


Introduction
Animal welfare can be defined as the physical and mental state of well-being of an animal and constitutes a balance between their positive and negative experiences (Spruijt et al. Reference Spruijt, Van den Bos and Pijlman2001; Ohl & van der Staay Reference Ohl and van der Staay2012; Rault et al. Reference Rault, Bateson, Boissy, Forkman, Grinde, Gygax, Harfeld, Hintze, Keeling, Kostal, Lawrence, Mendl, Miele, Newberry, Sandøe, Špinka, Taylor, Webb, Whalin and Jensen2025). One of the most commonly used frameworks to assess welfare is the Five Domains model (Mellor & Reid Reference Mellor and Reid1994; Mellor & Beausoleil Reference Mellor and Beausoleil2015; Mellor et al. Reference Mellor, Beausoleil, Littlewood, McLean, McGreevy, Jones and Wilkins2020). This framework posits that an animal’s mental experiences are a determining factor for its welfare state. These mental experiences are shaped by four physical domains: nutrition; environment; health; and behavioural interactions. For example, certain environments allow animals to engage in rewarding species-specific behaviours. In this specific case, the physical domains ‘environment’ and ‘behaviour’ would positively influence the mental state of the animal, thereby promoting good welfare.
Over the years, zoological institutions have increasingly prioritised animal welfare, gradually working towards the best welfare outcomes they can provide within their resource constraints. Ideally, the highest possible standard for animal welfare should be ensured, as it is not only ethically imperative, but also crucial for achieving the primary objectives of a modern zoo for conservation, education, research, as well as recreation (Fa et al. Reference Fa, Funk and O’Connell2011; Roe et al. Reference Roe, Mcconney and Mansfield2014; Beer et al. Reference Beer, Shrader, Schmidt and Yates2023; Greenwell et al. Reference Greenwell, Riley, Lemos de Figueiredo, Brereton, Mooney and Rose2023). Achieving these objectives depends on animals having positive welfare states. First, positive animal welfare is associated with greater reproductive success and longevity, which benefits conservation breeding programmes (Carlstead & Shepherdson Reference Carlstead and Shepherdson1994; Tidière et al. Reference Tidière, Gaillard, Berger, Müller, Bingaman Lackey, Gimenez, Clauss and Lemaître2016; Greggor et al. Reference Greggor, Vicino, Swaisgood, Fidgett, Brenner, Kinney, Farabaugh, Masuda and Lamberski2018; Edwards et al. Reference Edwards, Edes, Brown, Comizzoli, Brown and Holt2019). Second, animals displaying more species-typical behaviours aids the education goals of zoos and improves public sentiment towards zoos (Davey Reference Davey2006; McPhee & Carlstead Reference McPhee, Carlstead, Kleiman, Thompson and Baer2010; Ross et al. Reference Ross, Melber, Gillespie and Lukas2012; Hosey et al. Reference Hosey, Melfi and Pankhurst2013). Third, promoting species-typical behaviour improves future reintroduction success (Chappell & Thorpe Reference Chappell and Thorpe2022; Crates et al. Reference Crates, Stojanovic and Heinsohn2023). Lastly, positive welfare and display of species-typical behaviour are essential for producing reliable scientific insights, as they improve generalisability of results and minimise confounding variables (Poole Reference Poole1997; Prescott & Lidster Reference Prescott and Lidster2017). As such, stimulating species-typical behaviour to improve animal welfare is a critical step towards fulfilling the core missions of modern zoos.
Achieving optimal welfare in zoo settings is not without challenges. One major challenge is that animals are often housed in confined enclosures that do not fully resemble their natural environment and only have limited space and structural complexity. Enclosures that promote more natural behavioural patterns allow animals to make choices about how and where to move, rest, or explore, giving them a sense of control over their environment. For example, climbing structures, platforms, and varied substrates provide animals with multiple options for locomotion, which can promote expression of natural behaviours and reduce boredom because the environment is less predictable (Buchanan-Smith Reference Buchanan-Smith2011). However, enclosures that do not resemble a species’ natural environment can pose challenges to animal welfare because they over- or under-stimulate individuals, potentially leading to boredom, stress or frustration (Morgan & Tromborg Reference Morgan and Tromborg2007; Hosey et al. Reference Hosey, Melfi and Pankhurst2013; de Azevedo et al. Reference de Azevedo, Cipreste, Pizzutto and Young2023). This can impact the activity cycle of the animals living in such enclosures, leading them to perform different and less varied behavioural patterns than their wild counterparts (Kelly et al. Reference Kelly, Freeman, Rose, Baragli, Andrea and Henaut2025) or conspecifics in more appropriate enclosures (Neal Webb et al. Reference Neal Webb, Hau and Schapiro2018). In general, a diverse behavioural repertoire can be considered an indicator of positive welfare (Miller et al. Reference Miller, Pisacane and Vicino2016, Reference Miller, Vicino, Sheftel and Lauderdale2020; Hall et al. Reference Hall, Bryant, Staley, Whitham and Miller2021; but see Cronin & Ross Reference Cronin and Ross2019). In extreme cases, animals might develop abnormal repetitive behaviours when they are unable to cope with their environment, which may indicate impaired welfare (Mason Reference Mason, Mason and Rushen2006; Anthony et al. Reference Anthony, Dudzinski, Hill, Stafford, Tallo-Parra, Salas and Manteca2023). Looking beyond behavioural measures, poorly designed enclosures can also negatively affect the development of muscles and bones (Reilly & Franklin Reference Reilly and Franklin2016), as animals might experience less mechanical loading and less varied locomotion (Law & Kitchener Reference Law and Kitchener2020). Altogether, these factors highlight the need for careful enclosure design.
To avoid behavioural problems and welfare compromises, zoos should prioritise designing enclosures that cater to the behavioural needs of the animals. Such enclosures provide animals with sufficient challenge and do not restrict the natural behavioural repertoire of an individual (Morgan & Tromborg Reference Morgan and Tromborg2007; Hosey et al. Reference Hosey, Melfi and Pankhurst2013; Hall et al. Reference Hall, Bryant, Staley, Whitham and Miller2021). One way to accomplish this is to design enclosures that more functionally resemble the natural habitat of the species. Such enclosures are sometimes referred to as Functionally Appropriate Captive Environments (FACEs) (Reamer et al. Reference Reamer, Haller, Lambeth and Schapiro2017; Neal Webb et al. Reference Neal Webb, Hau and Schapiro2018). FACEs aim to mimic the natural environment for the species living in it, not necessarily for the human eye, by targeting biologically relevant elements that the animals interact with and allow them to perform their full behavioural repertoire. As such, they promote the occurrence of species-typical behaviours and simulate natural interactions with the environment (Fàbregas et al. Reference Fàbregas, Guillén-Salazar and Garcés-Narro2012; Bryan et al. Reference Bryan, Bremner-Harrison, Price and Wormell2017; Fieschi-Méric et al. Reference Fieschi-Méric, Ellis, Servini, Tapley and Michaels2022). One specific example comes from arboreal species, for whom climbing structures can help mimic elements of their natural environment. These structures provide physical stimulation by allowing animals to move more naturally in a three-dimensional space (Jensvold et al. Reference Jensvold, Sanz, Fouts and Fouts2001; Schmidt Reference Schmidt2011). For arboreal species, the design of enclosures and climbing structures therefore contributes to their overall welfare under human care.
One arboreal taxon that benefits from well-designed climbing structures is the orangutan (Tripp Reference Tripp1985; Perkins Reference Perkins1992; Roth et al. Reference Roth, Bionda and Sterck2017). The three species of orangutans (Pongo spp.) are the largest of all arboreal mammals (Delgado Jr & Van Schaik Reference Delgado and Van Schaik2000; Nater et al. Reference Nater, Mattle-Greminger, Nurcahyo, Nowak, de Manuel, Desai, Groves, Pybus, Sonay, Roos, Lameira, Wich, Askew, Davila-Ross, Fredriksson, de Valles, Casals, Prado-Martinez, Goossens, Verschoor, Warren, Singleton, Marques, Pamungkas, Perwitasari-Farajallah, Rianti, Tuuga, Gut, Gut, Orozco-terWengel, van Schaik, Bertranpetit, Anisimova, Scally, Marques-Bonet, Meijaard and Krützen2017). Their morphological adaptations allow them to navigate the forest canopy with efficiency: long forelimbs and short hindlimbs enable them to brachiate and sway between branches (Delgado Jr & Van Schaik Reference Delgado and Van Schaik2000). Furthermore, orangutans possess four very long digits and short opposable thumbs on both hands and feet, allowing for a sturdy grip while suspended (Delgado Jr & Van Schaik Reference Delgado and Van Schaik2000; Patel et al. Reference Patel, Orr and Jashashvili2020). These adaptations support a largely arboreal lifestyle. Wild orangutans spend approximately 15% of their time travelling, and this movement occurs almost entirely above ground (Morrogh-Bernard et al. Reference Morrogh-Bernard, Husson, Knott, Wich, Van Schaik, Van Noordwijk, Lackman-Ancrenaz, Marshall, Kanamori, Kuze, Bin Sakong, Mitra Setia and van Schaik2009). Key activities, such as reproduction, foraging and evading predators all depend on effective above-ground locomotion (Thorpe & Crompton Reference Thorpe, Crompton, Wich, Atmoko, Setia and van Schaik2008; Chappell & Thorpe Reference Chappell and Thorpe2022). In addition, orangutans are extremely energy-efficient (Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010), potentially due to their challenging environment, where they are often confronted with long periods of fruit scarcity (Vogel et al. Reference Vogel, Alavi, Watford, Brittain, Crowley, Naumenko, Aguado, Bransford, Zulfa, Moldawer, Farida, van Noordwijk, Setia, Utami Atmoko, Rothman and Raubenheimer2025). This also relates to their locomotion behaviour, which is characterised by energy-efficient strategies to move through the canopy (Thorpe et al. Reference Thorpe, Crompton and Alexander2007, Reference Thorpe, Holder and Crompton2009).
Wild orangutans employ a rich repertoire of locomotion types to move efficiently through complex canopy environments. During travel, they heavily rely upon suspensory forms of locomotion, such as torso-orthograde suspensory locomotion, where they move by hanging and using their forelimbs (Thorpe & Crompton Reference Thorpe and Crompton2005; Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). These forms of locomotion allow for efficient gap crossing, while providing sufficient stability by having contact with multiple supports (Thorpe & Crompton Reference Thorpe and Crompton2005; Thorpe et al. Reference Thorpe, Holder and Crompton2009). In addition, they use swaying to cross gaps between trees while travelling (Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). While feeding, they still heavily rely on suspensory locomotion, but they also use a rich repertoire of other locomotive behaviours that are relatively rare during travel, such as vertical climbing and descending to reach different parts of the tree or bipedal walking to reach for food at the ends of branches (Cant Reference Cant1987; Thorpe & Crompton Reference Thorpe and Crompton2006; Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). Quadrupedal walking is uncommon, accounting for only 8–18% of the time spent on movement and is mostly used in feeding contexts (Thorpe & Crompton Reference Thorpe and Crompton2006; Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). Importantly, the characteristics of climbing substrates, particularly their stiffness, strongly influence locomotion behaviours (Thorpe et al. Reference Thorpe, Holder and Crompton2009; Myatt & Thorpe Reference Myatt and Thorpe2011; van Casteren et al. Reference van Casteren, Sellers, Thorpe, Coward, Crompton and Ennos2013). For example, stiff structures facilitate quadrupedal walking and vertical climbing, while flexible structures encourage various forms of suspensory locomotion (Thorpe et al. Reference Thorpe, Holder and Crompton2009). Thus, depending on the structure of their environment, orangutans flexibly employ a wide range of locomotive behaviours.
Zoo-housed orangutans, however, show remarkable differences in locomotive behaviour compared to their wild counterparts. Unlike wild orangutans, they spend less time on movement and primarily move on the ground (Isler & Thorpe Reference Isler and Thorpe2003; Pearson et al. Reference Pearson, Davis and Litchfield2010). Furthermore, wild and zoo-housed orangutans differ in the frequencies with which they perform specific locomotive behaviours. For instance, zoo-housed orangutans often perform quadrupedal walking, which is rare in wild orangutans (Pearson et al. Reference Pearson, Davis and Litchfield2010; Roth et al. Reference Roth, Bionda and Sterck2017). In turn, suspensory locomotion — common in the wild — is relatively rare in zoos (Roth et al. Reference Roth, Bionda and Sterck2017). This can partly be attributed to the fact that zoo-housed orangutans are more terrestrial than their wild counterparts: while zoo-housed orangutans spend 30–50% of their time on the ground (Roth et al. Reference Roth, Bionda and Sterck2017), this percentage is lower than 5% for wild orangutans (Ashbury et al. Reference Ashbury, Posa, Dunkel, Spillmann, Atmoko, van Schaik and van Noordwijk2015). These differences in locomotion behaviour may even have important health consequences, such as obesity, for zoo-housed orangutans because of their slow metabolism (Gippoliti Reference Gippoliti2000; Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010). This risk increases when individuals reduce movement and rely more on energetically inexpensive forms of locomotion, such as quadrupedal walking (Hanna & Schmitt Reference Hanna and Schmitt2011; Pontzer Reference Pontzer2016). Also, ground-based movement places different pressures on their muscles and bones than above-ground locomotion (Isler & Thorpe Reference Isler and Thorpe2003). Over time, this may negatively affect the development of their musculoskeletal system and impair their climbing abilities (Sarmiento Reference Sarmiento1985; Isler & Thorpe Reference Isler and Thorpe2003; Chappell & Thorpe Reference Chappell and Thorpe2022). Indeed, a large panel of experts recently concluded that the differences in arboreal locomotion between wild and zoo-housed orangutans was one of the largest welfare risks for zoo-housed orangutans (Veasey et al. Reference Veasey, Armstrong, Butcher, Embury, Grossfeldt, Robbins and Thompson2025). Encouraging a diverse range of locomotion behaviours may thus improve their health and well-being.
Previous research suggests that enclosure design can affect locomotion behaviour of zoo-housed orangutans. For instance, enclosures with many horizontal trunks were found to increase the total amount of locomotion in zoo-housed orangutans compared to enclosures with smaller, flexible structures (Roth et al. Reference Roth, Bionda and Sterck2017). However, the enclosures with small, flexible structures did promote more diverse locomotion behaviour (Roth et al. Reference Roth, Bionda and Sterck2017). While these results suggest a trade-off between quantity (total amount) and quality (diversity) of locomotion, the previous study comprised a rather short observation period, immediately after one enclosure type was remodelled. As a result, the study may have missed behavioural patterns that emerge after long-term adaptation to the remodelled enclosures. The current study relies on focal observations to investigate the long-term effects of enclosure structure on the locomotion of zoo-housed Bornean orangutans (Pongo pygmaeus) at Apenheul Primate Park (Apeldoorn, The Netherlands). In this study, we focused on movement quantity and quality in two different enclosure types. One enclosure type contained a large number of stiff structures, such as horizontal and angled trunks. The other consisted mostly of thin, flexible structures, such as ropes.
First, we examined whether the amount of time spent on locomotion and other activities differed between the two enclosure types. Because orangutans are adapted to long periods of fruit scarcity and thus are characterised by extremely efficient locomotion behaviour (Thorpe et al. Reference Thorpe, Crompton and Alexander2007, Reference Thorpe, Holder and Crompton2009), we expected them to move more in enclosures with more trunks, as these structures allow for energy-efficient travel. Second, we investigated whether orangutans used different types of locomotion in the two enclosure types. Here, we expected more quadrupedal walking in enclosures with many trunks and more suspensory locomotion in those with many ropes. These forms of movement are typically associated with similar structures in the wild (van Casteren et al. Reference van Casteren, Sellers, Thorpe, Coward, Crompton and Ennos2013). Third, we explored the link between climbing structures and particular locomotive behaviours. Identifying such links could help guide the design of enclosures that promote more natural movement.
Materials and methods
Study site and subjects
In this study, seven zoo-housed Bornean orangutans were observed at Apenheul Primate Park in Apeldoorn, The Netherlands. Of these seven individuals, two were flanged males, three were adult females and two were subadults (Table 1). The study was conducted between February and September of 2024. All individuals were observed for 6 to 11 h in the inside enclosures, excluding time spent out of sight (Table 1).
All Bornean orangutans (Pongo pygmaeus) observed in this study (n = 7), including their demographic information and number of observation days and hours. Note that the observation times already exclude time out-of-sight

The animals were housed in four separate enclosures, each consisting of an inside enclosure connected to two outside islands (Figure 1). The inside enclosures each had a volume of 375 m3. In between these four inside enclosures there were eight separation enclosures, which could be closed off from the inside enclosures by automated doors. This created eight potential separation enclosures and allows for the animals to be moved around the enclosures. These separation enclosures were mostly inaccessible for the animals during the daytime, unless a keeper decided to open one of the 24 slide-doors connecting the enclosures. When temperatures allowed it (> 10°C when rainy and cloudy, > 5–6°C when sunny and without wind), the animals had access to both inside and outside enclosures during the day. Enrichment items, climbing structures and drinking water were present in both inside and outside enclosures, and feedings would also take place both inside and outside.
Satellite image showing the four different Bornean orangutan (Pongo pygmaeus) enclosures at Apenheul Primate Park (Google n.d.).

The building was originally designed to accompany a form of a fission-fusion holding system. However, in practice, the animals would be in the same composition most of the time due to the presence of two flanged males and some incompatible females. This resulted in all animals being kept in duos, except flanged male Amos, who was kept solitarily. The standard enclosure partners mostly consisted of mothers and their offspring but also included a pairing of an older flanged male and an older female. During 27 observations the composition deviated from the standard enclosure partners, most frequently combining Indah and Samboja with Sandy and Kevin. Individuals were rotated daily between the enclosures, together with their enclosure partner.
The animals were fed approximately three times within each daily observation time window (1000–1600h), but not according to a strict schedule. Additionally, they regularly received food enrichment (e.g. food puzzles, suspended food baskets or wooden blocks with holes for sticky edibles). During the observation window, food was presented twice in elevated food baskets and wooden blocks at different heights, and once just above ground level at the mesh in the inside enclosure. Drinking water was available at all times at ground level and at an elevated ridge in the inside enclosure, from taps attached to the wall. From the 6th of February to the 20th of March, the park was closed for visitors. The visitor season started on the 21st of March. Additionally, during the visitor season a fixed feeding presentation by caretakers was added to the feeding routine at 1300h. Nesting material was always present in the enclosures in the form of wood fibre, dispersed throughout the enclosure.
Detailed description of the inside enclosures
All inside enclosures were similar in size and basic design, with a height of 6.5 m and each with a floor surface area of approximately 58 m2 (Figure 2). The enclosures were designed to accommodate the orangutans’ need for vertical locomotion and were therefore layered at different height levels. A characterising feature of these inside enclosures is that the visitor viewing area was elevated. At the ground level was the zookeeper service area, on top of which the visitor area was located. Consequently, the visitor viewing windows only start approximately halfway of total height and extend to the ceiling. The bottom half of the inside enclosure was less visible for the visitors, providing space for the animals to avoid visitor interactions. The walls at the bottom half of the enclosures were covered with wood to facilitate climbing by the orangutans. At the bottom level of the enclosures a large mesh fence was present through which zookeepers could feed the animals and perform other standard procedures. At the visitors’ eye level, a broad ridge surrounded the edges of the enclosures, acting as an elevated piece of ground. This ridge, as well as the ground floor in the bottom half of the enclosures, was covered with biological substrate (biofloor consisting of biodegradable material such as wood-chips). Several metal rings were attached to the ceiling of the enclosures to enable mounting of different hanging structures.
Photographs depicting the two types of inside enclosures for Bornean orangutans (Pongo pygmaeus) at Apenheul Primate Park, showing (a) the large trunk-type, which is characterised by many large trunks and few ropes, and (b) multiple ropes-type, which is characterised by many ropes and a few suspended platforms.

In Apenheul, two main types of inside enclosures exist. The large trunk-type inside enclosures consisted of several angled vertical large trunks, along with some horizontally placed trunks. Additionally, some horizontal ropes stretched between the trunks and a hanging wooden platform was attached to the ceiling with ropes. Differences between the two enclosures of this type (A + B; see Figure 2[a]) include a large netting structure that hung diagonally from the walls towards the trunks in enclosure A whereas B contained a number of horizontally hanging fire hoses.
The inside enclosures of the multiple ropes type (C + D; see Figure 2[b]) consisted of roughly 25 ropes attached to the ceiling, hanging vertically with a large horizontally placed pipe-frame structure at the bottom. This pipe-frame was a large 4 × 3 m (length × width) square grid-shaped structure, in which some grids were filled with mesh or a wooden platform for resting. Additionally, above this frame, at least one hanging wooden platform was attached with ropes. These two inside enclosures were very similar but not identical, as enclosure C contained a second pipe structure in the top of the enclosure, whereas enclosure D did not have this. Furthermore, enclosure D contained a large diagonally hanging net, similar to the one in enclosure A, while this was absent in enclosure C.
Data collection
In the first two weeks of the study, preliminary observations were performed to practice the observation methods and promote familiarity with the behaviour variables and individuals. These observations were not included in the analyses. Behavioural data were collected between the 6th of February and the 13th of September 2024. Observations were conducted in the morning between 1000 and 1230h and in the afternoon between 1330 and 1600h. All individuals were observed approximately equally often during different times of day (Figure S1; Supplementary material). During the feeding presentation at 1300h no observations were performed. Focal animals were recorded continuously for 10 min. A Sony HDR-CX405 camcorder (Sony Corporation, Japan) was used to record the videos. A random number generator was used to select a focal animal, without allowing for an individual to be recorded twice in a row. The behaviours from the videos were subsequently coded via continuous sampling using the computer programme BORIS (Behavioral Observation Research Interactive Software) (Friard & Gamba Reference Friard and Gamba2016). In BORIS, additional information regarding the enclosure type and whether the observation occurred inside or outside were noted. Point behaviours were only scored again when they continued after 3 s had passed following its first occurrence. Videos could be slowed down or paused to ensure accurate recording of all behaviours. When an individual moved out of sight during the recording, the observation would only be used if the individual was in sight for at least 5 min. We only included observations in which the animal was in the inside enclosure for the full duration of the observation in the analyses, because the outside enclosures at Apenheul were very similar in structural design. In the period that the orangutans had access to the outside enclosure, approximately 50% of observations still took place exclusively inside.
Behaviour variables
The ethogram used in this study was adapted from Thorpe and Crompton (Reference Thorpe and Crompton2006) and Roth et al. (Reference Roth, Bionda and Sterck2017), which described positional behaviours (Table 2). These positional behaviours were sub-divided into locomotion and postural behaviours. Locomotion behaviours were scored immediately when they occurred, and a bout would last until the individual changed to a different positional behaviour. Postural behaviours were recorded but are beyond the scope of this paper. Concurrent with these positional behaviours, the structure upon which the behaviour was performed was recorded. Seven different structures were distinguished: Ropes, Pipes, Logs, Mesh, Ground, Platforms and Other.
Ethogram used for Bornean orangutans (Pongo pygmaeus) locomotion behaviours, adapted from Thorpe and Crompton (Reference Thorpe and Crompton2006) and Roth et al. (Reference Roth, Bionda and Sterck2017)

In addition to the positional behaviours, general time budget behaviours were recorded simultaneously (Table 3). Behaviours classed as ‘Other active’ included grooming and nest building.
Ethogram used for time budget behaviours of the Bornean orangutans (Pongo pygmaeus)

Statistical analysis
We performed all analyses in R version 4.3.1 (R Core Team 2023) through RStudio version 2023.09.1+494 (Posit Team 2023). All models were created in the Stan computational framework and accessed using the brms package (Bürkner Reference Bürkner2017, Reference Bürkner2018). In all models, we used treatment coding for categorical predictors. We ran each model with four chains and 3,000 iterations, 1,000 of which were warm-ups. We checked model convergence by inspecting the trace plots, histograms of the posteriors, and Gelman-Rubin diagnostics (Depaoli & van de Schoot Reference Depaoli and van de Schoot2017). Numbers of divergent transitions or other indications of model non-convergence were not excessive. Furthermore, we used emmeans (Lenth Reference Lenth2024) to obtain contrasts and pair-wise comparisons based on the posterior predictive distribution.
First, to investigate the effect of enclosure type on the time budget, we calculated the total proportion of time spent on foraging, resting, moving, and other activity per subject per enclosure type. On these data, we performed a Dirichlet regression, which can be used to model compositional data (i.e. dependent proportions that sum to one). It captures how predictors influence the relative distribution of components within a composition (Douma & Weedon Reference Douma and Weedon2019). Such an approach is ideal for analysis of time budget data, as the categories within time budgets are not independent. For example, when an individual is moving more, they have, by definition, less time to spend on the other behavioural categories. Dirichlet regression solves this problem by modelling the relative distribution of the categories. In the model, we specified the proportion of time spent on each of the four time-budget categories as dependent variables and included enclosure type as a predictor. Furthermore, we allowed intercepts and the slope for enclosure type to vary by subject through uncorrelated random effects. We specified Gaussian priors with M = 0 and SD = 1 for all predictors, and retained default priors for all other effects (Student’s t priors with 3 degrees of freedom for the Intercepts, half-Student’s t priors with 3 degrees of freedom for all variance parameters, and a Gamma prior with shape = 0.01 and rate = 0.01 for the phi-parameter).
Second, to investigate the occurrence of different locomotion behaviours as a function of enclosure type, we calculated the proportion of time spent on each locomotion behaviour per observation. Because some locomotion behaviours were rare and thus did not occur in every observation, our data contained many zeroes. Therefore, we performed a zero-inflated beta regression per locomotion behaviour. In these models, we specified the proportion of time spent on a locomotion type (i.e. angled climbing, vertical climbing, torso-orthograde suspensory locomotion, torso-pronograde suspensory locomotion, liana sway, quadrupedal walking, bipedal walking) as dependent variable and enclosure type as predictor. Furthermore, we allowed intercepts to vary by subject. We specified the same predictors and random effects for the zero-inflation (zi) parameter but only specified a random intercept by subject for shape (phi) parameter of the models. We specified Gaussian priors with M = 0 and SD = 1 for all fixed effects across all model components, and retained default priors for all other effects (Student’s t priors with 3 degrees of freedom for the Intercepts of the mean and phi, logistic priors for the Intercepts of the zi, half-Student’s t priors with 3 degrees of freedom for all variance parameters).
Third, to explore the relationship between structure type, locomotion behaviour, and enclosure type, we first calculated how many bouts of each locomotion behaviour were performed per substrate (log, mesh, rope, pipe, plateau, ground, other) per enclosure type at the subject-level. Then, we performed four categorical regressions that modelled the relative occurrence of each substrate level being used to perform a specific locomotion behaviour on. We ran these exploratory analyses only for angled climbing, vertical climbing, torso-orthograde suspensory locomotion, and quadrupedal walking. For the former three, because we found differences in their occurrence between enclosure types in the preceding analyses, and for the latter because we had the a priori expectation that quadrupedal walking would be less common in the rope enclosures. In each model, we specified the substrate used as the dependent variable, and enclosure type as the predictor. Furthermore, we allowed intercepts and the slope of enclosure to vary by subject through correlated random effects. Regarding priors, we specified Student’s t priors with 3 degrees of freedom, M = 0, SD = 2.5 for the intercepts and retained default priors for all other parameters (Student’s t priors with 3 degrees of freedom for the intercepts, half-Student’s t priors with 3 degrees of freedom for all variance parameters, and LKJ Cholesky priors with eta = 1 for correlations between random effects).
In Results, we report contrast estimates, their 95% credible interval, and the probability of direction (pd). This latter measure of effect existence reflects the probability of a parameter being strictly positive or negative and varies between 0.5 (no clear direction) and 1 (very clear direction) (Makowski et al. Reference Makowski, Ben-Shachar, Chen and Lüdecke2019).
Ethical statement
This study did not require official ethical approval, as this study was fully observational and did not interfere with daily practices. The project was internally approved by the zoo management and welfare coordinator and adhered to the ethical agreement for research at Apenheul Primate Park.
The observations in this study were performed in the publicly accessible areas of Apenheul Primate Park, during both the visitor and non-visitor seasons. The study was purely observational and did not alter the daily lives of the animals observed or interfere with daily practices. During the non-visitor season, the orangutan building was regularly visited by guided tours and by office personnel, meaning the orangutans were used to seeing humans in the non-visitor season as well. Furthermore, the observer (JG) tried to avoid disrupting the lives of the animals by avoiding any close interaction with the animals.
Results
Time budget (quantity of locomotion)
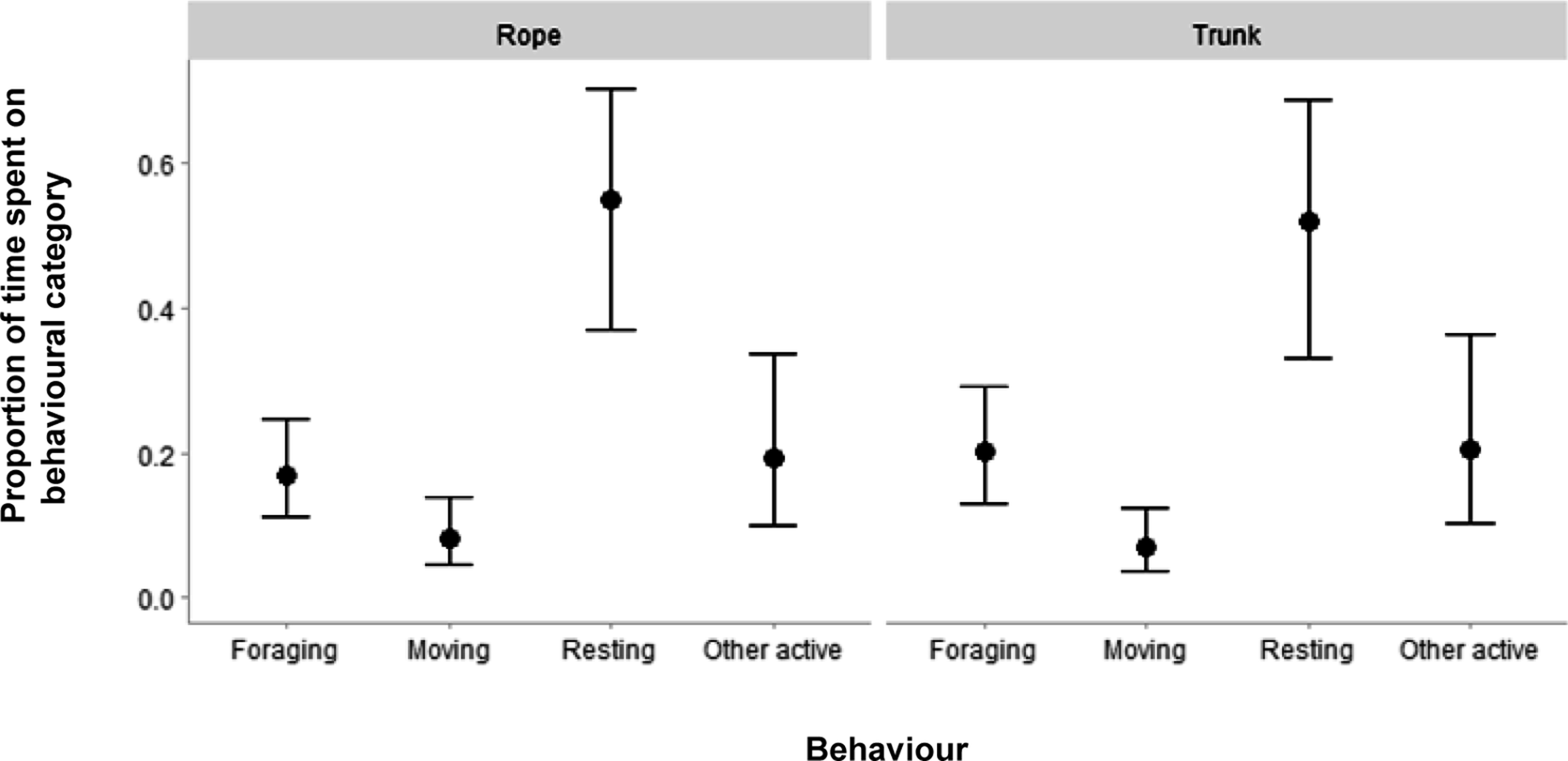
We investigated whether orangutans differed in their time budget between the Trunk and Rope enclosures by performing a Dirichlet regression with proportion of time spent on each time-budget category as dependent variable (Table S1; Supplementary material; Figure 3). The results showed no robust differences in time spent on movement between the enclosure types (ΔRope-Trunk = 0.012, 95%CrI [–0.024; 0.049], pd = 0.776), nor any differences for foraging (ΔRope-Trunk = –0.032, 95%CrI [–0.086; 0.021], pd = 0.895), resting (ΔRope-Trunk = 0.031, 95%CrI [–0.061; 0.118], pd = 0.783), and other active (ΔRope-Trunk = –0.010, 95%CrI [–0.084; 0.054], pd = 0.633).
Posterior predictions of the Dirichlet regression that models the proportion of time spent on time budget behaviours (foraging, moving, resting, and other active) for Bornean orangutans (Pongo pygmaeus) (n = 7) in two types of enclosures, either characterised by many ropes (‘Rope’) or by many trunks (‘Trunk’). Black dots show median posterior estimates and black lines reflect the 95% credible intervals.

Locomotion types (quality of locomotion)
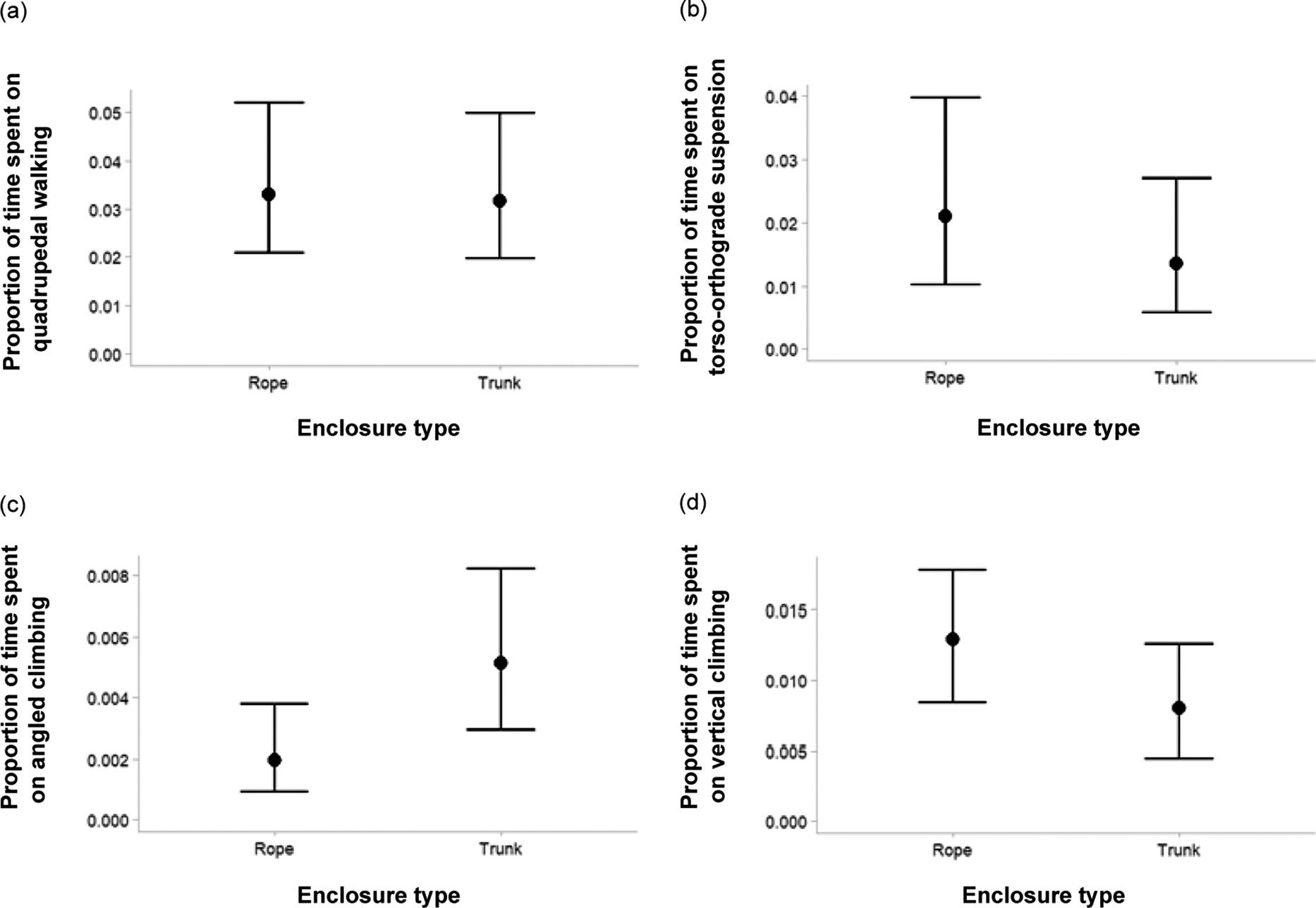
Next, we investigated how much time the orangutans spent on each of the seven locomotion types in each of the enclosure types (Table S2; Supplementary material). We used a multivariate zero-inflated beta regression model, with the proportion of observation time spent on each specific locomotion type as dependent variable. Orangutans did not differ in the time spent on quadrupedal walking between the Trunk and Rope enclosures (ΔRope-Trunk = 0.002, 95%CrI [–0.006; 0.009], pd = 0.657; Figure 4[a]). However, we did find a difference in time spent on torso-orthograde suspensory locomotion, with individuals spending more time on this locomotion type in the Rope enclosures than in the Trunk enclosures (ΔRope-Trunk = 0.008, 95%CrI [0.003; 0.016], pd = 0.999; Figure 4[b]). Furthermore, we found that orangutans showed more angled climbing in the Trunk than the Rope enclosures (ΔRope-Trunk = –0.003, 95%CrI [–0.005; –0.002], pd = 1.000; Figure 4[c]), but more vertical climbing in the Rope than the Trunk enclosures (ΔRope-Trunk = 0.005, 95%CrI [0.002; 0.008], pd = 0.999; Figure 4[d]). For the other three locomotion types, we found no substantial differences between the enclosure types (torso-pronograde suspensory locomotion: ΔRope-Trunk = –0.0002, 95%CrI [–0.002; 0.002], pd = 0.616; bipedal walking: ΔRope-Trunk = 0.001, 95%CrI [–0.003; 0.003], pd = 0.775; liana sway: ΔRope-Trunk = 0.001, 95%CrI [–0.001; 0.005], pd = 0.798).
Posterior predictions of the beta regressions that model the proportion of time spent on different locomotion types, namely (a) quadrupedal walking, (b) torso-orthograde suspension, (c) angled climbing, and (d) vertical climbing in Bornean orangutans (Pongo pygmaeus) (n = 7). Predictions are shown for the two types of enclosures, either characterised by many ropes (‘Rope’) or by many trunks (‘Trunk’). Black dots show median posterior estimates, black lines reflect the 95% credible intervals.

Substrate use
We explored the substrate use for four of the locomotion types: quadrupedal walking; torso-orthograde suspensory locomotion; angled climbing; and vertical climbing. To explore the use of different substrates for different locomotion types, we performed four categorical regression models (one for each locomotion type). Here, the specific substrate used at the locomotion bout level was the dependent variable. Thus, our analysis did not consider the duration of the locomotion behaviour.
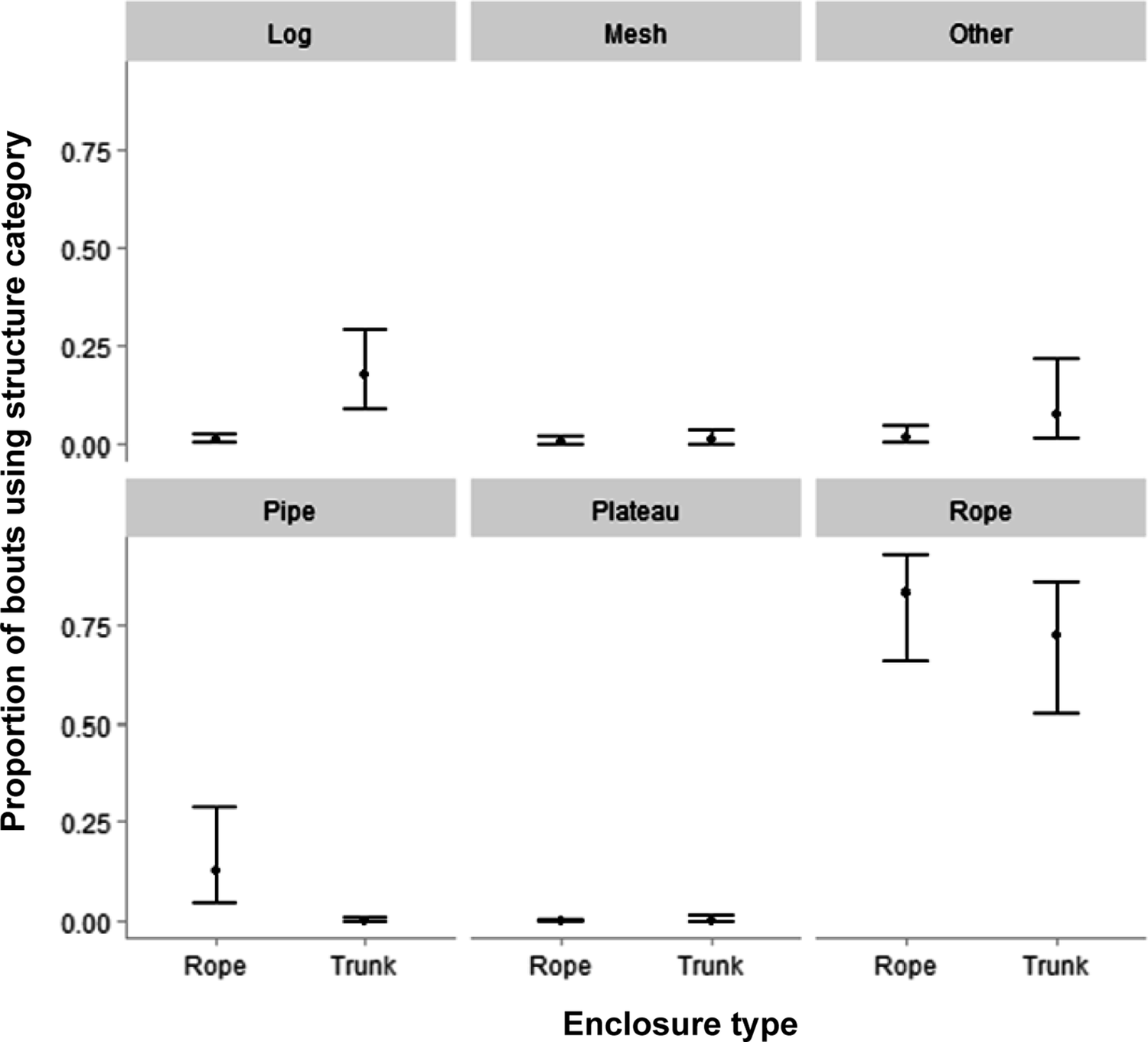
We found considerable variation in substrate use between locomotion types, but also between enclosure types (Figure 5). When looking at quadrupedal locomotion (Table S3, Figure S2; Supplementary material), we found that orangutans were more likely to perform this type of locomotion on ropes in the Trunk compared to the Rope enclosures (ΔRope-Trunk = –0.051, 95%CrI [–0.162; –0.001], pd = 0.976). However, for both enclosure types, quadrupedal walking is almost exclusively performed on ground substrate (Rope enclosure median proportion: 0.746; Trunk enclosure median proportion: 0.757). In addition, orangutans in the Rope enclosures also used pipes more often for quadrupedal locomotion, but this is a simple result of there not being any pipes available in the Trunk enclosures.
Posterior predictions from a categorical regression model of the number of bouts of torso-orthograde suspensory locomotion performed on different climbing structures in Bornean orangutans (Pongo pygmaeus) (n = 7). Predictions are shown for two enclosure types: enclosures characterised by many ropes (‘Rope’) and enclosures characterised by many trunks (‘Trunk’). Black dots show median posterior estimates, and black lines represent 95% credible intervals.

Regarding torso-orthograde suspensory locomotion (Table S4; Supplementary material; Figure 5), we found that it was most often performed on ropes in both enclosure types (Rope enclosure median proportion: 0.825; Trunk enclosure median proportion: 0.719). Interestingly, the use of ropes between enclosures was not robustly different, despite a median difference of almost 10 percentage points (ΔRope-Trunk = 0.107, 95%CrI [–0.119; 0.325], pd = 0.856). However, we found that orangutans were more likely to perform torso-orthograde suspensory locomotion on logs in the Trunk compared to the Rope enclosure (ΔRope-Trunk = –0.165, 95%CrI [–0.284; –0.077], pd = 1.00). Again, because the Trunk enclosures did not contain any pipes, a pair-wise comparison between enclosure types makes no sense here. Nevertheless, when housed in the Rope enclosures, orangutans performed torso-orthograde suspensory locomotion on pipes in almost 14% of their bouts.
Lastly, we explored substrate use for vertical and angled climbing (Tables S5 and S6, Figures S3 and S4; Supplementary material). Regarding vertical climbing, orangutans more often used ropes to perform this behaviour in the Rope than in the Trunk enclosures (ΔRope-Trunk = 0.319, 95%CrI [0.117; 0.519], pd = 0.978). We found no other clear differences between the enclosures, but orangutans regularly used logs (Rope enclosure median proportion: 0.203; Trunk enclosure median proportion: 0.330) and mesh (Rope enclosure median proportion: 0.230; Trunk enclosure median proportion: 0.395) to perform vertical climbing in both enclosure types. Angled climbing was almost exclusively performed on ropes (Rope enclosure median proportion: 0.660; Trunk enclosure median proportion: 0.509) and logs (Rope enclosure median proportion: 0.210; Trunk enclosure median proportion: 0.429) in both enclosures. Regarding between-enclosure differences, we found a difference only in the use of logs: orangutans performed angled climbing on logs more often in the Trunk than in the Rope enclosures (ΔRope-Trunk = –0.220, 95%CrI [–0.429; 0.008], pd = 0.972), although the credible interval slightly overlapped zero.
Discussion
Enabling animals to perform species-typical locomotion behaviour is essential for their well-being (Neal Webb & Schapiro Reference Neal Webb and Schapiro2023). Therefore, this study investigated differences in locomotion behaviour of zoo-housed Bornean orangutans in two different enclosure types at Apenheul Primate Park. These enclosures differed as regards the design of their climbing structures; one enclosure type contained many large firm substrates, while the other type contained more thin flexible substrates. The results show subtle differences between enclosures in the types of locomotion behaviour, but no difference in the total amount of movement. Specifically, torso-orthograde suspensory locomotion and vertical climbing occurred more often in the enclosures with a high number of vertical ropes.
Quantity of locomotion
The orangutans in all enclosure types studied spent less time on movement than has been observed in the wild. On average, the zoo-housed orangutans included in this study only spent 8% of the observed time on movement in the 6-h daily observation window, while in the wild orangutans have been observed to move for around 15% of the time in the full 12-h active period (Morrogh-Bernard et al. Reference Morrogh-Bernard, Husson, Knott, Wich, Van Schaik, Van Noordwijk, Lackman-Ancrenaz, Marshall, Kanamori, Kuze, Bin Sakong, Mitra Setia and van Schaik2009). Therefore, this study adds to the evidence of decreased levels of movement in zoo-housed orangutans, a commonly encountered issue in zoos likely due to the energy-efficient nature of orangutans (Isler & Thorpe Reference Isler and Thorpe2003; Pearson et al. Reference Pearson, Davis and Litchfield2010; Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010). Additionally, zoo-housed orangutans could be less motivated to move since they do not have to explore new areas or travel long distances to obtain resources. Such decreased levels of movement can pose health and welfare risks for the orangutans in zoos, as they are prone to obesity (Gippoliti Reference Gippoliti2000; Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010).
When comparing the enclosure types, we expected to observe more movement in the trunk enclosures than in the rope enclosures, because movement on horizontal trunks poses lower energetic costs for movement than on ropes. However, no meaningful differences in the amount of movement or any other time-budget behaviour were found. This was surprising, because previous research had shown that more movement occurred in an enclosure with a considerable number of horizontal trunks, compared to an enclosure with many ropes (Roth et al. Reference Roth, Bionda and Sterck2017). This contradiction is especially noteworthy, because the 2017 study in question actually took place in the same zoo, however only included the outside enclosures. Perhaps this indicates a difference between the inside and outside enclosures. However, this previous result could also have been a consequence of habituation to the new enclosures, as at the time of the previous study the trunk enclosures had only recently been remodelled. Previous research has indicated that orangutans do show signs of gradual habituation to their enclosure over a period up to three years, reducing the differences in the activity patterns between different enclosures (White et al. Reference White, Houser, Fuller, Taylor and Elliott2003). While Roth et al. (Reference Roth, Bionda and Sterck2017) did test for habituation, this was restricted to a period of seven weeks. Perhaps in the intervening years the orangutans did further habituate to the remodelled trunk enclosures, decreasing their exploratory behaviour and thus the amount of movement.
Quality of locomotion
We found a difference in the amount of torso-orthograde suspensory locomotion between enclosures. As expected, this locomotion behaviour occurred more often in the rope than in the trunk enclosures. This is in line with observations from the wild, that have linked this locomotion behaviour to thin flexible structures, such as lianas and vines (Thorpe et al. Reference Thorpe, Holder and Crompton2009; van Casteren et al. Reference van Casteren, Sellers, Thorpe, Coward, Crompton and Ennos2013). However, the zoo-housed orangutans still performed this type of locomotion less often than their wild counterparts, for whom it is the dominant form of movement (Thorpe & Crompton Reference Thorpe, Crompton, Wich, Atmoko, Setia and van Schaik2008). Yet our study confirms that in a zoo setting this locomotion behaviour is also associated with these thin flexible structures, as it nearly exclusively occurred on ropes. Interestingly, we found no robust difference between the enclosures regarding the proportion of bouts in which ropes were used to perform this behaviour. So, while torso-orthograde locomotion was more common in the rope enclosure, we cannot definitively attribute this difference to an increased usage of ropes. Moreover, our analyses on structure use were exploratory and looked only at bouts performed on each structure, but not the durations of these bouts. Nevertheless, ropes seem to facilitate torso-orthograde locomotion, given the large proportion of bouts performed on ropes.
While provision of plenty of thin flexible structures does increase the amount of torso-orthograde suspensory locomotion, it does still fail to stimulate natural levels of torso-orthograde locomotion in zoos. Thus, further efforts are necessary to stimulate orangutans to use more torso-orthograde suspensory locomotion. For example, providing foraging opportunities or other enrichment items that can only be reached by using the ropes could further increase the occurrence of this locomotion behaviour. Previous research has already stressed the importance of foraging opportunities in combination with movable structures, such as ropes, for the activity of zoo-housed orangutans (Tripp Reference Tripp1985). Additionally, provisioning enrichment items can alter the space use of animals in their enclosures, as has been observed in several species, including kinkajous (Potos flavus), brown bears (Ursus arctos), and chimpanzees (Pan troglodytes) (Yamanashi et al. Reference Yamanashi, Matsunaga, Shimada, Kado and Tanaka2016; Blount & Taylor Reference Blount and Taylor2000; Puehringer-Sturmayr et al. Reference Puehringer-Sturmayr, Fiby, Bachmann, Filz, Grassmann, Hoi, Janiczek and Frigerio2023). Furthermore, gorillas (Gorilla gorilla) and chimpanzees show very selective enclosure use and only occupy areas in which there is something useful, such as foraging or resting opportunities (Ross et al. Reference Ross, Calcutt, Schapiro and Hau2011). Therefore, dispersing resources and enrichment in such a way that orangutans are encouraged to move arboreally throughout the enclosure and engage with different elevated structures seems important. This strategy might at the same time increase locomotion quantity, as dispersion of food resources requires orangutans to move more to obtain the same amount of food. Thus, this strategy might promote a more naturalistic use of vertical space, increase the occurrence of torso-orthograde suspensory locomotion and possibly other locomotion types, such as tree swaying, and increase locomotion quantity overall.
Against our expectations, we found no meaningful difference in the proportion of time spent on quadrupedal walking between enclosure types. Quadrupedal walking is a locomotion behaviour that had been shown to be associated with firm structures in the wild (Thorpe et al. Reference Thorpe, Holder and Crompton2009). Therefore, we expected to find this behaviour more in the trunk enclosures. As mentioned before, quadrupedal walking is especially common in zoos, putting unnatural pressures on the muscles and skeleton of the orangutans (Isler & Thorpe Reference Isler and Thorpe2003). In the current, zoo-based study, quadrupedal walking was found to occur mostly on the ground. However, this gait also occurred on logs and plateaux. This shows that zoo-housed and wild orangutans use similar structures for quadrupedal walking, as previous studies have found that quadrupedal walking was strongly associated with firm structures in the wild (Thorpe et al. Reference Thorpe, Holder and Crompton2009; van Casteren et al. Reference van Casteren, Sellers, Thorpe, Coward, Crompton and Ennos2013).
Quadrupedal walking was found to be the most commonly used form of locomotion across both enclosure types. This result is in line with previous research on zoo-housed orangutans (Isler & Thorpe Reference Isler and Thorpe2003; Pearson et al. Reference Pearson, Davis and Litchfield2010). This is in stark contrast to the locomotive patterns displayed by wild orangutans, who rarely use quadrupedal walking (Cant Reference Cant1987; Thorpe & Crompton Reference Thorpe and Crompton2005, Reference Thorpe and Crompton2006; Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). The increased usage of quadrupedal walking and decreased usage of suspensory locomotion is likely a consequence of the energy efficient nature of orangutans and lack of motivations to leave the ground in zoos. Quadrupedal walking is relatively low in its energetic costs, which causes them to use quadrupedal walking more often when provided with structures allowing for this type of locomotion (Isler & Thorpe Reference Isler and Thorpe2003; Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010; Hanna & Schmitt Reference Hanna and Schmitt2011). Additionally, as quadrupedal walking occurred mostly on the ground, the overperformance of this behaviour is likely a consequence of the increased terrestriality of zoo-housed orangutans. This could be a consequence of the lack of motivation to ascend into the climbing structures, because there is for instance no need to avoid predators and there is only limited food presence at these heights. Therefore, in addition to providing more elevated feeding opportunities as motivators to use the climbing structures, making the ground floor unattractive could perhaps reduce the usage of this locomotion behaviour. Previous research indeed showed that in an enclosure with an unattractive ground floor, orangutans spent significantly more time in the elevated areas of the enclosure and were more active overall (Hebert & Bard Reference Hebert and Bard2000). Potentially, such interventions can also stimulate more species-typical and challenging locomotion behaviour, although it could also be difficult to implement from a practical or ethical perspective.
Lastly, we found differences in the occurrence of two climbing behaviours between the enclosures, with vertical climbing occurring more in the rope enclosures and angled climbing occurring more in the trunk enclosures. This is likely a logical consequence of the angle of the structures that are present in the respective enclosures. The ropes in the trunk enclosure were vertically aligned, while the trunks in the trunk enclosure were mostly angled. We found that both climbing behaviours were most often performed on ropes, but also frequently occurred on logs. Furthermore, vertical climbing occurred more on ropes in the rope enclosure. This indicates an association between this form of locomotion and vertical ropes. Vertical climbing has been shown to be one of the most energetically costly locomotion behaviours, while angled climbing is presumed to require somewhat less energy (Isler & Thorpe Reference Isler and Thorpe2003). Therefore, stimulating these behaviours could be beneficial, as they may provide the orangutans with a greater physical challenge thereby potentially enhancing their physical health. By increasing the number of vertical ropes, the most challenging locomotion behaviour could increase in occurrence.
Animal welfare implications and conclusion
The findings of this study could be used to improve the welfare of zoo-housed orangutans, as we have shown that orangutans displayed somewhat more species-typical and challenging forms of locomotion in the rope enclosure type. These consisted of torso-orthograde suspensory locomotion (the most commonly used locomotion in the wild) and vertical climbing, which is the most energy costly form of locomotion (Isler & Thorpe Reference Isler and Thorpe2003; Manduell et al. Reference Manduell, Morrogh-Bernard and Thorpe2011). The increased usage of these challenging species-typical behaviours could positively impact the welfare of the orangutans. Nevertheless, these natural locomotion behaviours were still less common than in wild orangutans. As discussed previously, further increasing the amount of thin flexible structures and reducing the attractiveness of the ground level can further stimulate orangutans to use more natural forms of locomotion and spend more time above-ground. However, such measures are likely to be effective only when paired with motivational stimuli. For example, providing a large portion of their daily food in elevated parts of the enclosure encourages orangutans to use structures that promote natural locomotion. Due to their energy-efficient nature (Pontzer et al. Reference Pontzer, Raichlen, Shumaker, Ocobock and Wich2010), orangutans may make little use of climbing structures if there is no clear goal to achieve. Thus, stimulating the orangutans by providing naturalistic climbing structures and relevant motivational stimuli can benefit their welfare, especially when combined with providing more mental challenges (Chappell et al. Reference Chappell, Phillips, van Noordwijk, Mitra Setia and Thorpe2015; Veasey et al. Reference Veasey, Armstrong, Butcher, Embury, Grossfeldt, Robbins and Thompson2025), and help to offset their efficient nature.
Fully mimicking the behavioural patterns of wild orangutans in zoos is almost impossible, as the constraints of captivity and other factors such as presence of caretakers and visitors will inevitably impact the behavioural repertoire of zoo-housed orangutans. That said, the extent to which the natural locomotion patterns should be replicated in zoos is a matter for debate, as there is also less functional requirement for certain behaviours in zoos (Veasey Reference Veasey2017). Stimulating natural locomotion should not be a goal per se but should be used as a tool to increase the welfare state of zoo-housed orangutans. While definitions of animal welfare in general emphasise the ability to perform species-typical behaviour, most do not include clearly defined thresholds for the quantity of such behaviours (Mellor & Reid Reference Mellor and Reid1994; Mellor & Beausoleil Reference Mellor and Beausoleil2015; Arndt et al. Reference Arndt, Goerlich and van der Staay2022). If an animal displays its full locomotive behavioural repertoire, but some behaviours are rarer than in the wild, this does not pose a welfare problem per se.
At the same time, quantity of locomotion has been shown to correlate with the overall scratching rate, inactivity and behavioural diversity in chimpanzees, where scratching and inactivity decreased and behavioural diversity increased with increased locomotion (Neal Webb & Schapiro Reference Neal Webb and Schapiro2023). While these three factors are often considered proxies for animal welfare, it is crucial to recognise the non-linear nature of the relationship between behavioural diversity and animal welfare (Cronin & Ross Reference Cronin and Ross2019). Despite this complex relationship, the observed correlations could imply that chimpanzees with high levels of locomotion experience a more positive welfare state. Therefore, Neal Webb and Schapiro (Reference Neal Webb and Schapiro2023) argued that, for chimpanzees, the amount of locomotion could perhaps be considered a positive welfare indicator in zoos. While such correlations are yet to be established for orangutans, similar relationships between locomotion and potential welfare proxies might exist in orangutans considering the aforementioned negative welfare implications of inactivity, such as reduced musculoskeletal health (Isler & Thorpe Reference Isler and Thorpe2003). Further research could investigate the relationship between the amount of locomotion, diversity of locomotion behaviour, and well-established welfare indicators.
In conclusion, enclosure design can stimulate more natural and challenging types of locomotion in zoo-housed Bornean orangutans. We found that orangutans performed more torso-orthograde suspensory locomotion when supplied with a large amount of vertically hanging ropes. However, this did not increase the time spent on movement and did not reduce the occurrence of quadrupedal walking. Therefore, further research is needed to investigate how these energy-efficient animals can be stimulated to move more, for example, through targeted placement of enrichment and resources critical for survival.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/awf.2026.10086.
Data availability statement
The complete datafile and analysis scripts are available through DataverseNL: https://doi.org/10.34894/NKB9A6.
Acknowledgements
We want to thank the orangutan caretakers at Apenheul for their support of our study. Furthermore, we thank four anonymous reviewers for their comments on the manuscript.
Competing interests
None.







 Open access
Open access