Introduction
Selective logging is a dominant activity in tropical forest areas, with half of global tropical forests either already explored or earmarked for future exploitation (Poker and MacDicken Reference Poker, Macdicken, Pancel and Köhl2016). Given the scope of this activity and the global importance of tropical forests for biodiversity, it is important to understand its multifaceted effects.
Numerous studies investigated the effects of logging on wildlife populations (Bicknell et al. Reference Bicknell, Struebig, Edwards and Davies2014; Burivalova et al. Reference Burivalova, Şekercioǧlu and Koh2014; Meijaard et al. Reference Meijaard, Sheil, Nasi, Augeri, Rosenbaum, Iskandar, Setyawati, Lammertink, Rachmatika, Wong, Soehartono, Stanley, O’brien, Lammertink, Rachmatika, Wong, Lammertink, Rachmatika, Wong, Soehartono, Stanley and O’brien2005), but few focused on the mechanisms by which logging affects them (Bousfield et al. Reference Bousfield, Cerullo, Massam and Edwards2020; Meijaard et al. Reference Meijaard, Sheil, Nasi, Augeri, Rosenbaum, Iskandar, Setyawati, Lammertink, Rachmatika, Wong, Soehartono, Stanley, O’brien, Lammertink, Rachmatika, Wong, Lammertink, Rachmatika, Wong, Soehartono, Stanley and O’brien2005). Population-level responses are the outcome of demographic processes, which may be indirectly affected by logging (Cosset et al. Reference Cosset, Gilroy and Edwards2019). For example, logging can affect animal feeding (Prosser et al. Reference Prosser, Gardner, Smith, Wern, Ambu and Goossens2016), movement (Cosset et al. Reference Cosset, Gilroy, Tomassi, Benedick, Nelson, Cannon, Messina, Kaputa, Fandrem, Madrid, Lello-Smith, Pavan, King, Fogliano, Hackney, Gerald, Chai, Cros, Chong, Tan, Chai, Cheoh and Edwards2021), and reproduction (Rivera et al. Reference Rivera, Politi, Bucher and Pidgeon2022), all of which can affect demography.
Body condition refers to the amount of energy reserves stored in fat and muscle tissues (Labocha et al. Reference Labocha, Schutz and Hayes2014; Stevenson and Woods Reference Stevenson and Woods2006), and is related to reproductive success (Atkinson and Ramsay Reference Atkinson and Ramsay1995; Ross et al. Reference Ross, Newman, Buesching, Connolly, Nakagawa and Macdonald2021), survival (Merila and Svensson Reference Merila and Svensson1997; Ross et al. Reference Ross, Newman, Buesching, Connolly, Nakagawa and Macdonald2021) and social status (Cotton et al. Reference Cotton, Small and Pomiankowski2006). Changes in body condition may arise due to disturbances such as selective logging and may provide early warnings of population responses (Irwin et al. Reference Irwin, Samonds, Raharison, Junge, Mahefarisoa, Rasambainarivo, Godfrey and Glander2019; Janin et al. Reference Janin, Léna and Joly2011).
The Lowland Tapir (Tapirus terrestris) is the largest terrestrial mammal in South America, where it plays key ecosystem roles (O’Farrill et al. Reference O’farrill, Galetti and Campos-Arceiz2013, Paolucci et al. Reference Paolucci, Pereira, Rattis, Silvério, Marques, Macedo and Brando2019). Tapirs are ecologically flexible (García et al. Reference García, Medici, Naranjo, Novarino and Leonardo2012, Vélez et al. Reference Vélez, Espelta, Rivera and Armenteras2017) and resilient to moderate disturbance. Although some studies reported tapir responses to logging (Carvalho-Jr. et al. Reference Carvalho-Jr, Nienow, Bonavigo and Haugaasen2021; Fragoso et al. Reference Fragoso, Mares, Schmidly, Mares and Schmidly1991; Harris et al. Reference Harris, Hallett, Davis, Carter, Singh, Roopsind, Maharaj and Bicknell2023; Tobler et al. Reference Tobler, Anleu, Carrillo-Percastegui, Santizo, Polisar, Hartley and Goldstein2018), no study to date has investigated potential sub-lethal effects of the activity on the species, such as changes in body condition.
Recently, Clauss et al. (Reference Clauss, Wilkins, Hartley and Hatt2009) and Pérez-Flores et al. (Reference Pérez-Flores, Calmé and Reyna-Hurtado2016) developed a protocol for scoring the body condition of tapirs, based on visual assessment of individuals. This presents an opportunity to non-invasively assess the body condition of wild animals (Pérez -Flores et al. Reference Pérez-Flores, Weissenberger, López-Cen and Calmé2020). Here, we apply this approach to camera-trap data to assess whether selective logging affects tapir body condition scores (hereafter, BCS) in Amazonian concessions. We tested three competing hypotheses regarding the impact of logging on tapir BCS. First, we considered a null hypothesis (H0) where tapir BCS remains unchanged, suggesting species resilience or that logging does not affect its food resources. Alternatively, we hypothesise that logging could lead to an increase in BCS (H1), as disturbances stimulate the growth of understorey vegetation, potentially increasing food availability. Conversely, logging could lead to a decrease in BCS (H2) if it depletes key resources such as large fruiting trees or increases the energetic costs of movement and predator avoidance.
Methods
Jamari National Forest (220,000 ha, 9.2154°S, 62.9395°W) is located in south-western Brazilian Amazonia. Climate is tropical monsoonal and vegetation cover is mostly lowland dense evergreen forest. Approximately half of Jamari has been allocated to reduced-impact logging concessions that are harvested under intensities of 10 and 15 m3 ha⁻1, in 20–25 year cycles. Further details about the area, including maps with the location of the concessions and camera stations, can be found in Carvalho Jr et al. (Reference Carvalho-Jr, Nienow, Bonavigo and Haugaasen2021, Reference Carvalho-Jr, Hawes and Haugaasen2022).
Camera-trap surveys were conducted between 2017 and 2020 as part of the Brazilian National Biodiversity Monitoring Program (Cronemberger et al. Reference Cronemberger, Ribeiro, Acosta, Andrade, Marini-Filho, Masuda, Mendes, Santos Nienow, Polaz, Reis, Sampaio, Souza and Tófoli2023). Sampling followed the TEAM camera trap protocol (Rovero and Ahumada Reference Rovero and Ahumada2017). A total of 190 camera-trap stations were distributed in four arrays (two in unlogged areas and two in logged areas) with a density of one station per 2 km2. At each site, a single camera (Bushnell Trophy Cam) was deployed for ∼30 days in the dry season (June–December). Cameras were placed at knee height and perpendicular to the ground. Each year, between 75 and 106 stations were sampled, totalling 379 deployments, 252 of them in unlogged and 127 in logged sites. The minimum distance between logged and unlogged sites at any given year was 8 km, which is more than twice the diameter of the tapir’s home range typically reported in the literature (2.7–3.25 km, Medici et al. Reference Medici, Mezzini, Fleming, Calabrese and Noonan2022; Noss et al. Reference Noss, Cuéllar, Barrientos, Maffei, Cuéllar, Arispe, Rúmiz and Rivero2003). The average time since logging at deployment sites was 3.16 years (range: 1–9).
Camera trap images were processed on the Wildlife Insights platform (Ahumada et al. Reference Ahumada, Fegraus, Birch, Flores, Kays, O’brien, Palmer, Schuttler, Zhao, Jetz, Kinnaird, Kulkarni, Lyet, Thau, Duong, Oliver and Dancer2020), and then, tapir photos were evaluated for body condition following the protocol proposed by Pérez-Flores et al. (Reference Pérez-Flores, Calmé and Reyna-Hurtado2016). Based on visual assessment of fat and muscles associated with the skeleton, we assigned a one to five score to each of six anatomical regions (head, neck, shoulders, ribs, spine, and pelvic bones). Scores were assigned to samples, defined as sets of photos taken <90 s apart. To ensure independence, we selected one sample per sampling station per year (the sample with the most visible anatomical regions). We use the average score of the visible anatomical regions as a BCS for each sample. The final averaged BCS therefore, ranged from one to five. All scores were assigned by the same person (GGCS).
We used the Pearson correlation coefficient to assess the consistency between scores of different anatomical regions. We used a Bayesian generalised linear mixed model (GLMM) to assess the effect of the binary variable treatment (unlogged/logged) on tapir BCS. To account for dependencies in the data structure, we included site and year as random effects, and we included a Gaussian process in the model to account for the possibility of a single individual being recorded at multiple neighbouring cameras. This explicitly models the spatial dependency between nearby locations, ensuring that observations from cameras within a single individual’s home range are not treated as entirely independent spatial units. The model was fitted using the brms R package (Bürkner Reference Bürkner2017).
Results
We obtained 68 independent samples from 56 sites. The average number of photos per sample was 9.5, ranging from three to 37 (disregarding an outlier with 80 photos). The most frequent number of visible anatomical regions was six (n = 30), followed by three (n = 11), four (n = 10), five and one (n = 6 for both), and two (n = 5). The most visible region was the ribs (n = 58, 85%), followed by neck (n = 56, 82%), head (n = 54, 80%), shoulder (n = 53, 78%), pelvis and spine (n = 39, 57% for both). The scores of all regions were highly correlated (all r > 0.67, mean r = 0.75).
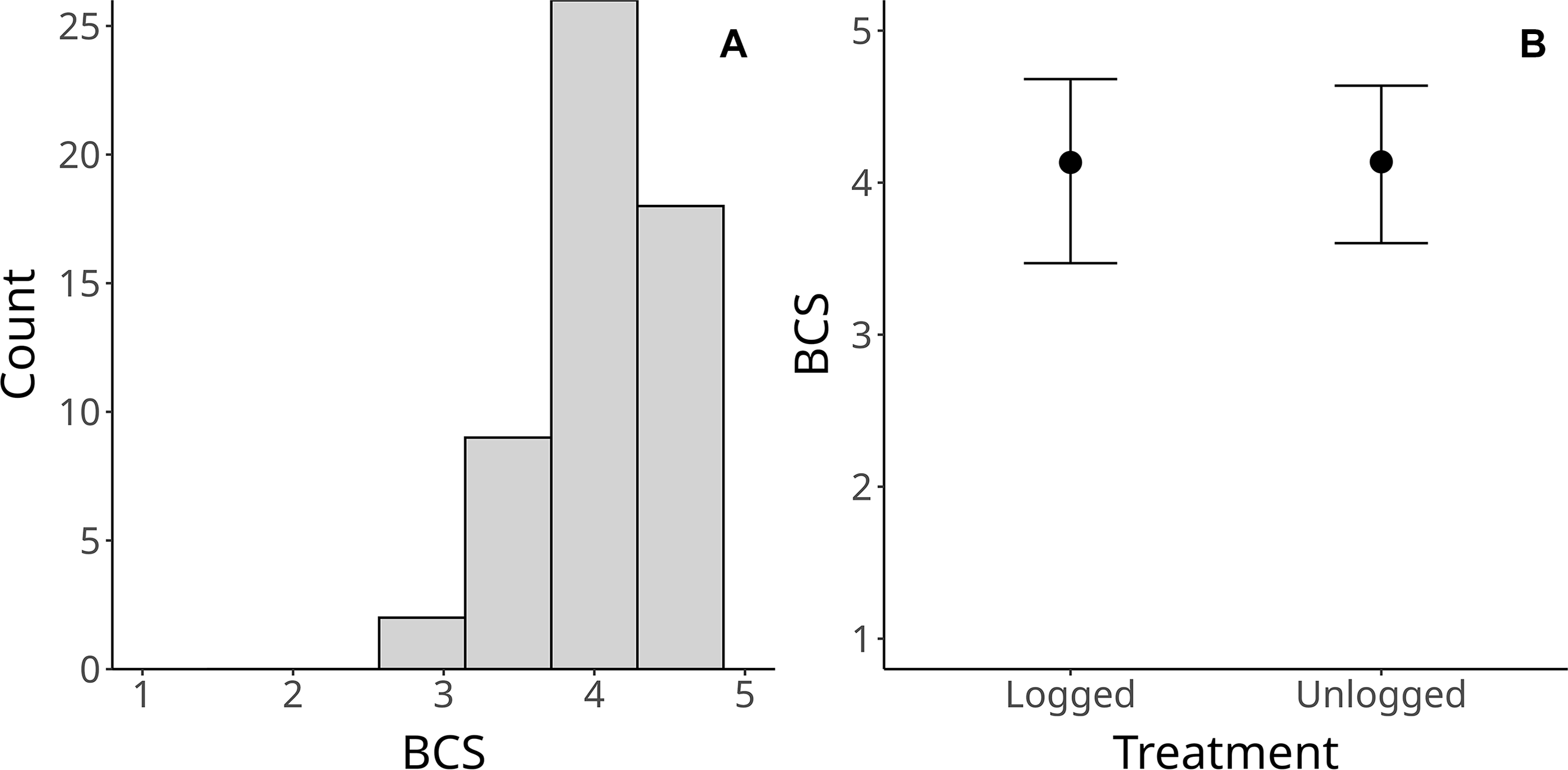
The average body condition across all samples was 4.2, ranging from three to five (Figure 1A). Forty-three BCS samples were from unlogged sites, and 25 from logged sites. According to the GLMM model, there was no significant difference between the scores from unlogged and logged sites (Figure 1B, mean BCS unlogged = 4.2, sd = 0.54; mean BCS logged = 4.3, sd = 0.56). As for the Gaussian process, the model estimated a spatial length-scale (ℓ) of 0.15 km, with a 95% Credible Interval of 0.04–0.43 km, which suggests that camera spacing (≥ 1.4 km) was sufficient to ensure independence.
(A) Histogram of tapir body condition scores (BCS) across all independent samples from Jamari National Forest, Brazilian Amazonia; (B) Conditional effects of treatment (unlogged/logged) on tapir BCS.

Figure 1. Long description
A histogram of tapir body condition scores across all independent samples from Jamari National Forest, Brazilian Amazonia. The histogram has five vertical bars representing body condition scores ranging from 1 to 5. The x-axis is labeled ‘BCS’ and the y-axis is labeled ‘Count’. The highest bar, representing a body condition score of 5, reaches a count of 25. The second highest bar, representing a body condition score of 4, reaches a count of approximately 18. The bars for body condition scores of 3, 2, and 1 have progressively lower counts. The distribution is skewed towards higher body condition scores. Additionally, a plot shows the conditional effects of treatment (unlogged/logged) on tapir body condition scores. The x-axis is labeled ‘Treatment’ with categories ‘Logged’ and ‘Unlogged’, and the y-axis is labeled ‘BCS’. Each treatment category has a data point with an error bar. The body condition scores for both logged and unlogged treatments are around 4, with overlapping error bars indicating similar body condition scores between the two treatments.
Discussion
We investigate the effect of selective logging on tapir body condition in Amazonian forest concessions. Our study joins others that used visual assessments to measure the BCS of tapirs (Pérez-Flores et al. Reference Pérez-Flores, Calmé and Reyna-Hurtado2016, Reference Pérez-Flores, Weissenberger, López-Cen and Calmé2020) and other wild species (Prosser et al. Reference Prosser, Gardner, Smith, Wern, Ambu and Goossens2016; Ramesh et al. Reference Ramesh, Kalle, Sankar and Qureshi2011; Smiley et al. Reference Smiley, Rittenhouse, Mong and Monteith2020; Wemmer et al. Reference Wemmer, Krishnamurthy, Shrestha, Hayek, Thant and Nanjappa2006; Zielke et al. Reference Zielke, Wrage-Mönnig and Müller2018).
Most tapirs were in good condition. This agrees with the few available reports of wild tapir body condition: >75% of tapirs with good condition in the Atlantic Forest and Pantanal (Medici et al. Reference Medici, Mangini and Fernandes-Santos2014), >70% with good condition in the Brazilian Cerrado (Fernandes-Santos et al. Reference Fernandes-Santos, Medici, Testa-José and Micheletti2020) and >90% of T. bairdii with above-average condition in tropical forests of Mexico (Pérez-Flores et al. Reference Pérez-Flores, Calmé and Reyna-Hurtado2016, but see Flores et al. Reference Pérez-Flores, Weissenberger, López-Cen and Calmé2020).
We did not find significant differences in BCS between unlogged and logged sites. This supports hypothesis H1, that reduced-impact logging does not affect food availability for the species, at least in the short term. This is consistent with the flexibility of tapirs in their diet (Henry et al. Reference Henry, Feer and Sabatier2000; Tobler et al. Reference Tobler, Janovec and Cornejo2010, Vélez et al. Reference Vélez, Espelta, Rivera and Armenteras2017), and the fact that they sometimes select secondary forests and open-canopy areas for browsing (Ferreguetti et al. Reference Ferreguetti, Tomas and Bergallo2017; Fragoso et al. Reference Fragoso, Mares, Schmidly, Mares and Schmidly1991; García et al. Reference García, Medici, Naranjo, Novarino and Leonardo2012; Salas Reference Salas1996). In fact, some studies reported positive effects of logging on large herbivores (Davies et al. Reference Davies, Heydon, Leader-Williams, Mackinnon, Newing, Fimbel, Grajal and Robinson2001; Morgan et al. Reference Morgan, Mundry, Sanz, Ayina, Strindberg, Lonsdorf and Kühl2018; Zwerts et al. Reference Zwerts, Sterck, Verweij, Maisels, Waarde, Geelen, Tchoumba, Zebaze and Kuijk2024), mostly because of the flush of palatable regrowth due to the increased amount of light reaching the understorey (Malhi et al. Reference Malhi, Riutta, Wearn, Deere, Mitchell, Bernard, Majalap, Nilus, Davies, Ewers and Struebig2022; Zwerts et al. Reference Zwerts, Sterck, Verweij, Maisels, Waarde, Geelen, Tchoumba, Zebaze and Kuijk2024).
Selective logging is a moderate disturbance compared to other anthropogenic stressors (Bousfield et al. Reference Bousfield, Cerullo, Massam and Edwards2020), so that tapir responses can differ when disturbances are more severe (e.g., Pérez-Flores et al. Reference Pérez-Flores, Weissenberger, López-Cen and Calmé2020). Furthermore, other factors besides food availability can affect tapir health, including pesticides and parasite load (Fernandes-Santos et al. Reference Fernandes-Santos, Medici, Testa-José and Micheletti2020; Mangini et al. Reference Mangini, Medici and Fernandes-Santos2012). Finally, we note that BCS is only one side of tapir responses to logging. A more complete assessment of logging impacts must consider additional aspects such as the distribution and abundance of the species, activity budgets, population dynamics, etc.
A potential caveat of this study is that tapirs were not uniquely identified, introducing a risk of pseudorepetition and, consequently, variance shrinkage. However, we believe this issue was solved, or at least minimised, by our conservative analytical framework, which included temporal filtering (restricting the dataset to a single record per location per year) and spatial modelling (including a Gaussian process to explicitly model the probability of neighbouring cameras recording the same individual).
In summary, we found that most tapirs in the study area had excellent body condition with no significant effects of logging on BCS. However, our results apply to the short term, as we cannot rule out that conditions may deteriorate in the future. Additional studies assessing the body condition of tapirs covering longer post-logging recovery periods or under alternative management scenarios are recommended. Beyond the immediate findings of this study, our results highlight the utility of the visual body condition score as a non-invasive, cost-effective tool for assessing the impacts of forest management in Amazonian forests.
Acknowledgements
We thank Instituto de Pesquisas Ecológicas, Serviço Florestal Brasileiro, Ecoporé, AMATA, MADEFLONA, Paulo H. Bonavigo, Camila M. Lemke and Rodrigo A. Nobre.
Funding statement
G.G.C.S. was supported by a PIBIC ICMBio scholarship (Fundep/UFMG project #30156). Fieldwork was supported by Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio; and Instituto de Pesquisas Ecológicas (project ‘Participatory monitoring of biodiversity in conservation units in the Amazon’, supported by USAID and the Gordon & Betty Moore Foundation).
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical statement
None.