


Management Implications
The habitat requirements and invasion dynamics of submerged aquatic vegetation (SAV) have important implications for future conditions along the Monongahela River in southwestern Pennsylvania. Hydrilla verticillata (hydrilla) tolerates low light, fluctuating flows, and a wide range of substrate types, allowing it to dominate both lentic and slower-moving lotic systems. In contrast, Myriophyllum spicatum (Eurasian watermilfoil) often expands in higher-light, moderately disturbed habitats and may proliferate where H. verticillata experiences seasonal dieback or incomplete canopy formation. Potamogeton crispus (curlyleaf pondweed), a cool-water species that grows through winter and often senesces by midsummer, can exploit early-season conditions before H. verticillata reaches peak biomass. Overlap among these ecological niches suggests the potential for shifts in species’ dominance through time, particularly under changing hydrologic conditions.
Collectively, these observations indicate that the upper Monongahela River now supports multiple invasive submerged aquatic plants, with H. verticillata currently the dominant invader. Continued monitoring will be necessary to track changes in species composition, evaluate potential competitive interactions among invasive macrophytes, and identify emerging management priorities for this reach of the river.
Introduction
Submerged aquatic vegetation (SAV) is an integral part of many aquatic systems. It provides cover for predatory fish (Korschgen and Green Reference Korschgen and Green1988; Rozas and Odum Reference Rozas and Odum1988), nursery waters for newly hatched fish and insect larvae (Miller et al. Reference Miller, Beckett and Blancher1987, Reference Miller, Kocovsky, Wiegmann and Miner2018), and food for macroinvertebrates (Valinoti et al Reference Valinoti, Ho and Armitage2011), and it slows erosion (Eulie et al. Reference Eulie, Walsh, Corbett and Mulligan2017). Numerous studies document both the positive and negative impacts of SAV on aquatic systems, noting that SAV type (species) is of great importance (Landry and Golden Reference Landry and Golden2018; Li et al. Reference Li, Weller, Gallegos, Jordan and Kim2007; Orth et al. Reference Orth, Dennison, Jonathan, Lefcheck, Gurbisz, Hannam, Keisman, Brooke, Moore, Murphy, Patrick, Testa, Weller and Wilcox2017, Reference Orth, Dennison, Wilcox, Batiuk, Landry, Gurbisz, Keisman, Hannam, Lefcheck, Murphy, Moore, Patrick, Testa, Weller, Merrittj and Hobaugh2022; Patrick et al. Reference Patrick, Weller, Li and Ryder2014, Reference Patrick, Weller, Orth, Wilcox and Hannam2018). In the Monongahela River system of southwestern Pennsylvania, conditions are suitable for SAV growth and it has become ubiquitous (Hofstra et al., Reference Hofstra, Clayton, Green and Auger1999; True-Meadows et al., Reference True-Meadows, Haug and Richardson2016; Boylen et al., Reference Boylen, Eichler and Madsen1999; Nichols and Shaw Reference Nichols and Shaw1986).
Hydrilla (Hydrilla verticillata (L. f.) Royle], a submerged, clonal aquatic plant, is widely recognized as one of the most aggressive invasive macrophytes in North America (Langeland Reference Langeland1996; USGS 2025). Its rapid stem elongation, prolific vegetative reproduction via stolons and turions, and capacity to form dense surface canopies allow it to alter habitat structure and competitively dominate freshwater plant communities (Harlan et al. Reference Harlan, Davis and Pesacreta1985). Recent taxonomic and genetic studies recognize three subspecies of H. verticillata in North America: ssp. verticillata, ssp. peregrina, and ssp. lithuanica (Foley et al. Reference Foley, Stebbins, Doherty, Tippery and Bugbee2024; Tippery Reference Tippery2023; Tippery et al. Reference Tippery, Bugbee and Weidman2020). Earlier literature commonly distinguished invasive populations based on reproductive form (monoecious vs. dioecious) (Steward Reference Steward1987); however, reproductive form does not necessarily correspond directly to subspecies. In the northeastern United States, H. verticillata populations have historically been attributed to the monoecious lineage, although genetic analyses indicate that multiple subspecies are present in the region.
First confirmed in 1989 in Connecticut from a pond at Mystic Seaport, H. verticillata (New London County) (Les et al. Reference Les, Mehroff, Cleland and Gabel1997) has since spread to inland lakes and the Connecticut River, reflecting a progressive expansion over nearly four decades. The taxon documented in the Connecticut River includes H. verticillata ssp. lithuanica based on genetic evidence (Tippery Reference Tippery2023). In the present study, specimens were identified to species using regional floristic keys (Crow and Hellquist Reference Crow and Hellquist2000).
While H. verticillata has been reported in portions of the upper Ohio River drainage (Department of Environmental Protection and the Pennsylvania Fish and Boat Commission, personal communication), peer-reviewed documentation of its presence in the Monongahela River in Pennsylvania is limited. The objective of this study was to document and verify the presence of H. verticillata within the mainstem of the Monongahela River in Pennsylvania and to provide baseline distributional information for this reach of the river. Here, we report the occurrence of Hydrilla verticillata ssp. peregrina (here called H. verticillata) and other aquatic invasive plants, consistent with broader patterns of northward expansion. Subspecies assignment to H. verticillata ssp. peregrina was based on morphological traits consistent with the taxonomic treatment of Tippery (Reference Tippery2023) and associated lineage descriptions. Genetic confirmation was not conducted.
Materials and Methods
The Monongahela River is best defined as a working river. It has been key in the economic development of the region and serves both commercial and recreational interests (Bell Reference Bell1921; Parker Reference Parker1999). The field survey reported in here was conducted from October to November 2025 among 14 sites (Figure 1). The survey was initiated in response to observations made during a related project in summer 2025 that documented the presence of H. verticillata in the Monongahela River from artificial reefs. Our fall survey was undertaken to independently verify its occurrence and to assess its distribution and relative extent in the lower Monongahela River, specifically within Allegheny, Washington, Greene, and Fayette counties, Pennsylvania (Figure 1).
Map depicting 14 sampling locations along the Monongahela River in southwestern Pennsylvania. Sampling locations are identified as: 1, Elizabeth boat ramp; 2, New Eagle boat ramp; 3, Monongahela Aquatorium; 4, Black Diamond boat ramp; 5, Monessen boat ramp; 6, Charleroi Community Park; 7, Speers boat ramp; 8, California boat ramp; 9, West Brownsville kayak access; 10, Engle’s Marina, Fredericktown, PA; 12, Rices Landing boat ramp; 13, Greensboro boat ramp; and 14, Point Marion boat ramp.

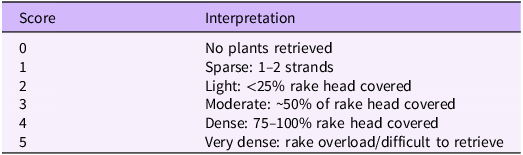
Submerged aquatic vegetation (SAV) was sampled using a double-sided metal rake mounted on a pole. Following the methods described by Madsen and Wersal (Reference Madsen and Wersal2017), the rake was lowered vertically to the substrate, rotated to collect vegetation, and lifted to the surface for identification. Abundance was scored using the scale described by Valley et al. (Reference Valley, Madsen and Welling2005; Table 1). Because this survey was intended as a rapid assessment, sampling intensity at each of the 14 sites was limited to approximately one to two rake drops.
Summary of rake fullness index used to assess density of submerged aquatic vegetation in the Monongahela River (Valley et al. Reference Valley, Madsen and Welling2005).

Boat ramps and landmarks (Figure 1) spanning an area from the towns of Point Marion, PA, to Elizabeth, PA (nearly river 106 km) were targeted during October and early November on the Monongahela River, including one major tributary, 10-mile Creek (Figure 1). In total, 14 collections were made. Records from all sources were summarized, and voucher specimens documenting species’ presence were prepared. Each species of SAV was identified, photographed, and catalogued into the herbarium collection at PennWest California.
To evaluate prior documentation of H. verticillata in this reach, we conducted database searches in fall 2025 using the Nonindigenous Aquatic Species (NAS) database maintained by the U.S. Geological Survey and searched herbarium specimen records available through the Mid-Atlantic Herbaria Consortium (Jacono et al. Reference Jacono, Richerson, Morgan, Pfingsten and Redinger2026; MAH 2025; USGS 2025). The Mid-Atlantic Herbaria Consortium verified voucher material housed at the Carnegie Museum of Natural History Herbarium (CM) (MAH 2025). These searches were intended to document any prior confirmed records of H. verticillata within the study reach before our 2025 survey.
Results and Discussion
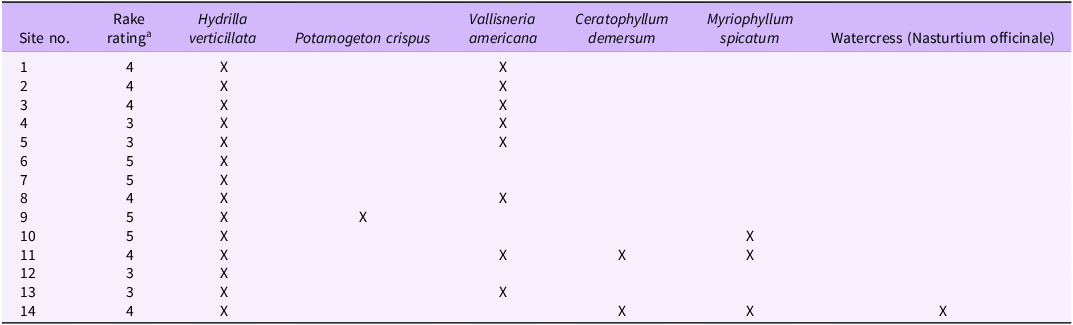
Our 2025 field surveys detected a consistent assemblage of submerged macrophytes consisting of H. verticillata, two additional non-native invasive species, and two native taxa (Table 2). Hydrilla verticillata was present at every site, often in high abundance, yielding an average rake score of 4 (Table 2), while Eurasian watermilfoil (Myriophyllum spicatum L.), curlyleaf pondweed (Potamogeton crispus L.), vallisneria (Vallisneria americana Michx.), and coontail (Ceratophyllum demersum L.) occurred less frequently and in lower abundances across sites.
Summary of recent collection made on the Monongahela River, PA, and the associated rake rating score of Hydrilla verticillata as determined by the methods describe by Madsen and Wersal (Reference Madsen and Wersal2017).

a Refer to Table 1 for community rake rating scoring criteria.
A 2016 voucher specimen from the Pennsylvania portion of the river provides the first curated physical confirmation of H. verticillata occurrence in this reach (Carnegie Museum Herbarium accession CM536399; Grund and Szoszorek Reference Grund and Szoszorek2016; Supplementary Table S1). Additional evidence of early presence in the upper drainage comes from a resident letter of September 4, 2017, to the Federal Energy Regulatory Commission (FERC) describing extensive H. verticillata mats in Cheat Lake, WV, that clogged boat propellers and coolant intakes, the text of which is publicly reproduced by the Upper Mon River Association (UMRA 2024). Cheat Lake lies within the upper Monongahela River basin and discharges downstream into the mainstem Monongahela River, representing a plausible upstream source population (Figure 1). Further indirect confirmation arises from Maryland Department of Natural Resources boat-inspection records at Deep Creek Lake, MD, where vessels last used on Cheat Lake, WV, carried attached H. verticillata fragments during late summer 2017 and 2018 (Bortz and Metheny Reference Bortz and Metheny2019). More recent documentation comes from a 2024 FERC Additional Information Request for the Lake Lynn Hydroelectric Project (Project No. 2459-279), which cites stakeholder reports of H. verticillata impacts and directs the licensee to characterize its distribution within Cheat Lake (FERC 2024).
Contemporary observations are also reflected in the U.S. Geological Survey (USGS) NAS database, which lists confirmed occurrences of H. verticillata at the Glassworks boat launch in Greene County, PA, cataloged as H. verticillata ssp. peregrina, as well as multiple provisional records from 2023 to 2024 in the Upper Monongahela River in Monongalia County, WV (USGS 2025). Although these records lack peer-reviewed population descriptions, together they demonstrate that H. verticillata has established throughout the Upper Monongahela River drainage. These historical and contemporary records suggest a long-standing but underdocumented presence of H. verticillata within the basin, with spread occurring along the navigational pools over the past decade or more.
During sampling from late October to early November, plants exhibited clear seasonal senescence, with declining aboveground biomass. Overwintering structures observed during this period consisted primarily of axillary turions; sediment tubers were not detected. These traits are consistent with monoecious H. verticillata populations documented from northern latitudes. Subspecies assignment to H. verticillata ssp. peregrina was based on morphological traits consistent with the taxonomic treatment of Tippery (Reference Tippery2023). Monoecious H. verticillata populations in northern North America have historically been associated with the Eurasian lineage referred to as Hydrilla verticillata ssp. peregrina (Madeira et al., Reference Madeira, Van, Steward and Schnell2000; Tippery et al. Reference Tippery, Bugbee and Weidman2020; Foley et al. Reference Foley, Stebbins, Doherty, Tippery and Bugbee2024). Although the observed traits are consistent with this lineage, morphological characteristics alone cannot reliably distinguish among Eurasian subspecies (e.g., H. verticillata ssp. peregrina vs. H. verticillata ssp. lithuanica).
We note that while Crow and Hellquist (Reference Crow and Hellquist2000) present an appropriate regional authority for species-level identification of H. verticillata, subspecies are not distinguished. In fact, there are no published identification keys that reliably differentiate H. verticillata subspecies based on vegetative morphology. Subspecies designations in Hydrilla are based on reproductive strategy, geographic provenance, and molecular lineage rather than on diagnostic morphological characters. Definitive subspecies assignment would therefore require molecular confirmation, which was beyond the scope of the present study.
Surveys conducted from the Pennsylvania–West Virginia border to Elizabeth, PA, documented H. verticillata at all 14 visited sites, indicating that the species is now widespread along this nearly 100-km section of the Pennsylvania portion of the Monongahela River. Hydrilla verticillata was consistently the dominant submerged macrophyte in terms of abundance and spatial extent (Table 2), a pattern consistent with its invasion dynamics elsewhere in North America (Langeland Reference Langeland1996; Rybicki and Landwehr Reference Rybicki and Landwehr2007). Its continuous distribution across multiple pool segments suggests a well-established, basin-level invasion rather than isolated colonization events.
At several sites, H. verticillata co-occurred with two invasive submerged macrophytes: M. spicatum and P. crispus. Although these species were less abundant than H. verticillata during our surveys, both are recognized aquatic invaders capable of expanding rapidly under suitable environmental conditions (Nichols and Shaw Reference Nichols and Shaw1986; Smith and Barko Reference Smith and Barko1990). Myriophyllum spicatum was present at multiple locations and should be considered a species of management concern moving forward. Potamogeton crispus occurred at fewer sites and at low abundance, a pattern typical of its cool-season phenology (Nichols and Shaw Reference Nichols and Shaw1986).
Myriophyllum spicatum was observed at multiple sites between the West Virginia–Pennsylvania border and the city of Monongahela, PA (Figure 1; Table 2; Supplementary Table S1). Although encountered at lower densities than H. verticillata, its repeated presence across pools indicates that it is established in the system. Herbarium vouchers collected over the past four decades document a persistent occurrence of M. spicatum in the Pennsylvania section of the Monongahela River (MAH 2025; Table 2), underscoring its long-established but historically underreported status. The USGS NAS database records corroborate this pattern, with confirmed entries in 2003 and 2016 (USGS 2025).
Potamogeton crispus was considerably more limited in distribution. It was detected only once during our recent survey as a small patch embedded within an H. verticillata bed. No voucher specimens of P. crispus were found in regional herbaria, and the species does not appear in USGS NAS records for the Monongahela River (USGS 2025). These observations indicate that P. crispus is either newly present, historically underreported, or present only at very low densities within this reach of the system. However, surveys were conducted on November 4, after the typical period of peak biomass for P. crispus, and seasonal senescence may have contributed to its limited detection.
Two native submerged aquatic plant species were documented within the Pennsylvania portion of the Monongahela River. Vallisneria americana was detected at several sites, typically occurring as scattered patches situated adjacent to the more expansive H. verticillata beds. Ceratophyllum demersum was considerably less common, occurring only at two sites as small, isolated patches. Neither native species formed large or continuous colonies, and both appeared restricted to localized areas embedded within H. verticillata–dominated nearshore habitats. At some sampling locations, floating mats of the filamentous algae Pithophora sp. (Roth) Wittrock were also observed interspersed with H. verticillata. Identification was based on microscopic examination of filament morphology and chloroplast characteristics.
This investigation was conducted as a rapid assessment to document the presence of H. verticillata and other submerged aquatic vegetation within the lower Monongahela River following initial observations in summer 2025. Sampling focused on publicly accessible boat ramps to enable efficient coverage of a broad river reach rather than to quantify population density, biomass, or fine-scale spatial distribution. Accordingly, these results should be interpreted as documentation of occurrence within the lower Monongahela River rather than a comprehensive distributional survey. Given evidence of earlier establishment in the upper Monongahela drainage (e.g., Cheat Lake and associated upstream records), future efforts should prioritize systematic, spatially explicit surveys of the mainstem Monongahela River to evaluate downstream expansion and population extent, as well as molecular analyses to confirm subspecies identity.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2026.10044
Funding statement
This research received no specific grant from any funding agency or the commercial or not-for-profit sectors.
Competing interests
The authors declare no conflicts of interest.




 Open access
Open access