu, Ouyang, Jiang, Hamdi, He and Frazier1990; Li et al. Reference Li, Ling, Jiang, Pan, Chen, Cai and Feng2009). Lapworthella ludvigseni Landing, Reference Landing1984 and Eccentrotheca kanesia Landing, Nowlan & Fletcher, Reference Landing1980 occur in the lower Watsonella crosbyi Zone of SE Newfoundland (Landing et al. Reference Landing, Myrow, Benus and Narbonne1989). Hence, the first tommotiids reported from Western Mongolia, South China and Avalonia are probably older than those from the base of the Tommotian Stage in its stratotype in the southeastern part of the Siberian Platform (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
3.g. Tianzhushanellids
Tianzhushanellids are bivalved and probably originally aragonitic shelly fossils assigned to the Family Tianzhushanellidae Conway Morris in Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990 with uncertain higher-rank taxonomy (see ‘Other fossils’ in Online Appendix 1 at http://journals.cambridge.org/geo). They were referred to molluscs (Parkhaev, Reference Parkhaev1998), but have more recently been regarded as the best candidates for calcitic-shelled stem-group brachiopods (Balthasar, Reference Balthasar2008; Li, Reference Li, Smith, O'Brien and Caron2009). New articulated material of Apistoconcha from strata correlative with the Cambrian Stage 3 in Australia seems to support its stem-group brachiopod position (Skovsted et al. Reference Skovsted, Holmer, Topper and Brock2010). The earliest undoubted representatives of the group are known from the SSF3 of China (Qian, Reference Qian1999; Li & Chen, Reference Li and Chen1992; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007) and occur at near the peak of the positive carbon isotope excursion ZHUCE in Sichuan (Brasier et al. Reference Brasier, Magaritz, Corfield, Luo, Wu, Ouyang, Jiang, Hamdi, He and Frazier1990, fig. 6; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
3.h. Brachiopods
The earliest reported brachiopod species with a calcium phosphate (organophosphatic) shell is the paterinid Aldanotreta sunnaginensis Pel'man, Reference Pel'man1977. It first appears on the Siberian Platform in the N. sunnaginicus Zone of the Tommotian Stage (Bengtson et al. Reference Bengtson, Fedorov, Missarzhevsky, Rozanov, Zhegallo and Zhuravlev1987; Ushatinskaya & Malakhovskaya, Reference Ushatinskaya and Malakhovskaya2001; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 4). The earliest Siberian brachiopod with a calcareous shell is Nochoroiella isitica Pel'man in Grigor'eva, Melnikova & Pel'man, Reference Grigor'eva, Melnikova and Pel'man1983 (assigned to obolellids, although its affinity with brachiopods was questioned: see online Appendices 1 & 2 at http://journals.cambridge.org/geo) and Obolella sp. from the D. lenaicus Zone of the Tommotian Stage (Figs 2, 4). Chemostratigraphic correlation with Siberia (cf. Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996) suggests that these occurrences are likely pre-dated by Khasagtina primaria Ushatinskaya, Reference Ushatinskaya1987 from Western Mongolia. The latter fossil is originally assigned to kutorginids, but its affinity to brachiopods needs revision (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 5).
3.i. Stenothecoids
Stenothecoids, a group of probably low-magnesium calcite (Zhuravlev & Wood, Reference Zhuravlev and Wood2008) enigmatic bivalved organisms with serial paired imprints sometimes present, occur in Cambrian Series 1–3 (Aksarina, Reference Aksarina and Selyatitskiy1968; Yochelson, Reference Yochelson1969; Runnegar & Pojeta, Reference Runnegar and Pojeta1974; Rozov, Reference Rozov1984; Pel'man, Reference Pel'man1985; online Appendices 1 & 2 at http://journals.cambridge.org/geo). In Siberia, the first reliable stenothecoids appear in the late Tommotian Stage of the Altai-Sayan Folded Area (Pel'man et al. Reference Pel'man, Aksarina, Koneva, Popov, Sobolev and Ushatinskaya1992; Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992), but the earliest stenothecoids overall (Stenothecoides sp. and S. yochelsoni) are reported respectively from Western Mongolia (Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982; Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996, fig. 13; Fig. 5; online Appendices 1 & 2 at http://journals.cambridge.org/geo) and the SSF3 Zone of South China (Yu, Reference Yu1996; online Appendices 1 & 2 at http://journals.cambridge.org/geo). Voronin et al. (Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982) defined the Stenothecoides Zone in Western Mongolia at a level correlated chemostratigraphically (cf. Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Fig. 5) with prominent positive peaks of the uppermost Nemakit–Daldynian Stage of the Siberian Platform. Consequently, like brachiopods, the first stenothecoids are reported from Cambrian Stage 2, probably below the lower Tommotian boundary (Fig. 2).
3.j. Mobergellids
Mobergellids are low conical or disc-shaped calcium phosphate problematic fossils with evidence of accretionary growth and paired radiating (possibly, muscular) imprints on the interior side (Bengtson, Reference Bengtson1968; Conway Morris & Chapman, Reference Conway Morris and Chapman1997; Skovsted, Reference Skovsted2003; Dzik, Reference Dzik2010). The first occurrence of Mobergella sibirica Skovsted, Reference Skovsted2003 (formerly Mobergella radiolata Bengtson, Reference Bengtson1968) in Siberia is known from the lower Dokidocyathus lenaicus–Tumuliolynthus primigenius Zone of the Tommotian Stage (Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Missarzhevsky, Reference Missarzhevsky1989; Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 4). Mobergella sp. is also reported from the upper D. regularis Zone of the Tommotian Stage (Repina et al. Reference Repina, Lazarenko, Meshova, Korshunov, Nikiforov and Aksarina1974), but was not illustrated. Fossils described as Mobergella, but without characteristic radiating imprints, are known from the middle Meishucunian Stage of China (Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007; online Appendix 1 at http://journals.cambridge.org/geo), although no definite record of mobergellids exists from China (Qian & Bengtson, Reference Qian and Bengtson1989; Streng & Skovsted, Reference Streng and Skovsted2006). The most reliable first appearance of the group is thus in the lower D. lenaicus Zone of the Tommotian Stage (upper part of the Cambrian Stage 2), within the carbon isotope trough between peaks III and IV of the Siberian reference scale (Brasier et al. Reference Brasier, Corfield, Derry, Rozanov and Zhuravlev1994a, Reference Brasier, Rozanov, Zhuravlev, Corfield and Derryb; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 4).
3.k. Other problematic shells and sclerites
Besides shells assigned herein to molluscs, hyoliths, halwaxiids, brachiopods, mobergellids, stenothecoids and tianzhushanellids, there are shells or plates of a calcareous or unknown original composition, often with evidence of accretionary growth. Some of these may represent parts of scleritomes similar to those of halkieriids or chitons (Conway Morris, McIlroy & Rushton, Reference Conway Morris, McIlroy and Rushton1998; Vendrasco et al. Reference Vendrasco, Li, Porter and Fernandez2009; see ‘Molluscs’ and ‘Other fossils’ in online Appendix 1 at http://journals.cambridge.org/geo). A number of such problematic shells are reported from China, where they are found in beds of the middle Meishucunian Stage (Bengtson, Reference Bengtson, Schopf and Klein1992b; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007). Most of them are treated as problematic molluscs by Bengtson (Reference Bengtson, Schopf and Klein1992b) and Li et al. (Reference Li, Steiner, Zhu, Zhu and Erdtmann2007), but some forms among them are alternatively regarded as ‘brachiopod-like’ or ‘problematic brachiopods’ by Qian, Li & Zhu (Reference Qian, Li and Zhu2001).
An array of problematic sclerites from Cambrian Series 1–2 is known from Siberia and elsewhere, and some of the sclerites can be assigned to larger groups discussed herein, such as protoconodonts, chancelloriids, halwaxiids, tommotiids and cambroclavids or to ecdysozoan cuticular sclerites and fragments of carapaces, but the others are more problematic (see Bengtson, Reference Bengtson, Schopf and Klein1992b; Qian et al. Reference Qian, Li, Zhu, Steiner and Erdtmann2004; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007; see ‘Other fossils’ in online Appendix 1 at http://journals.cambridge.org/geo).
3.l. Trilobites
The Class Trilobita are advanced arthropods (Phylum Arthropoda) with calcareous (low-magnesium calcite) carapaces (Wilmot & Fallick, Reference Wilmot and Fallick1989). The first occurrences of trilobites in the world (highlighted by thick horizontal lines within Stage 3 interval in Fig. 3) post-date or are estimated to be no earlier than those on the Siberian Platform, where their first undoubted appearance is just above the base of the Atdabanian Stage (Lieberman, Reference Lieberman2002; Hollingsworth, Reference Hollingsworth2005, Reference Hollingsworth2007, Reference Hollingsworth, Rábano, Gozalo and García-Bellido2008; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). The earliest trilobites (Family Archaeaspididae, Order Redlichiida) are represented by Profallotaspis Repina in Khomentovsky & Repina, Reference Khomentovsky and Repina1965 from Siberia and Fritzaspis Hollingsworth, Reference Hollingsworth2007 from Laurentia.
3.m. Bradoriids
Bradoriids known from Cambrian to lower Ordovician rocks worldwide have originally weakly or secondarily mineralized (phosphatized) carapaces (Jones & McKenzie, Reference Jones and McKenzie1980; Landing, Reference Landing, Nowlan and Fletcher1980; Butterfield, Reference Butterfield2003; Zhang, Reference Zhang2007; Zhang, Dong & Maas, Reference Zhang, Dong and Maas2011). These arthropods are represented by the Order Bradoriida Raymond, Reference Raymond1935 (Bradoriida sensu stricto) and Phosphatocopida Müller, Reference Müller1964 (Hou et al. Reference Hou, Siveter, Williams and Feng2002; Maas & Waloszek, Reference Maas and Waloszek2005; Williams et al. Reference Williams, Siveter, Popov and Vannier2007; Zhang, Reference Zhang2007). The former are regarded at present as stem-group crustaceans, whereas the latter are considered by some to be crown-group crustaceans (Hou et al. Reference Hou, Siveter, Williams, Walossek and Bergström1996, Reference Hou, Williams, Siveter, Siveter, Aldridge and Sansom2010; Shu et al. Reference Shu, Luo, Conway Morris, Zhang, Hu, Chen, Han, Zhu, Li and Chen1999). Although other stem-group crustaceans represented by non-mineralizing Isoxys zhurensis Ivantsov, Reference Ivantsov1990 are known as early as the middle Profallotaspis jakutensis Zone of the Atdabanian Stage on the southeastern Siberian Platform (Ivantsov, Reference Ivantsov1990), the first bradoriid Cambria Neckaja & Ivanova, Reference Neckaja and Ivanova1956 is reported in Siberia from the middle Atdabanian Delgadella anabara Zone (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 4). Coeval first occurrences of bradoriids are also known from South China (Hou et al. Reference Hou, Siveter, Williams and Feng2002), Gondwana (Hinz-Schallreuter, Gozalo & Liñán, Reference Hinz-Schallreuter, Gozalo and Liñán2008; Topper et al. Reference Topper, Skovsted, Brock and Paterson2011), Avalonia (Brasier, Reference Brasier, Cowie and Brasier1989c) and Baltica (Streng, Ebbestad & Moczydłowska, Reference Streng, Ebbestad and Moczydłowska2008).
3.n. Lobopodians
Lobopodians have been considered a Cambrian stem group of the Panarthropoda, including such phyla as Onychophora, Tardigrada and Arthropoda (see Budd, Reference Budd, Fortey and Thomas1997; Liu et al. Reference Liu, Shu, Han, Zhang and Zhang2007; Edgecombe, Reference Edgecombe2009). These animals were similar to modern onychophorans and are often regarded as members of the Phylum Lobopodia Snodgrass, Reference Snodgrass1938 (Hou & Bergström, Reference Hou and Bergström1995) or Tardipolypoda Chen & Zhou, Reference Chen and Zhou1997. Exceptionally preserved in the Chengjiang fossil lagerstätte, they are elongate animals with paired limbs and sclerites (trunk plates) situated along each side of the body (Chen, Hou & Lu, Reference Chen, Hou and Lu1989; Chen, Zhou & Ramsköld, Reference Chen, Zhou and Ramsköld1995; Ramsköld & Hou, Reference Ramsköld and Hou1991; Bergström & Hou, Reference Bergström and Hou2001; Hou et al. Reference Hou, Ma, Zhao and Bergström2004). The sclerites are thought to be of an original calcium phosphatic composition, although their biomineralization has been doubted by Steiner et al. (Reference Steiner, Li, Qian, Zhu and Erdtmann2007).
The first sclerites of Microdictyon sp. in Siberia are reported herein from the Emyaksin Formation of the Bol'shaya Kuonamka River, from beds correlated by carbon isotope chemostratigraphy with the upper Delgadella anabara Zone of the Atdabanian Stage (Kouchinsky et al., unpub. data; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). This occurrence is somewhat older than the previously reported occurrence from the upper Atdabanian Fansycyathus lermontovae Zone of Lena River (Varlamov et al. Reference Varlamov, Rozanov, Khomentovsky, Shabanov, Abaimova, Demidenko, Karlova, Korovnikov, Luchinina, Malakhovskaya, Parkhaev, Pegel, Skorlotova, Sundukov, Sukhov, Fedorov and Kipriyanova2008). It is correlated chemostratigraphically (Dillard et al. Reference Dillard, Pope, Coniglio, Hasiotis and Lieberman2007; online Appendices 1 & 2 at http://journals.cambridge.org/geo) with the regional first occurrence of Microdictyon sp. and Microdictyon cf. rhomboidale Bengtson, Matthews & Missarzhevsky, Reference Bengtson1986 in the lower Nevadella Zone of Laurentia (Bengtson, Matthews & Missarzhevsky, Reference Bengtson, Matthews, Missarzhevsky, Hoffman and Nitecki1986; Voronova et al. Reference Voronova, Drosdova, Esakova, Zhegallo, Zhuravlev, Rozanov, Sayutina and Ushatinskaya1987; Fig. 3) and, probably, with the first occurrence of M. sphaeroides Hinz, Reference Hinz1987 in the Camenella baltica Zone of Avalonia (Hinz, Reference Hinz1987) and Microdictyon depressum Bengtson in Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990 in the Abadiella huoi Zone of Australia. The first soft-bodied lobopodians with remains of sclerites are already diverse and well known from the Chengjiang fossil lagerstätte, which is time-equivalent with the upper Atdabanian Stage of Siberia.
3.o. Palaeoscolecids
The Class Palaeoscolecida Conway Morris & Robison, Reference Conway Morris and Robison1986 is known from the Cambrian Series 2 to the Upper Silurian. These fossils are morphologically similar to nematomorphs (Hou & Bergström, Reference Hou and Bergström1994) and priapulids (Conway Morris, Reference Conway Morris1997). They are broadly considered as a stem group of the Priapulida (Harvey, Dong & Donoghue, Reference Harvey, Dong and Donoghue2010) or Cycloneuralia (Budd, Reference Budd2001; Conway Morris & Peel, Reference Conway Morris and Peel2010). Complete preservation of their worm-like bodies is known, but far more often they occur as disarticulated calcium phosphate cuticular sclerites. The first palaeoscolecids, represented by the biomineralized trunk sclerites Hadimopanella apicata Wrona, Reference Wrona1982 are reported herein from the middle Nochoroicyathus kokoulini Zone (lower Judomia Zone) of northern Siberia (Kouchinsky et al., unpub. data; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 4). The earliest soft-bodied preservation of palaeoscolecids is known from the Sirius Passet fauna of Laurentia (Greenland; Conway Morris & Peel, Reference Conway Morris and Peel2010; online Appendices 1 & 2 at http://journals.cambridge.org/geo) and the Chengjiang fossil lagerstätte of South China (Hu, Reference Hu2005), which are time-equivalent with the upper Atdabanian of Siberia.
3.p. Echinoderms
The earliest fossils of the phylum Echinodermata are disarticulated and presumably originally high-magnesium calcite plates (Dickson, Reference Dickson2002, Reference Dickson2004). Some of these plates derive from brachiole-bearing echinoderms and thus can be assigned to eocrinoids. The ‘class’ Eocrinoidea Jaekel, Reference Jaekel1918 is paraphyletic and comprises stem-groups of other blastozoan clades (e.g. Sprinkle, Reference Sprinkle1973). The earliest echinoderm plates like these are reported herein from the upper Delgadella anabara Zone of the Atdabanian Stage from Siberia (Kouchinsky et al., unpub. data; Zamora et al. in press; online Appendices 1 & 2 at http://journals.cambridge.org/geo; Figs 2, 4). First echinoderm plates from the lower part of the Nevadella Zone of Laurentia are chemostratigraphically correlated with the upper part of the Delgadella anabara Zone of Siberia (Dillard et al. Reference Dillard, Pope, Coniglio, Hasiotis and Lieberman2007; Fig. 3). The earliest isolated echinoderm plates occur in lower Ovetian beds of West Gondwana (Spain) broadly correlated with the Delgadella anabara Zone of Siberia and also appear contemporaneously in South China and Australia (Zamora et al. in press).
3.q. Chordates (paraconodonts and agnathans)
Conodonts (including para- and euconodonts) are characterized by calcium phosphatic dental structures with dentin-like tissue (but see Kemp, Reference Kemp2002a, Reference Kempb). They are regarded as the earliest known biomineralized chordates (Donoghue, Forey & Aldridge, Reference Donoghue, Forey and Aldridge2000; Donoghue, Sansom & Downs, Reference Donoghue, Sansom and Downs2006). There is a microstructural similarity between the two sub-groups as well as a histological and morphological transition from paraconodonts to euconodonts, and therefore they are united as conodonts (Szaniawski & Bengtson, Reference Szaniawski and Bengtson1993, Reference Szaniawski, Bengtson and Szaniawski1998).
The oldest paraconodonts from Siberia are Westergaardodina cf. tricuspidata Müller, Reference Müller1959 and Furnishina sp. from the Paibian Stage-equivalent Kutugunian Horizon, which contains the upper Cambrian (Furongian Series) SPICE carbon isotope excursion (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Shabanov et al. Reference Shabanov, Korovnikov, Pereladov, Fefelov, Rozanov and Varlamov2008). Furnishina and Westergaardodina are known, however, from the Drumian Stage (within the Cambrian Series 3, or traditional Middle Cambrian) in most other crustal units. The oldest paraconodonts, represented by Furnishina sp. and Westergaardodina sp., are reported respectively from the Triplagnostus gibbus Zone of the uppermost part of Cambrian Stage 5 (lower Series 3) in Sweden (Müller, Reference Müller1959, Reference Müller1971; Dong, Reference Dong2004) and from Stage 5 deposits of Alaska (Dutro et al. Reference Dutro, Palmer, Repetski and Brosgé1984), but the former is much better preserved and constrained stratigraphically.
The earliest vertebrates with ossification in the dermal skeleton, that contains dentin-like tissue (Smith, Sansom & Repetski, Reference Smith, Sansom and Repetski1996; Karatajute-Talimaa, Reference Karatajute-Talimaa1997), are similar to the Ordovician agnathans and represented by disarticulated calcium phosphatic plates of Anatolepis sp. from the mid-Sunwaptan Stage of Laurentia (Smith & Sansom, Reference Smith and Sansom1995; Smith, Sansom & Cochrane, Reference Smith, Sansom, Cochrane and Ahlberg2001) corresponding to the upper part of the Cambrian Stage 9 (Furongian Series).
4. Discussion
There is no unambiguous evidence of the existence of bilaterian superphyla in the Precambrian (Budd, Reference Budd2008; Budd & Jensen, Reference Budd and Jensen2000, Reference Budd, Jensen, Donoghue and Smith2003), although some bilaterally symmetrical forms from the late Ediacaran, younger than 555 Ma, are interpreted as their representatives (e.g. Fedonkin & Waggoner, Reference Fedonkin and Waggoner1997). On the other hand, Ediacara-type fossils have repeatedly been reported from the Early Palaeozoic (Conway Morris, Reference Conway Morris1993; Jensen, Gehling & Droser, Reference Jensen, Gehling and Droser1998; Samuelson, Van Roy & Vecoli, Reference Samuelson, Van Roy and Vecoli2001; Zhang & Babcock, Reference Zhang and Babcock2001), but none of those is comparable with typical Ediacaran vendobionts in their structure, symmetry and growth pattern (Antcliffe & Brasier, Reference Antcliffe and Brasier2008; Zhuravlev et al. Reference Zhuravlev, Liñán, Gámez Vintaned, Debrenne and Fedorov2011).
The general succession of first appearances of biomineralized skeletal parts during the Cambrian radiation includes two main sets clustered geochronologically by high-rank phylogeny and reflects two successive phases of diversification of bilaterians (Fig. 2). Such a pulsed diversification accompanied a general increase in generic diversity of the biota towards the early Botoman maximum preceding the Botoman–Toyonian biotic crisis (Brasier et al. Reference Brasier, Corfield, Derry, Rozanov and Zhuravlev1994a; Zhuravlev & Wood, Reference Zhuravlev and Wood1996; Zhuravlev, Reference Zhuravlev, Zhuravlev and Riding2001; Li et al. Reference Li, Steiner, Zhu, Zhu and Erdtmann2007).
The first phase recognized herein is marked by a set of first appearances of biomineralization in the Terreneuvian Epoch (c. 541–521 Ma), mainly within the Fortunian Age. The latter embraces the first c.10 Ma of the Cambrian Period (Fig. 2). During the Terreneuvian Epoch such major skeletal groups as protoconodonts, halwaxiids, chancelloriids, hyoliths, molluscs, tommotiids, brachiopods, tianzhushanellids, stenothecoids, cambroclavids–paracarinachitids and mobergellids first appeared and diversified, as well as other forms with problematic affinities to these groups (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). All of them, except protoconodonts and possibly chancelloriids, can be comfortably placed within the total-group Lophotrochozoa. Among them, such extant biomineralizing lophotrochozoan phyla as Mollusca and Brachiopoda can be recognized. Annelida does not have undisputed biomineralized representatives during the Cambrian radiation (Conway Morris & Peel, Reference Conway Morris and Peel2008), but such problematic groups as hyolithelminths, hyoliths and halwaxiids may share that affinity (see Section 3 and remarks in online Appendix 1 at http://journals.cambridge.org/geo). By contrast, Bryozoa certainly appear relatively late, in the uppermost Cambrian, at the onset of the Ordovician radiation (Landing, English & Keppie, Reference Landing, English and Keppie2010).
Protoconodonts were among the first bilaterians to acquire hardened (probably mineralized calcium phosphatic) skeletal parts (presumably for active predation). These have been interpreted as grasping spines of the Chaetognatha, a protostomian phylum close to the Lophotrochozoa, but most likely its sister-group (Halanych, Reference Halanych2004; Dunn et al. Reference Dunn, Hejnol, Matus, Pang, Browne, Smith, Seaver, Rouse, Obst, Edgecombe, Sørensen, Haddock, Schmidt-Rhaesa, Okusu, Kristensen, Wheeler, Martindale and Giribet2008). They appeared in the fossil record at about the same time as the first calcareous sclerites of coeloscleritophorans, shells of molluscs and hyoliths. The majority of bilaterian skeletal fossils of the Terreneuvian Series belong to these latter groups. Their first appearances occurred in beds younger than the Precambrian–Cambrian boundary and the carbon isotope negative anomaly associated with this boundary (Narbonne, Kaufman & Knoll, Reference Narbonne, Kaufman and Knoll1994; Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Kimura et al. Reference Kimura, Matsumoto, Kakuwa, Hamdi and Zibaseresht1997; Li et al. Reference Li, Ling, Jiang, Pan, Chen, Cai and Feng2009), but not younger than those with Siberian δ13C feature Z and attributed to the upper Anabarites trisulcatus–lower Purella zones of the Nemakit–Daldynian Stage (Brasier, Khomentovsky & Corfield, Reference Brasier, Khomentovsky and Corfield1993; Brasier, et al. Reference Brasier, Corfield, Derry, Rozanov and Zhuravlev1994a, Reference Brasier, Rozanov, Zhuravlev, Corfield and Derryb). These first appearances can be correlated with the Anabarites trisulcatus–Protohertzina anabarica (SSF1) Assemblage Zone of the lower Meishucunian Stage of South China, where all of them except shells of molluscs occur in beds older than the tuffite with a combined depositional age of 535.2 ± 1.7 Ma (Zhu et al. Reference Zhu, Li, Hou, Pan, Wang, Deng and He2009).
Calcium phosphatic sclerites of tommotiids, probably members of the stem group of organophosphatic brachiopods, have a younger first appearance associated with the same rising trend, but still below highly positive peaks of the lower part of the Cambrian Stage 2. The first reported appearance of tommotiids is older than the estimated c. 530 Ma base of the Cambrian Stage 2 in Avalonia (Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007). Such groups as cambroclavids–paracarinachitids, other problematic shells with affinities close to molluscs or halwaxiids, and some problematic sclerites also appear during this interval (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
Brachiopods, apparently related to tommotiids, are another major group of lophotrochozoan animals with their first appearance in the Cambrian Stage 2. Together with tianzhushanellids, which probably represent stem-group calcareous brachiopods, and stenothecoids, their first occurrences are registered from Stage 2 strata older than or coeval with the Nemakit–Daldynian–Tommotian boundary of c. 525 Ma (Maloof et al. Reference Maloof, Porter, Moore, Dudás, Bowring, Higgins, Fike and Eddy2010a, Reference Maloof, Ramezani, Bowring, Fike, Porter and Mazouadb). Mobergella, Rhombocorniculum and some other problematic sclerites appeared during the upper part of Cambrian Stage 2 (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
Biomineralization in ecdysozoans and deuterostomians did not apparently occur during the first phase of the early Cambrian bilaterian radiation, but these events mark the second phase. Non-biomineralized ecdysozoans, however, existed before, as evidenced by: (1) fossilized embryos of Markuelia secunda Val'kov from the basal Tommotian Stage in Siberia, having already a sclerotized cuticle and representing the Scalidophora, a sub-group of the Cycloneuralia (Bengtson & Yue, Reference Bengtson and Yue1997; Dong et al. Reference Dong, Donoghue, Cheng and Liu2004, Reference Dong, Donoghue, Cunningham, Liu and Cheng2005, Reference Dong, Bengtson, Gostling, Cunningham, Walossek, Harvey, Kouchinsky, Val'kov, Repetski, Stampanoni and Donoghue2010); embryos and possibly related larvae of Pseudooides prima from the lower Meishucunian Stage of South China, considered as possible stem-group arthropods (Steiner et al. Reference Steiner, Zhu, Li, Qian and Erdtmann2004b) and (2) the arthropod-type trace fossils in pre-trilobitic strata (Crimes, Reference Crimes1987; Weber & Zhu, Reference Weber and Zhu2003; Weber, Steiner & Zhu, Reference Weber, Steiner and Zhu2007).
The second phase of the early Cambrian bilaterian radiation is thus marked by the first appearances of biomineralization in the Superphylum Ecdysozoa within Cambrian Stage 3, during < 5 Ma (Fig. 2). The earliest undoubted and widespread event in the second phase is the appearance of calcification in carapaces of trilobites at the base of Stage 3, succeeded by the first appearances of calcium phosphate carapaces of bradoriids (Phylum Arthropoda), sclerites of lobopodians (Phylum Tardipolypoda) and palaeoscolecids (?Phylum Priapulida) in that order, in the early and middle parts of Stage 3 (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2). The diversity of ecdysozoans and their dominance in the Burgess Shale-type fossil communities in upper Stage 3, by the conclusion of the Cambrian radiation, is well documented in such fossil lagerstätten as the Sirius Passet of north Greenland and the roughly time-equivalent Chengjiang of South China (e.g. Conway Morris & Peel, Reference Conway Morris and Peel2008, Reference Conway Morris and Peel2010).
The second phase also involves the Superphylum Deuterostomia, Phylum Echinodermata. The earliest biomineralizing representatives of echinoderms appear in the middle of Stage 3. Among them eocrinoids, members of the stem-group Echinodermata, can be recognized. The origin of their high-magnesium calcite stereom skeleton is the first documented skeletal biomineralization event among deuterostomians. Tunicates (Phylum Chordata) may have also existed at that time (Chengjiang fauna, Chen et al. Reference Chen, Huang, Peng, Chi, Wang and Feng2003, but see Bergström, Reference Bergström and Long2010), and some of them may have produced biomineralized spicules (see Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990). Non-mineralizing representatives of the Phylum Chordata were also present during the Cambrian radiation: hagfish-like fossils from Chengjiang are most likely the first vertebrates, but without traces of biomineralization (Shu et al. Reference Shu, Vannier, Luo, Chen, Zhang and Hu1999; Conway Morris, Reference Conway Morris2006; Bergström, Reference Bergström and Long2010). The first appearances of calcium phosphatic dental mineralization in problematic chordates/conodonts (i.e. paraconodonts) in the upper part of Cambrian Stage 5 (Donoghue, Reference Donoghue2002; Donoghue & Sansom, Reference Donoghue and Sansom2002; Kemp, Reference Kemp2002a, Reference Kempb; online Appendices 1 & 2 at http://journals.cambridge.org/geo) and dermal biomineralization in agnathan-like chordates by the end of the Cambrian (Cambrian Stage 9) (Smith & Sansom, Reference Smith and Sansom1995; Smith, Sansom & Cochrane, Reference Smith, Sansom, Cochrane and Ahlberg2001; Young, Reference Young2009) post-date both the Cambrian radiation and the first fossils interpreted as chordates by c. 10 and 25 Ma, respectively.
The fossil record of sponges and cnidarians suggests that they acquired skeletal biomineralization in the Precambrian (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2), but it was not until diverse bilaterian groups become abundant in the fossil record that radiation among skeletonized non-bilaterians like these occurred. Although solitary tubular forms with questionable affinities to cnidarians are already diverse during the first phase of the early Cambrian bilaterian radiation, biomineralized remains of sponges and spongiomorphs are sporadic until the Cambrian Stage 2, where corallomorphs also entered the fossil record (online Appendices 1 & 2 at http://journals.cambridge.org/geo; Fig. 2).
Distribution of calcium carbonate skeletal mineralogies from upper Ediacaran to lower Cambrian (Fig. 2) are shown to reflect fluctuations in the magnesium/calcium ratio in the ocean (Zhuravlev, Reference Zhuravlev1993; Ushatinskaya & Zhuravlev, Reference Ushatinskaya and Zhuravlev1994; Porter, Reference Porter2007; Zhuravlev & Wood, Reference Zhuravlev and Wood2008). First appearances of animal groups with aragonite skeletons occur mainly during the Fortunian Age. A few rare and speculative cases of low-magnesium calcite (LMC) biomineralization have been attributed to the Cambrian Stage 2, including Khasagtina primaria (kutorginid brachiopod?) and the earliest stenothecoids (Zhuravlev & Wood, Reference Zhuravlev and Wood2008; Fig. 2), but their mineralogy is only suggested from the inferred composition of younger forms, and hence alternative carbonate mineralogies cannot be excluded.
The second phase of the early Cambrian bilaterian radiation occurred after one of the major perturbances in the carbon cycle in the Phanerozoic Earth history, marked with a c. 10‰ negative shift in δ13C record in the middle of Stage 2 (online Fig. S1 at http://journals.cambridge.org/geo). The upper part of Stage 2, generally correlated herein with the Tommotian Stage of Siberia, represents a transitional interval with first appearances of high-magnesium calcite (HMC) skeletons, a transition that probably corresponds to decreasing Mg/Ca ratio of the ocean. The second phase, marked by the first appearance of trilobites (Fig. 2), is characterized by the onset of the LMC biomineralization in trilobites and brachiopods and a short-term switch to the calcite sea in the early Atdabanian (Zhuravlev & Wood, Reference Zhuravlev and Wood2008). Based on distribution of inorganic precipitates and the relative abundance of genera with aragonite and HMC v. LMC skeletal mineralogies, the remaining part of the second phase is characterized, however, as a reappearance of the aragonite sea (Zhuravlev & Wood, Reference Zhuravlev and Wood2008). Hence, despite an increasing number of genera with LMC skeletons during the second phase, the Cambrian radiation interval was generally aragonite and HMC favourable.
Acknowledgements
Artem Kouchinsky acknowledges support from the NASA Astrobiology Institute and the Swedish Research Council (Grant No. 621-2001-1751 to Stefan Bengtson and Grant No. 623-2003-207 to Artem Kouchinsky) at the initial stage of work in 2002–2005. Artem Kouchinsky was also supported later from the NordCEE (Nordic Centre for Earth Evolution) project (Danish National Research Foundation (Danmarks Grundforskningsfond)) grant to Prof. Donald Canfield. Igor Korovnikov (Institute of Oil-and-Gas Geology and Geophysics, Novosibirsk, Russia) and John Malinky are acknowledged for useful personal communication. We also thank Andrej Zhuravlev (Universidad de Zaragoza, Spain) for invaluable information and critical revision of this work. Ed Landing and Martin Brasier provided detailed reviews of the manuscript.
Appendix 1
Siberian first appearances in the Nemakit–Daldynian Stage (see Fig. 4)
S1. Sclerites of protoconodonts Protohertzina anabarica and P. unguliformis (Nemnekey; Khomentovsky & Karlova, Reference Khomentovsky, Karlova and Khomentovsky1991, Reference Khomentovsky and Karlova1993; projected onto the Siberian δ13C reference scale by Brasier, Khomentovsky & Corfield, 1993).
Remarks. Anabaritids are also reported from this level and below the Precambrian-Cambrian boundary negative excursion (see data in online Appendix 1 at http://journals.cambridge.org/geo, not shown in Fig. 4).
S2. Shelly molluscs Oelandiella sp. and Barskovia sp., halwaxiids Purella cristata, Siphogonuchites aff. triangularis, and orthothecid hyoliths Lophotheca socialis (Mt Konus; Khomentovsky, Val'kov & Karlova, 1990; projected onto the Siberian δ13C reference scale by Brasier, Khomentovsky & Corfield, 1993).
Remarks. Spicules of hexactinellids Protospongia sp. and ?sclerites of chancelloriids Chancelloria sp. are also reported from that level (Gonam; Khomentovsky & Karlova, Reference Khomentovsky and Karlova1993; projected herein onto the Siberian δ13C reference scale in accordance with Brasier, Khomentovsky & Corfield, 1993).
S3. Tubes of hyolithelminths Hyolithellus sp., sclerites of chancelloriids Chancelloria sp., problematic sclerites of Fomitchella cf. infundibuliformis (Gonam; Semikhatov & Serebryakov, Reference Semikhatov and Serebryakov1983; Khomentovsky et al. Reference Khomentovsky, Val'kov, Karlova, Nuzhnov, Khomentovsky, Yakshin and Karlova1983; Khomentovsky & Karlova, Reference Khomentovsky and Karlova1993; projected herein onto the Siberian δ13C reference scale in accordance with Brasier, Khomentovsky & Corfield, 1993).
Siberian first appearances in the Tommotian Stage (see Fig. 4)
S4. Tommotiid Sunnaginia imbricata, archaeocyaths, siliceous stauracts and pentacts of the hexactinellid sponges (Aldan; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Shabanov et al. Reference Shabanov, Korovnikov, Pereladov, Fefelov, Rozanov and Varlamov2008); problematic tubes Tommototubulus savitzkyi Fedorov, Reference Fedorov1986 (Aldan; Khomentovsky, Val'kov & Karlova, 1990).
S5. Calcium-phosphate brachiopods Aldanotreta sunnaginensis (Aldan; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Bengtson et al. Reference Bengtson, Fedorov, Missarzhevsky, Rozanov, Zhegallo and Zhuravlev1987; Ushatinskaya & Malakhovskaya, Reference Ushatinskaya and Malakhovskaya2001); problematic sclerites Tumulduria incomperta (Aldan; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Bengtson et al. Reference Bengtson, Fedorov, Missarzhevsky, Rozanov, Zhegallo and Zhuravlev1987; Rozanov et al. Reference Rozanov, Repina, Apollonov, Shabanov, Zhuravlev, Pegel’, Fedorov, Astashkin, Zhuravleva, Egorova, Chugaeva, Dubinina, Ermak, Esakova, Sundukov, Sukhov and Zhemchuzhnikov1992) and Archaeopetasus sp. (as opercula of Coleolella billingsi in Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969, pl. 7) (Aldan; Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969; Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990; Dzik, Reference Dzik1994); problematic tubes Coleolella billingsi (Sysoev, Reference Sysoev1962) (Aldan; Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983), ‘Coleolus’ trigonus Sysoev, Reference Sysoev1962 (Aldan; Sysoev, Reference Sysoev1962; Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969), and Coleoloides trigeminatus Missarzhevsky in Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969 (Aldan; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983).
S6. Spicules of the calcarean sponge Dodecaactinella sp. and thecae of corallomorph Cysticyathus tunicatus (Middle Lena; Kruse, Zhuravlev & James, 1995).
S7. Mobergella sibirica Skovsted, Reference Skovsted2003 (formerly Mobergella radiolata Bengtson, Reference Bengtson1968) (Aldan; Rozanov et al. Reference Rozanov, Missarzhevsky, Volkova, Voronova, Krylov, Keller, Korolyuk, Lendzion, Michniak, Pykhova and Sidorov1969; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Missarzhevsky, Reference Missarzhevsky1989; Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992).
S8. Calcium-carbonate brachiopods Nochoroiella isitica and Obolella sp. (Middle Lena; Grigor'eva, Melnikova & Pel'man, 1983; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov Reference Rozanov and Sokolov1984; Pel'man et al. Reference Pel'man, Aksarina, Koneva, Popov, Sobolev and Ushatinskaya1992;
S9. Problematic sclerites Rhombocorniculum insolutum (Middle Lena; Sokolov & Zhuravleva, Reference Sokolov and Zhuravleva1983; Rozanov & Sokolov, Reference Rozanov and Sokolov1984; Brasier, Reference Brasier, Cowie and Brasier1989b; Rozanov & Zhuravlev, Reference Rozanov, Zhuravlev, Lipps and Signor1992. Bol'shaya Kuonamka; Kouchinsky et al., unpub. data and herein).
Siberian first appearances in the Atdabanian Stage (see Fig. 4)
S10. Trilobites Profallotaspis sp. (Middle Lena; Rozanov & Sokolov, Reference Rozanov and Sokolov1984).
S11. Microdictyon sp. (Bol'shaya Kuonamka; Kouchinsky et al., unpub. data and herein).
S12. Eocrinoidea indet. (Bol'shaya Kuonamka; Kouchinsky et al., unpub. data and herein).
S13. Hadimopanella apicata (Bol'shaya Kuonamka; Kouchinsky et al., unpub. data and herein).
Appendix 2
Mongolian first appearances (see Fig. 5)
M1. Hexactinellid spicules (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 5; Braiser, Green & Shields, 1997); anabaritids A. trisulcatus and C. decurvatus (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996; Esakova & Zhegallo, Reference Esakova, Zhegallo and Rozanov1996; Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996) or Anabarites sp. (Esakova & Zhegallo, Reference Esakova, Zhegallo and Rozanov1996).
M2. Protoconodonts Protohertzina unguliformis; scaly shells of halwaxiids Purella sp.; orthothecid? hyoliths Pseudorthotheca bicostata Qian (indicated as P. bistriata in Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996, fig. 13, p. 386) (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, fig. 9; Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996, fig. 13).
M3. Tommotiids Camenella applanata and Camenella sp. (Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996, fig. 3 on p. 386; Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996), Camenella cf. baltica (Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982); problematic tubes Hyolithellus cf. vladimirovae (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996) or Hyolithellus sp. (Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982) and Coleolella billingsi (Brasier et al. Reference Brasier, Shields, Kuleshov and Zhegallo1996, figs 6, 7).
M4. Shelled molluscs Obtusoconus honorabilis and Granoconus trematus (Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996).
M5. Sclerites of chancelloriids Chancelloria sp. (Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982).
M6. Calcium carbonate-shelled brachiopod? Khasagtina primaria Ushatinskaya, Reference Ushatinskaya1987 (described as Kundatella sp. by Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982).
M7. Stenothecoides sp. (Voronin et al. Reference Voronin, Voronova, Grigor'eva, Drozdova, Zhegallo, Zhuravlev, Ragozina, Rozanov, Sayutina, Sysoeva and Fonin1982; Khomentovsky & Gibsher, Reference Khomentovsky and Gibsher1996, fig. 13).









