


1. Introduction
There are multiple ecological roles fulfilled by invertebrate organisms that make traces on bone substrates, as evidenced by the variety of forms this bioerosion can produce (Höpner & Bertling, Reference Höpner and Bertling2017). Organisms often bioerode in order to obtain nutrition. This nutrition may come from other soft-bodied organisms that inhabit the bone substrate, as with scavengers such as crabs that cause destruction of the bone while targeting worms, or microbial mats that cover the surface of the bone (Höpner & Bertling, Reference Höpner and Bertling2017). Organisms such as echinoids and gastropods, which produce surface traces rather than penetrative boreholes, often do so as a result of grazing on microbial mats as a source of nutrients, inadvertently marking the bone (Höpner & Bertling, Reference Höpner and Bertling2017). The bone itself can even be a source of nutrition, as with the bone-eating worm Osedax (Rouse et al. Reference Rouse, Goffredi and Vrijenhoek2004). Some bioeroding taxa also use the bones as a substrate or dwelling place, including boring clams and clionaid sponges (Höpner & Bertling, Reference Höpner and Bertling2017). By contrast, mechanical borers, such as most burrowing bivalves, are not nutrient miners and do not require a particular substrate, because they bore by rasping and not by dissolution (Tapanila et al. Reference Tapanila, Roberts, Bouare, Sissoko and O’Leary2004). Many of the clades of organisms that create surface etching or borings in bone also create similar traces in other hard substrates (Wisshak et al. Reference Wisshak, Knaust and Bertling2019). For example, sponges and clams have been known to bore into calcareous lithic substrates (Wisshak et al. Reference Wisshak, Knaust and Bertling2019), and echinoids graze on microbial mats and soft-bodied substrate colonizers, leaving traces on a variety of hard substrates today (Bromley, Reference Bromley1975). The most notable exception to this is Osedax, which is only found on vertebrate remains (Rouse et al. Reference Rouse, Goffredi and Vrijenhoek2004).
In the absence of preservation of soft tissues, or any surrounding sediment containing shell or exoskeletal remains, bioerosion on the surface of fossil bone can be used as a proxy for the presence of different invertebrate groups. Some taxa produce bioerosion with a highly distinctive morphology that does not vary much, whereas others produce a wider variety of morphologies when they etch or bore the bone (Pirrone et al. Reference Pirrone, Buatois and Bromley2014). Variation in the surface of the bone, such as degree of curvature, can also produce different traces, especially in the case of organisms such as echinoids and gastropods (Bromley, Reference Bromley1975; Pirrone et al. Reference Pirrone, Buatois and Bromley2014). Multiple factors must therefore be taken into account when identifying ichnotaxa and their invertebrate tracemakers.
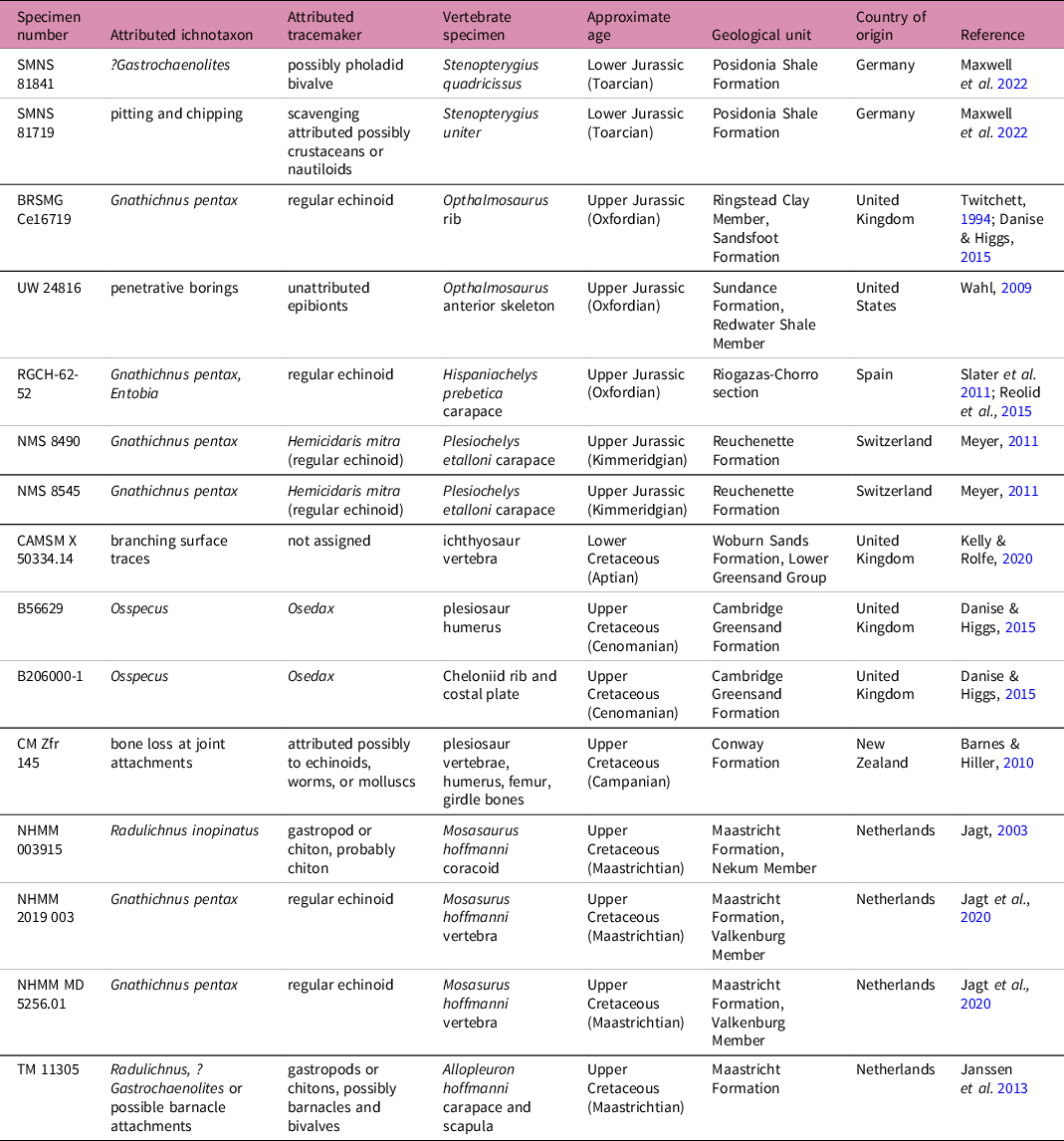
The taxa inferred to be present can potentially be used for partial reconstructions of community assemblages, inferences regarding localized environment (e.g. oxygen levels), identification of ecological roles (e.g. substrate colonizer, motile scavenger), and potentially broader ecological and climatic interpretations if trends emerge when diversity and prevalence of types of bioerosion are examined through deep time. Despite this potential of bioerosion as a rich source of taphonomic and ecological data, there is a dearth of literature describing traces made on bone substrates. For example, globally, there are only 13 previously published accounts, describing 15 instances of such bioerosion, on fossil marine reptiles (Table 1). This might indicate that such ecological interactions are seldom preserved. Here, coupled with a review of the literature, we survey large-bodied Jurassic and Cretaceous marine reptiles in four major UK museum collections. We describe and illustrate several different types of documented bioerosion, evaluate temporal patterns and demonstrate the potential utility of identifying bioeroding taxa in the fossil record for taphonomic and palaeoecological studies.
Instances of invertebrate bioerosion on Mesozoic marine reptile bone from the literature

2. Institutional abbreviations
BRSMG: Bristol Museum and Art Gallery, Bristol, United Kingdom; CAMSM: Cambridge Sedgwick Museum of Earth Sciences, Cambridge, United Kingdom; NHMUK: Natural History Museum, London, United Kingdom; OUMNH: Oxford University Museum of Natural History, Oxford, United Kingdom.
3. Bioerosion and Bioeroders
3.a. Invertebrate bioerosion in the fossil record of Mesozoic marine reptiles: a review
The literature on invertebrate bioerosion in Mesozoic marine reptile bone currently consists of 13 studies, describing 15 specimens, all from the Jurassic and Cretaceous, and predominantly from Europe (Table 1). In some cases, the identification of ichnotaxa and the associated tracemaker is clear and unequivocal, whereas in others, the identification is tentative. Many of these identifications have depended on studies of traces and tracemakers from other fossil bone types, or on analogous traces in present-day lithic substrates.
3.b. Osedax
First described from present-day whalefalls in 2004, the siboglinid annelid worm Osedax is found only in vertebrate remains in marine environments (Rouse et al. Reference Rouse, Goffredi and Vrijenhoek2004). These worms are characterized by the lack of a digestive tract and the adaptation of heterotrophic symbiosis with bacteria to gain nutrition from digesting lipids or collagen within vertebrate material (Rouse et al. Reference Rouse, Goffredi and Vrijenhoek2004; Rouse & Goffredi, Reference Rouse and Goffredi2023). Osedax boreholes are classified under the ichnogenus Osspecus (Higgs et al. Reference Higgs, Little, Glover, Dahlgren, Smith and Dominici2012). Osedax borings are generally characterized by only one opening leading to a single internal chamber, with interspecific variation in the morphology of the chamber (Higgs et al. Reference Higgs, Glover, Dahlgren, Smith, Fujiwara, Pradillon, Johnson, Vrijenhoek and Little2014). The chambers of the borings have branching structures and are approximately 3 mm–1 cm in diameter, with the entrance boreholes usually only ∼1 mm in diameter (Higgs et al. Reference Higgs, Glover, Dahlgren, Smith, Fujiwara, Pradillon, Johnson, Vrijenhoek and Little2014). This trace has been described from a plesiosaur humerus and a sea turtle rib and costal plate from the Cenomanian (early Late Cretaceous) of the UK and represents the only current evidence for the presence of Osedax in the Mesozoic (Danise & Higgs, Reference Danise and Higgs2015). Detailed comparison with CT-scans of Osspecus in present-day whalebone (Higgs et al. Reference Higgs, Glover, Dahlgren and Little2011; Higgs et al. Reference Higgs, Glover, Dahlgren, Smith, Fujiwara, Pradillon, Johnson, Vrijenhoek and Little2014: Figure 1) showed that the support for this Mesozoic identification of Osedax is robust (Danise & Higgs, Reference Danise and Higgs2015: Figure 2). As such, it seems that Mesozoic marine reptiles may have played a similar role to cetaceans for certain specialist taxa (Hogler, Reference Hogler1994). It is not clear how much further back into the Mesozoic Osedax traces might be found, or the extent of their prevalence throughout this time period.
3.c. Clionaid sponges
Clionaid sponge borings can be mistaken for those of Osedax because both create a series of round holes on the bone surface (Higgs et al. Reference Higgs, Little, Glover, Dahlgren, Smith and Dominici2012). The internal morphology of borings can help to distinguish between the two, with sponge borings interconnected by networks of channels beneath the cortical layer, unlike the single-entrance chambers of Osedax (Palmer & Plewes, Reference Palmer and Plewes1993; Higgs et al. Reference Higgs, Little, Glover, Dahlgren, Smith and Dominici2012; Higgs et al. Reference Higgs, Glover, Dahlgren, Smith, Fujiwara, Pradillon, Johnson, Vrijenhoek and Little2014). The round holes formed by sponges are often found in oyster shells (Breton et al. Reference Breton, Wisshak, Néraudeau and Morel2017), but they bore less commonly into bone substrates (Cione et al. Reference Cione, Acosta Hospitaleche, Pérez, Laza and César2010; Higgs et al. Reference Higgs, Little, Glover, Dahlgren, Smith and Dominici2012).
The ichnogenus Entobia is produced in the fossil record and today by clionaid sponges (Palmer & Plewes, Reference Palmer and Plewes1993; Bromley, Reference Bromley2004; Wilson, Reference Wilson and Miller2007; Breton et al. Reference Breton, Wisshak, Néraudeau and Morel2017). Entobia has been described in one fossil sea turtle from the Oxfordian (Late Jurassic) of Spain, based on the presence of its distinctive round boreholes (Slater et al. Reference Slater, Reolid, Schouten and Benton2011: pl. 1f). However, this specimen was not CT-scanned and the identification of the tracemaker was based entirely on surface holes; given the potential confusion with Osedax traces based on external morphology, an Entobia identification remains uncertain.
3.d. Echinoids
Echinoid grazing traces are known from oyster shell substrates (Palmer & Plewes, Reference Palmer and Plewes1993; Breton et al. Reference Breton, Wisshak, Néraudeau and Morel2017) and present-day aquaria and carbonate substrates (Bromley, Reference Bromley1975). These traces are classified under the ichnogenus Gnathichnus, which has a distinctive pentaradial symmetry due to the five mouthparts of echinoids (Bromley, Reference Bromley1975: Figure 1, pls 85, 86). Traces often overlap one another and cover an area of bone surface that has been grazed; clearer individual marks that do not overlap have a stellate geometry that shifts based on the curvature of the bone surface (Bromley, Reference Bromley1975; Wilson, Reference Wilson and Miller2007).
The traces caused by echinoids grazing on algal and bacterial mats are the most commonly described ichnotaxon from fossil bone, perhaps because of their distinctive morphology (Bromley, Reference Bromley2004; Wilson, Reference Wilson and Miller2007). Gnathichnus pentax has been described on a plesiosaur rib from the Oxfordian of the United Kingdom (Twitchett, Reference Twitchett1994; Danise & Higgs, Reference Danise and Higgs2015: Figure 1), in sea turtle bone from the Oxfordian (Late Jurassic) of Spain concurrently with Entobia (Slater et al. Reference Slater, Reolid, Schouten and Benton2011: pl. 1), in two sea turtle carapace pieces from the Kimmeridgian (Late Jurassic) of Switzerland (Meyer, Reference Meyer2011: Figure 5, 7) and in two mosasaur vertebrae from the Maastrichtian (latest Cretaceous) of the Netherlands (Jagt et al., Reference Jagt, Deckers, De Leebeeck, Donovan and Nieuwenhuis2020: Figure 4).
3.e. Gastropods and chitons
Gastropods and chitons grazing on microbial mats also produce surface traces, classified as Radulichnus, an ichnogenus that consists of linear traces formed by the abrasive radulae of these molluscs, producing large, curvate branching patterns on the surface of the substrate (Wisshak et al. Reference Wisshak, Knaust and Bertling2019). This type of surface bioerosion is sometimes distinctive (Jagt, Reference Jagt2003; Bromley, Reference Bromley2004). Radulichnus has been described as a sea turtle and mosasaur from the Maastrichtian of the Netherlands (Jagt, Reference Jagt2003: pls 1, 2; Janssen et al. Reference Janssen, Van Baal and Schulp2013: Figure 8). The sea turtle specimen also preserves other forms of pitting and bioerosion that are not clearly identifiable but might represent barnacle attachment scars (Janssen et al. Reference Janssen, Van Baal and Schulp2013).
3.f. Bivalves
A common type of boring in carbonate substrates and bones is the round hole made by burrowing bivalves. Extant bivalves produce these borings as dwelling holes to hide from predators (Wisshak et al. Reference Wisshak, Knaust and Bertling2019). These traces, classified as Gastrochaenolites, are often distinctively large (sometimes >10 mm in diameter) and form round holes with a tapering club shape that grows larger towards the terminus as the bivalve grows (Bromley, Reference Bromley2004). The tracemakers, pholadid bivalves, are mechanical borers and have been known to leave Gastrochaenolites traces in phosphatic clasts as well as in carbonate substrates (Tapanila et al. Reference Tapanila, Roberts, Bouare, Sissoko and O’Leary2004: Figure 4, 5). Such bivalve borers, therefore, probably treated fossil bone as they would any other phosphatic clast.
Examples of bivalve bioerosion on fossil marine reptile remains have been tentatively identified in an ichthyosaur from the Toarcian (Early Jurassic) of Germany (Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022), and alongside Radulichnus in a sea turtle from the Maastrichtian of the Netherlands (Janssen et al. Reference Janssen, Van Baal and Schulp2013). However, neither of these examples is definitive, because the pits are shallow and therefore not distinctly clavate internally.
3.g. Bioerosion traces of uncertain identification
There are many forms of bioerosion that are not readily identifiable, either because ichnotaxa have not been described or the tracemaker has not been identified, or because preservation does not allow for identification. Extensive bone loss formed through round, interconnected holes in limb ends or other joint attachment surfaces is a taphonomic mode that might be the result of multiple, closely-packed Gastrochaenolites borings, perhaps in combination with scavengers such as crabs destroying the bone to feed on the soft-bodied micro- or macro-organisms therein (Barnes & Hiller, Reference Barnes and Hiller2010). Other forms of bone loss causing chipping and wear at the edges of broad, flat bones might be due to a combination of invertebrate scavengers (Maxwell et al. Reference Maxwell, Cooper, Mujal, Miedema, Serafini and Schweigert2022).
4. Materials and Methods
4.a. Data collection
Specimens of ichthyosaurs, plesiosaurs and crocodyliforms from the Jurassic and Cretaceous of the United Kingdom were examined in four collections during 2022–2023: the Bristol Museum and Art Gallery (BRSMG), the Cambridge University Sedgwick Museum of Earth Sciences (CAMSM), the Oxford University Museum of Natural History (OUMNH) and the Natural History Museum, London (NHMUK). In total, we assessed over 15,000 specimens, with >75% of these from Jurassic strata. The bioeroded fossil bones in this dataset are mainly sourced from (1) the Callovian–Oxfordian Oxford Clay Formation (Hudson, Reference Hudson1978; Hudson & Martill, Reference Hudson and Martill1991); (2) the Kimmeridgian Kimmeridge Clay Formation (Oschmann, Reference Oschmann1988); (3) the Aptian–Albian Lower Greensand Group, comprising reworked fossil material from Late Jurassic to Early Cretaceous deposits (Ruffell & Wach, Reference Ruffell and Wach1991; Ruffell & Batten, Reference Ruffell and Batten1994); and (4) the Cenomanian Upper Greensand Formation, usually referred to locally and in the literature as the Cambridge Greensand, comprising reworked fossil material from Albian deposits (Unwin, Reference Unwin2001; Hart & Fox, Reference Hart and Fox2020).
The types of bioerosion documented and photographed in this study are macro-scale traces visible to the naked eye and produced by invertebrate tracemakers on bone substrates. These traces are generally either surface traces or penetrative boreholes, and have varying degrees of depth and trace morphology, depending on the tracemaker (Pirrone et al. Reference Pirrone, Buatois and Bromley2014).
4.b. Taxonomic and ichnotaxanomic identifications
Traces were identified based on previous descriptions of fossil and extant bioerosion on a variety of substrates. Where examples bear a strong resemblance to previously published descriptions, or where the morphology of the supposed tracemaker relates directly to the morphology of the trace, the identifications are considered to be unequivocal. For example, the pentaradial mouth of an echinoid is a highly likely candidate for any star-shaped traces, even if they do not strongly resemble previously described instances of Gnathichnus. With regard to bivalve borings, if the tracemaker is present in the dwelling hole or the complete boring is club-shaped (suggesting the continual growth of a boring clam), the trace is considered to be unequivocal. Tentative assignments are made where the boring or surface trace is weathered or equivocal with respect to either the morphology of the tracemaker or previous descriptions. Taxa that create surface borings, which are superficially similar, such as Entobia and Osspecus, are only tentatively assigned because additional examination of the internal morphology, beyond surface description, is required for unequivocal assignment. If there is a higher degree of uncertainty and the trace cannot be connected to one tracemaking taxon, then the traces are not assigned to an ichnotaxon.
5. Results
5.a. General observations
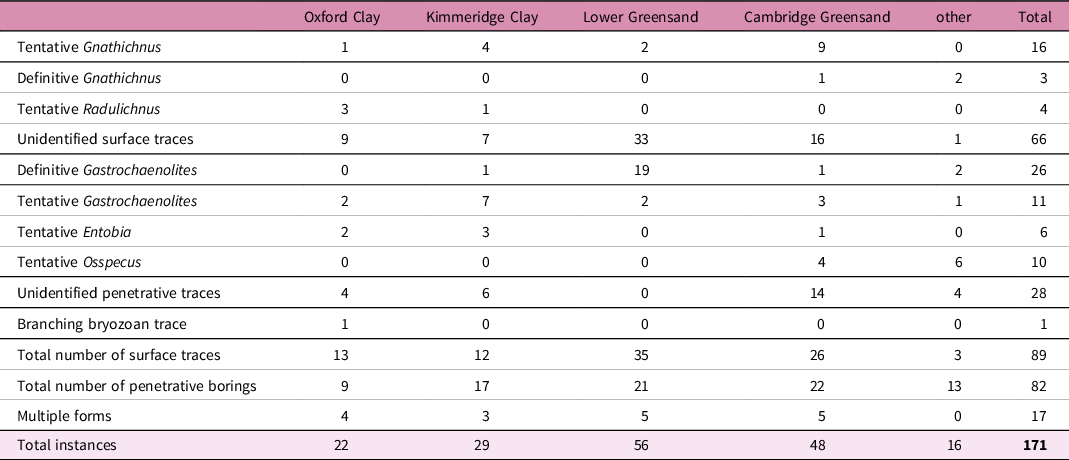
We identified 171 newly recognized instances of invertebrate bioerosion on 153 specimens of Jurassic and Cretaceous marine reptiles (online Supplementary Material at http://journals.cambridge.org/geo). This increases the known instances of invertebrate bioerosion on marine reptile bone by a factor of ten and implies that c. 1% of fossil bone that has been exposed on the seafloor prior to burial exhibits invertebrate traces. For the Jurassic stratigraphic units, 22 and 29 instances of bioerosion are sourced from the Oxford Clay and Kimmeridge Clay formations, respectively. In the Cretaceous, 56 and 48 instances are sourced from the Lower Greensand Group and Cambridge Greensand Formation, respectively. There are also 16 examples from two other Jurassic units and three other Cretaceous units, but because of the bias of collections and the distribution of fossil material in the United Kingdom, the data are largely from the four rock units mentioned above (Table 2).
Summary of the newly described instances of bioerosion

Nearly all of the specimens with bioerosion are from ichthyosaurs and plesiosaurs, with one specimen identified as a crocodyliform (BRSMG Cc17365), and one specimen representing either a crocodyliform or a plesiosaur (NHMUK PVR 629). It should also be noted that in one instance, a set of vertebrae belonging to both plesiosaurs and ichthyosaurs are catalogued under the accession number (CAMSM TN 1733), and it is not clear how many individual animals are present (online Supplementary Material at http://journals.cambridge.org/geo). Most of the marine reptile specimens housed in the UK collections, and therefore nearly all of those examined, are ichthyosaurs and plesiosaurs, so this taxonomic bias is unsurprising. The types of bioerosion described here occur indiscriminately on both plesiosaur and ichthyosaur bones (online Supplementary Material at http://journals.cambridge.org/geo). There are 107 instances of plesiosaur bone with bioerosion and 43 ichthyosaur bones with bioerosion, which broadly reflects the relative proportion of examined specimens belonging to the two groups (Table 2).
Bioerosion consists of examples of bones exhibiting identified and unidentified surface traces, bivalve borings and unidentified forms of boring and bone loss (see Table 2 for a summary). Taphonomic weathering processes and a lack of previous descriptions of traces can make the identification of bioerosion difficult at times, and only traces considered to be invertebrate bioerosion are included in the newly described instances presented in online Supplementary Material at http://journals.cambridge.org/geo. Thirty instances (∼18%) are definitively assigned to an ichnotaxon, and tentative identifications represent 47 instances (∼27% of the total data). A total of 94 instances of bioerosion (∼55%) are classified more generally into surface traces or borings that cannot readily be assigned to a specific ichnogenus. Sixteen (∼9%) of the specimens display multiple forms of bioerosion. The only ichnotaxa that are confidently identified are Gnathichnus and Gastrochaenolites, whereas Radulichnus, Entobia and Osspecus are only tentatively identified.
5.b. Surface traces
Surface traces make up ∼33% of the total dataset. Surface grazing traces have been identified with varying degrees of confidence. The only surface trace that is definitively identified here is Gnathichnus. There are 3 instances in which this ichnotaxon is confidently assigned, and 16 instances of tentative identification (Figure 1).
Examples of bioerosion are identified here as Gnathichnus. (a) and (b) Traces on a plesiosaur vertebra (CAMSM J.67977). (a) has been classified based on a resemblance to examples from the literature. See Bromley, Reference Bromley1975: Figure 1 and Jagt et al., Reference Jagt, Deckers, De Leebeeck, Donovan and Nieuwenhuis2020: Figure 4. (b) has been classified based on both the circled star-shaped trace and a resemblance to the bioerosion shown Bromley, Reference Bromley1975: pl. 85, Figure 2. (c) Star-shaped traces and scratch marks on an uncatalogued reptile element in the collections of CAMSM from the Lower Greensand. These traces have been classified based on the pentaradial shape of the circled individual traces and provide a good example of likely Gnathichnus, though the specimen is not included in the dataset, as the reptile taxon is undetermined. Scale bars are 10 mm.

There are four examples of surface traces tentatively identified as Radulichnus. These traces consist of small linear grooves but do not exemplify the broader meandering patterns in two parallel channels that are common in unequivocal cases of Radulichnus (Jagt, Reference Jagt2003: pl 1).
The vast majority of the documented surface traces (66 examples) are not assigned to any ichnotaxon because their morphology, though generally dendritic and suggestive of surface grazing by invertebrate organisms, is not distinct or is intermediate between previously described forms. There is a high degree of variability in curvature, depth of etching and extent of surface traces, which suggests a variety of taxa are responsible, but also complicates precise ichnotaxonomic identification.
Some surface traces resemble scratch marks and could have been made by a wide variety of scavengers. Others have a curvate branching morphology that matches that of certain calcifying substrate colonizers and might represent traces left behind when these organisms were weathered away (Figure 2). There is one example of a distinct trace left by a branching bryozoan adhering to the bone surface, which falls under the ichnogenus Finichnus (Taylor, Reference Taylor2013).
Examples of unidentified types of bioerosion. (a) A plesiosaur limb bone from the reworked Cenomanian Cambridge Greensand (NHMUK 35277), showing long, linear grazing traces. Given the scale of the traces, it is possible that these were created by the teeth of a vertebrate scavenger rather than an invertebrate tracemaker, but larger invertebrates such as crabs can make a variety of scraping traces. (b) A plesiosaur vertebra (NHMUK 46452) from the Aptian-Albian Lower Greensand, showing branching traces that match the morphology of the calcified branches adhered to the bone in (c), another plesiosaur vertebra (NHMUK PVR 2362) that is sourced from the same formation and collection. It is possible that calcifying organisms leave these traces. (d) A plesiosaur limb (CAMSM TN 1724) from the Cenomanian Cambridge Greensand showing deep branching surface traces. The roseate morphology of this trace is similar to others found in collections material, with some variability in depth and size, and bears a superficial resemblance to the sponge borings in belemnite guards described in Wisshak et al. Reference Wisshak, Titschack, Kahl and Girod2017: Figure 1 & 3. Scale bars are 10 mm.

5.c. Penetrative borings
Penetrative borings are described here in two forms, those that are more obviously bivalve borings (22% of the total dataset) and those that are unidentified forms of boring, pitting or bone loss (16% of the total dataset). In the data presented here, bivalve borings are usually from formations containing fossils that have been reworked. In total, 26 definitive, and 11 tentative, examples of Gastrochaenolites are described here. This is the only ichnotaxon that is morphologically distinct enough to have the number of definitive identifications exceed that of tentative identifications. These round borings range in size from ∼1 mm to >10 mm (Figure 3). At least two examples of these borings contain incontrovertible evidence for tracemaker identification because the bivalve that produced the hole remains in life position inside.
Examples of Gastrochaenolites. (a) A plesiosaur limb bone (CAMSM TN 1726) from the Cenomanian Cambridge Greensand with round borings. These boreholes bear a strong resemblance to bivalve borings in lithic substrates, such as (b) here. (b) an example of a lithic clast from the Lower Greensand (OUMNH K 37796), which has been bored by bivalves. Tapanila et al. Reference Tapanila, Roberts, Bouare, Sissoko and O’Leary2004: Figure 3 shows additional borings in lithics from the Eocene that strongly resemble (a) here. (c) A plesiosaur limb bone (CAMSM TN 3186) from the Lower Greensand. This bone bears a strong resemblance to the bivalve-bored Eocene fish bone in Tapanila et al. Reference Tapanila, Roberts, Bouare, Sissoko and O’Leary2004: Figure 3. A bivalve was found in life position in one of these borings. (d) A plesiosaur vertebra (OUMNH PAL-J.12321) from the Kimmeridgian. This boring is smaller in scale, showing that the morphology of bivalve traces remains the same regardless of the size or relative age of the bivalve creating the dwelling. In both (c) and (d), a bivalve was found in life position in one of the holes. Scale bars are 10 mm.

Boreholes and pitting, not obviously attributable to bivalve dwellings, are present in 28 instances (online Supplementary Material at http://journals.cambridge.org/geo). Many bones show varying degrees of pitting on the surfaces. It is possible that some of this pitting was caused by bivalves, but the morphology and size of the pits are highly variable, and it is not clear how many and what type of tracemakers are responsible.
The tracemakers of other penetrative borings, particularly small-scale ones (< ∼1 mm in diameter), are difficult to identify at the surface because of the lack of a visual line to the base of the boring and the fact that multiple taxa could potentially make similar small-scale borings. These borings cannot be identified to any meaningful taxonomic level without information on their internal morphology. Some degree of cortical layer erosion is also commonly present, perhaps due to networks of boreholes or collapsed chambers beneath the bone surface. Given the variable morphology and size range of these traces, several different taxonomic groups that have yet to be identified are probably responsible.
As with surface traces, some distinct but unidentified types of boring are recognised. For example, a deeply etched roseate pattern (Figure 1) might be a type of sponge boring, based on its resemblance to roseate sponge borings in belemnite guards (Wisshak et al. Reference Wisshak, Titschack, Kahl and Girod2017), but this has not been confirmed by a more detailed examination of the trace morphology.
Ten instances of tentative Osspecus have been recognised. It is not possible to confirm this identification unequivocally unless preservation is good and the specimens are CT-scanned to examine the internal morphology of the traces, but they bear a strong surficial resemblance to this ichnogenus.
5.d. Trends in bioerosion frequency through time
There are approximately twice the number of instances of bioerosion in the Cretaceous (115) specimens relative to those from the Jurassic (56) (see Table 2 for number of instances within each category of trace, formation and time bin). This ratio of instances of bioerosion holds true in the categories of both surface traces (28 in the Jurassic and 61 in the Cretaceous) and penetrative borings (28 in the Jurassic and 54 in the Cretaceous). The majority of the unidentified surface traces (33 instances) and nearly all of the definitive examples of bivalve borings (20 instances) are found in the Lower Greensand Group. Pitting and boring that are not identified are the most common (14 instances), and unidentified surface traces are still common (16 instances) in the Cambridge Greensand Formation. The only potential traces found solely in the Cretaceous are the tentatively recognized occurrences of Osspecus, which cannot be unequivocally identified without further investigation.
6. Discussion
6.a. Sampling
Although >75% of the examined specimens come from Jurassic deposits, ∼50% of the instances of bioerosion reported here are from Cretaceous specimens. This demonstrates that the relative prevalence of bioerosion in the Cretaceous is not an artefact of sample size. Thus, the disproportionate representation of bioerosion in the Cretaceous relative to the Jurassic in this dataset requires investigation and explanation.
6.b. Ecological factors
The trends in frequency and diversity of bioerosion in our dataset might have been influenced by the ‘Mesozoic radiation’ or ‘Mesozoic marine revolution’ (MMR) (Vermeij, Reference Vermeij1977). The Mesozoic is often cited as the period during which modern marine ecosystem structures emerged, and further examination of ecosystem structure throughout this time period may help to explain the breadth and timing of the ecological changes that occurred throughout the Jurassic and Cretaceous (Shaw et al. Reference Shaw, Coco, Wootton, Daems, Gillreath-Brown, Swain and Dunne2021; Buatois et al. Reference Buatois, Mángano, Desai, Carmona, Burns, Meek and Eglington2022). Shifts in marine ecological structure and food webs occurred throughout the Mesozoic, with an increase in productivity and resource availability to large apex predators in the Cretaceous relative to the Jurassic (Vermeij, Reference Vermeij1977; Hull, Reference Hull2017). The biotic turnovers and ecological restructuring continued through the Mesozoic, with a concentration of turnover in the mid-Cretaceous (Vermeij, Reference Vermeij1977; Rojas et al. Reference Rojas, Calatayud, Kowalewski, Neuman and Rosvall2021).
There are some previously observed trends in bioerosion assemblages that may be associated with the MMR, mainly in lithic substrates. These trends show a growing prevalence of infaunalization throughout the Mesozoic, with an increase in borings created as dwelling holes, potentially as a result of elevated levels of predation (Palmer & Plewes, Reference Palmer and Plewes1993; Taylor & Wilson, Reference Taylor and Wilson2003; Buatois et al. Reference Buatois, Mángano, Desai, Carmona, Burns, Meek and Eglington2022). Additionally, ichnotaxonomic diversity in lithic substrates increased throughout the Mesozoic (Palmer & Plewes, Reference Palmer and Plewes1993; Taylor, Reference Taylor2013). Grazing as a feeding mode also became more common later in the Mesozoic (Vermeij, Reference Vermeij1977). The presence or absence of similar trends in bioerosion on bone substrates has not previously been examined. The stratigraphic range of most of the forms of bioerosion identified here, with the exception of Radulichnus and Osspecus (which are unequivocally known only from the Cretaceous and later), covers the entire Mesozoic (Bromley, Reference Bromley2004), and all relevant bioeroding taxa apart from Osedax were present throughout the Mesozoic (the absence of descriptions of Radulichnus from the Jurassic does not imply that there was an absence of marine snails and chitons, given that both groups have a body fossil record from that period).
The potential influence of the MMR on the trends in our dataset is difficult to quantify because disentangling the influence of preservational bias in fossil vertebrates and invertebrates is a highly complicated issue. The exact timing of the events occurring throughout the MMR is also difficult to pin down (Buatois et al. Reference Buatois, Mángano, Desai, Carmona, Burns, Meek and Eglington2022). Trace fossils, being a subset of evidence for the overall presence of taxa, provide an additionally skewed representation of the abundance or diversity of the organisms responsible. Tracemakers cannot usually be identified to a fine taxonomic level, so it is not possible to tell the difference between a high-diversity and low-abundance assemblage and the inverse. Therefore, without additional data, it is not feasible to demonstrate that the observed correlation between the increase in abundance and diversity of taxa from the Jurassic through the Cretaceous associated with the MMR and the increase in frequency and prevalence of bioerosion in this particular dataset is causally linked.
6.c. The influence of taphonomy on trends in bioerosion
The Jurassic deposits (Oxford Clay and Kimmeridge Clay formations) are notably different in depositional environment to those of the Cretaceous (Lower Greensand Group and Cambridge Greensand Formation). The Oxford Clay and Kimmeridge Clay formations were deposited in shallow epicontinental seas with varying degrees of dysoxia in bottom waters (Hudson, Reference Hudson1978; Oschmann, Reference Oschmann1988; Hudson & Martill, Reference Hudson and Martill1991; Gallois, Reference Gallois2021). This could potentially have led to an underrepresentation of bioeroding invertebrate taxa. In the case of the Cretaceous deposits, which consist of sediment that has been eroded from the units below and then re-deposited at a later date (Unwin, Reference Unwin2001; Hart & Fox, Reference Hart and Fox2020; Kelly & Rolfe, Reference Kelly and Rolfe2020; Barrett, Reference Barrett2021), there were probably two distinct phases of bioerosion, one prior to initial burial of the bone material, and another after excavation and prior to re-burial.
The much higher prevalence of both surface traces and penetrative borings in the two targeted Cretaceous formations is probably the result of these burials and re-excavations. There are ten instances of multiple forms of bioerosion on bones from the Lower Greensand Group and Cambridge Greensand Formation, relative to the seven in the Oxford Clay and Kimmeridge Clay formations, reinforcing this point. When a bone is bioeroded prior to initial burial, and then has a second exposure on the sea floor at a later date, the second exposure is likely to produce additional traces, particularly from substrate colonizers, thereby potentially doubling the bioerosive traces on the surface of the bone. It stands to reason that much of the bioerosion in the Cretaceous stratigraphic units is therefore secondary. The high prevalence of Gastrochaenolites, which is generally described as a secondary trace in reworked deposits (Tapanila et al. Reference Tapanila, Roberts, Bouare, Sissoko and O’Leary2004), in the Lower Greensand Group supports this conclusion. The heavy degree of unidentified pitting in the Cambridge Greensand could reflect a similar phenomenon, representing either the weathered traces of boring clams or pitting caused by a weakening of the bone surface because of the secondary attachment of substrate colonizers such as oysters and sponges.
Osedax is the only bioeroding form that is not seen in the Jurassic rock units. Present-day examples of Osedax from shallow shelf environments analogous in depth to Jurassic epicontinental seas are from well-oxygenated waters (Glover et al. Reference Glover, Källström, Smith and Dahlgren2005), suggesting that lower oxygen levels might be a factor in limiting instances of these borings in the Jurassic units studied here. Alternatively, Osedax might be absent in the Jurassic because it had not yet originated or diversified. The latter interpretation is in keeping with molecular divergence time estimates and the earliest fossil evidence of the bone-eating worm that suggests it originated in the middle to Late Cretaceous (Vrijenhoek et al. Reference Vrijenhoek, Johnson and Rouse2009; Danise & Higgs, Reference Danise and Higgs2015; Taboada et al. Reference Taboada, Riesgo, Bas, Arnedo, Cristobo, Rouse and Avila2015). Determining whether Osedax was genuinely absent in the Jurassic, or if it has just not been found in our samples because we have only examined low-oxygen Jurassic environments to date, will require further fossil discoveries, a more extensive survey of museum collections, and/or visualization of the internal structures of boring traces from additional geographic localities and depositional environments that might have been made by Osedax or clionaid sponges.
In short, we suggest that the combined impacts of the low oxygen levels of the Kimmeridge Clay and the Oxford Clay formations potentially suppressing high levels of bioerosion, and the double exposure of the fossils from the Lower Greensand Group and the Cambridge Greensand Formation, are probably responsible for the differences in prevalence of bioerosion seen between units and time bins in the Jurassic and Cretaceous of the UK. Although large-scale and long-term macroevolutionary patterns associated with the Mesozoic Marine Revolution are also predicted to have had an impact on the prevalence of invertebrate bioerosion, a larger and more diverse dataset will be required in order to separate this factor from the more local and specialist factors relating to differences in environmental setting.
7. Conclusions
We present newly collected data on 153 bioeroded specimens of marine reptile bones, primarily from four Jurassic and Cretaceous UK stratigraphic units. This dataset significantly enlarges (approximately by an order of magnitude) the previous literature detailing invertebrate bioerosion on marine reptile bone, which comprises only 15 instances, and demonstrates that bioerosion is far more prevalent than previously realised. The bioerosion present in the two Cretaceous stratigraphic units is approximately double that present in the Jurassic formations, despite greater sampling from the latter period. This difference is probably due to two factors: lower oxygen levels in the Jurassic depositional environments suppressing higher levels of bioerosion, and the double exposure to bioeroding organisms in the Cretaceous units, where fossils are eroded out, reworked, and later re-deposited. Further work on specimens from a broader geographical range and additional ichnotaxonomic description will be required to quantify trends in the data more definitively and to parse the drivers behind these patterns.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0016756823000651
Acknowledgements
The authors would like to thank the following collections managers for access and assistance with loans: Matt Riley (CAMSM), Hilary Ketchum (OUMNH), Marc Jones (NHMUK), Susie Maidment (NHMUK) and Debbie Hutchinson (BRSMG). PDM’s contribution was supported by grants from The Royal Society (UF160216, URF\R\221010). We also thank two anonymous reviewers for their comments on the manuscript draft.
Author contributions
SJT conceived and designed the project, wrote the first draft of the manuscript, collected and analysed the data, prepared the figures and interpreted the results. PU and PDM contributed to writing the manuscript and helped with interpretation and conceptualization of the results.
Competing interests
The authors declare no conflicts of interest.






 Open access
Open access