



1. Introduction
Development is the process through which reproducible forms unfold from a genetic blueprint. This reproducibility in the face of natural genetic and environmental variation is often explained as a buffering of the developmental process. Waddington used the concept of canalisation to explain the general observation that wild-type forms are better buffered to variation than mutant forms (Waddington, Reference Waddington1942). Underpinning this concept is the control of developmental programmes by networks of genes connected by regulatory interactions. Canalisation is an emergent property of such complex gene regulatory networks (Siegal & Bergman, Reference Siegal and Bergman2002).
The reproducible form of Arabidopsis (Arabidopsis thaliana) flowers is controlled by the combinatorial action of MADS-box transcription factors acting in specific whorled domains in the floral meristem (Coen & Meyerowitz, Reference Coen and Meyerowitz1991). According to the floral quartet model, petals form on the floral meristem via the activity of a tetrameric protein complex containing APETALA1 (AP1), APETALA3, PISTILLATA and SEPALLATA MADS-box transcription factors (Honma & Goto, Reference Honma and Goto2001). The combination of these transcription factors activates a petal-specific gene regulatory network that is itself embedded in a larger network controlling flower formation (Chen et al., Reference Chen, Yan, Fu and Kaufmann2018). Regulatory interactions within this gene network ensure that four petals form reproducibly in Arabidopsis flowers in spite of natural genetic variation (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Therefore, petal number is robust in Arabidopsis.
By contrast, petal number is decanalised in a close relative of Arabidopsis called Cardamine hirsuta. Individual flowers on a single plant display a variable number of petals between zero and four, and this phenotype varies in response to natural genetic and environmental variation (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017; Monniaux et al., Reference Monniaux, Pieper and Hay2016; Pieper et al., Reference Pieper, Monniaux and Hay2016). Therefore, petal number is not robust in C. hirsuta. This loss of robustness was traced to divergence of the MADS transcription factor AP1, which relaxed its epistasis over quantitative trait loci (QTL) that cause petal number to vary (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Previous work showed that transferring the Arabidopsis AP1 genomic locus into C. hirsuta was sufficient to canalise petal number by masking the phenotypic effects of at least nine QTL in the C. hirsuta genome (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Therefore, AP1 divergence in C. hirsuta altered its epistatic interactions with genes in the floral regulatory network and was the likely mechanism by which cryptic variation was exposed in C. hirsuta, contributing to the evolution of variable petal number.
Epistasis plays a large role not only in trait evolution, but also in domestication and breeding. Genomes contain vast stores of potential cryptic variants, whose phenotypic effects are only exposed after mutating genes in epistatic interactions with them (Bergman & Siegal, Reference Bergman and Siegal2003; Paaby & Rockman, Reference Paaby and Rockman2014). The exposure and subsequent selection for or against cryptic variants have helped shape developmental traits during crop domestication and modern breeding (Soyk et al., Reference Soyk, Benoit and Lippman2020). For example, in tomato, phenotypic effects of cryptic alleles of MADS-box genes, with both negative and positive effects on yield, were exposed by epistatic interactions during breeding for jointless fruit (Soyk et al., Reference Soyk, Lemmon, Oved, Fisher, Liberatore, Park, Goren, Jiang, Ramos and van der Knaap2017, Reference Soyk, Lemmon, Sedlazeck, Jimenez-Gomez, Alonge, Hutton, Van Eck, Schatz and Lippman2019). Therefore, complex gene regulatory networks buffer developmental processes and modification of these regulatory interactions, through evolution, domestication or breeding, contributes to phenotypic variation.
Arabidopsis flowers achieve a reproducible number of floral organs through the precise control of gene expression domains in space and time (Neumann et al., Reference Neumann, Xu, Smaczniak, Schumacher, Yan, Bluthgen, Greb, Jonsson, Traas and Kaufmann2022; Refahi et al., Reference Refahi, Zardilis, Michelin, Wightman, Leggio, Legrand, Faure, Vachez, Armezzani and Risson2021). How such patterning processes function in decanalised C. hirsuta flowers to produce a variable number of petals is unclear. To gain insight into this process, we studied the activity of a patterning module comprising the growth-promoting hormone auxin and growth-repressing transcription factors CUP-SHAPED COTYLEDON1,2 (CUC1,2) that belong to the NAM-ATAF-CUC (NAC)-domain family (Aida et al., Reference Aida, Ishida, Fukaki, Fujisawa and Tasaka1997). This patterning module is broadly important for plant development, and functions in various developmental contexts and different species (Berger et al., Reference Berger, Harpaz-Saad, Brand, Melnik, Sirding, Alvarez, Zinder, Samach, Eshed and Ori2009; Bilsborough et al., Reference Bilsborough, Runions, Barkoulas, Jenkins, Hasson, Galinha, Laufs, Hay, Prusinkiewicz and Tsiantis2011; Furutani et al., Reference Furutani, Vernoux, Traas, Kato, Tasaka and Aida2004; Galbiati et al., Reference Galbiati, Sinha Roy, Simonini, Cucinotta, Ceccato, Cuesta, Simaskova, Benkova, Kamiuchi and Aida2013; Heisler et al., Reference Heisler, Ohno, Das, Sieber, Reddy, Long and Meyerowitz2005; Kierzkowski et al., Reference Kierzkowski, Runions, Vuolo, Strauss, Lymbouridou, Routier-Kierzkowska, Wilson-Sanchez, Jenke, Galinha and Mosca2019). In Arabidopsis flowers, CUC1,2 transcription factors establish boundaries that demarcate the positions of four petal primordia on the adjacent floral meristem (Baker et al., Reference Baker, Sieber, Wellmer and Meyerowitz2005), which are marked by the formation of auxin maxima (Chandler et al., Reference Chandler, Jacobs, Cole, Comelli and Werr2011; Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2012; Reference Lampugnani, Kilinc and Smyth2013; Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). How CUC transcription factors instruct the patterning of auxin maxima has been previously described in the context of leaf development (Bilsborough et al., Reference Bilsborough, Runions, Barkoulas, Jenkins, Hasson, Galinha, Laufs, Hay, Prusinkiewicz and Tsiantis2011; Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024; Kierzkowski et al., Reference Kierzkowski, Runions, Vuolo, Strauss, Lymbouridou, Routier-Kierzkowska, Wilson-Sanchez, Jenke, Galinha and Mosca2019). In C. hirsuta, CUC1 regulates the expression of auxin-related genes, including the direct transcriptional activation of WAG serine/threonine AGCVIII kinases that regulate the polarity of the plasma membrane-localised PIN-FORMED1 (PIN1) auxin transporter (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024). Polar auxin transport relies on the export of auxin out of cells by PIN transporters to create auxin asymmetries that are central to plant growth and development. Specifically, the convergence of PIN1 polarities create a maximum auxin response that can be read out as activation of DR5 or degradation of DII reporters (Brunoud et al., Reference Brunoud, Wells, Oliva, Larrieu, Mirabet, Burrow, Beeckman, Kepinski, Traas and Bennett2012; Ulmasov et al., Reference Ulmasov, Murfett, Hagen and Guilfoyle1997) and trigger outgrowth. Thus, through the direct regulation of auxin targets, the CUC1 transcription factor in C. hirsuta contributes to PIN1 repolarisation, enabling new PIN1 convergences that underlie organogenesis (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024).
In this study, we used genetics, time-lapse confocal imaging and quantitative analysis of early flower development (Rambaud-Lavigne & Hay, Reference Rambaud-Lavigne and Hay2020) to investigate CUC/auxin patterning of decanalised petal number in C. hirsuta. We found that CUC1,2 genes regulate petal number by forming boundary regions between sepals and promoting the convergence of PIN1 polarities to create auxin maxima in these boundaries. The positioning of auxin activity maxima in the sepal whorl, rather than in the adjacent whorl on the floral meristem, is a key difference to Arabidopsis. Therefore, the same patterning module is deployed differently to initiate petals in C. hirsuta versus Arabidopsis. Modifying cellular growth, by manipulating CUC activity or ambient temperature, altered the amount of space between sepals in which discrete auxin maxima formed to trigger petal initiation. In this way, growth is integral to the patterning process controlling petal initiation in C. hirsuta and easily influenced by environmental and genetic variation, resulting in variable patterning outputs.
2. Results
2.1. CUC genes regulate petal number in C. hirsuta by modifying growth and auxin patterning
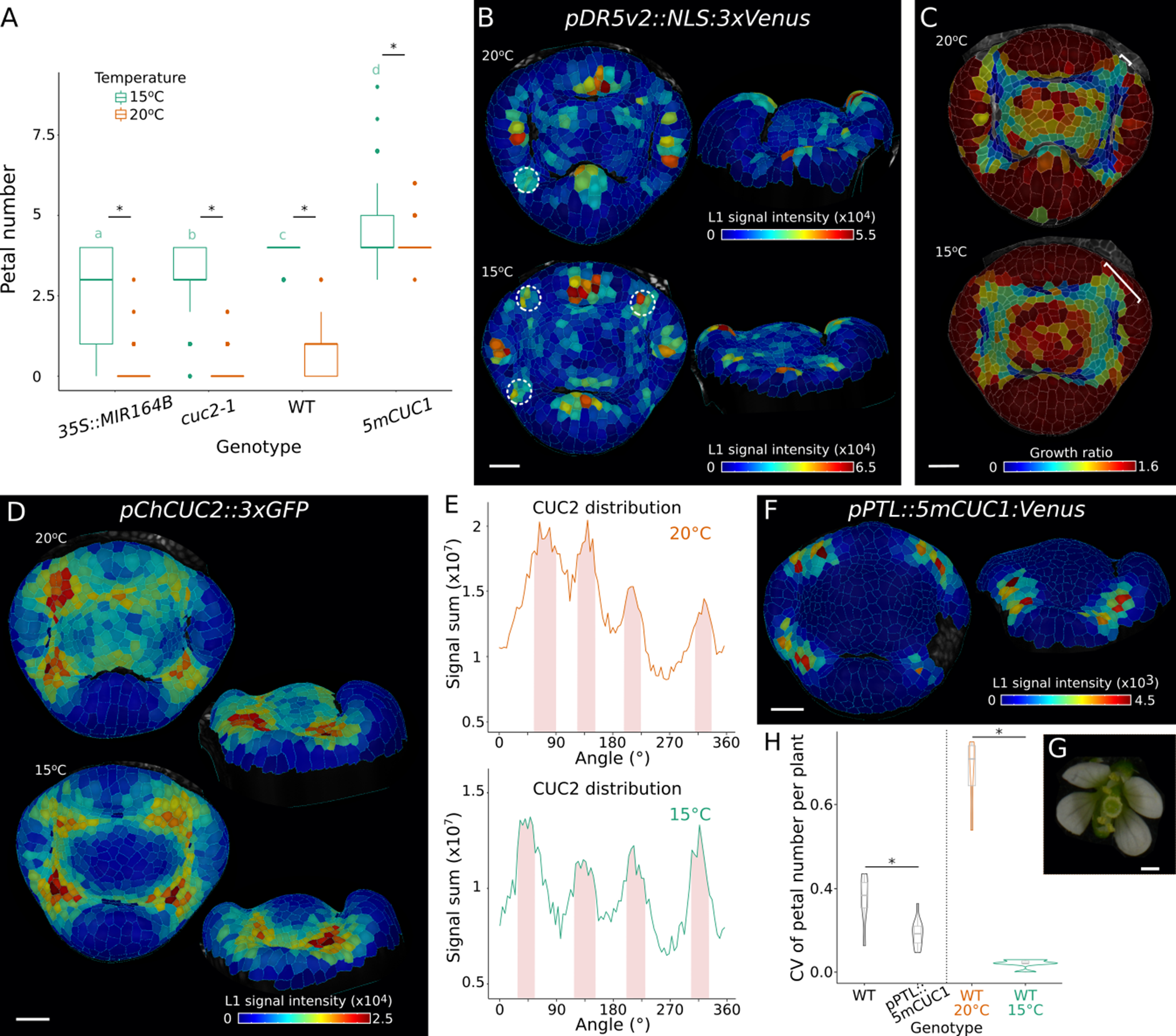
To investigate the role of CUC genes in regulating petal number in C. hirsuta, we analysed different genotypes with loss or gain of CUC function. In order to reduce CUC activity, we used cuc2-1 mutants (Rast-Somssich et al., Reference Rast-Somssich, Broholm, Jenkins, Canales, Vlad, Kwantes, Bilsborough, Dello Ioio, Ewing and Laufs2015) and the previously described 2x35S::MIR164B C. hirsuta line where overexpression of the Arabidopsis MICRORNA164B gene results in post-transcriptional degradation of both CUC2 and CUC1 (Blein et al., Reference Blein, Pulido, Vialette-Guiraud, Nikovics, Morin, Hay, Johansen, Tsiantis and Laufs2008). To increase CUC activity, we transformed C. hirsuta with 5mCUC1: a previously described miR164-resistant genomic clone of Arabidopsis CUC1 (Mallory et al., Reference Mallory, Dugas, Bartel and Bartel2004). Reduced CUC activity resulted in loss of petals in both cuc2-1 and 2x35S::MIR164B flowers (Figure 1a,b). Adjacent sepals were fused to varying extents in 2x35S::MIR164B (Supplementary Figure S1), but not cuc2-1 flowers. Cardamine hirsuta 5mCUC1 flowers with increased CUC1 activity had significantly more petals than the wild type (Figure 1a,b). 5mCUC1 flowers had a median petal number of four and up to six petals were occasionally observed (Figure 1a,b). These phenotypes are consistent with previous observations in Arabidopsis (Laufs et al., Reference Laufs, Peaucelle, Morin and Traas2004; Mallory et al., Reference Mallory, Dugas, Bartel and Bartel2004), and indicate that the variable petal number in wild-type C. hirsuta can be modulated by altering the level of CUC activity.
CUC activity regulates petal number in Cardamine hirsuta by modifying patterns of growth and auxin maxima. (a) Representative inflorescence (top) and flower (bottom) of C. hirsuta 2x35S::miR164b, cuc2-1, WT and 5mCUC1. (b) Boxplots of average petal number in C. hirsuta 2x35S::miR164b (n = 84 flowers, four plants), cuc2-1 (n = 127 flowers, six plants), WT (n = 38 flowers, two plants) and 5mCUC1 (n = 104 flowers, five plants). Petal number differs significantly between genotypes (Welch one-way ANOVA, p = 3.56e−103), and different letters denote statistical significance at p < 0.05 using a Games–Howell pairwise comparison as post hoc analysis. (c-d) Representative floral primordia surface reconstructions of C. hirsuta 2x35S::miR164b (n = 9), WT (n = 30) and 5mCUC1 (n = 7) (top and side view projections) showing cell area extension (heat map:cell growth ratio) during 10 h of growth (c) and DR5v2::NLS:3xVenus signal (heat map:average epidermal cell signal intensity in arbitrary units) at floral stage 4 (d). Arrows in (c) point to one inter-sepal boundary in each image, in top and side views. White dashed circles in (d) indicate DR5 expression maxima in inter-sepal boundaries. (e-h) Radial quantification of DR5 signal (sum of epidermal signal intensity in arbitrary units) in stage 4 floral primordia of C. hirsuta WT (e, f), 2x35S::miR164b (g) and 5mCUC1 (h). In grey: 95% confidence interval. Pink shading: petal regions at approximately 45°, 135°, 225° and 315°. Plots in (e, g, h) correspond to the samples shown in (d), and in (f) to the mean of 10 WT samples. Scale bars: 2 mm (a: inflorescences), 1 mm (a: flowers), 20 μm (c, d).

To investigate the effects of CUC activity on petal number, we considered the dual role of CUC genes to both specify boundary regions and to promote organogenesis via the formation of auxin maxima. We observed that boundary regions between sepals were reduced in 2x35S::MIR164B and enlarged in 5mCUC1 flowers, when compared to wild-type flowers (Figure 1c and Supplementary Figure S1). To quantify these phenotypes at a cellular level, we used time-lapse confocal imaging and MorphoGraphX software (Barbier de Reuille et al., Reference Barbier de Reuille, Routier-Kierzkowska, Kierzkowski, Bassel, Schupbach, Tauriello, Bajpai, Strauss, Weber and Kiss2015) to compute the amount of cellular growth that occurred during 10 h of development in stage 4 flowers (Figure 1c). In 2x35S::MIR164B flowers, we found that cellular growth was increased in the sepal whorl, particularly in the regions between sepals, resulting in overgrowth of the inter-sepal boundaries (Figure 1c). Conversely, cellular growth was reduced in broad regions between sepals in 5mCUC1 flowers, creating larger inter-sepal boundaries and narrower sepals (Figure 1c). Thus, altering the level of CUC activity influences the amount of space available between sepals for petals to form in C. hirsuta flowers.
We have previously shown that peaks of auxin activity maxima associated with initiating petals are often absent from the floral meristem, but are rather located in the inter-sepal boundaries of C. hirsuta flowers (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). This is in stark contrast to Arabidopsis where these auxin activity peaks are always located on the floral meristem (Figure 4b) (Chandler et al., Reference Chandler, Jacobs, Cole, Comelli and Werr2011; Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2012; Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013; Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). To investigate whether the formation of auxin activity maxima varies in response to CUC activity in C. hirsuta flowers, we analysed a DR5v2::nls:3xVenus (DR5) reporter in our time-lapse series (Figure 1d). In 5mCUC1 flowers, we consistently observed discrete foci of DR5 expression in each of the four inter-sepal boundaries (Figure 1d). On the other hand, foci of DR5 expression were absent from inter-sepal boundaries of 2x35S::MIR164B flowers (Figure 1d). To visualise DR5 distribution more quantitatively, we plotted DR5 signal intensity radially in the approximately circular floral primordia (Figure 1e–h). The most intense DR5 peaks are at sepal tips in all flowers, and petal initiation regions lie between these peaks (pink shading, Figure 1e–h). In wild-type flowers, weak DR5 signal of varying intensity was observed between sepals (Figure 1e), but these weak, variable peaks were barely evident when the signal was averaged across 10 different samples (Figure 1f). DR5 peaks were absent from the inter-sepal boundaries of 2x35S::MIR164B flowers (Figure 1g), whereas strong peaks of DR5 signal were present between each sepal in 5mCUC1 flowers (Figure 1h). Therefore, gain or loss of CUC activity is associated with the presence or absence of auxin activity peaks in inter-sepal boundaries. In summary, CUC1,2 genes regulate C. hirsuta petal number by specifying slow-growing boundaries between sepals that are competent to form auxin maxima.
2.2. Ambient temperature regulates petal number in C. hirsuta
We have previously shown that petal number in wild-type C. hirsuta is regulated by differential growth and maturation of floral buds in response to ambient temperature (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017). Slower growth and prolonged maturation time at lower temperatures produced larger boundaries between sepals and more petals (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017). Since increased CUC activity also created larger inter-sepal boundaries and more petals (Figure 1), we investigated the interaction between CUC genes and ambient temperature. When we grew CUC loss- and gain-of-function genotypes at 15°C, we found a significant increase in petal number in all genotypes, compared to plants grown at 20°C (Figure 2a). In particular, cuc2-1 and 2x35S::MIR164B flowers produced a median number of three petals at 15°C compared to zero petals at 20°C (Figure 2a). This suggests that the growth and maturation processes regulated by ambient temperature are not dependent on CUC activity. In line with this conclusion, significant differences in petal number between all CUC genotypes were observed at both 15°C and 20°C (Figure 2a).
Ambient temperature regulates petal number independent of CUC activity in Cardamine hirsuta. (a) Boxplots of average petal number in C. hirsuta 2x35S::miR164b, cuc2-1, WT and 5mCUC1 grown at 15°C (green) and 20°C (orange). At 15°C, 2x35S::miR164b (n = 187 flowers from 8 plants), cuc2-1 (n = 150 flowers from 6 plants), WT (n = 150 flowers from 6 plants), 5mCUC1 (n = 242 flowers from 10 plants); at 20°C the same data as in Figure 1b was used. Two-way ANOVA shows that petal number differs significantly between genotypes (*, p = 2.04e−223), growth temperatures (p = 4.56e−178) and genotype:temperature interactions (p = 1.90e−78); different letters denote statistical significance at p < 0.05 for genotypes grown at 15°C using Tukey’s HSD test as post hoc analysis. (b, d) Expression of pDR5v2::NLS:3xVenus (b, n = 11 at 20°C, n = 6 at 15°C) and pChCUC2::3xGFP (d, n = 2 at 20°C, n = 3 at 15°C) in representative floral primordia of C. hirsuta grown at 20°C and 15°C (top and side view projections). Heat maps show average epidermal cell signal intensity in arbitrary units. White dashed circles in (b) indicate DR5 expression maxima in inter-sepal boundaries. (c) Maps of cell area extension on representative floral primordia surface reconstructions of C. hirsuta WT grown at 20°C and 15°C (heat map: cell growth ratio, n = 30 at 20°C, n = 17 at 15°C) during 14 and 21 h of growth respectively. Brackets indicate the width of one inter-sepal boundary in each image. (e) Radial quantification of ChCUC2 signal in samples shown in (d). Pink shading: petal regions at approximately 45°, 135°, 225° and 315°. (f) Expression of pPTL::5mCUC1:Venus in representative floral primordia of C. hirsuta (n = 2). Heat maps show average epidermal cell signal intensity in arbitrary units. (g) Representative flower of C. hirsuta expressing pPTL::5mCUC1:Venus. (h) Violin plots of coefficients of variation (CV) of petal number per plant in C. hirsuta pPTL::5mCUC1:Venus (n = 330 flowers, 15 plants) and in the corresponding WT (n = 128 flowers, 6 plants) and in WT grown at 20°C (n = 78 flowers, 4 plants) or 15°C (n = 150 flowers, 6 plants). CV differs significantly between genotypes (Wilcoxon test: WT-pPTL::5mCUC1:Venus p = 0.0154; WT20°C–WT15°C p = 2.02e−46), * statistical significance at p < 0.05. Scale bars: 20 μm (b, c, d, f), 0.5 mm (g).

We reasoned that the increase in wild-type petal number at 15°C might be associated with more regular formation of DR5 peaks between sepals, similar to our findings in 5mCUC1 flowers (Figure 1d,h). To investigate this, we used time-lapse confocal imaging to compare DR5 expression in stage 4 flowers of plants grown at 15°C or 20°C (Figure 2b). We observed more discrete foci with higher DR5 signal intensity in inter-sepal boundaries at 15°C (Figure 2b). We also measured the amount, duration and distribution of growth in these flowers (Figure 2c). The duration of growth between similar developmental stages was much longer at 15°C, indicating slower growth, and cells with low amounts of growth were distributed in broader regions between sepals at 15°C compared to 20°C (Figure 2c), matching previous results (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017). Therefore, DR5 peaks form more consistently in the larger inter-sepal regions of flowers grown at 15°C, similar to 5mCUC1 flowers.
To determine the pattern of CUC2 transcription in C. hirsuta flowers, and whether it is altered by ambient temperature, we analysed stage 4 flowers of a C. hirsuta pChCUC2::3×GFP transcriptional reporter (ChCUC2) grown at 20°C and 15°C (Figure 2d,e). At this stage, ChCUC2 is expressed between sepals and excluded from sepals (Figure 2d). At 20°C, the intensity of ChCUC2 expression varied between different inter-sepal regions (Figure 2e), and showed a broad, background level of expression throughout the floral meristem (Figure 2d). At 15°C, the signal intensity of ChCUC2 was more similar between each sepal (Figure 2e), with maximum intensity at the boundary between the sepal whorl and the meristem (Figure 2d). ChCUC2 expression was absent from the floral meristem at 15°C, creating a sharper differential between the expression in inter-sepal boundaries and lack of expression throughout the rest of the floral primordium (Figure 2d,e).
Altogether, these results indicate that DR5 and ChCUC2 resolved to expression domains with less overlap in the larger inter-sepal boundaries formed in C. hirsuta flowers at 15°C compared to 20°C. Within each inter-sepal boundary, ChCUC2 expression was focused adjacent to the floral meristem (Figure 2d) while DR5 maxima formed towards the periphery of the flower (Figure 2b). These expression patterns were associated with a significantly lower coefficient of variation for wild-type petal number at 15°C compared to 20°C (Figure 2h). Therefore, more space between sepals seems to allow more reproducible patterning of petal initiation.
To test whether increasing CUC activity specifically in inter-sepal boundaries can reduce the variability in C. hirsuta petal number, we generated pPTL::5mCUC1 transgenics. We had previously shown that an Arabidopsis PETALLOSS (PTL) transgene expressed specifically in inter-sepal boundaries of C. hirsuta flowers (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Therefore, we used this PTL promoter to ectopically express 5mCUC1 (Figure 2f). In plants grown at 20°C, petal number was increased in pPTL::5mCUC1 flowers (Figure 2g and 3.4 ± 0.8 petals vs. 2.3 ± 1.1 petals in WT, mean ± SD) and the coefficient of variation was significantly lower (Figure 2h). Therefore, petal number variation was reduced by specifically increasing CUC1 activity in the inter-sepal domains of C. hirsuta flowers.
2.3. AUX1 and PIN1 localise to petal initiation regions in C. hirsuta flowers
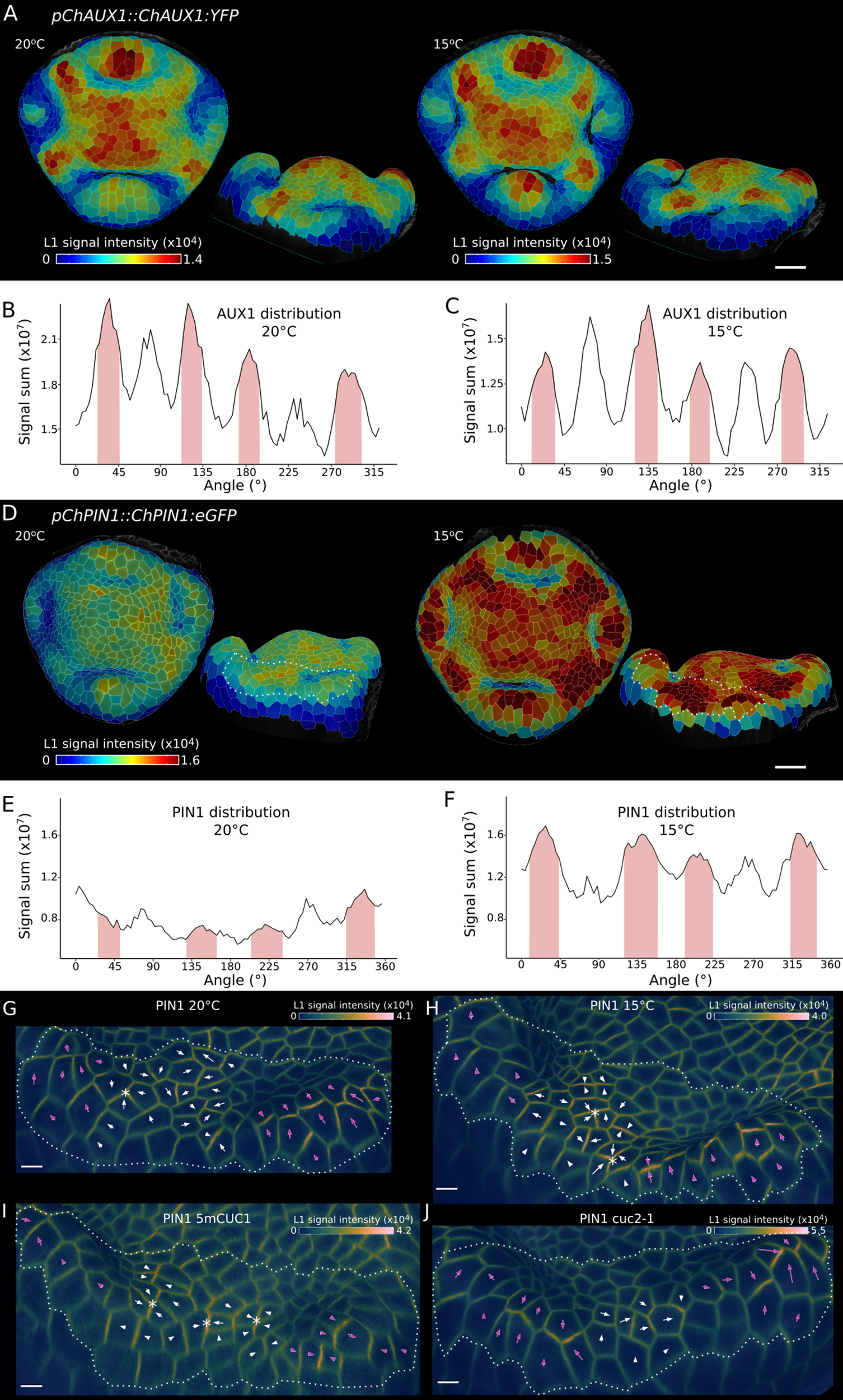
To further investigate the patterning of petal initiation by auxin in C. hirsuta flowers, we analysed expression of the auxin efflux protein PIN1 and the auxin influx protein AUX1 (Bennett et al., Reference Bennett, Marchant, Green, May, Ward, Millner, Walker, Schulz and Feldmann1996; Galweiler et al., Reference Galweiler, Guan, Muller, Wisman, Mendgen, Yephremov and Palme1998). Both proteins had been previously shown to play a role in the local accumulation of auxin that triggers petal initiation in Arabidopsis (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). To localise PIN1, we used a previously described C. hirsuta pChPIN1::ChPIN1:eGFP (ChPIN1) fusion protein that complements C. hirsuta pin1 mutants (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024). To localise AUX1, we generated a functional C. hirsuta pChAUX1::ChAUX1:YFP116 (ChAUX1) fusion protein with an in-frame fusion of YFP at amino acid 116 of ChAUX1, as described for Arabidopsis AUX1–YFP fusions (Swarup et al., Reference Swarup, Kargul, Marchant, Zadik, Rahman, Mills, Yemm, May, Williams and Millner2004). We quantified cellular signal of ChAUX1 and ChPIN1 to compare expression at the tissue level in stage 4 flowers of wild-type plants grown either at 20°C or 15°C (Figure 3a–f). At this stage, ChAUX1 is expressed in inter-sepal regions, where petals initiate, as well as the tips of medial sepals and the floral meristem (Figure 3a). Ambient temperature had little effect on ChAUX1 expression (Figure 3a–c). In contrast, ChPIN1 expression was increased at 15°C with the most intense signal in inter-sepal regions (Figure 3d–f). Therefore, both proteins are expressed at the right time and place to potentially contribute to petal initiation in C. hirsuta, and increased ChPIN1 expression was associated with the increased petal number observed at 15°C.
AUX1 and PIN1 localise to petal initiation regions in Cardamine hirsuta flowers. (a, d) pChAUX1::ChAUX1:YFP116 (a) and pChPIN1::ChPIN1:eGFP (d) expression in representative floral primordia of C. hirsuta grown at 20°C (a: n = 1, d: n = 5) and 15°C (a: n = 4, d: n = 4), shown in top and side view projections. Heat maps show average epidermal cell signal intensity in arbitrary units. Segmentations based on AUX1 signal at the plasma membrane rather than PI staining in (a). White dashed lines in (d) outline the inter-sepal regions shown in (g) and (h). (b-c, e-f) Radial quantification of pChAUX1::ChAUX1:YFP116 (b, c) and pChPIN1::ChPIN1:eGFP (e, f) signal (sum of epidermal signal intensity in arbitrary units) in C. hirsuta grown at 20°C (b, e) and 15°C (c, f). Petal regions are indicated on plots at approximately 45°, 135°, 225° and 315° (pink shading). (g-j) Plasma membrane-localised signal of pChPIN1::ChPIN1:eGFP in inter-sepal regions of representative C. hirsuta samples grown at 20°C (g, n = 5) and 15°C (h, n = 4), and 5mCUC1 (i, n = 2) and cuc2-1 (j, n = 1) grown at 20°C (side view projections). Heat maps show epidermal signal intensity at the cell edges in arbitrary units. Arrows estimate cell polarity of the signal using vectors computed by MGX with arrowheads added manually to face the most intense signal. Pink arrows are in sepal domains and white ones are in inter-sepal domains. Scale bars: 20 μm (a, d), 5 μm (g–j).

Since PIN1 proteins transport auxin in a polar manner (Galweiler et al., Reference Galweiler, Guan, Muller, Wisman, Mendgen, Yephremov and Palme1998), we analysed the polarity of the plasma membrane localisation of ChPIN1 in inter-sepal boundary cells of the same stage 4 flowers described earlier (Figure 3g,h). We computed the direction of ChPIN1 polarity in each cell as previously described (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024), and indicated this direction using arrows to point to the most intense signal. Coordinated fields of ChPIN1 polarities pointed to the direction of sepal tips in all flowers (magenta arrows, Figure 3g–j and Supplementary Figure S2). Between these fields, we observed divergent ChPIN1 polarities (white arrows, Figure 3g–j) that occasionally formed convergence points (asterisks, Figure 3g–j). Such convergence points formed more frequently between sepals in wild-type flowers grown at 15°C compared to 20°C (asterisks, 15/16 boundaries formed convergence points at 15°C, Figure 3h, vs. 7/16 at 20°C, Figure 3g and Supplementary Figure S2). We also observed a higher frequency of ChPIN1 convergence points in the large inter-sepal regions of 5mCUC1 flowers (asterisks, Figure 3i, 8/8 boundaries formed convergence points, Supplementary Figure S2) and a lower frequency between sepals in cuc2-1 flowers (Figure 3j, 1/4 boundaries formed convergence points). Since auxin maxima are formed by the convergence of PIN1 polarities (Heisler et al., Reference Heisler, Ohno, Das, Sieber, Reddy, Long and Meyerowitz2005), these observations match the higher incidence of DR5 maxima between sepals in 5mCUC1 flowers (Figure 1d) and wild-type flowers with higher petal number at 15°C (Figure 2b). However, the observed ChPIN1 convergences in flowers with low petal number (Figure 3g,j) suggest that formation of a PIN1 convergence is not sufficient per se to form a stable peak of auxin activity that leads to petal initiation. In summary, higher petal number in 5mCUC1 and 15°C grown flowers was associated with more frequent formation of PIN1 convergence points and auxin maxima in larger inter-sepal boundaries.
2.4. Robust petal number in Arabidopsis
In comparison to C. hirsuta, petal number is canalised in Arabidopsis. To compare the localisation of CUC2, auxin activity maxima, PIN1 and AUX1 in this robust patterning system, we analysed stage 4 flowers of Arabidopsis. At this stage, the cellular signal of pAtAUX1::AtAUX1:YFP116 (AtAUX1) (Swarup et al., Reference Swarup, Kargul, Marchant, Zadik, Rahman, Mills, Yemm, May, Williams and Millner2004) was intense in inter-sepal boundaries, as previously reported (Supplementary Figure S3) (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). An Arabidopsis pAtCUC2::nls:tdTomato (AtCUC2) transcriptional reporter was expressed in inter-sepal boundaries and excluded from sepals (Figure 4a). In comparison to C. hirsuta, petal initiation sites marked by expression of the auxin activity reporter DR5v2::nls:3xVenus (DR5) were located on the floral meristem (Figure 4b). In this way, no overlap occurred between AtCUC2 transcription in inter-sepal boundaries and sites of petal initiation. We next investigated the expression and polarity of Arabidopsis pAtPIN1::AtPIN1:GFP (AtPIN1) associated with the formation of DR5 peaks on the floral meristem. We found the highest AtPIN1 signal located throughout the sepal whorl (Figure 4c). Coordinated fields of AtPIN1 polarities pointed to the direction of sepal tips (white arrows, Figure 4d). Immediately adjacent to the inter-sepal domains, we observed additional fields of AtPIN1 polarity running to the direction of the floral meristem, and therefore towards sites of petal initiation (grey arrows, Figure 4d).
Robust petal number in Arabidopsis. (a, c, h) Heat maps quantifying the epidermal signal in representative samples of pAtCUC2::NLS:tdTomato in WT (a, n = 10) and pAtPIN1::AtPIN1:GFP in WT (c, n = 4) and ptl-1 (h, n = 2) stage 4 floral primordia of Arabidopsis thaliana. Colour bars: epidermal signal intensity in arbitrary units. Arrows in (a) point to AtCUC2 expression in one inter-sepal boundary in top and side views. White dashed lines in (c) and (h) outline the inter-sepal regions shown in (d) and (i). (b, g) pDR5v2::NLS:3xVenus (green) expression in representative samples of WT (b, n = 6) and ptl-1 (g, n = 3) Arabidopsis flowers at stage 4; cells outlined with PI staining (red). Top and side views are shown, white dashed circles in side view (b) indicate DR5 expression maxima on the flanks of the floral meristem. (d, i) Projections of plasma membrane-localised signal of pAtPIN1::AtPIN1:GFP in inter-sepal regions of Arabidopsis WT (d) and ptl-1 (i), shown as side views. Heat maps show epidermal signal intensity at the cell edges in arbitrary units. Arrows estimate cell polarity of the signal using vectors computed by MGX with arrowheads added manually to face the most intense signal. White arrows point to sepal tips, grey arrows point to FM. (e) Representative inflorescences (top) and flowers (bottom) of Arabidopsis WT and ptl-1. (f) Boxplots of petal number in Arabidopsis WT (n = 150 flowers, six plants) and ptl-1 (n = 150 flowers, six plants). Letters denote statistically significant differences between means based on Welch one-way ANOVA (p = 1.68e-80). Scale bars: 20 μm (a, b, c, g, h), 5 μm (d, i), 2 mm (inflorescences) and 0.5 mm (flowers) (e). Image in (b) reproduced from Monniaux et al. (Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018).

Previous work had established that auxin is a mobile petal initiation signal in Arabidopsis (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). It had been shown that increasing auxin biosynthesis in the inter-sepal domain in ptl mutants was sufficient to restore petal initiation in the adjacent whorl on the floral meristem (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). Therefore, we investigated the localisation of AtPIN1 in the ptl-1 allele, which shows almost complete petal loss (Figure 4e,f). As previously described, the loss of petals in ptl-1 flowers was associated with the absence of DR5 maxima at petal initiation sites on the floral meristem (Figure 4g) (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). The AtPIN1 signal was reduced throughout the sepal whorl in ptl-1 compared to wild type and conspicuously low in the adjacent whorl where petals would normally form (Figure 4h). Coordinated fields of AtPIN1 polarities were visible in the sepal whorl, pointing to the direction of sepal tips (white arrows, Figure 4i). However, AtPIN1 signal was not visible in the adjacent whorl (Figure 4i). Therefore, ptl-1 did not show the fields of AtPIN1 polarity observed in wild-type flowers that putatively transport auxin from inter-sepal regions to sites of petal initiation on the floral meristem (Figure 4i).
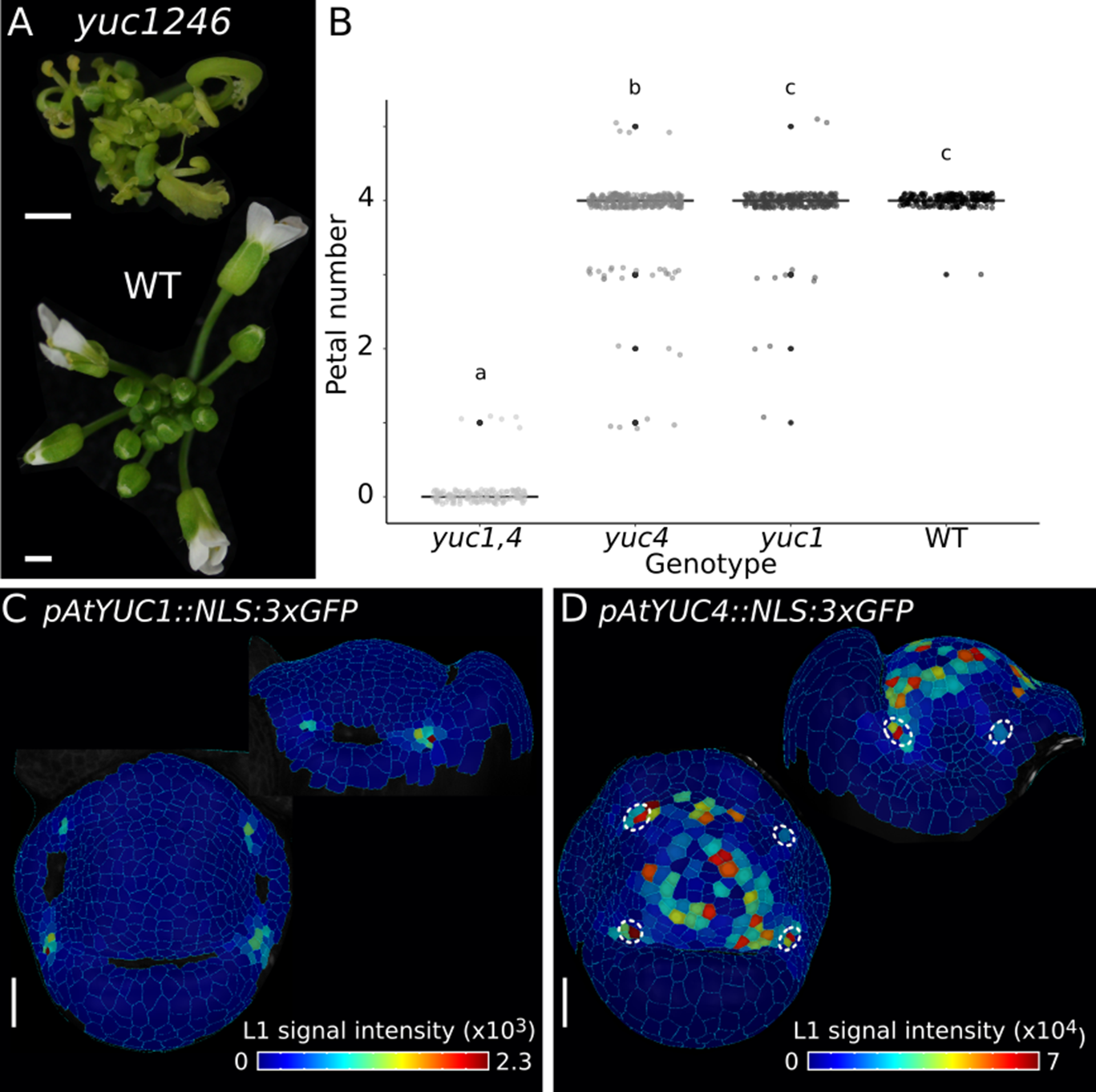
Previous studies have used the bacterial auxin biosynthesis gene iaaH under the control of the PTL promoter to restore petal initiation in ptl flowers (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). These experiments demonstrated the sufficiency of local auxin biosynthesis for petal initiation. However, it was not clear where endogenous auxin biosynthesis occurred in developing flowers. To investigate this question, we analysed YUCCA (YUC) genes, which are involved in the main auxin biosynthesis pathway (Cheng et al., Reference Cheng, Dai and Zhao2006; Reference Cheng, Dai and Zhao2007; Stepanova et al., Reference Stepanova, Robertson-Hoyt, Yun, Benavente, Xie, Dolezal, Schlereth, Jurgens and Alonso2008). Quadruple yuc1 yuc2 yuc4 yuc6 mutants show strong floral defects, including loss of petals (Figure 5a) (Cheng et al., Reference Cheng, Dai and Zhao2006). To assess the contribution of individual YUC genes to petal formation, we first imaged transcriptional reporters for YUC1, YUC4 (Zhang et al., Reference Zhang, Runions, Mentink, Kierzkowski, Karady, Hashemi, Huijser, Strauss, Gan and Ljung2020) and YUC2, YUC6 (Galvan-Ampudia et al., Reference Galvan-Ampudia, Cerutti, Legrand, Brunoud, Martin-Arevalillo, Azais, Bayle, Moussu, Wenzl and Jaillais2020). We failed to detect pYUC6::GFP expression in young flowers and found only faint expression of pYUC2::GFP in internal floral tissues near the pedicel (Supplementary Figure S4). In contrast to this, pYUC1::NLS:3xGFP (YUC1) expression was confined to a very specific domain in stage 4 flowers, consisting of a few cells between sepals (Figure 5c). At this stage, pYUC4::NLS:3xGFP (YUC4) was expressed more broadly in the floral meristem and excluded from the sepal whorl (Figure 5d). We observed high YUC4 expression adjacent to inter-sepal zones at sites of petal initiation on the floral meristem (Figure 5d). To assess whether these two genes regulate petal number, we analysed single and double yuc1 yuc4 mutants. We found no difference in petal number between wild-type and yuc1; a small, but significant reduction of petal number in yuc4 and almost complete loss of petals in yuc1 yuc4 double mutants (Figure 5b). Therefore, YUC1 and YUC4 act redundantly to control petal initiation. The expression domains of YUC1 and YUC4 suggest that two sites of local auxin production may be relevant for petal initiation – one defined by YUC1 expression in the inter-sepal boundary, and another site in the adjacent whorl defined by YUC4 expression. Therefore, based on genetic analysis and gene expression patterns, it is likely that YUC1 and YUC4 contribute to local auxin synthesis during petal initiation in developing flowers.
Auxin biosynthesis genes YUC1 and YUC4 contribute to petal number in Arabidopsis. (a) Representative inflorescences of Arabidopsis WT and yuc1246 mutant. (b) Boxplots of petal number in yuc1yuc4 (n = 137 flowers, 6 plants), yuc4 (n = 278 flowers, 12 plants), yuc1 (n = 261 flowers, 12 plants) and WT plants (n = 206 flowers, 9 plants). Letters denote statistically significant differences (p < 0.05) between means based on Welch one-way ANOVA (p = 0, Tukey’s HSD post hoc test). P values are 0.659 (Col-0-yuc1), 0 (Col-0-yuc1yuc4), 3.88e−4 (Col-0-yuc4), 0 (yuc1-yuc1yuc4), 1.34e−2 (yuc1-yuc4) and 0 (yuc1yuc4-yuc4). (c-d) Heat maps quantifying epidermal signal in representative samples of pAtYUC1::NLS:3xGFP (c, n = 5) and pAtYUC4::NLS:3xGFP (d, n = 4) in Arabidopsis stage 4 flowers (top and side view projections). Colour bars: epidermal signal intensity in arbitrary units. White dashed circles in (d) indicate YUC4 expression in petal initiation regions on the floral meristem. Scale bars: 1 mm (a), 20 μm (c, d).

In summary, invariant petal number in Arabidopsis is associated with clear patterning by CUC2 expression defining inter-sepal boundaries and the formation of auxin maxima in the adjacent whorl on the floral meristem. Auxin promotes petal initiation at these floral meristem sites in Arabidopsis (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013), and here we implicate polar auxin transport by PIN1, and local auxin biosynthesis by YUC1 and YUC4 in this process.
3. Discussion
Our analysis shows that CUC1,2 genes regulate petal initiation in C. hirsuta by creating boundaries of reduced cellular growth between sepals and promoting the convergence of PIN1 polarities to create auxin maxima in these boundaries. This positioning of auxin maxima between sepals, rather than in the adjacent whorl on the floral meristem, distinguishes petal formation in C. hirsuta from Arabidopsis. In C. hirsuta, the space available between sepals for discrete auxin maxima to form is critical for petal initiation. In this way, growth of the inter-sepal region is an integral part of the patterning process controlling petal initiation, and subject to variation by environmental and genetic factors that cause petal number to vary in C. hirsuta (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017; Pieper et al., Reference Pieper, Monniaux and Hay2016).
We have shown that increasing or decreasing CUC activity in C. hirsuta shifts the distribution of petal number to a higher or lower mean, respectively, than the wild type. This is different to the canalising effect of transferring AP1 from Arabidopsis into C. hirsuta, which produces invariant petal number (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). The effect of this Arabidopsis AP1 transgene is associated with an expanded expression domain that extends onto the floral meristem in C. hirsuta. In comparison, the endogenous domain of C. hirsuta AP1 expression is restricted to the sepal whorl and petal initiation occurs between sepals (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Yet, according to the ‘ABC’ model, petal formation requires not only AP1, but also ‘B’ genes, such as APETALA3 (AP3) and PISTILLATA, which are excluded from the sepal whorl in Arabidopsis (Coen & Meyerowitz, Reference Coen and Meyerowitz1991). We verified that a functional Arabidopsis AP3 fusion protein was restricted to the floral meristem in Arabidopsis, but extended out into inter-sepal regions in C. hirsuta flowers (Supplementary Figure S5), which matched the endogenous expression domain of C. hirsuta AP3 (Supplementary Figure S5). Therefore, a potential overlap exists between AP1 and AP3 expression in the inter-sepal boundaries where petals initiate in developing C. hirsuta flowers.
Our findings indicate that CUC genes play a dual role in promoting the competence of inter-sepal regions to give rise to petals in C. hirsuta. First, CUC genes create boundary regions of reduced cellular growth that maintain the competence to proliferate (Bhatia et al., Reference Bhatia, Wilson-Sanchez, Strauss, Vuolo, Pieper, Hu, Rambaud-Lavigne and Tsiantis2023; Vernoux et al., Reference Vernoux, Kronenberger, Grandjean, Laufs and Traas2000). This is a general feature of CUC action; for example, during early leaf development in C. hirsuta, CUC2 limits the growth, but not proliferation, of cell populations along the leaf margin to pattern leaflet boundaries (Bhatia et al., Reference Bhatia, Wilson-Sanchez, Strauss, Vuolo, Pieper, Hu, Rambaud-Lavigne and Tsiantis2023). Second, CUC activity promotes the convergence of PIN1 polarities, which are responsible for creating auxin maxima. For example, CUC1 acts in C. hirsuta leaves to repolarise PIN1 via the direct transcriptional activation of WAG kinases that phosphorylate PIN1 (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024). Testing whether CUC1 regulates PIN1 polarity via the same mechanism in C. hirsuta flowers will be an important follow-up to this study.
The positioning of auxin maxima in inter-sepal boundaries is likely to create conflicts that impair petal initiation in C. hirsuta. On the one hand, auxin maxima trigger organogenesis, but on the other hand, CUC activity specifies boundaries that should be devoid of organ formation. Similar conflicts were proposed to contribute to the failure of pin1 inflorescences to initiate flowers. Naked pin1 apices were shown to express early markers of organ initiation, such as AINTEGUMENTA and LEAFY, together with CUC2, creating a hybrid organ/boundary identity at the meristem periphery (Vernoux et al., Reference Vernoux, Kronenberger, Grandjean, Laufs and Traas2000). We found that petal initiation was more likely to succeed in C. hirsuta when larger boundary regions were created between sepals, either by slow growth and prolonged maturation at low ambient temperatures or increased CUC1 activity in these regions. We observed the convergence of PIN1 polarities more frequently and more regular spatial patterning of auxin maxima in these larger inter-sepal regions. This suggests that organ versus boundary identities may have the space to resolve with less overlap in a larger tissue context. Moreover, we found that the effect of low ambient temperature to increase petal number and reduce its variability was independent of CUC1,2 gene activity. Therefore, additional factors that contribute to auxin patterning may have more space to resolve in larger inter-sepal regions.
A key feature of robust petal initiation in Arabidopsis is the formation of auxin maxima on the floral meristem. It was previously shown that auxin is a mobile signal for petal initiation and that factors in inter-sepal regions, such as AUX1 and PTL, influence the availability of auxin to accumulate in the adjacent whorl (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013). Our findings indicate how polar transport and local synthesis of auxin may contribute to this process. We observed a divergent field of PIN1 polarity in Arabidopsis flowers that may provide a putative transport route for auxin from the inter-sepal region to the adjacent whorl on the floral meristem. This PIN1 localisation was absent from the floral meristem in C. hirsuta flowers and in ptl-1 mutants that lacked petals. Based on previous work (Hu et al., Reference Hu, Wilson-Sanchez, Bhatia, Rast-Somssich, Wu, Vlad, McGuire, Nikolov, Laufs and Gan2024), we hypothesise that CUC may instruct the repolarisation of PIN1 in inter-sepal boundaries of Arabidopsis flowers to initiate new auxin fluxes towards petal initiation sites on the floral meristem. Testing this hypothesis may help to further understand how auxin controls robust petal initiation in Arabidopsis.
CUC activity has also been shown to influence the expression of YUCCA auxin biosynthetic genes (Abley et al., Reference Abley, Sauret-Gueto, Maree and Coen2016; Yamada et al., Reference Yamada, Tanaka, Miyazaki and Aida2022). Our genetic analysis indicates that YUC1 and YUC4 contribute to petal initiation in Arabidopsis. YUC1 expression suggests that auxin is likely synthesised in inter-sepal regions, which may provide a local source of auxin for polar transport to petal initiation sites via PIN1. Furthermore, the expression of YUC1 in inter-sepal regions of developing flowers also matched previous reports for the auxin biosynthetic genes TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 (TAA1) and TAR2 (Yadav et al., Reference Yadav, Kumar, Mahajan, Sahu, Singh and Yadav2023). On the other hand, YUC4 expression suggests that auxin is also likely to be synthesised locally at petal initiation sites on the floral meristem. Therefore, auxin synthesised at a distance in inter-sepal regions and also locally on the floral meristem are both likely to contribute to the formation of auxin maxima for petal initiation in Arabidopsis flowers. Comparing the function and distribution of YUCCA genes in C. hirsuta flowers may help to further understand the variable initiation of petals in this species.
In summary, decanalised petal number in C. hirsuta is associated with a shift in petal initiation events from the floral meristem to inter-sepal boundaries. This is underpinned by AP1 gene divergence in C. hirsuta, which resulted in loss of AP1 expression from the floral meristem and its loss of epistasis over petal number QTL (Monniaux et al., Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018). Attempting to initiate petals in boundary regions likely exposed this developmental process to the influence of many genetic loci that are otherwise cryptic in a robust system. Identifying the genes underlying petal number QTL (Monniaux et al., Reference Monniaux, Pieper and Hay2016; Reference Monniaux, Pieper, McKim, Routier-Kierzkowska, Kierzkowski, Smith and Hay2018; Pieper et al., Reference Pieper, Monniaux and Hay2016), and studying the natural variation and genetic basis for petal number plasticity in response to temperature, will be important avenues of future research to gain insight into these questions.
4. Materials and methods
4.1. Accessions and plant material
Plant material and accessions used in this study are described in Table 1. ap3-7 is a Col-0 allele identified in seed stock CS6565 with a GT>GA mutation in the splice site of intron 4. The described mutation for ap3-6 was not present in this stock. Therefore, we named the allele ap3-7. Genotyping primers f324/f325 were used to amplify a 342-bp fragment that is cut by MboI in the ap3-7 mutant, and remains uncut in the wild type. yuc4-1 yuc1 double mutants were identified from selfed yuc4-1 yuc1/+ plants using genotyping primers lr063/lr064, lr066/lr067, LBb1.1 and Spm32.
Plant material used in this study.

4.2. Transgenic plant construction
The wild-type genotype in C. hirsuta is the reference Oxford (Ox) accession, herbarium specimen voucher Hay 1 (OXF) (Hay & Tsiantis, Reference Hay and Tsiantis2006) and in Arabidopsis, the reference Col-0 accession. All plasmids were transformed into wild-type plants by Agrobacterium tumefaciens (strains GV3101 or C58)-mediated floral dip (Clough & Bent, Reference Clough and Bent1998).
The pChAUX1::ChAUX1:YFP116 plasmid contains an in-frame fusion of YFP after amino acid 116 of ChAUX1, as described for functional Arabidopsis AUX1-YFP fusions (Swarup et al., Reference Swarup, Kargul, Marchant, Zadik, Rahman, Mills, Yemm, May, Williams and Millner2004). A 3.4-kb ChAUX1 promoter sequence (3.4 kb before ATG) was amplified from C. hirsuta genomic DNA with primers m272/m434, digested with SalI/XmaI and subcloned into pBJ36. A 450-bp sequence of the 5′ end of gChAUX1 (starting at ATG) was amplified with primers m276/m430, and YFP coding sequence (715 bp) was amplified with primers m431/m432. Both fragments were fused by overlapping PCR so that YFP was inserted after amino acid 116 of gChAUX1. This fragment was fused by overlapping PCR with a 2.8-kb sequence of the 3′ end of gChAUX1 (amplified with primers m433/m277). The whole fragment was subcloned into pBJ36 by XmaI/BamHI digestion and validated by sequencing. Functionality of the construct was tested by complementing Arabidopsis aux1-22 mutants: five independent lines had a single transgene copy (IDna Genetics, Norwich, UK) and rescued the root gravitropism phenotype of aux1-22 seedlings. This functional construct was transformed into C. hirsuta.
The following three constructs were generated by GreenGate cloning (Lampropoulos et al., Reference Lampropoulos, Sutikovic, Wenzl, Maegele, Lohmann and Forner2013). pAtPTL::5mCUC1:Venus: a 2.8-kb pAtPTL promoter sequence (consisting of 1.3 kb before the ATG, the 5′UTR, the first exon and intron and eight codons of the second exon), that drives functional PTL expression (Lampugnani et al., Reference Lampugnani, Kilinc and Smyth2013), was amplified from Arabidopsis genomic DNA. 5mCUC1 was amplified from pCUC1::5mCUC1 plasmid (Mallory et al., Reference Mallory, Dugas, Bartel and Bartel2004). Entry vectors were cloned into the binary vector pGGZwf01 (Perez-Anton et al., Reference Perez-Anton, Schneider, Kroll, Hofhuis, Metzger, Pauly and Hay2022). pAtAP3::AtAP3:GFP: a 696-bp AtAP3 sequence was amplified from Arabidopsis cDNA as two fragments with primers m478/m472 and m476/m479 to mutate an internal BsaI site. This entry vector was cloned with pGGA002 (Addgene plasmid #48813; Lampropoulos et al., Reference Lampropoulos, Sutikovic, Wenzl, Maegele, Lohmann and Forner2013), which contains a 1.269-kb pAtAP3 promoter sequence, into the binary vector pGGZ003. Fifteen independent lines were generated in C. hirsuta and 24 in Arabidopsis ap3-7. Lines that complemented the sterility of homozygous ap3-7 mutants were analysed for transgene copy number (IDna Genetics, Norwich, UK) and three lines with either one or two transgene copies were imaged in T2 and T3 plants. pChAP3::ChAP3:GFP: a 927-bp pChAP3 promoter sequence was amplified from C. hirsuta genomic DNA with primers m469/m470. A 696-bp ChAP3 sequence was amplified from C. hirsuta cDNA as two pieces with primers m480/m475 and m476/m481 to remove an internal BsaI site. Entry vectors were cloned into the binary vector pGGZ003. Functionality of the construct was tested by complementing Arabidopsis ap3-7 mutants. This functional construct was transformed into C. hirsuta, six independent lines were analysed for transgene copy number (IDna Genetics, Norwich, UK) and three lines with either one or two transgene copies were imaged in T2 and T3 plants.
The pChCUC2::3xGFP:3 ′ plasmid contains 2.6-kb C. hirsuta CUC2 promoter sequence (2.6 kb before ATG) and 2.6-kb 3′ sequence (2.6 kb after STOP) amplified from C. hirsuta genomic DNA and fused with 3xGFP. Fragments were subcloned into pBS KS+ as follows: 2.6-kb 5′ CUC2 promoter in SacI and BamHI sites, 3xGFP in BamHI and PstI sites, and 2.6-kb 3′ CUC2 in PstI and SalI sites; then amplified using primers with NotI overhangs and cloned into pMLBART. The pAtCUC2::NLS:tdTomato reporter was generated by fusing a 3.2-kb Arabidopsis CUC2 promoter sequence (3.2 kb before ATG), amplified from pAtCUC2::AtCUC2:Venus plasmid (Heisler et al., Reference Heisler, Ohno, Das, Sieber, Reddy, Long and Meyerowitz2005), to tdTomato and cloned into pMLBART-hyg.
All primers used in this study are listed in Table 2.
4.3. List of primers used.
Primers used in this study.

4.4. Plant growth conditions and petal number scoring
Seeds were sown on soil, stratified 10 days in the dark at 4°C, germinated for 7 days in short days (SD) and transferred to long-day (LD) conditions in the greenhouse. SD: 8 h light (22°C), 16 h dark (20°C), LD: 16 h light (22°C), 8 h dark (20°C). Plants grown at 15°C were sown on soil, stratified 10 days in the dark at 4°C and transferred into a growth cabinet with LD conditions: 16 h light (16°C, 70% relative humidity), 8 h dark (14°C, 70% relative humidity). Petal number was scored every second day in open flowers that were removed one by one with tweezers from the primary shoot. The first 25 flowers on the main shoot were scored. Due to the seasonal variability of petal number in C. hirsuta (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017), data presented in each petal number plot always come from a single batch of experiments.
4.5. Confocal laser scanning microscopy
Floral primordia were staged as previously described in Arabidopsis (Smyth et al., Reference Smyth, Bowman and Meyerowitz1990) and C. hirsuta (McKim et al., Reference McKim, Routier-Kierzkowska, Monniaux, Kierzkowski, Pieper, Smith, Tsiantis and Hay2017). Time-lapse imaging was performed using plants grown as described earlier. Upon bolting, primary inflorescence meristems were cut and dissected under water. Flowers were removed one by one until the inflorescence meristem and young floral primordia up to stage 3 were uncovered. Dissected shoots were transferred to ½ MS supplemented with 1.5% plant agar (Duchefa), 1% sucrose and 0.1% plant preservative medium (Plant Cell Technology), with pH adjusted to 5.7 with KOH. Shoots were oriented in order to lay the inflorescence meristem or the floral meristem of interest flat and fixed with a drop of warm 1% agarose. Cell walls were stained with 0.1% propidium iodide (PI, Sigma) for 5 min before each observation, then rinsed twice in H2O and immersed in H2O at least 10 min before observation in order to transiently stop growth that would impair the acquisition of large image stacks. Water-immersed meristems were imaged from the top, on the day of the dissection and 14, 24 and 38 h after the initial time point in order to image stages 3 to 6 of floral development. Between each acquisition, the samples were placed into a LD growth chamber (at standard 20°C temperature or at 15°C when required). Confocal imaging was performed using an upright Leica SP8 confocal laser scanning microscope equipped with a long working distance water immersion objective (HC FLUOTAR L 25x/0.95 W) (Leica) and HyD hybrid detectors (Leica). Excitation was achieved with OPS lasers at 488 nm for GFP, 514 nm for Venus and YFP and 552 nm for tdTomato and PI. The emission fluorescence was collected at 500–520 nm for GFP, 520–540 nm for Venus and YFP, 575–590 nm for tdTomato and 630–650 nm for PI. Unidirectional scans (16 or 12 bit) of a 1024 × 1024 pixel region of interest were acquired at 600 Hz with a pixel size of 150–200 nm and a z-step size of 500 nm. Line averaging was either absent or set to 2.
4.6. Quantitative image analysis
Confocal image stacks were analysed with MorphoGraphX software (Barbier de Reuille et al., Reference Barbier de Reuille, Routier-Kierzkowska, Kierzkowski, Bassel, Schupbach, Tauriello, Bajpai, Strauss, Weber and Kiss2015): image stacks were loaded and the sample surface was detected and reconstructed in a 2.5D object by generating a mesh. All top- and side-view snapshots of flower meristems show quantitative data projected on 2.5D meshes. To quantify gene expression, the epidermal (2–5 μm) signal was projected onto the reconstructed surface and a heat map of signal intensity (border or interior signal depending on the reporter) averaged per cell was generated.
To quantify signal intensities at different positions relative to the centre of the floral meristem, the stacks were manually aligned in MorphoGraphX in order to place the z-axis at the centre pointing upward, the x-axis representing the 0° position and pointing to the right. Angles increased counter clockwise, within the xy plane. All images were aligned the same way, with a lateral sepal at position 0°, the adaxial sepal at 90°, the second lateral sepal at 180° and the abaxial sepal at 270°. The function ‘Export Histogram Circular’ (in Stack -> Analyses) was used with the following parameters: bin number: 90, voxel value threshold %: 1, voxel distance min (μm): 0, voxel distance max (μm): 50 and weight by volume: yes. Cell signal intensities were exported as a csv file and imported into R for statistical analysis and visualisation.
To quantify cell growth with MorphoGraphX, we tracked cell lineages between two consecutive time points, computed growth ratios and displayed the data as a colour map on the second time point. In order to capture floral stage 4, samples grown at 20°C were imaged three times over 24 h of development and samples grown at 15°C were imaged three times over 43 h of development. Consecutive time points corresponding to a 10-h interval (Figure 1c), a 14-h interval (20°C, Figure 2c) or a 21-h interval (15°C, Figure 2c) were analysed.
To quantify PIN1 polarity with MorphoGraphX, the signal projected on the surface was colour coded based on its intensity (for visualisation) and the process ‘compute signal orientation’ was used. The displayed output consisted in min and max signal intensity axes drawn inside each segmented cell. Screenshots of regions of interest were taken with and without these axes being displayed. ImageJ (version 1.51i or earlier) was used to manually draw the max axis in cells of interest and to assign an arrowhead towards the edge with the most PIN1 expression based on visual comparison between the two edges of the computed max axis.
4.7. Statistical analysis
Petal number was statistically analysed with R Studio version 1.3.1093 or earlier (R Team, 2020). Throughout the article, average values are mean ± SD and boxplots show quartiles (box and whiskers), median (bold line), outliers (dots). A Welch one-way ANOVA followed by a Games–Howell post hoc test for pairwise comparisons was used in Figure 1b. A two-way ANOVA followed by a Tukey’s HSD test was used to test the influence of both temperature and genotype on petal number in Figure 2a. A Welch one-way ANOVA was used in Figure 4f and a one-way ANOVA followed by a Tukey’s HSD test was used for pairwise comparisons in Figure 5b. The coefficient of variation was calculated as the ratio of the standard deviation to the mean in Figure 2h, and a Wilcoxon rank sum test was used to compare two independent groups of samples.
4.8. Photographs of inflorescences and flowers
The photographs of inflorescences and individual flowers were taken with a Canon compact camera equipped with a macro lens EF 50 mm 1:25 and a lens macro 1:1 (Canon).
Open peer review
To view the open peer review materials for this article, please visit http://doi.org/10.1017/qpb.2025.10015.
Acknowledgements
We thank M. Tsiantis for comments and sharing materials; P. Laufs and T. Vernoux for sharing materials; R. Dello Ioio for generating 5mCUC1 C. hirsuta transgenic lines; W. Faigl for technical assistance; N. Bhatia and S. Strauss for advice and D. Wilson Sanchez for advice and sharing materials.
Competing interest
The authors declare none.
Data availability statement
Raw data, confocal stacks in tiff format as well as MorphoGraphX segmentation meshes are made available at: https://doi.org/10.17617/3.93
Author contributions
AH conceived and designed the study. LR-L performed experiments and analysed data. LR-L, MM, ZH and SM generated materials. AH and LR-L wrote the article.
Funding statement
This work was supported by the Deutsche Forschungsgemeinschaft FOR2581 on Plant Morphodynamics (A.H., grant number HA 6316/2-1). M.M. and S.M. were supported by the European Molecular Biology Organization Long Term Fellowships, S.M. by a National Science and Engineering Research Council of Canada Post-Doctoral Fellowship.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/qpb.2025.10015.






 Open access
Open access
Comments
In this study we take advantage of the high experimental tractability of the Arabidopsis relative, C. hirsuta, to study variation in a trait – petal number - that is invariant in Arabidopsis. As such, this work is timely and important as it increases our understanding of phenotypic diversity that is not accessible for study in Arabidopsis.
Here we use state-of-the-art live imaging protocols and quantitative image analysis (MorphoGraphX) to comparatively address the dynamics of petal development in space and time.
Our study addresses the relationship between genotype and developmental phenotypes and, as such, is relevant to geneticists, developmental biologists, evolutionary biologists, plant biologists, and researchers using quantitative approaches.
Many thanks for considering our work.