


The past decade has seen a major shift in our thoughts about the origins of eating disorders. Anorexia nervosa has traditionally been regarded as a disorder with social and cultural explanations, rather than a developmental or biological disorder (Reference Nasser, Katzman and GordonNasser et al, 2000). For about a century from the first formalisation of the concept of anorexia nervosa by Lasègue gue the disorder was regarded as a non-specific, environmentally responsive neurosis of women with its roots in the dysfunctional family. Only in the 1970s did anorexia nervosa emerge as a neurotic syndrome in its own right. At about the same time, bulimia nervosa was first described by Russell (Reference Russell1979), and since then the prevalence has risen, sharply outnumbering cases of anorexia nervosa. Descriptions of binge eating disorder followed soon after. By the 1980s, eating disorders were thought of as an extreme manifestation of societal obsessions with thinness, with common subclinical syndromes present in the population. In the 1990s this causality shifted to gender dynamics, focusing on power and self-determination, rather than being related to the biology of female gender. Current sociocultural explanations include worldwide cultural dynamics, such as cultures in transition (‘Westernising’ societies) and confused gender identities. However, the biggest change in the perception of eating disorders came in the 1990s, when converging evidence from twin and family studies showed that they have a genetic component.
Although there is no doubt that these sociocultural explanations are important and relevant, anorexia, bulimia and obesity are perhaps better regarded as heterogeneous disorders with complex multifactorial aetiology, involving the interaction of genes and the environment, particularly social factors. However, little is known about this aetiology, particularly its biological components, and this is severely hampering the development of new treatments.
THE PHENOTYPE: WHAT ARE EATING DISORDERS?
The diagnosis of eating disorders is a tricky business: diagnosis can be unstable, with clinical features changing over time, often switching from anorexia to bulimia, and there is frequently comorbidity with other diagnoses such as anxiety and depression (Reference Fairburn and HarrisonFairburn & Harrison, 2003). Risk factors can be common to the two disorders, or distinct, such as dieting. This combination of overlapping and distinct features makes research into fixed risk factors such as genes and environmental events difficult to perform.
So how can nosology be improved to better reflect the underlying structure of eating disorders? There is a lack of empirical evidence for DSM–IV criteria for eating disorders, and some suggest that they may not adequately describe the key symptoms (Reference Hebebrand, Casper and TreasureHebebrand et al, 2004). On the one hand, the modern criteria used to define the illness have been criticised for overemphasis on pathoplastic features of eating disorders such as fear of fat, which are absent in many cases; on the other hand, the tendency to bunch eating disorders together can mask important differences, such as in eating behaviour itself. For example, binge eating occurs across the weight spectrum from anorexia nervosa to obesity. Both eating behaviours and body composition have been used for diagnostic purposes. In clinical studies, anorexia nervosa – the restricting type – emerges as a distinct phenotype, and is now subtyped into restricting and binge–purging subtypes, but transitions between these subtypes are common. Family studies have found four classes of eating disorders, which cluster in families: restricting anorexia nervosa, with and without obsessive–compulsive symptoms, a combination of purging anorexia and bulimia nervosa, and finally bulimia nervosa with self-induced vomiting (Reference Keel, Fichter and QuadfliegKeel et al, 2004).
Stable phenotypes: a developmental perspective
Given the temporal instability between clinical syndromes which obfuscates research into fixed markers such as genes, there has been a move to examine more stable features such as personality and neurobiological traits. These features precede the onset of the eating disorder and are present in childhood, making them developmental in origin (Reference Connan, Campbell and KatzmanConnan et al, 2003). Childhood perfectionism appears to be a particularly strong antecedent risk factor (Reference Anderluh, Tchanturia and Rabe-HeskethAnderluh et al, 2003). Some enduring personality traits appear linked to specific types of eating behaviours, whereas others are more general (Reference Karwautz, Rabe-Hesketh and CollierKarwautz et al, 2002). For example, individuals with both constrained (anorexia) and disinhibited (bulimia) eating share the personality trait of high harm avoidance. However, novelty-seeking personality is linked specifically to those who show disinhibited eating behaviour, as is exploratory excitability, impulsivity and extravagance (Reference Vervaet, van Heeringen and AudenaertVervaet et al, 2004). Impulsivity in binge eating is particularly marked in the context of negative affect. In contrast, constrained eating (as seen in restricting anorexia nervosa) is linked to compulsivity. It is therefore interesting to note that including obsessionality as part of the behavioural phenotype of anorexia nervosa may be a powerful tool for genetic linkage analysis (Reference Devlin, Bacanu and KlumpDevlin et al, 2002). These personality traits may be linked to the underlying neurobiological template.
Compulsivity may be linked to perceptual and cognitive inflexibility. People with anorexia nervosa have reduced flexibility in the acute state and, for some measures, also after recovery (Reference Tchanturia, Anderluh and MorrisTchanturia et al, 2004). This appears to be a familial trait not shared by those with bulimia nervosa, which is associated with childhood obsessive–compulsive traits. Thus, inflexibility might be an endophenotype linked to constrained eating. In turn, this inflexibility might possibly be linked to a difficulty in extinguishing learnt behaviours, a hypothesis raised by Strober (Reference Strober2004).
Clues also come from neurobiology. The complex links between personality, biology and environment have been the particular focus of work by Steiger and colleagues, particularly how serotonergic factors, personality trait variations and developmental typologies converge (Reference SteigerSteiger, 2004). One particular serotonin receptor, 5-HT2A, has been found to have abnormal levels in recovered patients, with different diagnostic groups showing differing alterations (Reference Kaye, Frank and MeltzerKaye et al, 2001). Thus, abnormalities in central serotonin function vary across the eating disorder spectrum. The fact that these persist after recovery suggest that serotonin function might be a traits marker for eating disorders.
GENES
In the 1980s twin studies in anorexia nervosa led to a radical re-evaluation of the long-standing emphasis on the social and cultural explanations of eating disorders, and it is now accepted by most that eating disorders have a genetic component (Reference Bulik, Sullivan and WadeBulik et al, 2000). Although it is not possible to draw firm conclusions on the exact contribution of genes and environment to eating disorders, it is reasonable to suggest that they contribute to the disorder in roughly equal measure.
Many traits related to eating disorders also have a heritable component. These include binge eating, self-induced vomiting, drive for thinness, dietary restraint and disinhibition (Reference Bulik, Sullivan and WadeBulik et al, 2000), several of which are quantitative traits which can be measured in the normal population. The realisation that eating disorders have in part a genetic basis has led to molecular studies aimed at finding genetic vulnerability factors. The driving force behind this is that almost nothing is known about the aetiology of eating disorders, and the discovery of susceptibility genes would be an important step in its unravelling.
Molecular genetic analysis has focused on both genome-wide linkage analysis in multiply affected families in order to try to identify chromosomal loci harbouring genes, and on candidate gene studies. Genome-wide linkage analysis has not found loci for the broad diagnostic category of anorexia and bulimia nervosa, but analysis of pedigrees with at least one pair of relatives with restricted anorexia nervosa identified linkage to chromosome 1p34 (Reference Grice, Halmi and FichterGrice et al, 2002), consistent with the idea that the latter might be a distinct phenotype.
Quantitative genetics has also been used to try to overcome current diagnostic limitations. Drive for thinness and obsessionality tracked most closely with anorexia nervosa, providing novel genetic loci on chromosomes 1 for a composite measure and 13 for drive for thinness (Reference Devlin, Bacanu and KlumpDevlin et al, 2002). Bulimia nervosa, in particular the form with self-induced vomiting, has been linked to chromosome 10 (Reference Bulik, Devlin and BacanuBulik et al, 2003), strengthening the idea that bulimia with self-induced vomiting might be a distinct phenotype. Interestingly, this position is also a known locus for obesity, consistent with the elevated family history of obesity seen in bulimia nervosa.
Association studies have focused on the genes specifically relating to the neurochemistry of eating behaviour. One such candidate is the serotonin receptor 5-HT2A gene HTR2A. Candidate gene association analysis has identified a putative risk allele in this gene, and this has been replicated in some but not all studies. It is possible that heterogeneity within clinical groups might explain differences between studies, as there is the suggestion that HTR2A is associated with constrained rather than disinhibited eating behaviour. Tantalisingly, HTR2A maps to a chromosome 13 linkage peak for the drive for thinness phenotype (Reference Devlin, Bacanu and KlumpDevlin et al, 2002).
The brain-derived neurotrophic factor (BDNF) gene has also been implicated as a susceptibility gene for anorexia nervosa. This protein is involved in regulating feeding behaviour in the hypothalamus, including the regulation of serotonin levels, and low levels are implicated in depression. A multicentre European study found strong evidence that a polymorphism in this gene increases susceptibility to all eating disorder diagnoses, perhaps through its involvement in affective symptoms (Reference Ribases, Gratacos and Fernandez-ArandaRibases et al, 2004).
An alternative approach is to examine models of eating disorders in animals. Feeding behaviour is similar among mammals, and many animals – including mice and pigs – display behaviours reminiscent of human eating disorders (Reference Owen, Treasure and CollierOwen et al, 2001). For example, mice and rats given food for only 2 h per day and allowed access to a running wheel develop behavioural hyperactivity and weight loss analogous to anorexia nervosa. This points to fundamental mammalian behaviours that might explain some of the phenotypes seen in human eating disorders, and which could be dissected by genetic analysis in rodents and used to model new forms of treatment (Reference Exner, Hebebrand and RemschmidtExner et al, 2000).
FUTURE RESEARCH
Increasingly, the consensus is that eating disorders are complex disorders consisting of both genetic and social factors, with a developmental component strongly linked to adult illness (Fig. 1). This means that integrated research into their aetiology, which includes genetic, biological, developmental and social perspectives, is now realistic. However, many difficulties remain – not least the problem of defining these disorders in a way that reflects their underlying features. This approach is beginning to take effect, with the recognition that restricting anorexia may be a distinct illness with its own biological basis. However, many commonalities between anorexia and bulimia remain, including risk factors that appear to be common to all such diagnoses. Despite this progress, the study of gene–environment interaction has barely been touched to date. There are many questions that remain to be answered regarding the aetiology of eating disorders. Which biological pathways are involved? Can a set of valid diagnoses reflecting underlying aetiology be constructed? Why are those affected predominantly women? How can the dual influence of genes and societal pressures be successfully reconciled? What is the exact nature of the developmental component? How can treatment be improved? The answers to these questions should become apparent in the coming decade.
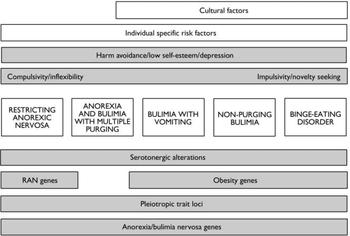
Fig. 1 Empirical structure of eating disorder. Cultural factors include pervasive factors such as diet and cultural attitudes to weight and shape. Cognitive style and personality are influenced by both genes and environment. Correlations exist between biological factors (serotonin function), environment (adverse childhood experiences), personality (e.g. impulsivity) and affect. All of these factors are influenced and mediated by genes. RAN, restricting anorexia nervosa.
DECLARATION OF INTEREST
None.


eLetters
No eLetters have been published for this article.