


Introduction
Global climate change continues to increase temperatures and the frequency of extreme events such as droughts, floods, and heat waves (IPCC, Reference Lee and Romero2023). Climate analyses and forecasts indicate that mid- and low-latitude areas will be affected by more severe droughts and more intense temperature fluctuations, generating conditions that negatively impact crops, including grapevines. The changes that have occurred up to the present have affected the phenology, physiology, yields, and fruit quality (Domínguez et al., Reference Domínguez, Cirrincione, Deis and Martínez2024; Fraga et al., Reference Fraga, Malheiro, Moutinho-Pereira and Santos2013; Gambetta et al., Reference Gambetta, Herrera, Dayer, Feng, Hochberg and Castellarin2020). Several crop management approaches mitigate these effects; however, the rapid pace of climate change exceeds the capacity of current grapevine management practices (IPCC, Reference Lee and Romero2023).
Moreover, these different crop management approaches have been shown to have limitations primarily due to biological constraints. Among the longer-term options, genotype modification (more tolerant cultivars and rootstocks), selection of new cultivars, and genetic improvement are among the most efficient options (Chen et al., Reference Chen, Fei, Pang, Krstic, Clingeleffer, Howell, Chen and Zhang2024; Serrano-Parra et al., Reference Serrano Parra, Martínez Gascueña, Alonso, Cebrián Tarancón, Mena Morales and Chacón Vozmediano2024). The use of rootstocks is a useful strategy to enhance tolerance to pests, salinity, soil types, and drought (Serrano-Parra et al., Reference Serrano Parra, Martínez Gascueña, Alonso, Cebrián Tarancón, Mena Morales and Chacón Vozmediano2024). However, various rootstocks have shown variable responses depending on the grafted scion. Studies conducted with rootstocks 1103P and 101-14MGt demonstrated that, under severe stress (20% of soil water capacity), 1103P activated a drought evasion strategy by stomatal conductance reduction through increased abscisic acid (ABA) in the roots. Meanwhile, 101-14MGt maintained greater photosynthesis, adopting a tolerance strategy (Bianchi et al., Reference Bianchi, Ricciardi, Pozzoli, Grossi, Caramanico, Pindo, Stefani, Cestaro, Brancadoro and De Lorenzis2023). Knowing the different mechanisms by which different rootstocks respond to drought is essential for improved selection. However, no comprehensive synthesis currently integrates xylem anatomy, aquaporin regulation, and rootstock genetics to explain drought adaptation in Vitis. This review aims to synthesise current knowledge on grapevine rootstock responses to drought and heat stress by integrating physiological, hydraulic, molecular, and breeding knowledge with a focus on the mechanisms underlying drought and heat tolerance.
Advances in rootstock breeding
In recent decades, the breeding of grapevine rootstocks has intensified to address the effects of climate change and soil pests. Three initiatives stand out for their international relevance: the M series in Italy, the RG series in Spain, and the C series in Australia. The M series, developed by the University of Milan in collaboration with local nursery growers, is the result of more than twenty years of selection and has been evaluated at multiple sites across the Italian peninsula. Its objective is to combine tolerance to water and heat stress with improvements in vigour and oenological quality. Rootstocks 161-49C, 41-BMgt, and 420A Mgt modulate sugar accumulation and pH during ripening, providing advantages under high-temperature conditions. In contrast, 140Ru and 5-BB give rise to musts with higher pH and sugar content, which is a disadvantage for quality wine production (Santesteban et al., Reference Santesteban, Rekarte, Torres, Galar, Villa-Llop, Visconti, Intrigliolo, Escalona, de Herralde and Miranda2023). Among the M series, M4 shows moderate vigour and greater water-use efficiency, accelerating Cabernet Sauvignon ripening compared to 1103P (Corso et al., Reference Corso, Vannozzi, Ziliotto, Zouine, Maza, Nicolato, Vitulo, Meggio, Valle, Bouzayen, Müller, Munné-Bosch, Lucchin and Bonghi2016). The RG series, developed by the Vitis Navarra Nursery, has evaluated different genotypes (RG2–RG9) in Syrah and Tempranillo for four years. Some rootstocks demonstrated a good balance between yield and phenolic quality, while in water deficit conditions they did not outperform commercial reference rootstocks (Marin et al., Reference Marín, Miranda, Abad, Urrestarazu, Mayor, Villa-Llop and Santesteban2023). The C series, developed by CSIRO (Australia), was designed to resist phylloxera and nematodes, as well as extreme heat and drought conditions (Chen et al., Reference Chen, Fei, Pang, Krstic, Clingeleffer, Howell, Chen and Zhang2024). C20 and C114 showed good performance under heat and drought conditions (Clingeleffer et al., Reference Clingeleffer, Morales, Davis and Smith2019).
Over the past few decades, advances in genomics and transcriptomics have transformed rootstock selection and breeding strategies. Multiple crosses have enabled genome assemblies from different Vitis species and cultivars targeting water and heat tolerance (Cantu et al., Reference Cantu, Massonnet and Cochetel2024). At the population level, association studies and QTL analyses (GWAS/QTL mapping) have identified genomic regions associated with root morphology, yield, and water stress tolerance-related traits, facilitating the incorporation of markers into marker-assisted selection (MAS) programmes and genomic prediction. Transcriptomics analyses (RNA-seq) have been used for its contributions to the temporal and tissue-specific resolution required to elucidate molecular mechanisms (e.g., regulation of aquaporins, antioxidant pathways, and hormonal signalling networks (ABA/ET/JA)) in response to drought and high temperatures (Labarga et al., Reference Labarga, Mairata, Puelles, Martín, Albacete, García-Escudero and Pou2023). A comparative study of rootstocks and scions has shown that the rootstock genotype and phenotype can drive changes in the roots and vascular tissue of the scion, such as the regulation of aquaporins and transcription factors (Harris et al., Reference Harris, Pratt, Kovacs, Klein, Kwasniewski, Londo, Wu and Miller2023). In grapevine, editing protocols have been successfully implemented (including DNA-free methods using ribonucleoprotein delivery) and have reported successful edits that confer resistance to pathogens and allow for the modification of metabolic pathways and stress responses. However, direct application to grapevine rootstocks remains limited (Cantu et al., Reference Cantu, Massonnet and Cochetel2024; Najafi et al., Reference Najafi, Bertini, D’Incà, Fasoli and Zenoni2022). Future research priorities include (1) characterising the performance of diverse rootstocks under heat and drought conditions and expanding integrated databases and (2) exploiting the genetic diversity of Vitis species by identifying favourable alleles for incorporation into conventional marker-assisted breeding programmes.
Physiological mechanisms underlying tolerance
Plants have tolerance mechanisms that allow them to mitigate the effects of drought and heat. One key strategy is the regulation of stomatal opening. In hot and dry climates, stomatal closure is vital for preserving water in plant tissues and also prevents the formation of air bubbles (embolisms) in the xylem that could block water transport from the soil throughout the plant (Alsina et al., Reference Alsina, Smart, Bauerle, de Herralde, Biel, Stockert, Negron and Save2011). To date, the strategies used by rootstocks to tolerate thermal and water stress are not yet fully known (de Souza et al., Reference de Souza, Gindro, Verdenal, Spring, Spangenberg and Zufferey2022).
Studies on the rootstocks 1103P and 101-14 Mgt, grafted with Cabernet Sauvignon, under different levels of water deficit (80%, 50%, and 20% soil water content, SWC), demonstrated that under severe water deficit, the rootstock genotype significantly influenced scion gas exchange. The 1103P rootstock displayed a drought-avoidance strategy, characterised by reduced stomatal conductance and photosynthetic inhibition, associated with an increase in root ABA content, which promoted stomatal closure in leaves. By contrast, 101-14 Mgt maintained relatively high photosynthetic rates under the same conditions (Martin-StPaul et al., Reference Martin-StPaul, Delzon and Cochard2017; Padgett-Johnson et al., Reference Padgett-Johnson, Williams and Walker2000).
Other genotypes, however, show slight or no stomatal regulation, which can result in an increased risk of dehydration or a marked reduction in plant growth (Zang et al., Reference Zhang, Marguerit, Rossdeutsch, Ollat and Gam-betta2016). Based on this behaviour, plants, and consequently rootstocks, can be classified as isohydric or anisohydric. Rootstocks such as 110R exhibited isohydric behaviour, whereas 1103P and SO4 were typically anisohydric. The latter allow greater variation in plant water status while keeping their stomata open for longer periods under drought conditions, thereby showing a tolerance-type response to water stress. Each strategy has advantages and disadvantages depending on the environment and growing conditions.
Rootstocks influence not only vine water status but also yield, grape quality, and the vine’s resilience under increasingly challenging climatic scenarios (Ollat et al., Reference Ollat, Peccoux, Papura, Esmenjaud, Marguer-it, Tandonnet, Bordenave, Cookson, Barrieu, Rossdeutsch, Geros, Manuela, Gil and Delrot2016). Comparing 140Ru, RG8, 110R, and SO4 grafted with Tempranillo, the highest values of net assimilation and gas exchange were found in 140Ru. Under these conditions, it presented a lower negative water potential.
The RG8 rootstock is classified as vigorous and exhibits high hydraulic conductivity, with K plant values of 75 mmol H2O plant−1 s−1 MPa−1 at a predawn water potential of −0.19 MPa and 21 mmol H2O plant−1 s−1 MPa−1 at −0.33 MPa, together with high gas-exchange rates. In contrast, the 110R rootstock showed lower net assimilation and stomatal conductance than the other rootstocks, with K plant values of 56 mmol H2O plant−1 s−1 MPa−1 at a predawn water potential of −0.20 MPa and 11 mmol H2O plant−1 s−1 MPa−1 at −0.30 MPa. Nevertheless, net assimilation rates remained relatively high, averaging 10.1 µmol CO2 m−2 s−1. Finally, water-use efficiency is used to classify rootstock drought tolerance and is related to stomatal behaviour, root morphology, and hydraulic conductance (Buesa et al., Reference Buesa, Torres, Tortosa, Marín, Villa-Llop, Douthe, Santesteban, Medrano and Escalona2023; Villalobos-González et al., Reference Villalobos-González, Muñoz-Araya, Franck and Pastenes2019; Zúñiga et al., Reference Zúñiga, Ortega-Farías, Fuentes, Riveros-Burgos and Poblete-Echeverría2018). With this information, it is possible to choose rootstocks according to water availability.
Root system morphology and hydraulics
Root system morphology regulates plant water status (Alonso-Forn et al., Reference Alonso-Forn, Buesa, Flor, Sabater, Medrano and Escalona2025; Hochberg et al., Reference Hochberg, Herrera, Cochard and Badel2016; Martin-StPaul et al., Reference Martin-StPaul, Delzon and Cochard2017). The diameter and density of xylem vessels have been studied in 5 rootstocks: Fercal, RG4, 110R, 140Ru, and SO4. This study found important differences between these rootstocks that could double the diameter and densities of vessels between different rootstocks. Fercal presented diameters of 48 µm with densities of 55 vessels.mm−2, while RG4 with 79 µm and 40 vessels.mm−2, 110R 90 µm and 25 vessels.mm−2, 140Ru 50 µm and 65 vessels.mm−2, and SO4 58 µm 75 vessels.mm−2 (Ramsing et al., Reference Ramsing, Gramaje, Mocholí, Agustí, Cabello Sáenz de Santa María, Armengol and Berbegal2021). Based on these data, Fercal had 11.4% of the total cross-sectional area occupied by vessels: RG4 9.94%, 110R 4.42%, 140Ru 16.59%, and SO4 26.07%. There does not appear to be any relationship between this area and its drought tolerance, as 140Ru and 110R are reported to have the highest tolerance. Rootstocks obtained from crosses between Vitis berlandieri × NovoMexicana, V. riparia, V. riparia × V. berlandieri and V. rupestris also showed statistically different xylem vessel diameters (40–64 µm), with densities ranging from 45 to 80 vessels.mm−2 (Bauerle et al., Reference Bauerle, Centinari and Bauerle2011; Ramsing et al., Reference Ramsing, Gramaje, Mocholí, Agustí, Cabello Sáenz de Santa María, Armengol and Berbegal2021). The characteristics of xylem vessels determine hydraulic efficiency and resistance to embolism (De Herralde et al., Reference De Herralde, del Mar Alsina, Aranda, Savé and Biel2006; Pouzoulet et al., Reference Pouzoulet, Pivovaroff, Santiago and Rolshausen2014; Sorek et al., Reference Sorek, Greenstein, Netzer, Shtein, Jansen and Hochberg2021; Zambonini et al., Reference Zambonini, Savi, Rosner and Petit2024). Under drought conditions or high-water demand, embolism can occur in xylem vessels, interrupting water flow to the rest of the plant. While hydraulic efficiency is the capacity of xylem vessels to transport water from the root to aerial organs in the soil-plant-atmosphere continuum along a water potential gradient (Quintana-Pulido et al., Reference Quintana-Pulido, Villalobos-Gonzalez, Muñoz, Franck and Pastenes2018). This hydraulic efficiency depends on hydraulic conductance (Kh), modulated by genotype and water regime, as well as by root morphology. Fine and dense roots have been shown to promote better absorption under drought conditions by exploring larger soil volumes. In addition, deep root systems increase access to water reserves. However, suberisation of the endodermis and exodermis can limit hydraulic conductance under conditions of water deficit (Alsina et al., Reference Alsina, Smart, Bauerle, de Herralde, Biel, Stockert, Negron and Save2011; Birouste et al., Reference Birouste, Zamora-Ledezma, Bossard, Pérez-Ramos and Roumet2014). Water uptake at the cellular level is mediated by aquaporins, thereby regulating this flow (Barrowclough et al., Reference Barrowclough, Peterson and Steudle2000).
Rootstock-scion interactions alter the scion’s hydraulic characteristics, such as the anatomy of the xylem vessels (diameter, number, and density), which subsequently affects the scion’s hydraulic conductivity and gas exchange. While these anatomical characteristics are expressed in the scion’s xylem, they reflect the influence of the rootstock genotype through physiological and anatomical modulation. For example, Santarosa et al. (Reference Santarosa, de Souza, Mariath and Lourosa2016) and Loupit and Cookson (Reference Loupit and Cookson2020) demonstrated that rootstock-scion interactions modify the characteristics of the scion’s xylem vessels (diameter, density, and total xylem area) in grafted grapevines, indicating that these characteristics are observable in the scion but are conditioned by the rootstock genotype (Pérez-Álvares et al., Reference Pérez-Álvarez, Intrigliolo, Martínez-Moreno, García-Sánchez, Parra, Alfosea-Simon and Buesa2023; Zufferey et al., Reference Zufferey, Spring, Verdenal, Dienes, Belcher, Lorenzini, Koestel, Rösti, Gindro, Spangenberg and Viret2017). Additionally, comparative studies under water deficit have shown rootstock-dependent differences in scion hydraulic conductivity correlated with xylem anatomy and vessel frequency, implying that anatomical and functional hydraulic differences measured in the scion originate from rootstock effects (de Souza et al., Reference de Souza, Gindro, Verdenal, Spring, Spangenberg and Zufferey2022).
Furthermore, comparative studies under water deficit conditions have shown that differences in scion hydraulic conductivity (Kpetiole and whole-plant Kh) dependent on the rootstock correlate with xylem anatomy and vascular frequency, implying that the anatomical and functional hydraulic differences measured in the scion originate from rootstock effects. Comparisons of hydraulic conductivity between rootstocks (1103P, 101-14Mgt, Kober 5BB, 3309C, and 1103P) found that the 101-14Mgt rootstock had the highest hydraulic conductance. This has been associated with its greater vigour. Greater water availability leads to greater biomass production if all other factors are optimal. On the other hand, Kober 5BB or 3309C presented high petiolar Kh values under mild water stress conditions (de Souza et al., Reference de Souza, Gindro, Verdenal, Spring, Spangenberg and Zufferey2022). The relationship between the biomass produced by photosynthesis and the amount of water consumed is known as water-use efficiency (WUE) (Bianchi et al., Reference Bianchi, Grossi, Tincani, Simone Di Lorenzo, Brancadoro and Rustioni2018; Bianchi et al., Reference Bianchi and Brancadoro2021). WUE is an indicator of the degree of plant adaptation to water deficit. It generally increases under drought conditions (Pou et al., Reference Pou, Flexas, Alsina Mdel, Bota, Carambula, de Herralde, Galmes, Lovisolo, Jimenez and Ribas-Carbo2008); existing variations between rootstocks: 110R, M1, and M4 increase their WUE under limiting conditions, while SO4 maintains lower values (Frioni et al., Reference Frioni, Biagioni, Squeri, Tombesi, Gatti and Poni2020; Mirás-Avalos et al., Reference Mirás-Avalos and Araujo2021).
Under the same study conditions, 1103P showed a stem potential of −0.85 (MPa), net assimilation 15 µmol CO2.m−2.s−1, and WUEi 70 µmol CO2⋅mol H2O. Similar values were found for SO4 at −0.84 MPa; 17 µmol CO2.m−2.s−1 and 69 µmol CO2⋅mol H2O. The least efficient rootstocks were 140Ru, 110R, and RG8, with values of 51.6 μmol CO2·mol H2O, 58.52 μmol CO2·mol H2O and 50 μmol CO2·mol H2O, corresponding to a WUE and stem potential of −0.85; −0.69; −0.57 MPa.
Studies conducted in Syrah and Sangiovese showed that low-vigour rootstocks increased water efficiency (Edwards et al., Reference Edwards, Betts, Clingeleffer and Walker2022), while in Sangiovese, M4 improved both water efficiency and grape quality under drought conditions (Frioni et al., Reference Frioni, Biagioni, Squeri, Tombesi, Gatti and Poni2020). Overall, these results confirm that selecting rootstocks that enhance vineyard water-use efficiency represents a highly effective strategy to mitigate the impacts of drought and heat waves in viticulture. Maintaining root biomass under low water availability conditions permits plants to increase their water absorption capacity, as is the case with the M4 rootstock. Contrarily, SO4 reduced root growth (Pérez-Alvarez et al., Reference Pérez-Álvarez, Intrigliolo, Martínez-Moreno, García-Sánchez, Parra, Alfosea-Simon and Buesa2023). The morphological and physiological characteristics of the roots become more important when the vineyard is irrigated using drip irrigation. In the case of Riparia Gloire, it did not modify root diameter, showing low root system plasticity (Bonarota et al., Reference Bonarota, Toups, Bristow, Santos, Jackson, Cramer and Barrios-Masias2024).
Molecular mechanism
Aquaporins are transmembrane proteins that facilitate the transport of water and small solutes. They are classified into several subfamilies, including plasma membrane intrinsic proteins (PIPs), which can also transport H2O2 and boron (Hove and Bhave, Reference Hove and Mand Bhave2011; Priya et al., Reference Priya, Dhanker, Siddique, HanumanthaRao, Nair, Pandey, Singh, Varshney, Prasad and Nayyar2019). Another family comprises tonoplast intrinsic proteins (TIPs). Five types have been identified in grapevines, where they facilitate water movement between the cytosol and the vacuole and act as cytoplasmic osmoregulators by facilitating the transport of small ions and solutes (Zwieniecki and Secchi, Reference Zwieniecki, Secchi, Chaumont and Tyerman2017). A third family present in grapevines is the nodulin 26-like intrinsic proteins (NIPs), aquaglyceroporins, with seven types found in the plasma membrane.
Aquaporin expression varies depending on both the level of water stress and genotype. In grapevines grafted onto 1102P, the aquaporins VvPIP and VvTip have been reported to be upregulated, whereas they are downregulated in plants grafted onto 161-49C (Labarga et al., Reference Labarga, Mairata, Puelles, Martín, Albacete, García-Escudero and Pou2023). Overexpression of aquaporins in 1103P improved drought tolerance by enhancing water uptake capacity under stress conditions (Galmés et al., Reference Galmés, Pou, Alsina, Tomàs, Medrano and Flexas2007; Haider et al., Reference Haider, Zhang, Kurjogi, Pervaiz, Zheng, Zhang, Lide, Shangguan and Fang2017). Aquaporin expression also plays an important role during post-drought recovery, VvTIP1 and VvTIP2;1 increased after rewatering following a drought stress (Galaz et al., Reference Galaz, Pérez-Donoso and Gambardella2024). Under severe and prolonged water stress, aquaporin activation ensures continued water absorption, making the rootstock more tolerant to drought, as is the case with 1103P (Galmés et al., Reference Galmés, Pou, Alsina, Tomàs, Medrano and Flexas2007; Zarrouk et al., Reference Zarrouk, Garcia-Tejero, Pinto, Genebra, Sabir, Prista, David, Loureiro-Dias and Chave2016).
In addition, a positive correlation has been detected between VvTIP2;1 expression and stomatal conductance under water stress conditions (Galaz et al., Reference Galaz, Pérez-Donoso and Gambardella2024), highlighting the role of TIP aquaporins in regulating cellular osmoregulation. Table 1 shows the various types of aquaporins, their location, general function, and their participation in regulation under water stress conditions.
Classification, localisation, and physiological roles of aquaporins involved in water stress regulation

The aquaporins facilitate the movement of water and solutes from the vacuole to the cytoplasm during periods of high-water demand or recovery from drought (Maurel et al., Reference Maurel, Verdoucq, Luu and Santoni2008; Vandeleur et al., Reference Vandeleur, Sullivan, Athman, Jordans, Gilliham, Kaiser and Tyerman2009). Besides water, these aquaporins can transport other polyalcohols such as mannitol and myo-inositol that act as compatible solutes (Aubert et al., Reference Aubert, Gout, Bligny and Douce1994). These osmolytes promote the osmotic adjustment, allowing continued water absorption under drought conditions.
Rootstocks 1103P and 110R show a greater capacity for osmotic adjustment under drought conditions, maintaining cellular integrity and the transpiration rate in isohydric cultivars. Additionally, greater drought resistance leads to greater aquaporin expression in the meristematic and root elongation zones, allowing for greater water uptake (Siefritz et al., Reference Siefritz, Tyree, Lovisolo, Schubert and Kaldenhoff2002). Accordingly, Cabernet Sauvignon grafted onto 1103P and 101-14 showed increased expression of aquaporins VvPIP2;1 and VvPIP2;2, which has been associated with anisohydric behaviour (Galaz et al., Reference Galaz, Pérez-Donoso and Gambardella2024; Sade et al., Reference Sade, Moshelion, Chaumont and Tyerman2017; Zarrouk et al., Reference Zarrouk, Garcia-Tejero, Pinto, Genebra, Sabir, Prista, David, Loureiro-Dias and Chave2016), maintaining their water balance compared to 161-49C (Gambetta et al., Reference Gambetta, Herrera, Dayer, Feng, Hochberg and Castellarin2020).
Aquaporin-mediated hydraulic regulation is closely linked to phytohormonal signalling. Rootstocks indirectly regulated osmotic adjustment capacity through the coordinated control of aquaporin expression and ABA balance. The interaction between rootstock-scion and drought tolerance is further modulated by plant hormones such as ABA and ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC). Under drought conditions, increased ABA induces stomatal closure and a decrease in hydraulic conductance (Tombesi et al., Reference Tombesi, Nardini, Frioni, Soccolini, Zadra, Farinelli, Poni and Palliotti2015). Plants grafted onto 161-49C exhibit higher ABA concentrations in leaves under water stress compared to those grafted onto 1103P, which presented lower amounts, but was accompanied by high levels of the ethylene precursor, ACC (Fahad et al., Reference Fahad, Hussain, Bano, Saud, Hassan, Shan, Khan, Khan, Chen, Wu, Tabassum, Chun, Afzal, Jan, Jan and Huang2015; Labarga et al., Reference Labarga, Mairata, Puelles, Martín, Albacete, García-Escudero and Pou2023; Salvi et al., Reference Salvi, Manna, Kaur, Thakur, Gandass, Bhatt and Muthamilarasan2021). In 1103P a high ACC/ABA ratio has been associated with enhanced drought tolerance (Duan et al., Reference Duan, Li, Guoqiao, Su-Zhou, Li, Merkeryan, Liu and Liu2021; Karaca and Sabir, Reference Karaca and Sabir2018).
Hydraulic properties, regulated by aquaporin activity, are influenced by ABA through both direct and indirect effects. ABA accumulation in leaves under water stress compared to those grafted onto 1103P, which exhibited lower ABA levels but higher ACC concentration (Fahad et al., Reference Fahad, Hussain, Bano, Saud, Hassan, Shan, Khan, Khan, Chen, Wu, Tabassum, Chun, Afzal, Jan, Jan and Huang2015; Galaz et al., Reference Galaz, Pérez-Donoso and Gambardella2024; Shatil et al., Reference Shatil-Cohen, Attia and Moshelion2011). ABA also regulates several metabolic pathways, such as stimulating the phenylpropanoid pathway in red grape cultivars (Deis et al., Reference Deis, Cavagnaro, Bottini, Wuilloud and Silva2011; Martínez-Lüscher et al., Reference Martínez-Lüscher, Cabodevilla, Abdullah, Iglesias, Zamarreño, García-Mina, Morales, Goicoechea and Pascual2025).
Rootstocks differ in their regulation of stomatal conductance and leaf water potential. For instance, 110R regulates stomatal conductance to maintain a relatively constant leaf water potential, prevents dehydration by controlling ABA synthesis, and exhibits isohydric behaviour (Clingeleffer et al., Reference Clingeleffer, Morales, Davis and Smith2019; Perez-Álvarez et al., Reference Pérez-Álvarez, Intrigliolo, Martínez-Moreno, García-Sánchez, Parra, Alfosea-Simon and Buesa2023; Serra et al., Reference Serra, Strever, Myburgh and Deloire2013). In contrast, SO4 tends to exhibit anisohydric behaviour under specific conditions, similar to 1103. The 1103P rootstock stands out for maintaining its drought tolerance through the overexpression of aquaporins (VvPIP and VvTIP) and changes in phytohormone concentrations (lower ABA under stress), while the opposite occurs at 161-49C (Labarga et al., Reference Labarga, Mairata, Puelles, Martín, Albacete, García-Escudero and Pou2023).
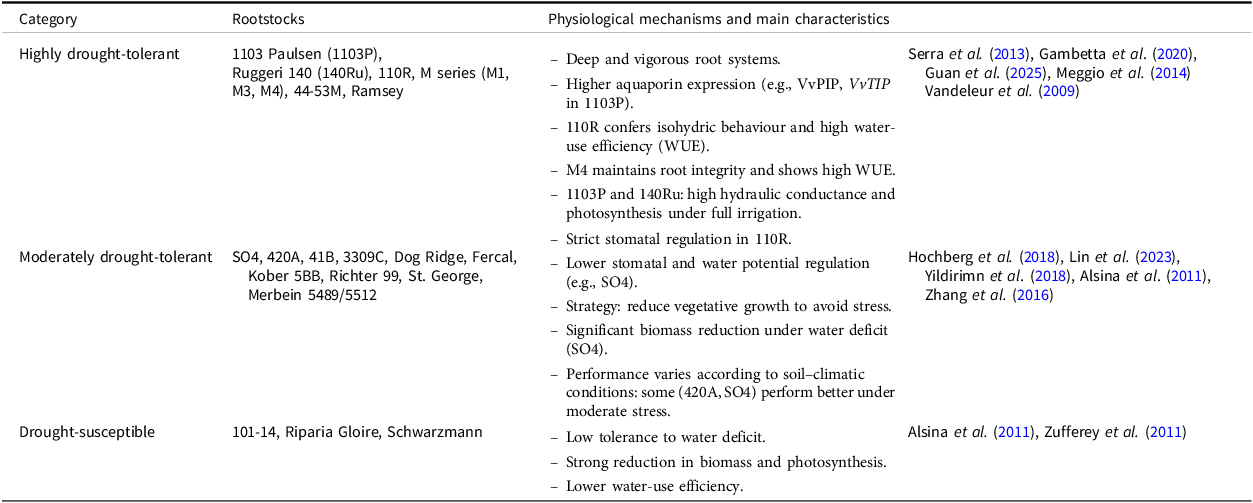
Table 2 shows physiological mechanisms and characteristics that confer varying degrees of drought tolerance. In addition to hydraulic and hormonal regulation, drought responses involve the activation of transcription factors such as the MYB family, WRKY, NAC, and bHLH, which are involved in the activation of stress response pathways. For example, VhMYB2 positively regulates salinity and drought tolerance mechanisms, though VhWRKY44 enhances drought tolerance. Epigenetic regulation also modulates gene expression under stress, as shown by altered methylation of the VvNCED1 promoter, which affects its expression and contributes to stress adaptation. The expression of VvNAC17 is modulated by changes in DNA methylation under water deficit and is associated with various functions such as osmotic regulation, activation of the antioxidant system, and increased flavonoid metabolism (Ju et al., Reference Ju, Yue, Min, Wang, Fang and Zhang2020).
Shows the most recently studied rootstocks and their physiological mechanisms and characteristics that confer varying degrees of drought tolerance

Heat tolerance
Extreme or prolonged heat affects net assimilation, transpiration, and respiration, impacting plant growth and yield in ways that may be irreversible (Krishankumar et al., Reference Krishankumar, Hunter, Alyafei, Hamed, Subramaniam, Ramlal, Kurup and Amiri2025). High temperatures decrease net assimilation, and depending on the temperature, respiration can increase up to 38–42°C and then begin to decline (Sharma et al., Reference Sharma, Kumar, Shahzad, Ramakrishnan, Sidhu, Bali, Handa, Kapoor, Yadav, Khanna, Bakshi, Rehman, Kohli, Khan, Parihar, Yuan, Thukral, Bhardwaj and Zheng2020). At temperatures above 45°C, thylakoid membrane fluidity increases, and total chlorophyll content decreases, reflecting the damage caused by high temperatures (Guo et al., Reference Guo, Li, García-Caparros, Wang and Liang2025). In warm-climate agricultural regions, heat stress is often associated with water stress. The reverse is also very common in these regions where drought is a frequent situation (Hirayama et al., Reference Hirayama and Shinozaki2010). These high temperatures produce cellular damage through the oxidation of proteins, lipids, and nucleic acids, affecting cellular function. These oxidations are due to the reactive oxygen species produced as a result of high temperatures. Naturally, there are defence mechanisms such as the activation of heat shock transcription factors that produce protective proteins or chaperones (Chen et al., Reference Chen, Fei, Pang, Krstic, Clingeleffer, Howell, Chen and Zhang2024). These proteins prevent protein denaturation, thus allowing the plant to maintain its essential metabolism (Priya et al., Reference Priya, Dhanker, Siddique, HanumanthaRao, Nair, Pandey, Singh, Varshney, Prasad and Nayyar2019). However, the molecular capacity to tolerate high temperatures depends on multiple factors, such as genetics, temperature levels and periods, nutritional and water status, etc. Heat-tolerant Vitis varieties such as Vitis davidii cv. Tangwei showed higher expression of a transcription factor (HSFB1) than Vitis vinifera (Chen et al., Reference Chen, Liu, Li, Yuan, Mu, Wang, Li, Duan, Fan, Liang and Wang2023). As a result of high temperatures, plants respond on various physiological levels. One modification detected is the change in phenology. Increased temperatures have led to earlier budburst, up to 16 days, and ripening, up to 24 days (Domínguez et al., Reference Domínguez, Cirrincione, Deis and Martínez2024; Martínez-Lüscher et al., Reference Martínez-Lüscher and Sand Kurtural2021; Venios et al., Reference Venios, Korkas, Nisiotou and Banilas2020). This situation produced the ripening now coincides with the hottest period of the season, decoupling the synthesis of sugars and phenolic compounds and reducing phenolic concentration, which affects wine quality (Deis et al., Reference Deis, de Rosas and Cavagnaro2012; Domínguez et al., Reference Domínguez, Cirrincione, Deis and Martínez2024; Fraga et al., Reference Fraga, Malheiro, Moutinho-Pereira and Santos2013; Sadras et al., Reference Sadras and Moran2012). Photosynthesis is another physiological process that is very sensitive to high temperatures. Under high-temperature conditions, net assimilation and efficiency are reduced. This is due to decreased enzymatic activity; damage to photosystem II, protein denaturation, rubisco inactivation, and chlorophyll degradation occur (Edwards et al., Reference Edwards, Unwin, Kilmister and Treeby2017; Greer, Reference Greer2017; Greer, Reference Greer2018). Through transpiration or evaporative cooling, leaf temperature can be kept below maximum limits, favouring carbon assimilation. This leads to increased water loss. Consequently, rootstocks with higher hydraulic conductivity are better able to sustain transpiration, resulting in lower leaf temperatures and mitigating the adverse effects of heat stress. On the other hand, plants can increase or decrease stomatal density in response to temperature and CO2, affecting water-use efficiency (Bertolino et al., Reference Bertolino, Caine and Gray2019). Another cellular strategy is the accumulation of osmocompatible metabolites such as sucrose, fructose, sorbitol, proline, and organic acids to decrease or maintain cellular osmotic potential and, consequently, water potential, absorb water, and maintain cellular functions (Gouot et al., Reference Gouot, Smith, Holzapfel and Barril2019; Raza et al., Reference Raza, Bhardwaj, Atikur Rahman, García-Caparrós, Habib, Saeed, Charagh, Noriega and Pérez2019). Some of these signals are produced by ABA, whose concentrations increase rapidly in response to heat and water stress, which enhances antioxidant enzymes and regulates carbohydrate content and energy balance. Ethylene plays a modulatory role in heat stress tolerance by enhancing the expression of VpERF1 and VpERF2. These genes encode transcription factors belonging to the ethylene response factor (ERF) family, which is involved in plant responses to biotic and abiotic stresses (Li et al., Reference Li, Chen and Grierson2021). VpERF1 appears to be more involved in the response to abiotic stress than other transcription factors, and it binds to DRE (Dehydration-Responsive Element) in genes involved in dehydration tolerance (Sun et al., Reference Sun, Zhao, Gan, Ren, Fang, Karungo, Wang, Chen, Li and Xin2016). The jasmonates, induced by high solar radiation and high temperatures and changes in the lipid composition of the plasma membrane, initiate signalling by inducing the expression of heat tolerance genes. Meanwhile, melatonin acts as a broad-spectrum antioxidant (Li et al., Reference Li, Lv, Wu, Xie, Zhang, Zhang and Xu2023), improving thermotolerance by increasing the expression of HSPs (heat shock proteins) and the accumulation of phenolic compounds. These proteins act as molecular chaperones, collaborating in the correct folding of proteins, preventing the aggregation of denatured proteins under heat or drought conditions, maintaining protein and membrane stability, and hormonal signalling (ABA and ethylene oxide), thus generating tolerance to heat stress (Berka et al., Reference Berka, Kopecká, Berková, Brzobohatý and Černý2022; Corso and Bonghi, Reference Corso and Bonghi2014). Salicylic acid provides thermotolerance by reducing electrolyte escape, stabilising Ca2+, maintaining H-ATPase activity, Rubisco activity, and PSII functionality (Chai et al., Reference Chai, Liu, Xiang, Meng, Sun, Cheng, Liu, Duan, Xin and Li2019; Gutierrez-Gamboa et al., Reference Gutiérrez-Gamboa, Zheng and Martínez de Toda2019; Noronha et al., Reference Noronha, Silva, Silva, Frusciante, Diretto and Gerós2022; Raza et al., Reference Raza, Bhardwaj, Atikur Rahman, García-Caparrós, Habib, Saeed, Charagh, Noriega and Pérez2019). Rootstocks permit vines to coordinate their response and recovery, maintaining shoot growth by modulating water uptake and signalling various responses to the leaves. The M series rootstocks (M1, M3, and M4) exhibit high tolerance to drought and heat. In particular, M3 is described as heat- and drought-tolerant, and M4 is drought-tolerant.
The M4 rootstock, by improving drought tolerance, confers a greater capacity to maintain water balance and stomatal conductance, allowing for improved photosynthetic performance even at high temperatures (Meggio et al., Reference Meggio, Prinsi, Negri, Lorenzo, Lucchini, Pitacco, Failla, Scienza, Cocucci and Espen2014; Prinsi et al., Reference Prinsi, Negri, Failla, Scienza and Espen2018). The RG series rootstocks were developed with the aim of creating rootstocks adapted to Mediterranean conditions characterised by warm temperatures and prolonged drought events (Marin et al., Reference Marín, Santesteban, Dayer, Villa-Llop, Abad, Gambetta, Torres-Ruiz and Torres2022). RG8 maintained acidity during ripening in warm areas, while RG10 delayed phenolic ripening, making it an excellent choice under extremely hot conditions (Marin et al., Reference Marín, Miranda, Abad, Urrestarazu, Mayor, Villa-Llop and Santesteban2023). The C series was developed to improve tolerance to heat and water stress. The C20 and C114 rootstocks performed well in hot and dry climates (Chen et al., Reference Chen, Fei, Pang, Krstic, Clingeleffer, Howell, Chen and Zhang2024).
Conclusion
The use of rootstocks represents a fundamental strategy for addressing the challenges posed by climate change in viticulture. This review highlights that rootstocks differ markedly in their capacity to tolerate drought and heat through distinct morphological, physiological, and molecular mechanisms (Dal Santo et al., Reference Dal Santo, Zenoni, Sandri, De Lorenzis, Magris, De Paoli, Di Gaspero, Del Fabbro, Morgante, Brancadoro, Grossi, Fasoli, Zuccolotto, Tornielli and Pezzotti2018). 1103 P rootstock exhibits enhanced drought tolerance linked to high hydraulic conductivity, anisohydric behaviour under stress, and the overexpression of aquaporins, whereas 110R shows a more isohydric strategy associated with tighter stomatal regulation and ABA-mediated control for water status.
Future research should therefore prioritise an integrative approach that simultaneously assesses root system traits, hydraulic conductivity, vessel anatomy, aquaporin expression, hormonal signalling, and gas exchange responses in defined scion rootstock combinations. Studies conducted under both well-watered conditions and controlled drought and high-temperature scenarios (27–30°C) will be critical for clarifying the mechanism that confers resilience. Based on this knowledge, genotypes can be selected for new crosses and the development of rootstocks with greater tolerance to drought and extreme heat (Jiao et al., Reference Jiao, Zeng, Huang, Zhang, Mao and Chen2023), while maintaining yield and grape quality. Different rootstocks can improve one or more responses to these stresses, marking important differences between rootstocks. In this context, the diversity of available options – from widely used rootstocks such as 110R, 1103P, and 140Ru to the newer M and RG series – offers a valuable opportunity to enhance vineyard resilience. Strategic rootstock selection, supported by mechanistic understanding, emerges as a key tool for sustainable viticulture under increasingly water and heat-limited conditions.


 Open access
Open access