1 Introduction
Fossils are a rich source of information about the history of the Earth and can be used to test hypotheses about mass extinctions, biogeography, the diversification of clades and communities, the effects of climate change on evolution and extinction, and the effects of evolution and extinction on climate. Fossils are also a finite natural resource, an important aspect of a nation’s natural heritage, and a didactic medium that engages public audiences in discourse about the history of life on Earth. The fossil record of the National Park System (NPS) exists precisely at the intersection between fossils as a body of scientific information and fossils as public natural heritage, and the National Park Service balances the multidimensional significance of federal fossil resources through its four core management activities: inventory, monitoring, research, and curation.
Here, we present a comprehensive inventory of Cenozoic paleobotanical resources that details the spatial, stratigraphic, taxonomic, and temporal scope and significance of fossil plants preserved in 74 of the 288 National Park Service areas with paleontological resources. This inventory is the first installment of a complete inventory of Cenozoic, Mesozoic, and Paleozoic paleobotany of the NPS, and the product of a collaborative effort among four student–mentor pairs and NPS scientists sponsored by the Paleontology in the Parks Fellowship Program of the National Park Service and Paleontology Society.
1.1 Cenozoic Paleobotany in North America
The Cenozoic paleobotanical record of North America preserves evidence of the modernization of plant families and terrestrial ecosystems, extinct plant lineages and plant communities without modern ecological analogs, and the response of plants to global events such as the Cretaceous–Paleogene (K–Pg) mass extinction, Paleocene–Eocene Thermal Maximum (PETM), and climatic optima. This history has been written in part based on fossil plants now under the stewardship of the NPS, and provides a rich context for interpreting the significance of new or understudied NPS fossil plant resources. In this section, we summarize key patterns present in the Cenozoic paleobotanical record of the USA in relation to four themes with particular relevance for interpreting NPS resources: recovery from mass extinction, angiosperm diversification, climate change, and intercontinental dispersal. In doing so, we highlight the paleobotanical resources of NPS units that have contributed or have the potential to contribute to a greater understanding of the evolutionary, climatic, and physiographic history of North America.
1.1.1 Extinction Recovery
The collision of a bolide asteroid with the Yucatan Peninsula about 66 Ma prompted a restructuring of terrestrial ecosystems through instantaneous and widespread extinction of species followed by heterogeneous pathways of recovery (Alvarez et al., Reference Alvarez, Alvarez, Asaro and Michel1980; McElwain and Punyasena, Reference McElwain and Punyasena2007; Nichols and Johnson, Reference Nichols and Johnson2008; Wilf et al., Reference Wilf, Carvalho and Stiles2023). Mass extinction among plants at the K–Pg boundary has been documented from latest Cretaceous and earliest Paleocene outcrops in the Williston Basin in North Dakota (Wilf and Johnson, Reference Wilf and Johnson2004), the Denver Basin in Colorado (Barclay et al., Reference Barclay, Johnson, Betterton and Dilcher2003; Lyson et al., Reference Lyson, Miller and Bercovici2019), and the Raton and San Juan basins in southern Colorado and northern New Mexico (Tschudy et al., Reference Tschudy, Pillmore, Orth, Gilmore and Knight1984; Wolfe and Upchurch, Reference Wolfe and Upchurch1986, Reference Wolfe and Upchurch1987; Flynn and Peppe, Reference Flynn and Peppe2019). Palynological sequences across the K–Pg boundary indicate a spike in fern spore abundance and paucity of angiosperm pollen immediately above the iridium anomaly that marks the boundary and extinction of 20–30% of pollen taxa (Tschudy et al., Reference Tschudy, Pillmore, Orth, Gilmore and Knight1984; Johnson et al., Reference Johnson, Nichols, Attrep and Orth1989; Barclay et al., Reference Barclay, Johnson, Betterton and Dilcher2003). Macrofloral assemblages offer greater taxonomic resolution than palynofloras, and comparison of latest Cretaceous and earliest Paleocene leaf assemblages indicates even higher plant extinction rates, around 50% (Wolfe and Upchurch, Reference Wolfe and Upchurch1986; Wilf and Johnson, Reference Wilf and Johnson2004; Lyson et al., Reference Lyson, Miller and Bercovici2019).
The North American forests that emerged in the first ca. 5 Ma ensuing the K–Pg mass extinction differed markedly in physiognomic and floristic attributes from the latest Cretaceous forests that they replaced, and showed geographic heterogeneity among themselves. Macrofossil evidence from the Raton and Williston basins suggests an increased importance of deciduous trees in Early Paleocene mid-latitude floras based on community-level shifts in leaf functional traits (Wolfe and Upchurch, Reference Wolfe and Upchurch1987; Blonder et al., Reference Blonder, Royer, Johnson, Miller and Enquist2014), and this inference meshes well with the prominence of families such as Betulaceae, Cornaceae, Juglandaceae, and Platanaceae, which are common in the temperate deciduous forests of the Northern Hemisphere today (Manchester, Reference Manchester2014). These lineages, alongside taxodiaceous conifers, Cercidiphyllaceae, and Hammamelidaceae, were the dominant components of backswamp and alluvial ridge habitats during the first several million years following the bolide impact (Wing, Reference Wing, Janis, Scott and Jacobs1998), and have been interpreted to comprise a low-diversity and spatially homogenous flora spanning from southern Alberta to central Colorado (Johnson and Hickey, Reference Johnson and Hickey1990; McIver and Basinger, Reference McIver and Basinger1993; Wing, Reference Wing, Janis, Scott and Jacobs1998; Barclay et al., Reference Barclay, Johnson, Betterton and Dilcher2003; Wilf and Johnson, Reference Wilf and Johnson2004; Manchester, Reference Manchester2014). However, recent reports from earliest Paleocene strata in the San Juan Basin and the Corral Bluffs site in the Denver Basin have documented higher diversity floras within the first million years of the Paleocene (Flynn and Peppe, Reference Flynn and Peppe2019; Lyson et al., Reference Lyson, Miller and Bercovici2019). These floras comprise a distinct set of taxa, including the earliest legume fossil, and have physiognomic attributes comparable to modern seasonal tropical forests.
Yukon–Charley Rivers National Preserve and Denali National Park and Preserve in central Alaska, and Big Bend National Park in southern Texas each include fossiliferous sequences of Upper Cretaceous and Lower Paleocene rocks (see Section 3 for further details). Increased monitoring, reconnaissance, and research in these units has the potential to further increase our understanding of the geographic heterogeneity in species’ responses to the extinction event and subsequent ecological recovery.
1.1.2 Angiosperm Diversification
The Cenozoic is a time interval characterized by diversification within extant families of flowering plants (Ramírez-Barahona et al., Reference Ramírez-Barahona, Sauquet and Magallón2020; Zuntini et al., Reference Zuntini, Carruthers and Maurin2024) and their rise to ecological dominance (Wing and Boucher, Reference Wing and Boucher1998; Carvalho et al., Reference Carvalho, Jarmillo and de la Parra2021). The Cenozoic strata of the USA contribute to our understanding of this diversification through documenting extinct genera with known (e.g., Florissantia; Manchester, Reference Manchester1992) or unknown (e.g., Palibinia; Manchester et al., Reference Manchester, Judd and Kodrul2024) familial affinities, and early occurrences of extant genera that persist (e.g., Alnus; Forest et al., Reference Forest, Savolainen and Chase2005) or have gone extinct (e.g., Pterocarya; Stults et al., Reference Stults, Tiffney and Axsmith2022) from the continent. Collectively, these occurrences illustrate that North American vegetation has been deeply affected by extinction, and that the extant flora is composed of families and genera with heterogeneous ages of origin and evolutionary histories.
Fossil plants provide insight into extinction by preserving combinations of morphological characters that cannot be observed today and by occurring beyond the geographic range of their nearest living relatives. Extinct genera with morphologies consistent with living angiosperm families but not matching any extant genus within them formed a prominent component of Paleogene North American floras and cooccurred with or in some cases predated the first occurrences of confamilial extant genera. Paleobowditchia, an extinct genus in the Fabaceae subfamily Papilionoideae, whose type locality is in the Lamar River Formation at Yellowstone National Park (Herendeen et al., Reference Herendeen, Cardoso, Herrera and Wing2022), attests to the latter pattern, and Cedrelospermum, an extinct genus of the Ulmaceae that cooccurs with Ulmus in the fossiliferous shales of Florissant Fossil Beds National Monument (Manchester, Reference Manchester1989a), attests to the former. Several extant families appeared and disappeared from the North American flora over the course of the Cenozoic, and others lost phylogenetic diversity (number of higher taxa below family rank) through the extinction of genera that persist outside of North America today. The Cercidiphyllaceae and Trochodendraceae, monogeneric families now restricted to eastern Asia, are represented by extant and extinct genera in the North American fossil record that grew as common components of Paleocene–Miocene floras of the Great Plains, Rocky Mountains, and Pacific Northwest (Crane et al., Reference Crane, Manchester and Dilcher1991; Manchester et al., Reference Manchester, Crane and Dilcher1991), including those preserved in Paleocene strata of Theodore Roosevelt National Park in North Dakota (Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991), and Eocene strata of Glacier National Park in Montana (Smith, Reference Smith2023). The Juglandaceae, represented in North America today by Juglans and Carya, were once represented by three additional extant genera that today occur only in Eurasia (Manchester, Reference Manchester1989b), as well as numerous extinct genera (Manchester, Reference Manchester1989b, Reference Manchester1991). Thus, despite its ecological importance in the extant North American flora, the Juglandaceae represent a prominent example of a lineage that lost phylogenetic diversity in North America over the course of the Cenozoic.
Extinction did not affect all lineages equally over the course of the Cenozoic, and the US fossil record also illustrates the capacity of plant families and genera to persist and diversify over multimillion-year timescales. Acer and Quercus are diverse and ecologically dominant genera today, with evolutionary roots deep in the Cenozoic: Occurrences in the Late Paleocene–Early Eocene Copper Lake Formation in Katmai National Park and Preserve, Alaska (Acer; Wolfe and Tanai, Reference Wolfe and Tanai1987) and the Middle Eocene (44 Ma) Clarno Nut Beds of John Day Fossil Beds National Monument (Quercus; Manchester, Reference Manchester1994) are among the earliest records of each genus. At the family level, fossil histories of the Betulaceae and Rosaceae indicate a net accumulation of higher-level taxa throughout the Cenozoic, a marked contrast to the trajectory of phylogenetic diversity in the Juglandaceae. The Rosaceae today are among the most diverse families in North America (ca. 700 species), and appear to have first established this diversity during the Eocene (DeVore and Pigg, Reference DeVore and Pigg2007). Latest Early Eocene floras from the Okanogan Highlands of eastern Washington and British Columbia include the oldest known Prunus and Oemleria flowers (Benedict et al., Reference Benedict, DeVore and Pigg2011), along with early or first occurrences of other rosaceous taxa (Wolfe and Wehr, Reference Wolfe and Wehr1988), and the Late Eocene flora of Florissant Fossil Beds National Monument includes early occurrences of Rosa, Cercocarpus, and several additional rosaceous taxa (Manchester, Reference Manchester2001). In the Betulaceae, all five extant North American genera have persisted since the Paleogene (Crane, Reference Crane1989; Forest et al., Reference Forest, Savolainen and Chase2005), and some (e.g., Carpinus, Corylus) show a similar pattern of stratigraphic occurrence to the Rosaceae, with first occurrences in the Okanogan Highlands (Pigg et al., Reference Pigg, Manchester and Wehr2003; Pigg and DeVore, Reference Pigg and DeVore2010).
1.1.3 Climate Change
Global climate of the Cenozoic transitioned from a high-CO2 greenhouse world that lacked polar ice sheets in the Paleocene and Eocene to a low-CO2 world with southern ice sheets beginning in the late Eocene and northern ice sheets beginning in the Pliocene (Zachos et al., Reference Zachos, Dickens and Zeebe2008; Westerhold et al., Reference Westerhold, Marwan and Drury2020; Cenozoic CO2 Proxy Integration Project (CenCOPIP) Consortium, 2023). Superimposed on this general trend are rapid and transient warming episodes, such as PETM and other Early Eocene hyperthermal events, prolonged warm periods such as Early and Middle Eocene climatic optima and the Middle Miocene Climatic Optimum, and abrupt shifts in mean climate state such as the Eocene–Oligocene Transition (EOT), Middle Miocene Climatic Transition, and glacial–interglacial cycles of the Quaternary. Fossil floras of appropriate stratigraphic position document two principal patterns in the response of plants to this climatic history: (1) rapid or cyclic climate changes are matched by rapid plant migration, leading to large taxonomic turnover but low rates of extinction; and (2) prolonged or abrupt and directional climate changes are associated with long-term shifts in plant community composition mediated by immigration and/or local extinction and speciation.
The response of North American plant communities to climatic perturbations lasting less than a million years is best known from two time intervals: the PETM, a period of ca. 200,000 years beginning at the Paleocene–Eocene boundary (56 Ma) that was characterized by 4–8°C of greenhouse warming (McInerney and Wing, Reference McInerney and Wing2011); and the last ca. 800,000 years, characterized by large fluctuations in polar ice volume and atmospheric CO2 levels (Petit et al., Reference Petit and Raynaud1999; Lüthi et al., Reference Lüthi, Le Floch and Bereiter2008). Floral records from the Bighorn Basin in Wyoming indicate that the predominant response of woody plants to the climate changes of the PETM was through reversible range shifts; however, local extirpations and immigrations occurred as well, giving forth to novel earliest Eocene plant associations (Wing et al., Reference Wing, Harrington and Smith2005; Wing and Currano, Reference Wing and Currano2013; Korasidis and Wing, Reference Korasidis and Wing2023). A similar response of eastern North American vegetation to cycles of icesheet expansion and retraction during the Late Pleistocene and Holocene has been documented from pollen cores taken from a large network of glacial lakes, including one within Cape Cod National Seashore in Massachusetts (Winkler, Reference Winkler1985). Collectively, these records illustrate that the formation and dissolution of plant associations reflect individualistic interactions between species and climatic factors (Jackson and Overpeck, Reference Jackson and Overpeck2000).
In contrast, plant responses to climatic changes occurring over multimillion-year timescales are characterized by widespread extinction and directional shifts in floristic composition and/or vegetation structure. This pattern is well-documented in the Eocene floras (Leopold and MacGinitie, Reference Leopold, MacGinitie and Graham1972; Wing, Reference Wing1987) of the Rocky Mountain Region that record a progressive loss of Late Paleocene and Early Eocene taxa, many with East Asian biogeographic affinities (e.g., Metasequoia; Wing, Reference Wing1987) and influx of taxa with neotropical biogeographic affinities (e.g., Myrtaceae; Manchester et al., Reference Manchester, Dilcher and Wing1998). This shift in floristic composition has been correlated with regional drying in the Rocky Mountain Region and differs from the Eocene record of the Pacific Northwest, which retained a wetter climate and tropical to subtropical “East Asian” lineages up until the Late Eocene–Early Oligocene (Leopold and MacGinitie, Reference Leopold, MacGinitie and Graham1972; DeVore and Pigg, Reference DeVore and Pigg2010). The Middle Eocene (44 Ma) Clarno and Early Oligocene (33 Ma) Bridge Creek floras from Oregon, preserved in John Day Fossil Beds National Monument, illustrate major changes in the makeup of the Pacific Northwest flora associated with a gradual trend of climatic cooling and drying in the Middle–Late Eocene, punctuated by the abrupt climatic deterioration of the EOT (Retallack, Reference Retallack, Prothero and Berggren1992; Wolfe, Reference Wolfe, Prothero and Berggren1992). This period of climate change has been interpreted as a major factor in the regional replacement of evergreen angiosperms by deciduous angiosperms and conifers, and the regional extinction of numerous taxa that persist today in the eastern USA and eastern Asia (MacGinitie, Reference MacGinitie1953; Meyer and Manchester, Reference Meyer and Manchester1997).
1.1.4 Intercontinental Dispersal
Three land connections influenced the potential for intercontinental dispersal via biogeographic corridors during the Cenozoic in North America: the Bering Land Bridge (BLB) between Alaska and Russia; the North Atlantic Land Bridge (NALB) between eastern North America and Western Europe; and the Isthmus of Panama that connected the Americas. The BLB lay at high northern latitudes, likely between 60 and 80°N, and served as a viable corridor throughout the Cenozoic, despite variability in the degree of continuity and local climate of land across the Bering Strait (Tiffney and Manchester, Reference Manchester2001). Migration across the NALB is hypothesized to have occurred across a northern pathway, Svalbard to Greenland to Ellesmere Island (McKenna, Reference McKenna1975, Reference McKenna, Bott, Saxov, Talwani and Thiede1983), and a southern pathway, Scotland to the Faroe Islands to Iceland to Greenland to North America (Tiffney, Reference Tiffney1985; Tiffney and Manchester, Reference Manchester2001; Denk et al., Reference Denk, Grímsson, Zetter, Símonarson, 96Landman and Harries2011). Current tectonic and paleobotanical evidence (reviewed by Denk et al., Reference Denk, Grímsson, Zetter, Símonarson, 96Landman and Harries2011) suggests that migration via the southern NALB occurred up until the Late Miocene (10–6 Ma). The Isthmus of Panama formed a persistent terrestrial landscape between Central and North America around 3.5 Ma, and was the result of a complex sequence of events, beginning with the formation of a large island in the Late Eocene (Jaramillo, Reference Jaramillo, Hoorn, Perrigo and Antonelli2018). The geologic history of the land bridge and the frequency of biotic interchange prior to its formation is an active area of debate (Montes et al., Reference Montes, Cardona and Jaramillo2015; O’Dea et al., Reference O’Dea, Lessios and Coates2016).
The inference of migration routes for taxa that now or once occurred on multiple Laurasian (North America, Europe, Asia) continents are based on the geo-stratigraphic distribution of fossil taxa and the present distribution of their living relatives (Manchester, Reference Manchester1999). The fossil record of North America includes immigrant taxa with earliest stratigraphic occurrences in Europe or Asia (e.g., Betula), supporting dispersal into North America by the NALB or BLB, as well as migrant taxa with earliest occurrences in North America that subsequently occur in Asia and Europe (e.g., Cornus). Among extant lineages, legacies of intercontinental dispersal have contributed to modern Holarctic distributions (e.g., Acer), and Asian–North American disjunctions (e.g., Nyssa). Differential extinction following range expansion via past dispersal corridors has also produced patterns of paleo-endemism among taxa that were widespread as fossils but only persist on one continent (e.g., Eucommia in Asia, Comptonia in North America), and “range-reversal” for taxa that persist on a different continent from their earliest fossil occurrence (e.g., Cyclocarya).
Early and Middle Eocene floras preserved in several Alaskan national parks provide insight into the vegetation structure near the migration corridor of the BLB and provide the opportunity to catch modern Holarctic taxa in the act of migration. Early Eocene plants from Wrangell–St. Elias National Park and Preserve include lineages that are mainly tropical and evergreen today, such as the Icacinaceae, Lauraceae, and Annonaceae (Wolfe, Reference Wolfe1966, Reference Wolfe1977), implying that the greenhouse climate of the Early Cenozoic allowed some members of these families to reach high latitudes despite the increased seasonality of insolation. Further systematic and paleoecological study of the floras within Alaskan national parks has the potential to improve our understanding of the origin of modern biogeographic patterns among Northern Hemisphere plants and of ecosystem function within non-analog high-latitude evergreen floras.
2 Scope of the Review
Our inventory focuses on occurrences of plant macrofossils (megafossils) in Cenozoic sedimentary rock formations and unlithified sediments that fall within the boundaries of national parks, historical parks, scenic and heritage trails, monuments, preserves, recreation areas, and other types of units administered by the National Park Service. We define plant macrofossils as macroscopic physical evidence of past plant life (Santucci et al., Reference Santucci, Tweet, Visaggi and Hodnett2024) and include permineralizations, charcoalifications, and compressions and impressions of leaves, flowers, fruits, seeds, stems, branches, and wood. In addition, we selectively include records of plant microfossils, such as pollen, spores, phytoliths, and dispersed cuticle from cores and outcrops and plant ichnofossils and related sedimentary structures, such as root traces and other rhizoliths. The microfossil and ichnofossil records are here reported in instances where their presence indicates potential for macrofossil discovery or has contributed to paleoenvironmental interpretation of plant macrofossil assemblages. We exclude macrofossil occurrences documented from packrat middens (reviewed by Tweet et al., Reference Tweet, Santucci and Hunt2012a) and provide less thorough coverage of the substantial body of literature devoted to the Quaternary pollen record.
To compile the NPS Cenozoic plant fossil inventory, we searched peer-reviewed scientific journals, NPS archives (internal park communications, internal reports, etc.), geologic maps, paleontological inventory reports, USGS internal memos (“E&R reports”), popular science articles, museum collection databases, and other relevant databases such as the Alaska Paleontological Database (www.alaskafossil.org/) and the Florissant Fossil Database (https://flfo-search.colorado.edu/). Potential records were checked against NPS boundaries to determine whether a given occurrence was within or outside a given park unit, and for confirmed fossils, we conducted a secondary literature search to the best of our abilities to find associated geochronologic, stratigraphic, taphonomic, and taxonomic information. We present park-wide summaries for the Quaternary and each epoch of the Paleogene and Neogene in Supplementary Tables 1–6 (Supplementary Tables are available to access online at www.cambridge.org/matel-et-al). Text summaries are presented later, organized geographically by the seven NPS legacy regions, and within regions by the 32 Inventory and Monitoring networks.
3 Park Inventories
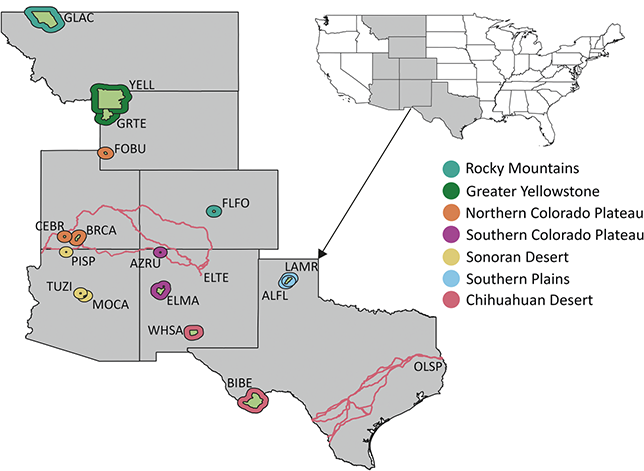
3.1 Alaska Region
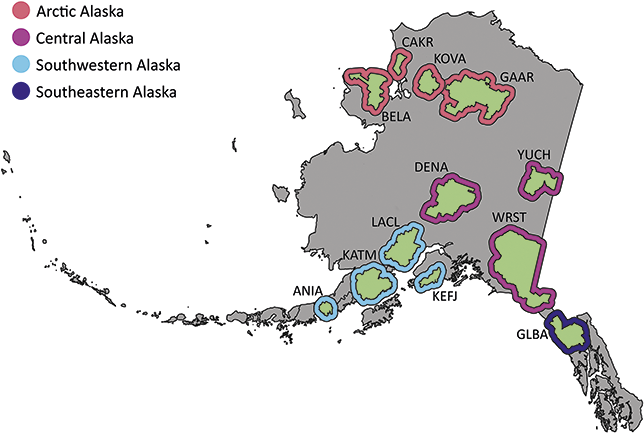
The Alaska Region includes 16 NPS units divided among four inventory and monitoring networks and encompasses 54 million acres, approximately two-thirds of the land area managed by the NPS (Figure 1). Of these 16 parks, 11 contain confirmed records of Cenozoic plant macrofossils, and Yukon–Charley Rivers National Preserve likely contains undocumented Cenozoic plant macrofossils (Kenworthy and Santucci, Reference Kenworthy and Santucci2003; Santucci and Kenworthy, Reference Santucci and Kenworthy2008; Elder et al., Reference Elder, Santucci, Kenworthy, Blodgett and McKenna2009; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011).
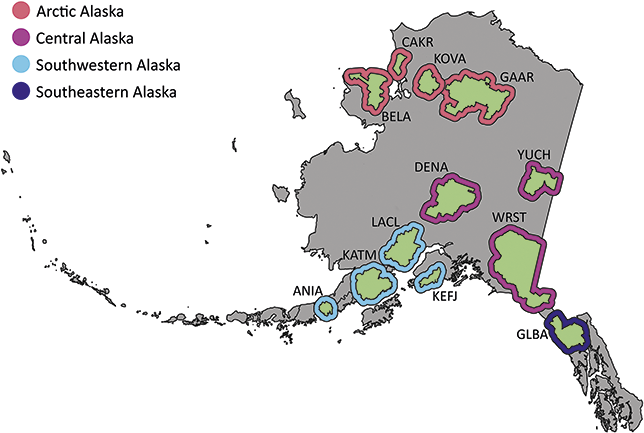
Figure 1 Map of the 11 NPS units with confirmed Cenozoic paleobotanical resources in the Alaska Region and one unit (YUCH) with potential resources. Interior green shading illustrates the area enclosed by the perimeter of park boundaries plus a 3 km buffer, and exterior shading colored by inventory and monitoring represents the area enclosed by the perimeter of park boundaries plus a 30 km buffer. Unit acronyms are generally formed from the first four letters of one-word unit names (e.g., Katmai National Park and Preserve = KATM) and the first two letters of the first two words for multiple-word unit names (e.g., Bering Land Bridge National Preserve = BELA).
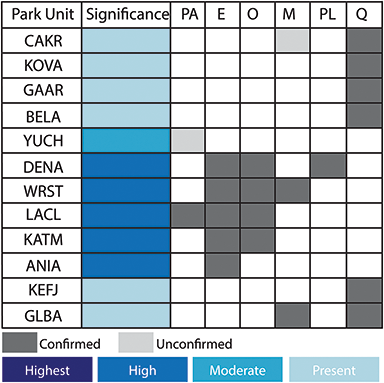
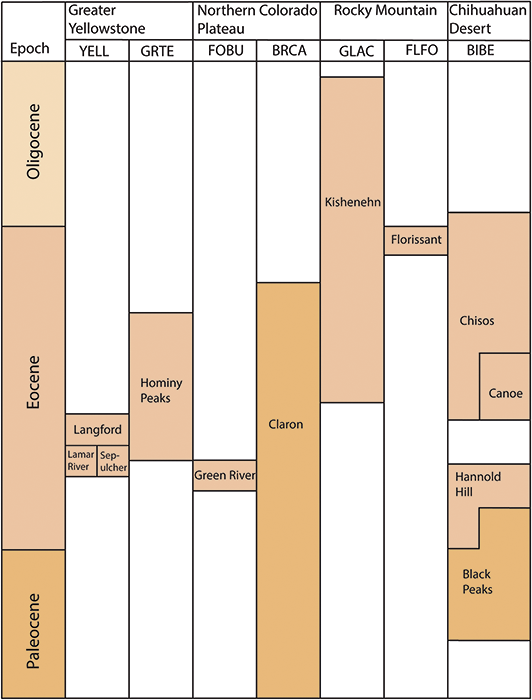
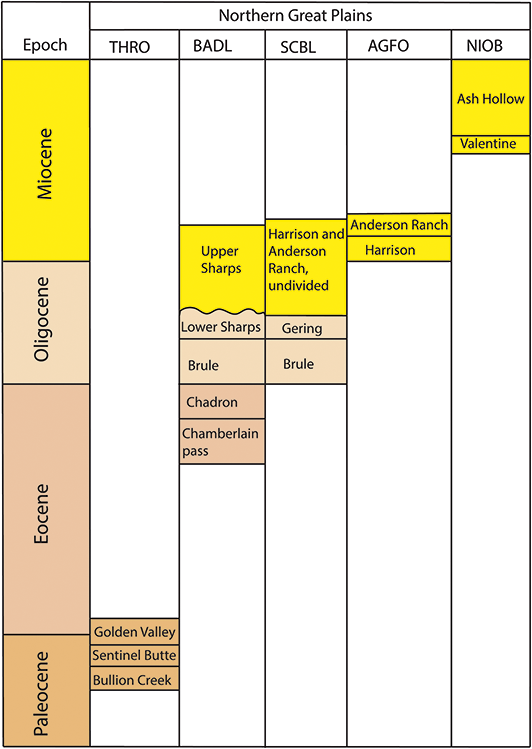
Alaskan national parks preserve highly significant paleobotanical resources that provide insight into the evolution of the Alaskan flora over the whole of the Cenozoic (Figure 2) but have especially enriched the paleobotanical understanding of arctic floras during the Eocene and Oligocene (Figure 3). Early Eocene floras from the Kulthieth Formation in Wrangell–St. Elias National Park and Preserve as well as floras from the Kushtaka Formation to the south document “boreotopical” forests, a non-analog vegetation type characterized by rainforest-like leaf physiognomy and the presence of some lineages now restricted to the tropics and subtropics of Asia (Wolfe, Reference Wolfe1977, Reference Wolfe, Boulter and Fisher1994a, Reference Wolfe1994b; Wing, Reference Wing, Janis, Scott and Jacobs1998; Graham, Reference Graham1999). Middle and Late Eocene floras are known from Aniakchak National Monument and Preserve and Katmai National Park and Preserve on the Alaskan Peninsula (Hollick, Reference Hollick1936; Wolfe, Reference Wolfe1966), as well as from the Katalla and Rex Creek floras of Central Alaska (Wolfe, Reference Wolfe, Prothero and Berggren1992), and illustrate a step in the modernization of the flora, with an increased presence of the broadleaf deciduous lineages (e.g., Betulaceae) characteristic of modern North American forests. Oligocene and Miocene floras, known from NPS units on the Alaskan Peninsula and Central Alaska as well as a number of sites outside of park boundaries (Wolfe, Reference Wolfe1977; Wing, Reference Wing, Janis, Scott and Jacobs1998), represent further modernization of the flora by complete loss of broadleaf evergreen lineages and their replacement by broadleaf deciduous plants and conifers, with an apparent peak in the diversity of the broadleaf deciduous component during the Early Miocene (Wolfe and Tanai, Reference Wolfe and Tanai1980).
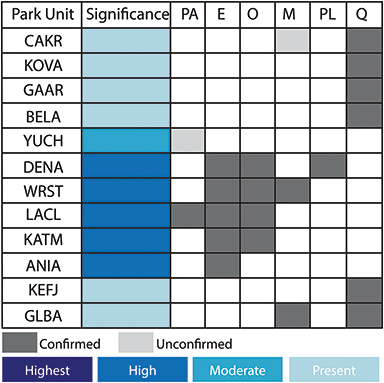
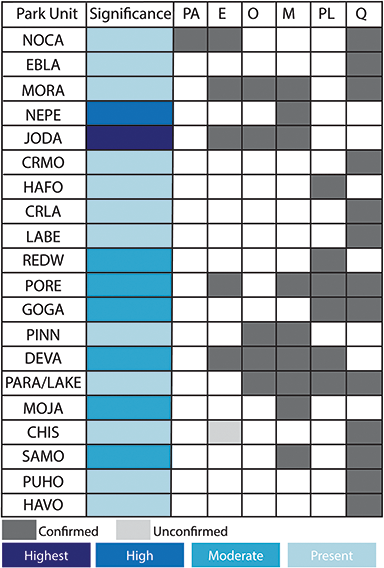
Figure 2 Summary of the temporal distribution and significance of Cenozoic paleobotanical resources in the Alaska Region. The significance rating of individual parks represents the aggregate significance of all resources within that park and was scored based on criteria such as holotype specimens, plant fossil abundance and preservation quality, and number of publications referencing primary paleobotanical data from the park.
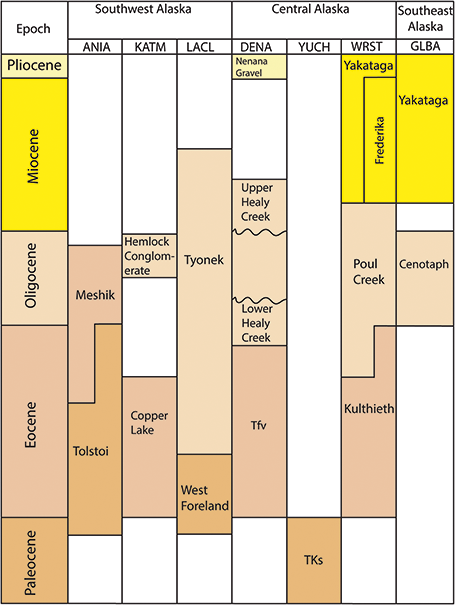
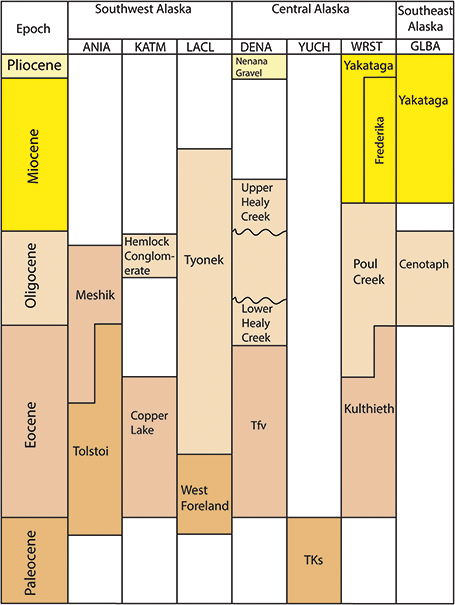
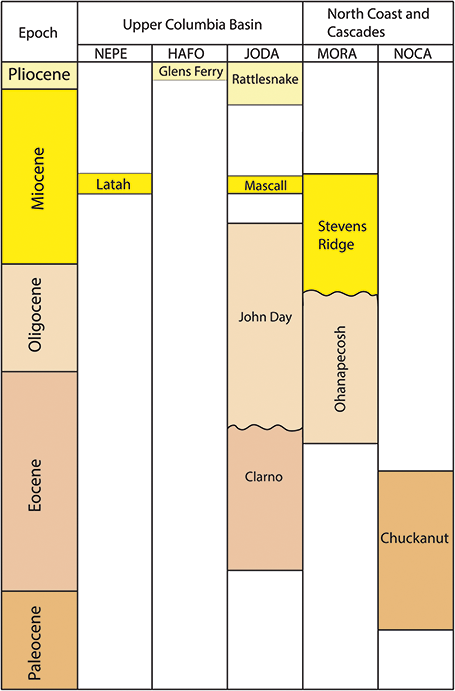
Figure 3 Stratigraphic chart of Tertiary plant-fossil–bearing formations in the Alaska Region.
Numerous additional Alaskan paleofloras are preserved outside of the NPS and add important context for interpreting and managing park resources. Much of the primary documentation of these resources comes from the monographic works of Swiss paleobotanist Oswald Heer and USGS paleobotanists Arthur Hollick and Jack Wolfe (Heer, Reference Heer1869; Hollick, Reference Hollick1930, Reference Hollick1936; Wolfe, Reference Wolfe1966, Reference Wolfe1977; Wolfe et al., Reference Wolfe, Hopkins and Leopold1966), and the Tertiary (used here for convenience for pre-Quaternary Cenozoic) record in Alaska has been reviewed several times (Wolfe, Reference Wolfe and Graham1972; Wing, Reference Wing, Janis, Scott and Jacobs1998; Graham, Reference Graham1999).
3.1.1 Arctic Alaska Inventory and Monitoring Network
Cape Krusenstern National Monument
Cape Krusenstern National Monument contains approximately 650,000 acres of land along the northwestern coast of Alaska (Figure 1). The monument was established in 1978 to protect a series of beach ridges along the southern coast (Lanik et al., Reference Lanik, Swanson and Karpilo2019). Unidentified fossil wood recovered from these ridges is the only report of paleobotanical material within the park (Mason and Ludwig, Reference Mason and Ludwig1990).
Kobuk Valley National Park
Kobuk Valley National Park preserves approximately 1.75 million acres of the Kobuk River Valley and borders Noatak National Preserve to the north (Figure 1). The only report of Cenozoic paleobotanical material within the park comes from Epiguruk Bluffs, a locality on the Kobuk River containing fossil vertebrates, mollusks, and wood. Wood samples used for radiocarbon dating constrain the age of the deposits to 37,000–14,000 years before present (ybp) (Hamilton and Ashley, 1983; Hamilton et al., Reference Hamilton, Ashley, Reed and van Etten1984, Reference Hamilton, Ashley, Reed and Schweger1993; Lanik et al., Reference Lanik, Swanson and Karpilo2019).
Gates of the Arctic National Park and Preserve
Gates of the Arctic National Park and Preserve (GAAR) lies entirely north of the Arctic Circle and is the nation’s second largest national park, encompassing nearly 8.5 million acres (Figure 1). Paleobotanical material collected from GAAR includes plant fragments from unnamed lacustrine sediments on the north side of Noatak Valley just north of Okturak Creek (Forester, Reference Forester1979) and fossil wood from overbank deposits of the west bank of the Killik River (Repenning, Reference Repenning1974).
Bering Land Bridge National Preserve
Bering Land Bridge National Preserve (BELA) covers 2.8 million acres of tundra on the Seward Peninsula of Alaska (Figure 1). Quaternary paleobotanical resources within BELA have been discovered in eroding permafrost sediments (pingo deposits), tephra deposits of the Devil Mountain maar exposed on banks of thermokarst lakes, and in gravel deposits along the banks of the Noxapaga River (Spackman, Reference Spackman1951; Goetchus and Birks, Reference Goetcheus and Birks2001; Wetterich et al., Reference Wetterich, Grosse and Schirrmeister2012). Just east of BELA in the vicinity of Lava Camp Mine, is a volcanic lava flow dated to 5.7 ± 0.2 Ma (Late Miocene) that preserved a forested floodplain (Hopkins et al., Reference Hopkins, Matthews, Wolfe and Silberman1971). The museum collection at BELA holds a fossil leaf and fossil liverwort specimen, but these are not linked to locality or age information (Lanik et al., Reference Lanik, Swanson and Karpilo2019).
Lava Camp Mine: The Lava Camp Mine locality (USGS 11190) in the Imuruk River Valley just east of the park boundary preserves plant macrofossils in alluvial deposits that were buried under an upper Miocene (5.7 Ma) basaltic lava flow (Hopkins et al., Reference Hopkins, Matthews, Wolfe and Silberman1971). The macrofossil assemblage represents a floodplain forest composed largely of Picea and Betula. Its taxonomic composition is similar to approximately contemporaneous deposits from the Cook Inlet Region, and together these floras represent the Clamgulchian (Late Miocene–Pliocene) Alaskan time-stratigraphic stage (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Hopkins et al., Reference Hopkins, Matthews, Wolfe and Silberman1971). Macrofossil types include needle leaves, ovulate cones, and seeds; taxa include Picea (three spp.), Pinus, Tsuga, Betula, Symphoricarpos, Vaccinium, Carex, and Cyperus (Hopkins et al., Reference Hopkins, Matthews, Wolfe and Silberman1971). The locality also preserves a diverse insect and arachnid assemblage consisting of at least 83 species (Elias et al., Reference Elias, Kuzmina and Kiselyov2006). Parts of the southeast corner of the park contain upper Tertiary (Neogene) volcanic deposits (map unit Qtv; NPS, 2015 [unpublished digital geologic map]), which might preserve similar communities.
Noxopaga River: Fossil wood identified as Populus and Picea was recovered from a 12 m (40 ft) section of Quaternary gravel deposits exposed on the west bank of the Noxopaga River, between Goose and Bussard creeks (Spackman, Reference Spackman1951).
Pingo Deposits: Wetterich et al. (Reference Wetterich, Grosse and Schirrmeister2012) collected samples of pingo deposits from Whitefish maar, Devil Mountain maar, and North and South Killeak maars in the Cape Espenberg lowlands (within BELA) of the Seward Peninsula and from nine other localities (outside BELA) in Arctic and Central Alaska. The combined age range of these samples spans 46,615–8,986 ybp, and the oldest sediments containing identifiable plant macrofossils were constrained to 32,870 ± 220 ybp. The macrofossil assemblage includes twigs, leaves, seeds, and fruits representing gymnosperms and woody and herbaceous angiosperms. Common plant genera include Betula, Carex, Hippuris, Potamogeton, Potentilla, and various members of the Ericaceae.
Devil Mountain Lake Tephra: The Devil Mountain Lake tephra buried approximately 2500 km2 (970 mi2) of land surface, reaching a maximum thickness of 1 m (3 ft) over an approximately 1200 km2 (460 mi2) region (Goetchus and Birks, Reference Goetcheus and Birks2001). The tephra was subsequently capped by loess, preserving an in situ tundra landscape, and the sequence has recently become exposed along the banks of thermokarst lakes. Plant macrofossils and organic material used for radiocarbon dating were sampled from twelve different localities in the Cape Espenberg–Devil Mountain area and produced an age range of 18,260–17,700 radiocarbon ybp (ca. 21,500 calendar ybp). This age range places the deposits within the thermal minimum of the Last Glacial Maximum (28,000–14,000 ybp). The macrofossil assemblages are interpreted to represent a sedge-dominated, herb-rich tundra grassland based on the relative ground cover of moss, graminoids, forbs, and shrubs in point-transect collections and the composition of seeds and fruits isolated from sieved sediment. Representative taxa include Carex, Salix, Papaver, and Potentilla (Goetchus and Birks, Reference Goetcheus and Birks2001).
3.1.2 Central Alaska Inventory and Monitoring Network
Yukon–Charley Rivers National Preserve
Yukon–Charley Rivers National Preserve (YUCH) encompasses approximately 2.5 million acres (Figure 1). This area protects about 161 km (100 mi) of the Yukon River and the entire Charley River basin. The park’s geology spans nearly 1.3 billion years of Earth’s history, from the Precambrian to the Cenozoic (Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). The park does not contain any confirmed Cenozoic paleobotanical resources; however, it does contain exposures of Upper Cretaceous and Paleocene rocks (map unit TKs; Figure 4), which have yielded Paleocene plant fossils just outside of the park’s southern boundary (Hollick, Reference Hollick1930, Reference Hollick1936; Mertie, Reference Mertie1942) and Cretaceous plant fossils from within the park (Knoll, Reference Knoll and Young1976; Tiffney, Reference Tiffney and Young1976; Fiorillo et al., Reference Fiorillo, Fanti, Hults and Hasiotis2014).

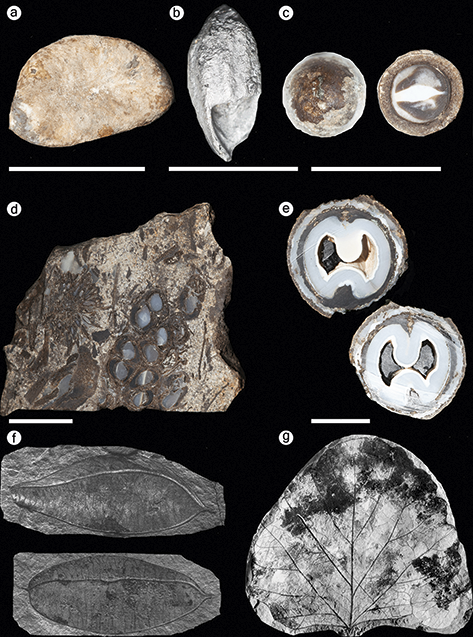
Figure 4 Fossil plants from the Alaska Region. (a) Paratinomiscium conditionalis, a leaf belonging to the Menispermaceae recovered from the Yakutat Bay locality in Wrangell–St. Elias National Park and Preserve. Image from Hollick (Reference Hollick1936). (b) Corylus harrimani Knowlton collected from Katmai National Park and Preserve. Scale bar = 5 cm. (c) Ptermospermites alaskana collected from Katmai National Park and Preserve. Scale bar = 5 cm. (d) Holotype of Acer disputabilis collected from Katmai National Park and Preserve. Scale bar = 1 cm. (e) Myrica banksiaefolia var. curta collected from Katmai National Park and Preserve. Scale bar = 1 cm. (f) Specimen of Picea harrimani collected from Katmai National Park and Preserve. Scale bar = 1 cm. (g) Holotype specimen of Aesculus arctica collected from Katmai National Park and Preserve. Scale bar = 5 cm. (h) Holotype specimen of Hicoria magnifica collected from Katmai National Park and Preserve. Scale bar = 5 cm. (i) Alnus evidens, recovered from the Kukak Bay locality in Katmai National Park and Preserve. This taxon was formerly placed in the genus Corylus. Image from Hollick (Reference Hollick1936).
Unnamed unit (TKs): Plant macrofossils and palynomorphs are preserved in a continuous Upper Cretaceous to lower Cenozoic depositional sequence mapped as unit TKd (Foster, Reference Foster1976) or, more commonly, as TKs (Mertie, Reference Mertie1930; Knoll, Reference Knoll and Young1976; Dover and Miyaoka, Reference Dover and Miyaoka1988; Miyaoka, Reference Miyaoka1990). This unit is laterally continuous from the southeast corner of the park (near Eagle, Alaska) to the northwest of the park (south of Circle, Alaska). It forms a belt approximately 145 km (90 mi) long and parallel to the course of the Yukon River, and its Paleogene deposits are thickest and most laterally extensive in its southern and eastern regions (Mertie, Reference Mertie1942; Tiffney, Reference Tiffney and Young1976). The subdivision of the sequence represented by map unit TKs into smaller stratigraphic units is likely warranted but has been hindered by the unit’s large areal extent, variable lithology, and widely spaced fossil localities (Tiffney, Reference Tiffney and Young1976).
At least 28 plant macrofossil localities are documented from TKs in the Eagle–Circle district (Mertie, Reference Mertie1942). Many of the early discoveries of fossil localities from this formation occurred in the southeastern region of YUCH and just outside the park’s southern boundary (Hollick, Reference Hollick1930, Reference Hollick1936; Mertie, Reference Mertie1942). Among these localities are several Early Cenozoic sites along the banks of Seventymile Creek and one of its tributaries, Bryant Creek, which have yielded well-preserved angiosperm leaf fossils, several of which were described as new species (Hollick, Reference Hollick1936). Geologic maps indicate that there is significant potential to find similar localities within the southeastern region of YUCH, which is approximately 8 km (5 mi) to the north.
Denali National Park and Preserve
Denali National Park and Preserve (DENA), encompasses approximately 6 million acres and protects the large glaciers of the Alaska range as well as a diverse megafauna (Figure 1). The park’s geology is complex, and study and interpretation by the NPS, USGS, and academic researchers are ongoing (Csejtey et al., Reference Csejtey, Mullen and Cox1986; Tomsich et al., Reference Tomsich, McCarthy, Fowell and Sunderlin2010; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). Cenozoic paleobotanical material has been recovered from at least three different formations within DENA: an unnamed Lower Eocene map unit labeled unit Tfv, the Upper Oligocene Healy Creek Formation, and the Upper Miocene to Pliocene Nenana Gravel (Figure 4; Csejtey et al., Reference Csejtey, Mullen and Cox1986; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). The Cantwell Formation, once considered to be Paleocene in age, has also yielded plant fossils within DENA; however, recent studies (Tomisch et al., Reference Tomsich, McCarthy, Fowell and Sunderlin2010) indicate a Late Cretaceous age for the Cantwell Formation.
Fluviatile and subordinate volcanic rocks (Eocene?), unit Tfv: While developing a geologic map for the Healy Quadrangle, Csejtey et al. (Reference Csejtey, Mullen and Cox1986) documented two plant fossil localities within DENA that are ascribed to an unnamed map unit labeled Tfv. These localities are in the eastern-central area of the park between Costello Creek and the western fork of the Chulitna River (see Csejtey et al., Reference Csejtey, Mullen and Cox1986, for further details). The proposed Eocene age of this unit is based only on its sparse plant fossils and correlation with other Early Cenozoic Alaskan fossil assemblages. Plant fossils from this unnamed unit were found in a shale interbed within conglomerate and include needles of Metasequoia occidentalis and fragments of broad-leaved dicotyledons (Csejtey et al., Reference Csejtey, Mullen and Cox1986). This sample was studied by paleobotanist Jack Wolfe, who suggested that the dicotyledonous component represented a plant community younger than the Cantwell Formation, which he then believed to be Paleocene in age (J. A. Wolfe, pers. comm. in Csejtey et al., Reference Csejtey, Mullen and Cox1986).
Healy Creek Formation: The Healy Creek Formation is the basal unit of the Usibelli Group (also known as the Tertiary coal-bearing group, or map unit Tcu of Nenana Coalfield), which encompasses five formations that span from the Oligocene to Miocene (Figure 4; Wahrhaftig et al., Reference Wahrhaftig, Wolfe, Leopold and Lanphere1969). A collection of plant fossils assigned to this group was made at the top of the section exposed at Dunkle Coal Mine, in the same east-central region of DENA, which also yielded plant fossils from the unnamed unit (unit Tfv) already discussed (Csejtey et al., Reference Csejtey, Mullen and Cox1986). This assemblage includes twelve species described from leaf compressions and impressions. Cercidiphyllum crenatum and Alnus evidens are notable from a biostratigraphic standpoint and suggest that the assemblage belongs to the Angoonian (Late Oligocene) Alaskan time-stratigraphic stage (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966). Other taxa present in this collection include the fern Onoclea, the gymnosperm Metasequoia cf. glyptostroboides, and broad-leaved angiosperms such as Betula, Salix, Populus, and Corylus. Elsewhere in DENA, Bruce Reed collected an assemblage including Metasequoia cf. glyptostoboides, Alnus evidens, “Carya” magnifica, Equisetum, and undetermined species of Alnus and Populus at USGS localities 11123–11125. The localities are assigned to an unnamed unit composed of conglomerate, sandstone, and siltstone interpreted as Oligocene in age on the basis of its plant fossils (Wolfe, Reference Wolfe1968). It is possible that this assemblage was recovered from unmapped exposures of the Healy Creek Formation.
Outside of DENA, in the Nenana Coalfield, the Healy Creek Formation has yielded similar assemblages to those recovered from the park; these assemblages also include Ulmus and Quercus (Wahrhaftig et al., Reference Wahrhaftig, Wolfe, Leopold and Lanphere1969). In the Nenana Coalfield, the Healy Creek deposits are in conformable contact with other paleobotanically productive formations of the Usibelli Group, and there is potential to discover similar sequences in the northeastern region of DENA where the Usibelli Group is exposed in the Savage River drainage and near Sable Pass.
Nenana Gravel: The Nenana Gravel includes conglomerate, sandstone, claystone, and lignite that accumulated during uplift and erosion of the Alaska Range in the Pliocene. The unit is exposed along portions of the park road in the park’s northeast corner (Csejtey et al., Reference Csejtey, Mullen, Cox and Stricker1992; Wilson et al., Reference Wilson, Dover and Bradley1998; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). Within DENA, the only paleobotanical materials recovered from the gravel to date are a Picea cone and carbonized wood in bajada deposits (Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). Leopold and Liu (Reference Leopold and Liu1994) reported carbonized plant fragments and fossil fruits of Trapa from probable deposits of the Nenana Gravel northeast of the park near Ferry, Alaska.
Wrangell–St. Elias National Park and Preserve
Wrangell–St. Elias National Park and Preserve (WRST) in southeast Alaska is the largest national park in the USA, encompassing more than 13 million acres (Figure 1). The park was established to conserve a region that includes three major mountain ranges – the Chugach, St. Elias, and Wrangell mountains – numerous glaciers, and wildlife habitat ranging from temperate rainforest to tundra. The park also contains significant Cenozoic paleobotanical resources. In 1905, R. S. Tarr was the first to collect plant fossils within what is now WRST, from outcrops of the Paleocene–Eocene Kulthieth Formation along Esker Stream near Yakutat Bay. The collection from this locality was later studied by USGS paleobotanists Arthur Hollick and Jack Wolfe, yielding type specimens of several species (Figure 4; Hollick, Reference Hollick1936; Wolfe, Reference Wolfe1977). Later collecting efforts, also led by the USGS, yielded plant specimens from three additional formations: the Eocene–Miocene Poul Creek Formation, the Miocene Frederika Formation, and the Miocene–Pleistocene Yakataga Formation (Figure 4; Wolfe, Reference Wolfe1977; Fremd et al., Reference Fremd, Dunn, Rickabaugh, Graham and Rosenkrans2003; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011).
Kulthieth Formation: Fossil plants from the Kulthieth Formation were recovered in a calcareous sandstone layer at USGS paleobotanical locality 3879 along Esker Stream, which flows into Yakutat Bay in the southeast corner of the park (Hollick, Reference Hollick1936). The stratigraphic position of this locality within the Kulthieth Formation is uncertain. Wolfe (Reference Wolfe1977) initially referred the Kulthieth Formation to the Middle Eocene based on mollusk fossils occurring in intertonguing marine rocks (Marincovich and McCoy, Reference Marincovich and McCoy1984), but its planktonic microfossils indicate an Early Eocene age, potentially within the Early Eocene Climatic Optimum (Wolfe, Reference Wolfe, Boulter and Fisher1994a). The fossil assemblage collected at Esker Stream includes ferns, gymnosperms, and angiosperms represented by leaf compressions and impressions and one fossil endocarp (Figure 4). This locality produced type specimens of several species that belong to angiosperm families that do not occur in Alaska today: Celastrus comparabilis Hollick in the Celastraceae, Paratinomiscium conditionalis in the Menispermaceae (Figure 4), and Paleophytocrene elytraeformis in the Icacinaceae; it also produced the lectotype specimen of an extinct species of wood fern, Dryopteris alaskana. Wolfe (Reference Wolfe1977) considered the fossil flora collected from the Kulthieth Formation to represent “Paratropical Rainforests” based on its high proportion of large and entire-margined leaves and the presence of the families Annonaceae and Icacinaceae, which mainly occur in the tropics today. Climate reconstructions based on leaf physiognomy of 54 species from the Kulthieth Formation estimate that the assemblage lived under the warmest climate of any Alaskan Paleogene flora, with a mean annual temperature of 19.4°C (66.9°F) (Wolfe, Reference Wolfe, Boulter and Fisher1994a).
Poul Creek Formation: Fossil plants from the Poul Creek Formation were collected from USGS paleobotanical locality 11185, which occurs near Marvine Glacier in the southeast corner of the park (Wolfe, Reference Wolfe1977). Molluscan biostratigraphy of the Poul Creek Formation suggests that its sediments were deposited between the Late Eocene and Early Miocene (Figure 3), and the presence of Alnus evidens at USGS 11185, an indicator of the Angoonian (Late Oligocene) floral stage in Alaska, suggests an Oligocene depositional age for the formation’s fossil-plant–bearing strata (Wolfe, Reference Wolfe1977; Marincovich and McCoy, Reference Marincovich and McCoy1984; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). Elsewhere, the Poul Creek Formation is known as a marine unit, but USGS 11185 occurs along a fault in which no marine fossils were found (Stoneley, Reference Stoneley1967; Wolfe, Reference Wolfe1977). The plant remains recovered from the Poul Creek Formation are fossil leaves assigned to the angiosperms Alnus evidens and Cercidiphyllum crenatum and the gymnosperm Metasequoia glyptostroboides (Wolfe, Reference Wolfe1977). Outside of WRST, a well-preserved permineralized walnut, Juglans lacunosa, was illustrated from Poul Creek locality USGS 11182 (figure 48 p, q, in Manchester, Reference Manchester1987).
Frederika Formation: The Frederika Formation occurs in the Wrangell Volcanic Fields, and outcrops extend into the northern and eastern regions of the park (Richter et al., Reference Richter, Preller, Labay and Shew2006). The valley walls of a westward-flowing tributary of Frederika Creek (Skolai Creek) are the type area of the Frederika Formation and the location of USGS localities 9933, 9935, and 9927, each of which falls within park boundaries (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Wolfe and Tanai, Reference Wolfe and Tanai1987). The Frederika Formation preserves paleobotanical material in a variety of depositional contexts, including paleosol sequences, thick coal beds, lacustrine deposits, and silty tuffaceous sandstones (Fremd et al., Reference Fremd, Dunn, Rickabaugh, Graham and Rosenkrans2003). The plant fossils recovered from the Frederika Formation in the Skolai Creek area include angiosperms common in temperate deciduous forests, such as Acer, Alnus, Betula, and Populus, as well as the conifers Pinus and Metasequoia (Wolfe, Reference Wolfe1966; Santucci et al., Reference Santucci, Blodgett, Elder, Tweet and Kenworthy2011). Collectively, this assemblage suggests a Miocene age (Wolfe, Reference Wolfe1966). Wolfe (Reference Wolfe1964) remarked that the Pinaceae are better represented in the Frederika Formation than in contemporaneous floras of Cook Inlet and speculated that the difference in plant composition could be linked to the greater topographic relief and elevation of the Wrangell Mountains during the Miocene.
Yakataga Formation: The Yakataga Formation mainly comprises marine sequences but includes two paleobotanical localities (USGS 11183 and 11184) established in strata that have a similar lithology to the marine beds (Wolfe, Reference Wolfe1977). Both localities are in the southeastern area of the park, near Yakutat Bay. The locality USGS 11183 occurs in exposures near Haenke Glacier and USGS 11184 occurs in the Pinnacle Hills area. Floral and faunal biostratigraphy of the Yakataga Formation exposures that produced the plant-fossil–bearing horizons suggest that they were deposited in the Miocene (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Wolfe, Reference Wolfe1977). Respectively, USGS 11183 and 11184 were referred to the Seldovian (Early to Middle Miocene) and Homerian (Middle to Late Miocene) floral stages based on the presence of Alnus appsii and Carpinus cobbii (Wolfe, Reference Wolfe1970, Reference Wolfe1977; Addicott et al., Reference Addicott, Winkler and Plafker1978). Fossil leaves of the fern Osmunda and the gymnosperm Metasequoia glyptostroboides were also recovered from the Yakataga Formation in WRST, and a fossil Fagus leaf was recovered from the Yakataga Formation in Glacier Bay National Park and Preserve (see Section 3.1.4) (Wolfe, Reference Wolfe1977).
3.1.3 Southwest Alaska Inventory and Monitoring Network
Lake Clark National Park and Preserve
Lake Clark National Park and Preserve (LACL) encompasses 4 million acres of a dynamic natural landscape renowned for its coastline along Cook Inlet, glaciers, glacial lakes, rugged mountain peaks, and two active volcanoes, Mount Iliamna and Mount Redoubt (Figure 1). The park preserves one of the most important Mesozoic marine fossil localities in Alaska at Fossil Point, located on the Cook Inlet coast in Tuxedni Bay (Kenworthy and Santucci, Reference Kenworthy and Santucci2003). Younger rocks of the Paleocene–Lower Eocene West Foreland Formation and the Oligocene–Middle Miocene Tyonek Formation also crop out along the Cook Inlet coast and contain plant macrofossils (Figure 3; Hollick, Reference Hollick1936; Wolfe et al., Reference Wolfe, Hopkins and Leopold1966).
West Foreland Formation: Exposures of the West Foreland Formation on the north shore of Chinitna Bay in the Iniskin-Tuxedni Region have produced fossil wood and leaves from locality USGS 3505 (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Ruga et al., Reference Ruga, Lanik and Hults2020). The West Foreland Formation is considered to be Late Paleocene–Early Eocene in age based on its plant fossils (Magoon et al., Reference Magoon, Adkison and Chmelik1976a, citing an oral comm. with Wolfe). Calderwood and Fackler (Reference Calderwood and Fackler1972) considered it to be the basal formation of the Kenai Group, a sequence of botanically fossiliferous terrestrial deposits spanning the Upper Paleocene to the Pliocene. The holotype specimen of Ginkgo reniformis var. conformis Hollick was collected from USGS 3505, and other taxa also found at this locality include Ginkgo biloba, Metasequoia glyptostroboides, and Taxodium tinajorum (Hollick, Reference Hollick1936; Wolfe et al., Reference Wolfe, Hopkins and Leopold1966). Fossil tree stumps exposed on the north shore of Chinitna Bay were discovered in 2019. They have not been studied taxonomically (or otherwise) and should be a management priority because they are at high risk of being lost to erosion (Ruga et al., Reference Ruga, Lanik and Hults2020). An additional macrofossil locality of the West Foreland Formation has been found just north of the park boundary near Redoubt Point (Magoon et al., Reference Magoon, Adkison and Egbert1976b), and there is potential to find additional localities in the Iniskin-Tuxedni Region within LACL.
Tyonek Formation: USGS paleobotanical localities 9760, 9887, 11344, 11355, and 11367 were established from exposures of the Tyonek Formation within park boundaries at Redoubt Point (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Wolfe and Tanai, Reference Wolfe and Tanai1987). Like the Kukak Bay localities in Katmai National Park and Preserve (see below), these deposits are considered to be Upper Oligocene based on the presence of taxa characteristic of the Angoonian floral stage, such as Alnus evidens and Corylus harrimanii (Wolfe et al., Reference Wolfe, Hopkins and Leopold1966; Wolfe and Tanai, Reference Wolfe and Tanai1980). Other taxa collected from the Redoubt Point flora include the conifers, Glyptostrobus europaeus and Metasequoia glyptostroboides, and angiosperms, Vaccinium, Populus cf. kenaina, Carpinus cappensis, and the holotype specimen of Acer kenaicum. Wolfe (Reference Wolfe1994b) included seventeen leaf morphotypes from the Redoubt Point assemblage in a foliar physiognomy-based paleoclimate reconstruction and concluded that the assemblage grew in the coolest climate of any Paleogene Alaskan flora, with a mean annual temperature of 9°C (48.2°F).
Katmai National Park and Preserve
Katmai National Park and Preserve (KATM) encompasses approximately 4 million acres on the Alaska Peninsula (Figure 1). It is about 160 km (100 mi) northeast of Aniakchak National Monument and Preserve and about 240 km (150 mi) southwest of Lake Clark National Park and Preserve. The park contains a multitude of paleontological resources and has rich Mesozoic marine and Paleogene terrestrial records (Harrington et al., Reference Harrington, Lanik, Hults and Druckenmiller2023). Fossil plants were first discovered from what is now KATM in exposures at Kukak Bay, now mapped as the Upper Oligocene Hemlock Conglomerate. This collection was made by DeAulton Saunders, a member of the June–July 1899 Harriman Expedition funded by E. H. Harriman, and was studied by the paleobotanist F. H. Knowlton (Knowlton, Reference Knowlton, Emerson, Palache, Dall, Ulrich and Knowlton1904). Continued exploration of the region has led to the discovery of dozens of other plant fossil localities inside KATM, belonging to the Hemlock Conglomerate and the Paleocene–Lower Eocene Copper Lake Formation (Magoon et al., Reference Magoon, Adkison and Egbert1976b; Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996).
Copper Lake Formation: Fossiliferous exposures of the Copper Lake Formation have been found in the northeastern area of the park near Cape Douglas and at Dumpling Mountain in the western region of the park (Blodgett et al., Reference Blodgett, Santucci and Tweet2016). The fossil flora at Cape Douglas is preserved in sandstone and siltstone layers that overly a basal conglomerate. The age of the plant-fossil–bearing horizons is interpreted as Early Eocene on the basis of its taxonomic composition (Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996, citing a written comm. from J. A. Wolfe). The plant fossils recovered from the Copper Lake Formation at Cape Douglas are leaf compressions and impressions that include three species of ferns, the gymnosperm Sequoia langsdorfii, and six species of angiosperms. Taxa named from this flora include the fern Osmunda dubiosa and the angiosperms Myrica banksiaefolia var. curta, Crataegus alaskensis, and Acer douglasense (Hollick, Reference Hollick1936; Wolfe and Tanai, Reference Wolfe and Tanai1987; Figure 4).
The assemblage of fossil leaves and wood recovered at Dumpling Mountain is also interpreted to date from the Late Paleocene or Early Eocene, and its minimum age is constrained to the Late Eocene based on an intruding diorite dike dated to 39.63 ± 0.17 Ma (Parrish et al., Reference Parrish, Fiorillo, Jacobs, Currano and Wheeler2010). Twelve distinct leaf morphotypes were identified from 45 collected specimens and include several species – such as Cercidiphyllum genetrix, Glyptostrobus europaeus, and Platanus raynoldsii – that were widespread in Alaska and the western USA during the Paleogene (Parrish et al., Reference Parrish, Fiorillo, Jacobs, Currano and Wheeler2010). Notably, none of the morphotypes described from Dumpling Mountain represent species identified from the Cape Douglas localities that occur about 160 km (100 mi) to the east, raising the intriguing possibility that the two assemblages represent compositionally distinct but contemporaneous ecological communities. The fossil wood specimens discovered at Dumpling Mountain are well preserved and belong to four species of conifers, including a new species of Pinus Section Parrya, and the platanaceous wood genus Platanoxylon (Parrish et al., Reference Parrish, Fiorillo, Jacobs, Currano and Wheeler2010).
Hemlock Conglomerate: Fossil plants from the Hemlock Conglomerate have been collected from several localities along the southeastern coast of KATM, including the historical Kukak Bay locality (Knowlton, Reference Knowlton, Emerson, Palache, Dall, Ulrich and Knowlton1904) and a reference section of the Hemlock Conglomerate measured at Cape Nukshak (Keller and Reiser, Reference Keller and Reiser1959). These collections include leaves of ferns, gymnosperms, and angiosperms, as well as conifer and angiosperm reproductive structures (Knowlton, Reference Knowlton, Emerson, Palache, Dall, Ulrich and Knowlton1904; Hollick, Reference Hollick1936; Wolfe and Tanai, Reference Wolfe and Tanai1987). Common taxa include the gymnosperms Picea, Pinus, Sequoia, and Taxodium and the angiosperms Acer, Aesculus, Corylus, and Alnus (Figure 4). The collections also include holotypes of 12 taxa (Supplementary Table 7).
Aniakchak National Monument and Preserve
Aniakchak National Monument and Preserve (ANIA), located on the Alaskan Peninsula, was established to preserve Aniakchak Caldera and its surrounding lands (Figure 1). The park contains 600,000 acres and is rich in paleontological resources from the Jurassic, Cretaceous, and Paleogene (Kenworthy and Santucci, Reference Kenworthy and Santucci2003). The Paleogene resources include plant fossils of the Tolstoi and Meshik formations (Hollick, Reference Hollick1936; Wolfe, Reference Wolfe1980).
Tolstoi Formation: The Tolstoi Formation crops out widely on the Alaska Peninsula, and the age of its strata ranges from Early Paleocene to Middle Eocene (Wolfe, Reference Wolfe1980; Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996). At its type section near Pavlof Bay, about 240 km (150 mi) southwest of ANIA, the Tolstoi Formation represents a warm to subtropical shallow marine environment and contains marine mollusks and dinoflagellates indicative of a Middle Eocene age (Marincovich, Reference Marincovich, Filewicz and Squires1988; Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996). At its reference section near Ivanof Bay, about 135 km (85 mi) southwest of ANIA, and in exposures to the northeast (including those in the park), the Tolstoi Formation represents fluvial floodplain and delta sequences and contains a diverse flora indicative of a Paleocene–Eocene age and a warm, wet environment (Wolfe, Reference Wolfe1981; Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996). The plant fossil localities of the Tolstoi Formation within ANIA occur throughout the formation’s stratigraphic range and are widely spaced, from the southeastern boundary near Elephant Head Point to the northeastern boundary, where the park borders the Alaskan Peninsula National Wildlife Refuge along Pumice and Painter creeks (Hollick, Reference Hollick1936; Wolfe, Reference Wolfe1980; Kenworthy and Santucci, Reference Kenworthy and Santucci2003).
In the northeastern area of the park, plant macrofossil localities occur near Painter and Pumice creeks. Sample P-23 was collected by W. R. Smith in 1922 from a mountain southwest of Pumice Creek and yielded the lectotype of the fern Dryopteris meyeri, the holotype specimen of Acer disputabilis (Figure 4), and other angiosperm leaves assigned to Ulmus braunii, Populus arctica, and Zizyphus hyperboreus (Hollick, Reference Hollick1936). The USGS paleobotanical localities 11577–11580 were discovered in several intervals of a stratigraphic section near Painter Creek and yielded an assemblage including a tree fern, Cyathea inequalateralis; a conifer, M. occidentalis; and the angiosperms Pterocarya nigelloides, Viburnum variabilis, and undetermined species of Acer and Platycarya (Wolfe, Reference Wolfe1980). Wolfe (Reference Wolfe1980) suggested that these localities are Early to Middle Eocene in age and represent the youngest strata of the Tolstoi Formation. Two more localities are present in the central region of the park near Jaw Mountain. Sample P-25 was collected by W. R. Smith in 1922 and includes leaf fossils of the angiosperm family Rhamnaceae (Z. hyperboreus and Paliurus colombi) and a fossil flower of an extinct species of Hydrangea (Hydrangea alaskana). Locality USGS 11632 was established near a peak 4.0 km (2.5 mi) south of Jaw Mountain and yielded a leaf belonging to a member of the Menispermaceae (Cocculus flabella) and needles of the conifers Metasequoia and Glyptostrobus. This assemblage is characteristic of the lower Tolstoi deposits and interpreted to be late Paleocene in age (Wolfe, Reference Wolfe1980). Exposures of the Tolstoi Formation near Elephant Head Point on Cape Kumlik in the southeastern region of ANIA produced a small collection of plant fossils, which includes undetermined species of Alnus and Vitis (Wolfe, Reference Wolfe1980).
Meshik Formation: The Meshik Formation or Meshik Volcanics is a volcanic unit that contains intervals of volcaniclastic sedimentary rock (Detterman et al., Reference Detterman, Miller, Yount and Wilson1981). One such interval, comprising sandstone and water-deposited tuff, is the source of USGS paleobotany locality 11640. These beds, exposed near Aniakchak Crater in the western region of Aniakchak National Monument, have yielded a diverse, angiosperm-rich flora including Metasequoia, Cercidiphyllum, Quercus, Pterocarya, Ulmus, Cornus, various Rosaceae, and the holotype specimen of an extinct species of maple, Acer dettermanii (USNM 396014) (Wolfe and Tanai, Reference Wolfe and Tanai1987; Wolfe, Reference Wolfe, Boulter and Fisher1994a). Wolfe (Reference Wolfe1980) interpreted this assemblage as Middle Eocene in age, an estimate that falls in the early part of the age range suggested by K-Ar dates of the tuff (43.1 ± 1.3 to 24.9 ± 0.49 Ma; Detterman et al., Reference Detterman, Case, Miller, Wilson and Yount1996). Wolfe (Reference Wolfe, Boulter and Fisher1994a) used 34 leaf morphotypes from this site to estimate its paleoclimate based on leaf physiognomy and characterized the assemblage as a notophyllous broad-leaved evergreen forest with a mean annual temperature of 13.7°C (56.7°F). Detterman et al. (Reference Detterman, Miller, Yount and Wilson1981) also reported well-preserved petrified logs, some occurring in original growth position, in exposures of the Meshik Formation on the south shore of Kujulik Bay just south of the southern park boundary.
Kenai Fjords National Park
Kenai Fjords National Park (KEFJ) is located on the Kenai Peninsula in Alaska, preserves about 600,000 acres, and includes the 780 km2 (300 mi2) Harding Ice Field, one of four ice caps in North America (Figure 1). Potentially in situ logs and woody debris were discovered near Exit Glacier, about 20 km (12 mi) northwest of Seward, Alaska. Initial radiocarbon dating of the woody material from KEFJ indicates an age of 900 years, suggesting that it may represent a forest community buried by an advancing glacier (Kenworthy and Santucci, Reference Kenworthy and Santucci2003).
3.1.4 Southeast Alaska Inventory and Monitoring Network
Glacier Bay National Park and Preserve
Glacier Bay National Park and Preserve (GLBA), located in the northeastern part of southeastern Alaska, preserves about 3.3 million acres of land rich in floral and faunal biodiversity and is home to globally recognized tidewater glaciers (Figure 1). Although the area of the park is extensively glaciated, plant macrofossils have been discovered from outcrops of the Oligocene Cenotaph Formation and Miocene–Pleistocene Yakataga Formation on Cenotaph Island, and abundant subfossil Holocene wood has been discovered from Muir Inlet (Figure 3).
Cenotaph Formation: The Cenotaph Formation is an Oligocene volcanic unit interbedded with marine and terrestrial sedimentary rocks that crops out in the Lituya Bay Region, including on Cenotaph Island and on the mainland south of the bay. The plant fossil locality 75-Apr-37 occurs in horizons of the type section of the Cenotaph Formation on Cenotaph Island and has produced angiosperm leaves attributed to Macclintockia pugetensis, Magnolia reticulata, and undetermined species of Pterocarya and the Lauraceae (Wolfe, Reference Wolfe1976). M. pugetensis, an extinct species of uncertain familial affinity, is restricted to the Kummerian (Oligocene) floral stage in Alaska, and therefore provides strong biostratigraphic correlation with other plant-fossil–bearing units from the Gulf of Alaska (Wolfe, Reference Wolfe1977). Plafker (Reference Plafker1967) included the Cenotaph Formation within the Topsy Formation, which consists of Middle Oligocene–Upper Miocene shallow marine deposits that are conformably overlain by the Yakataga Formation.
Yakataga Formation: Within GLBA, the Yakataga Formation is exposed along the coast between Lituya Bay and Icy Point, on Cenotaph Island in Lituya Bay, and north and south of Fairweather Glacier (Santucci and Kenworthy, Reference Santucci and Kenworthy2008). Exposures on the south shore of Cenotaph Island (USGS paleobotany locality 11186) produced a single leaf fossil assigned to Fagus (beech) and are the only source of paleobotanical material from the Yakataga Formation within GLBA. Elsewhere within and beyond the boundaries of GLBA, the Yakataga Formation is known as a marine unit and has produced fossils of foraminifera, bivalves, and gastropods that suggest a Middle Miocene–Pleistocene age (MacNeil et al., Reference MacNeil, Wolfe, Miller and Hopkins1961; Santucci and Kenworthy, Reference Santucci and Kenworthy2008).
Muir Inlet: An early reference to the sub-fossil wood and standing stumps present in what is now called Muir Inlet was made by the naturalist John Muir in the book Travels in Alaska (Reference Muir1915). Tree-ring cores and radiocarbon age estimates from these trees have been compiled with records from living trees in GLBA to reconstruct forest succession and paleoclimate in the region over about the past 10,000 years (Lawson et al., Reference Lawson, Wiles and Finnegan2007; Lawson and Wiles, Reference Lawson and Wiles2009).
Park Collections: The park collections include three pieces of basal peat deposits and two pieces of petrified wood. The peat was collected from a terrace above Echo Creek north of Lituya Bay and dated to 40,000 ybp. Wood specimens were collected by C. V. Janda from the west side of Adams Valley and from the beach at Dundas Bay and identified as belonging to a conifer and an angiosperm, respectively (Santucci and Kenworthy, Reference Santucci and Kenworthy2008).
3.2 Pacific West Region
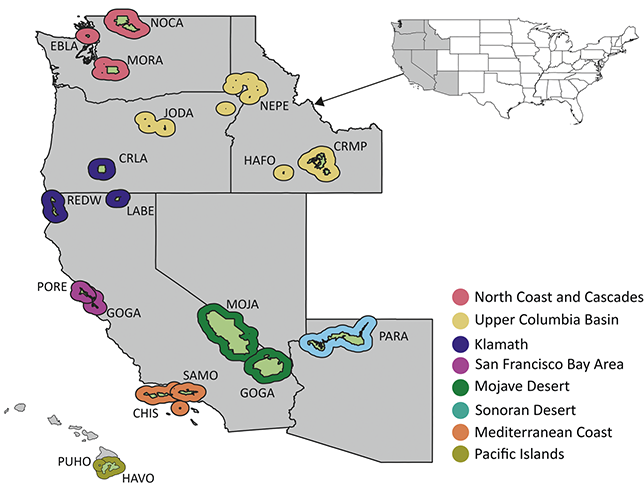
The Pacific West Region includes 64 NPS units in the states and territories of American Samoa, California, Guam, Hawai’i, Idaho, Nevada, Oregon, and Washington. The parks of this region are clustered in seven inventory and monitoring networks (Figure 5): the Klamath Network (KLMN), the Mediterranean Coast Network (MEDN), the Mojave Desert Network, the North Columbia and Cascades Network, the Pacific Islands Network, the San Francisco Bay Area Network (SFAN), and the Upper Columbia Basin Network. More than 25 geologic formations spanning the Paleocene to Holocene from 20 park units in the Pacific West Region have confirmed paleobotanical resources.
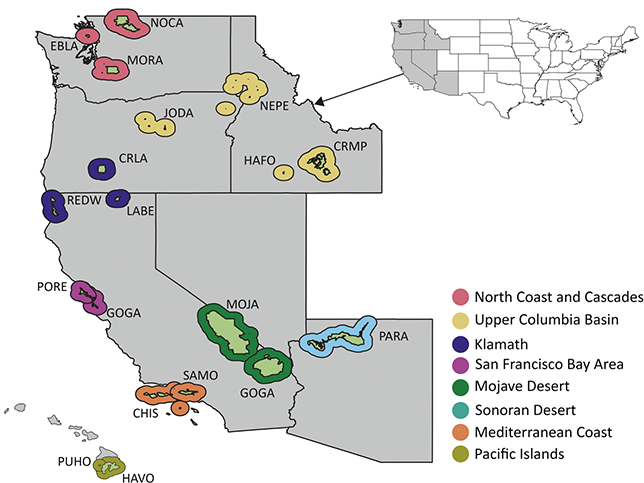
Figure 5 Map of NPS units with Cenozoic paleobotanical resources in the Pacific West Region, colored by inventory and monitoring network. See Figure 1 legend for details on the park symbology and acronyms.
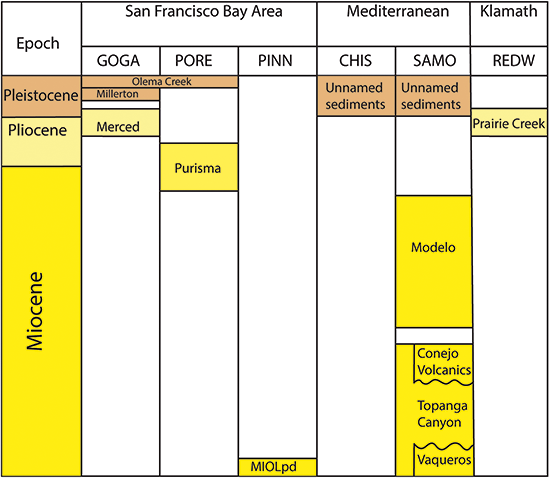
Significant resources from the Pacific West Region include classic paleobotanical localities such as the Eocene Clarno Nut Beds in John Day Fossil Beds National Monument and the Miocene Whitebird site of Nez Perce National Historical Park, as well as a number of lesser-known Plio–Pleistocene sites in the KLMN, MEDN, and SFAN networks of coastal California (Figure 6). The anatomically preserved fruits and seeds of the Clarno Nut Beds represent a phase in the vegetational history of the Pacific Northwest when the flora included a mixture of plants with familial affinities to groups mainly known from tropical and subtropical areas today (e.g., Icacinaceae, Menispermaceae) as well as to lineages characteristic of temperate North American forests (e.g., Platanaceae). The insight into the Eocene vegetational history of the Pacific West provided by the Clarno Nut Beds is complemented by assemblages preserved outside of NPS units, such as the floras of the Puget Group (Burnham, Reference Burnham1994) and Chuckanut Formation (Mustoe and Gannaway, Reference Mustoe and Gannaway1997) in Washington state and the Goshen and Comstock floras of Oregon (see Wing, Reference Wing, Janis, Scott and Jacobs1998; Graham, Reference Graham1999; and DeVore and Pigg, Reference DeVore and Pigg2010 for reviews of Eocene Pacific Northwest floras).
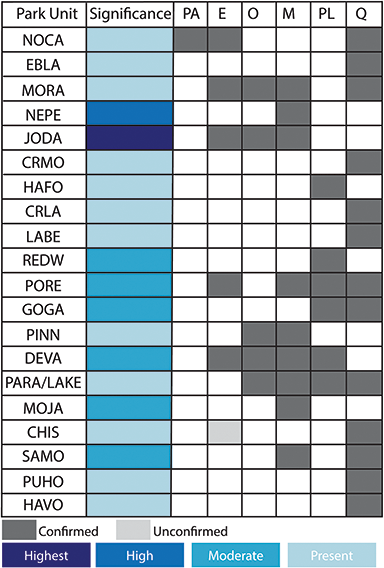
Figure 6 Summary of the temporal distribution and significance of Cenozoic paleobotanical resources in the Pacific West Region. See Figure 2 legend for explanation of significance ratings.
The Miocene Whitebird site, deposited ca. 30 million years later in western Idaho, includes a greater proportion of broadleaf deciduous angiosperms and conifers characteristic of the extant North American flora, though not necessarily of the Pacific Northwest. Miocene floras from the Columbia Plateau occurring outside of NPS areas are well-documented (Chaney and Axelrod, Reference Chaney and Axelrod1959; Graham, Reference Graham and Graham1965), and recent work has placed them into a stratigraphic context to illustrate changes in floral composition during the Middle Miocene Climatic Optimum (Lowe, Reference Lowe2024).
Fossil plants from the Plio–Pleistocene are preserved in Redwood National Park, Golden Gate National Recreation Area, Point Reyes National Recreation Area, Channel Islands National Park, and Santa Monica Mountains National Recreation Area. These assemblages mainly include genera and species that still occur along the California coast today and have been used to trace the evolutionary history of regional vegetation types such as the California closed-cone pine forest (Axelrod, Reference Axelrod and Philbrick1967).
3.2.1 North Coast and Cascades Inventory and Monitoring Network
Northern Cascades National Park Service Complex
The Northern Cascades National Park Service Complex (NOCA) is situated in the state of Washington (Figure 5) and was established to facilitate recreational enjoyment of the North Cascade Mountains and associated glaciers, watercourses, and ecosystems. Cenozoic paleobotanical resources are preserved in NOCA in three contexts: lower Cenozoic sandstones, Quaternary lake sediments, and Quaternary mass wasting deposits.
Chuckanut Formation/Tertiary Sandstones: Sediments of the Chuckanut Formation were deposited in a coastal plain during the Paleocene and Eocene and are well-exposed throughout northwestern Washington and southwestern British Colombia (Figure 7; Mustoe et al., Reference Mustoe, Dillhoff and Dillhoff2007; Fay et al., Reference Fay, Kenworthy and Santucci2009). Within NOCA, non-diagnostic plant fragments and a fossil log have been found in strata that are variously included within the Chuckanut Formation (Staatz et al., Reference Staatz, Tabor and Weis1972) or within an informal sedimentary unit comprised of sandstones and conglomerates (Tabor et al., Reference Tabor, Haugerud, Hildreth and Brown2003). Outside of NOCA, in Chuckanut outcrops near Bellingham, Washington, the formation has produced abundant plant fossils, including palm leaves and foliage of tree ferns, gymnosperms, and woody angiosperms (Mustoe and Gannaway, Reference Mustoe and Gannaway1995; Mustoe et al., Reference Mustoe, Dillhoff and Dillhoff2007; Mustoe, Reference Mustoe2019). These fossil assemblages are preserved in three different members of the Chuckanut Formation and the stratigraphic succession of plant communities they represent is interpreted to reflect a local transition from a warmer and wetter climate to a dryer and cooler climate, potentially related to the climatic deterioration of the EOT or episodes of local mountain-building (Mustoe and Gannaway, Reference Mustoe and Gannaway1997). Well-preserved plant fossils come from fine-grained portions of the Chuckanut Formation, and if such strata are present within NOCA, they are likely stratigraphically close to the fossiliferous sandstone.
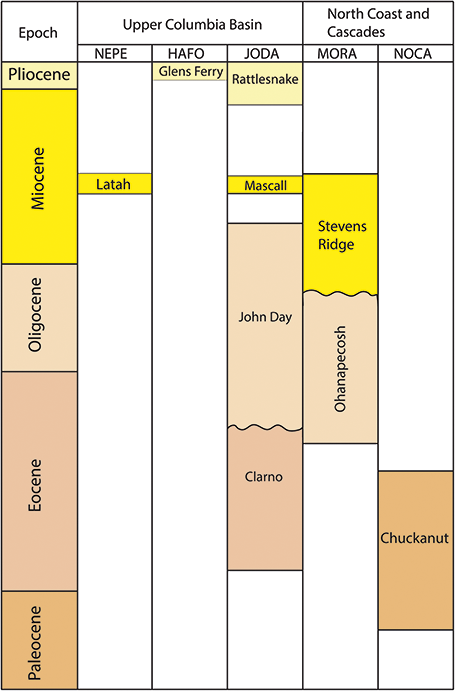
Figure 7 Stratigraphic chart of Tertiary plant-fossil–bearing formations in the Upper Columbia Basin and Northern Coast and Cascades Inventory and Monitoring Networks.
Quaternary sediments: Plant macrofossils, pollen, and charcoal have been recovered from Ridley and Thunder lakes and Skagit Valley, all located in the northeastern corner of the park. The cores from Ridley and Thunder lakes preserve a paleoenvironmental record of the past 14,000 years, and their floras include many plants that inhabit the region today such as Abies, Chamaecyparis, Picea, Pinus, Tsuga, Alnus, Betula, and Fraxinus (Spooner et al., Reference Spooner, Brubaker and Foit2008; Fay et al., Reference Fay, Kenworthy and Santucci2009). The macrofossils from Skagit Valley document vegetational changes between 24,000 and 17,110 radiocarbon ybp (Riedel, Reference Riedel2007; Fay et al., Reference Fay, Kenworthy and Santucci2009). Older sediments of this interval are dominated by conifer foliage of Pinus albicaulis, Picea engelmannii, and Abies lasiocarpa, and leaves of Salix and fragments of Selaginella become more abundant in the younger sediments. Finally, trees buried in growth position are preserved in ca. 7,000-year-old lake deposits that formed after a landslide dammed the Upper Skagit River (Riedel et al., Reference Riedel, Pringle and Schuster2001).
Ebey’s Landing National Historical Reserve
Ebey’s Landing National Historical Reserve (EBLA), located on Whidbey Island of the San Juan Archipelago in Washington state, encompasses about 20,000 acres and includes two state parks and the historic town of Coupeville (Figure 5). The surficial geology of EBLA is dominated by unconsolidated Quaternary sediments that were deposited by glacial and fluvial processes (Fay et al., Reference Fay, Kenworthy and Santucci2009). The paleobotanical material preserved in these sediments is limited to a few minor occurrences. Sediments accumulated during the Olympia nonglacial interval (ca. 60,000–20,000 ybp) preserve pollen, pinecones, branches, leaf prints, and in situ tree roots (Troost, Reference Troost2002); wood found in an estuary, possibly of Douglas fir, was radiocarbon dated to 1740–1790 or 1810–1960 CE (Polenz et al., Reference Polenz, Slaughter, Dragovich and Thorsen2005a, Reference Polenz, Slaughter and Thorsen2005b).
Mount Rainier National Park
Mount Rainier National Park (MORA) preserves approximately 235,000 acres of land surrounding the peak of Mount Rainier or Tahoma, in the central portion of the Cascade Range of southwestern Washington (Figure 5). The park contains paleobotanical resources in volcaniclastic strata of the Ohanapecosh and Stevens Ridge formations deposited between the Eocene and Miocene and in Holocene lake sediments and mass-wasting deposits.
Ohanapecosh Formation: The Ohanapecosh Formation is comprised of up to 3,000 m (9,800 ft) of volcanic and volcaniclastic sediments within MORA (Fay et al., Reference Fay, Kenworthy and Santucci2009). The only paleobotanical material yet recovered from these strata within MORA are fragments of wood and leaves, but layers of well-preserved fossil leaves and a forest buried by a lahar deposit are known from correlative strata outside the park. Localities nearby MORA that have yielded significant fossil plant assemblages include a site 600 m (2,000 ft) west of the park boundary on the north side of North Puyallup River, sites on Osborne Mountain and at Catt Creek 3 km (2 mi) southwest of MORA, and a lahar deposit that preserves permineralized wood 24 km (15 mi) west of MORA, near the town of Mineral, Washington (Fay et al., Reference Fay, Kenworthy and Santucci2009). The leaf floras collected near the park were determined by Roland Brown to include woody angiosperms such as Alnus, Betula, Castanea, Cercidiphyllum (katsura tree), Cinnamomum, Laurus, and Platanus, as well as the palm Sabal, the gymnosperm Metasequoia, and the fern Thelypteris (Brown, Reference Brown1956; Fisher, Reference Fisher1957). The fossil leaves preserved in the lower and upper strata of the Ohanapecosh Formation along Catt Creek are the basis of the Late Eocene–Oligocene age assigned to this unit (Figure 7). Fay et al. (Reference Fay, Kenworthy and Santucci2009) suggested that similar layers may be accessed within MORA near Round Pass and in the saddle just north of Tumtum Peak.
The park collections of MORA contain six paleobotanical specimens that represent conifer needles and wood and angiosperm leaves with a possible affinity to the Fagaceae (Regan Dunn, pers. comm., 2009). Only one of these specimens (MORA 13164) has associated geographic information, and this specimen was recovered in 2006 from the Ohanapecosh River upstream from the Stevens Canyon landslide (Fay et al., Reference Fay, Kenworthy and Santucci2009). The Ohanapecosh Formation is widely exposed in this drainage basin and is a potential candidate for the source of this specimen.
Stevens Ridge Formation: The Stevens Ridge Formation is predominantly composed of quartz-bearing ash flows and unconformably overlies the Ohanapecosh Formation. A freshwater gastropod (Viviparus) of Late Oligocene or Miocene age collected from a quarry about 13 km (8 mi) south of MORA was tentatively correlated with the Stevens Ridge strata exposed in the park (Fiske et al., Reference Fiske, Hopson and Waters1963). Conifer needles are present at several localities of the Stevens Ridge Formation within MORA (Fiske et al., Reference Fiske, Hopson and Waters1963).
Holocene lake sediments and mass wasting deposits: Paleobotanical resources preserved in Holocene lake sediments and mass wasting deposits facilitate the study of recent landscape evolution in MORA by recording local vegetational change and by constraining the age of glacial and tectonic events that lead to mass wasting. Macrofossils sieved from sediment cores taken from three lakes on the south side of Mount Rainier have been used to establish a 6,000-year record of forest succession (Dunwiddie, Reference Dunwiddie1983, Reference Dunwiddie1985, Reference Dunwiddie1986, Reference Dunwiddie1987), and Holocene wood preserved in lahars has been used to constrain the age of numerous postglacial lahars by radiocarbon dating and dendrochronology (Pringle and Scott, Reference Pringle and Scott2001; Pringle, Reference Pringle2008).
3.2.2 Upper Columbia Basin Inventory and Monitoring Network
Nez Perce National Historical Park
Nez Perce National Historical Park (NEPE) includes 38 sites in Idaho, Montana, Oregon, and Washington that commemorate the stories and history of the Nimiipuu or Nez Perce people (Figure 5). Six of these sites are on land owned by the National Park Service, and one of these six sites, the Whitebird Battlefield, the location of the first battle of the Nez Perce Flight of 1877, includes exposures of the fossiliferous Latah Formation.
Latah Formation: The classic White Bird locality of the Latah Formation is exposed in a highway roadcut that traverses the Whitebird Battlefield site of NEPE. The Latah Formation is characterized as a sequence of lake beds, ash, sands, and silts interbedded in the Colombia River Basalt, and the formation’s fossiliferous strata outcrop extensively in Idaho and eastern Washington (Kirkham and Johnson, Reference Kirkham and Johnson1929). A tuff dated from the top of the section of the Latah Formation at White Bird provides a minimum age of 14.3 ± 0.1 Ma, but a slightly older age of 15.5–15.8 Ma is likely, based on correlation with the Bully Creek Formation in eastern Oregon (Nash and Perkins, Reference Nash and Perkins2012; Figure 7). The paleobotanical resources of the Latah Formation in the vicinity of Whitebird, Idaho, were first documented by Kirkham and Johnson (Reference Kirkham and Johnson1929), and their original specimens and localities have been featured in several subsequent studies, notably Berry (Reference Berry1930, Reference Berry1932, Reference Berry1934), Ashlee (Reference Ashlee1932), Brown (Reference Brown1935, Reference Brown1937), Herendeen and Dilcher (Reference Herendeen and Dilcher1991), and Manchester et al. (Reference Manchester, Crane and Dilcher1991). Of the localities mapped by Kirkham and Johnson (Reference Kirkham and Johnson1929), stations 8, 9, 12, and 14 (hereafter, White Bird site) are within or adjacent to NEPE, and station 14, which yielded the most species, is probably equivalent to USGS 8444.
Macrofossils from the Latah flora recovered from the White Bird site include attached and isolated foliage, fruits, and seeds that represent 97 species and at least 24 families (Figures 8f and 8g; Kirkham and Johnson, Reference Kirkham and Johnson1929; Ashlee, Reference Ashlee1932). The flora is dominated numerically and in number of species by angiosperms; the genera with the greatest representation at White Bird localities are Acer (maple), Quercus, Populus, and Betula (Berry, Reference Berry1932). Gymnosperms from White Bird include Glyptostrobus, Juniperus, Pinus (pine), Taxodium, and Tsuga, and pteridophytes include Equisetum, Polystichum, and Pteris (Ashlee, Reference Ashlee1932). Several genera represented in the paleoflora, including Cassia, Castanea, Euonymus, Fagus, Glyptostrobus, Liquidambar, Magnolia, Malva, Nyssa, Ostrya, and Sophora, are not found in the region today (Berry, Reference Berry1932).
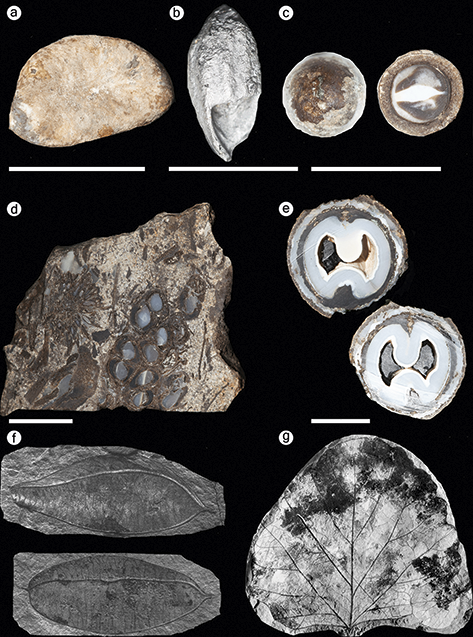
Figure 8 Fossil plants from the Pacific West Region. All scales = 1 cm. (a–e) Fossils from the Clarno Nut Beds of John Day National Monument. (a) Holotype specimen of Anonaspermum bonesii representing the seed of an extinct member of the tropical plant family Annonaceae. (b) Holotype specimen of Nyssa scottii. (c) Holotype of Laurocarpum nutbedensis representing the fruit of an extinct member of the tropical plant family Lauraceae. (d) Holotype specimen of Fortunearites endressii representing the infructescence of an extinct member of the Hamamelidaceae. (e) Holotype of Anonmyocarpa ovoideae representing a fruit of unknown familial affinity. (f) Specimen of Caesalpinia spokanensis collected from the Whitebird site of the Latah Formation in Nez Perce National Historic Park. (g) Specimen of Vitis washingtonensis from the Whitebird site.
The White Bird site has yielded holotype specimens of Juniperus sabinoides, Ceratophyllum praedomersum, Aristolochia whitebirdensis, and Viburnum whitebirdensis, and paratypes of Caesalpinia spokanensis, Zizyphoides auriculata, and Nordenskioldia interglacialis (Ashlee, Reference Ashlee1932; Herendeen and Dilcher, Reference Herendeen and Dilcher1991; Manchester et al., Reference Manchester, Crane and Dilcher1991). C. spokanensis (Figure 8) includes a fossil fruit collected from White Bird that served as the holotype for the species Cercis idahoensis, now a synonym of C. spokanensis (Berry, Reference Berry1930; Brown, Reference Brown1937; Herendeen and Dilcher, Reference Herendeen and Dilcher1991). Z. auriculata and N. interglacialis are respectively based on leaf and reproductive fossils that co-occur at Miocene localities throughout the Pacific Northwest and are thought to belong to the same taxon (Manchester et al., Reference Manchester, Crane and Dilcher1991). Furthermore, these leaves and fruits are, respectively, congeneric with the taxa Zizyphoides flabellum and Nordenskioldia borealis, which occur in association at Paleocene localities in North America, Greenland, and Norway, and at Paleocene and Eocene localities in eastern Asia (Manchester et al., Reference Manchester, Crane and Dilcher1991).
Outside of NEPE, the Latah Formation has produced several other notable fossil assemblages in eastern Washington and western Idaho, including the Spokane and Cour d’Alene (Knowlton, Reference Knowlton1926), Clarkia (Smiley and Rember, Reference Smiley, Rember and Smiley1985; Rember, Reference Rember1991), Juliaetta (Kirkham and Johnson, Reference Kirkham and Johnson1929), and Grand Coulee (Berry, Reference Berry1932) floras. Collectively, these floras document the diversity, composition, and structure of Pacific Northwest forests during the Miocene Climatic Optimum (Lowe, Reference Lowe2024), and in the case of the Clarkia flora, offer rare insight into the paleobiochemistry of plant organs through the exceptional preservation of leaf cuticular and mesophyll tissues (Logan et al., Reference Logan, Boon and Eglinton1993), as well as pigmentation (Niklas and Brown, Reference Niklas and Brown1981), secondary metabolites (Otto et al., Reference Otto, Simoneit and Rember2003), and potentially DNA (Golenberg et al., Reference Golenberg, Giannasi and Clegg1990).
John Day Fossil Beds National Monument
The John Day Fossil Beds National Monument (JODA) in central Oregon consists of three separate protected areas: the Clarno, Painted Hills, and Sheep Rock units. Together they provide access to a succession of Eocene to Miocene fossiliferous strata and volcanic rocks, including the Clarno and John Day formations, the Picture Gorge Basalt, the Mascall Formation, and the Rattlesnake Formation. A geological field trip guide (Dillhoff et al., Reference Dillhoff, Dillhoff, Dunn, Myers and Strömberg2009) provides a useful overview of the local geology and paleontology. The Clarno Unit includes Eocene floras of the Clarno Nut Beds, Hancock (Mammal) Quarry, and Hancock Canyon. The Painted Hills Unit includes several localities of the Early Oligocene Bridge Creek flora studied by Newberry (Reference Newberry1898), Lesquereux (Reference Lesquereux1883), Knowlton (Reference Knowlton1902), Chaney (Reference Chaney1927), and Meyer and Manchester (Reference Meyer and Manchester1997). The Sheep Rock Unit, where the National Monument is headquartered, includes the Cant Ranch flora (Meyer and Manchester, Reference Meyer and Manchester1997) and is adjacent to the Miocene Mascall flora (Knowlton, Reference Knowlton1902; Chaney, Reference Chaney1927).
Clarno Formation: The Clarno Formation comprises terrestrial volcanic and volcaniclastic rocks deposited between ca. 54 and 39 Ma (Figure 7) and contains a diverse and well-preserved flora based on wood, leaf, fruit, seed, and palynomorph fossils (Figure 8; Bestland and Retallack, Reference Bestland and Retallack1994; Manchester, Reference Manchester1994; Wheeler and Manchester, Reference Wheeler and Manchester2002). The flora of the Clarno Formation preserves subtropical and warm temperate fossil plants including horsetails, ferns, cycads, ginkgo, pine, sequoia, palms, banana relatives, and numerous other angiosperms representing both extant and extinct genera.
The Nut Beds site near Hancock Field Station in the Clarno Unit of JODA was so-named by amateur fossil collectors who discovered the location in the 1930s with agatized walnuts and various other nut and seeds preserved as casts, molds, and permineralizations. The fruit and seed flora was initially monographed by Scott (Reference Scott1954), who recognized 10 species, plus six indeterminate additional morphotypes. Continued amateur collecting through the 1970s resulted in greatly expanded collections, such that Manchester (Reference Manchester1994) was able to discriminate 145 genera and 173 species representing a warm temperate to subtropical environment. The flora includes numerous extinct genera, but also early records of genera still living today, such as Juglans, Quercus, Celtis, Magnolia, Iodes, Vitis, and Ampelocissus. Woody vines are particularly well represented, including multiple genera of the families Menispermaceae, Icacinaceae (e.g., Iodes), and Vitaceae (e.g., Ampelocissus, Vitis).
The Nut Beds site is unusual in having various plant parts preserved in association with vertebrate remains. Bones and teeth of several mammals and a crocodilian have been recognized (Hanson, Reference Hanson, Prothero and Emry1996). In addition to bones, nuts, and seeds, the site provides the richest known Eocene petrified wood flora worldwide (Wheeler and Manchester, Reference Wheeler and Manchester2002), including 66 genera and 76 species. Fossil leaves are well preserved in the Nut Bed siltstones, with excellent details of venation, but are very fragmentary and time-consuming to collect, requiring gluing of adjacent fragments to obtain complete leaves (Manchester, Reference Manchester1981; Smith, Reference Smith2023). A survey of dispersed pollen and spores has also been conducted (Smith, Reference Smith2023).
The Hancock Quarry, also referred to as Mammal Quarry, dates to about 40 Ma, 4 million years younger than the Clarno Nut Beds (Bestland and Retallack, Reference Bestland and Retallack1994). The quarry was excavated in the 1960s and 1970s for vertebrate fossils and also yielded fruits and seeds (McKee, Reference McKee1970). The flora includes Juglans, Vitis, Icacinaceae, and Menispermaceae. One of the moonseeds was originally treated as Diploclisia, a genus today found in Asia, but more recently recognized as Stephania jacquesii (Han et al., Reference Han, Manchester, Fu, Jin and Quan2018). Stephania is native to the paleotropics and Australasia today (POWO, 2023).
Lahar deposits exposed in Hancock Canyon yield petrified wood, leaves, and occasional fruits. These were investigated by Wheeler and Manchester (Reference Manchester2014). Cycad leaves of genus Ctenis were found in the Hancock Canyon deposits adjacent to Hancock Field Station as well as in the Nut Beds (Erdei and Manchester, Reference Erdei and Manchester2015).
John Day Formation: The John Day Formation is composed of tuffs and alluvial deposits and is divided into four members that span from the Late Eocene to the Early Miocene (Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). The Big Basin Member contains fossiliferous Upper Eocene and Lower Oligocene strata, the latter of which preserve the Bridge Creek flora. The Early Oligocene Bridge Creek flora was first recognized based on collections made by pioneering paleontologist and reverend Thomas Condon from the John Day Formation near the Painted Hills, an area now within John Day National Monument. Two other Bridge Creek localities, Cove Creek and Iron Mountain, are within or immediately adjacent to JODA.
The collections were initially studied by Newberry (Reference Newberry1898), Lesquereux (Reference Lesquereux1883), and Knowlton (Reference Knowlton1902), and subsequently by Chaney (Reference Chaney1927) and Meyer and Manchester (Reference Meyer and Manchester1997). The Bridge Creek flora at Painted Hills and Cant Ranch includes a temperate assemblage with Metasequoia, Alnus, Fagus, Quercus, Acer, Tilia, and a few extinct genera such as Asterocarpinus (Betulaceae) and Florissantia (Malvaceae). In the Cove Creek flora, the taxa represented by the greatest number of specimens are Alnus and Paracarpinus, a fossil leaf genus associated with Asterocarpinus. The Iron Mountain assemblage is similar to the other sites, including abundant fossils of Metasequoia, Alnus, Acer, Asterocarpinus, and Paracarpinus, but it also includes unique elements such as Paleophytocrene, an extinct genus of Icacinaceae; Zingiberopsis, a fossil genus of the ginger family (Zingiberaceae); and a relatively high abundance of Toxicodendron (Meyer and Manchester, Reference Meyer and Manchester1997). The Iron Mountain assemblage has the greatest richness of the three sites, preserving a minimum of 44 species, compared to 35 in the Cove Creek assemblage and 38 in the Painted Hill assemblage.
Mascall Formation: The Mascall Formation dates to ca. 15 Ma and comprises rhyolitic tuffs with associated sands and gravels (Bestland and Retallack, Reference Bestland and Retallack1994). Paleofloras from the Mascall Formation are primarily found outside of JODA, with the exception of unidentified plant fossils reported from the Mascall Overlook in the park’s Sheep Rock Unit (Rensberger, Reference Rensberger1976; Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). A number of woody angiosperm and gymnosperm taxa have been recovered from the Mascall Formation near JODA, including Ginkgo, Sequoia, Quercus, Alnus, Salix, Acer, Ficus, and Rhus (Chaney, Reference Chaney1925; Chaney and Axelrod, Reference Chaney and Axelrod1959).
Rattlesnake Formation: The Rattlesnake Formation is an Upper Miocene unit composed of fanglomerates and is present in the Sheep Rock Unit of JODA. Paleobotanical material is comparatively sparse and includes reports of Platanus, Ulmus, and Salix fossils (Fremd et al., Reference Fremd, Bestland and Retallack1994; Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005).
Craters of the Moon National Monument and Preserve
Craters of the Moon National Monument and Preserve (CRMO) in Idaho was established to preserve a portion of the lava flows in the Craters of the Moon Lava Field (Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). These lava flows contain abundant tree molds, “lava trees” – upright, cylindrical molds with the impression of bark on the inside – and carbonized wood. More than 140 tree molds have been discovered and mapped in the northern end of the park (Owen and Melander, Reference Owen and Melander2014). The Blue Dragon and Trench Mortar Flat lava flows have the largest number of tree molds, and Stearns (Reference Stearns1928) also reported tree molds within CRMO from the “northwest flow from Big Craters,” Owl Cavern flow, and Black flow (Stearns, Reference Stearns1928; Owen and Melander, Reference Owen and Melander2014). Carbonized wood was found in some of the tree molds and has been used for carbon-14 dating, revealing the dates of the flows with wood to span from 15,000 to 1,700 ybp (Bonnichsen and Breckenridge, Reference Bonnichsen and Breckenridge1982; Kuntz et al., Reference Kuntz, Lefevre, Champion and Skipp1989; Owen and Melander, Reference Owen and Melander2014). Upright lava trees are up to 1.5 m (5 ft) tall and have been found at Trench Mortar Flat, the Tree Mold Flat lava flows, and the wilderness area of CRMO (Stearns, Reference Stearns1928).
Hagerman Fossil Beds National Monument
The Hagerman Fossil Beds National Monument (HAFO) in Hagerman, Idaho, is renowned for its late Pliocene vertebrate faunas that include the “Hagerman Horse” (Equus simplicidens), the earliest recognized member of the extant genus Equus (Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). Paleobotanical material collected from HAFO is limited to pollen and rare occurrences of fossil wood.
Glenns Ferry Formation: The Pliocene Glenns Ferry Formation represents deposition in lacustrine, floodplain, and fluvial environments and contains basalt flows, silicic volcanic ash, and basaltic pyroclastic deposits that constrain the age of the unit to the Pliocene (ca. 5–3 Ma) (Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). Fossil wood has been occasionally found in the Glenns Ferry Formation but has not been formally studied. One well-preserved specimen showing anatomical detail was reported in the National Park Service Morning Report for December 23, 1993 (Kenworthy et al., Reference Kenworthy, Santucci, McNerney and Snell2005). Palynomorphs isolated from sediments of this unit reveal a community of conifers with sparse hardwoods and many steppe elements in the Chenopodiaceae and Asteraceae (Leopold and Denton, Reference Leopold and Denton1987). The palynomorph assemblage becomes increasingly rich in grass pollen over time and, alongside the coeval vertebrate fauna, documents the transition of this region from forests to an open steppe and grassland environment (Leopold and Denton, Reference Leopold and Denton1987).
3.2.3 Klamath Inventory and Monitoring Network
Crater Lake National Park
Crater Lake National Park (CRLA) in the Cascade Range of southwestern Oregon preserves a caldera lake that formed after the eruption and collapse of Mount Mazama 7,700 ybp. Carbonized tree stumps are preserved in Pleistocene glacial till deposits, and carbonized wood is present in Holocene pumice fall deposits and ignimbrite deposits (Santucci and Kenworthy, Reference Santucci and Kenworthy2009). The estimated age of the eruption that produced Crater Lake is based on radiocarbon dating of charcoal twigs preserved in ignimbrite (Bacon, Reference Bacon2008).
Lava Beds National Monument
Lava Beds National Monument in the Cascade Range of northern California preserves flows that contain tree molds, charcoal, and charred wood. Three such flows have been identified inside the monument, namely the Valentine Cave (2,260 ybp), Black Crater (3,080 ybp), and Callahan flows (1,120 ybp) (KellerLynn, Reference KellerLynn2014). Charred wood and charcoal from these flows were used for radiocarbon dating (KellerLynn, Reference KellerLynn2014).
Redwood National Park and State Parks
Redwood National Park (REDW) was established to preserve temperate rainforests along the Pacific coast of northern California that include the world’s tallest trees, the coast redwood (Sequoia sempervirens). The park encompasses four units – Redwood National Park, and Jedediah Smith, Del Norte, and Prairie Creek state parks – that are cooperatively managed by the state of California and the federal government. The Prairie Creek State Park unit of REDW contains a Late Pliocene flora based on tree and root casts, logs, leaf impressions, carbonized leaves, cones, and peat (Criswell, 1968; Trexler, Reference Trexler1989).
Prairie Creek Formation: Plant fossils of the Prairie Creek Formation have been recovered from at least six horizons within a 244 m (800 ft) section of conglomerate, sandstone, and siltstone exposed on the coastal cliffs of the Gold Bluff area within Prairie Creek State Park (Santucci and Kenworthy, Reference Santucci and Kenworthy2009). The sequence is interpreted to have been deposited during the Late Pliocene and Early Pleistocene (Figure 9) in a fluvial floodplain of the ancestral Klamath River, and its fossil assemblage comprises eighteen species of plants, an unidentified fossil vertebrate, and two species of freshwater pelecypods (Criswell, 1968). The Prairie Creek flora includes the liverwort Marchantia, the fern Dryopteris, herbaceous and woody angiosperms – including Acer, Alnus, and Betula – and gymnosperms, including S. sempervirens. Casts of trees infilled with sand and associated with carbonized wood are also present but have not been identified (Trexler, Reference Trexler1989).
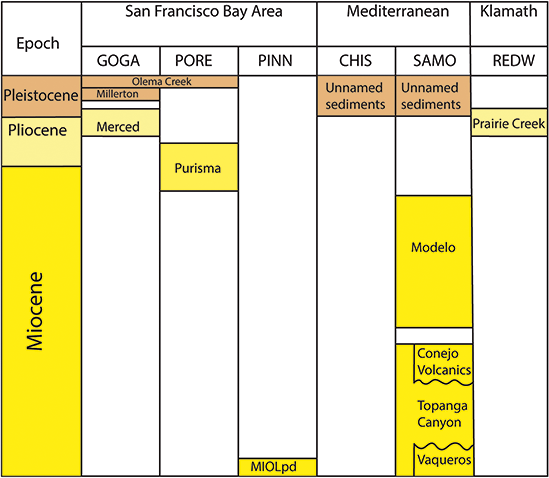
Figure 9 Neogene plant-fossil–bearing formations in the Klamath, Mediterranean Coast, and San Francisco Bay Area Inventory and Monitoring Networks.
3.2.4 San Francisco Bay Area Inventory and Monitoring Network
Point Reyes National Seashore
Point Reyes National Seashore (PORE) is situated along the coast north of San Francisco, California, and was established to preserve about 70,000 acres of beaches, sea cliffs, coastal dunes, grasslands, and Douglas fir forests (Elder et al., Reference Elder, Nyborg, Kenworthy and Santucci2008). Plant fossils have been noted from several formations exposed within PORE, including the Merced, Millerton, and Olema Creek formations that have also yielded plant fossils in the Golden Gate National Recreation Area (see below), but few of these occurrences have been described in detail.
Point Reyes Conglomerate: The Eocene Point Reyes Conglomerate is exposed on the western portion of the Point Reyes Peninsula and is the oldest geologic unit exposed within the boundaries of the national seashore. The conglomerate contains carbonized plant remains and seeds in silty micaceous sand horizons (Galloway, Reference Galloway1977).
Purisma Formation: The Upper Miocene–Pliocene Purisma Formation (ca. 6.7–4 Ma) is the uppermost unit in a sequence of three formations, which as a whole grade from glauconite-bearing greensands upward to shales and white siltstone and mudstones (Galloway, Reference Galloway1977; Elder et al., Reference Elder, Nyborg, Kenworthy and Santucci2008). Axelrod (Reference Axelrod, Lipps and Moores1971) described pinecones collected from strata of the Purisma Formation exposed along Drakes Bay within PORE.
Olema Creek Formation: The Pleistocene Olema Creek Formation is composed of interbedded estuarine and alluvial sediments deposited on a coastal plain, and its section commonly includes fine-grained overbank and backswamp deposits with abundant carbonaceous material (Grove et al., Reference Grove, Colson, Binkin., Dull and Garrison1995; Figure 9). The backswamp deposits are interpreted as such based on the occurrence of peat beds that suggest the presence of standing water. The peat beds preserve plant macrofossils and pollen. The macrofossil assemblage includes unidentified leaves, roots, branches, and cones, and palynomorphs include aquatic and riparian taxa such as Potamogeton, Cyperaceae, Salix, and Alnus (Grove et al., Reference Grove, Colson, Binkin., Dull and Garrison1995). Thermoluminescence dating constrains the age of the Olema Creek Formation deposits of Tomales Bay to about 130,000 ybp (Grove et al., Reference Grove, Colson, Binkin., Dull and Garrison1995).
Golden Gate National Recreation Area
The Golden Gate National Recreation Area (GOGA) encompasses about 80,000 acres in the vicinity of San Francisco, California, with exceptional exposures of late Cenozoic marine and terrestrial rocks that include three paleobotanically productive formations (Henkel et al., Reference Henkel, Elder, Santucci and Clites2015; Figure 9).
Merced Formation: The Merced Formation includes terrestrial and marine deposits from the late Pliocene to Pleistocene. Fossil pinecones from sandstones of the Merced Formation exposed in eroding sea cliffs at Mussel Rock and Thornton Beach were first collected and studied in the early 1930s (Dorf, Reference Dorf1933; Mason, Reference Mason1934), and pinecones from Thornton Beach have been collected as recently as 2012 (Henkel et al., Reference Henkel, Elder, Santucci and Clites2015). Collections from a site (locality 159 in Dorf, Reference Dorf1933) 3 km (2 miles) north of Mussel Rock produced paratype specimens of Pseudotsuga sonomensis and Pinus masoni that appear to be closely related to extant Pseudotsuga taxifolia and Pinus muricata (Dorf, Reference Dorf1933). Strata unconformably underlying the Merced Formation from another site near Mussel Rock produced the type specimen of Pinus lawsoniana, implying a minimum Pliocene age for this species (Axelrod, Reference Axelrod and Philbrick1967). Routine monitoring of these fossiliferous and actively eroding sea cliffs should be a priority for paleontological resource management in GOGA.
Millerton Formation: The Millerton Formation comprises Upper Pleistocene marine and freshwater clays, silts, sands, gravels, and conglomerate (Henkel et al., Reference Henkel, Elder, Santucci and Clites2015). Within GOGA, the formation is exposed on the shore of Tomales Bay and has yielded a rich floral record from outcrops on the eastern side of the bay (Elder et al., Reference Elder, Nyborg, Kenworthy and Santucci2008). The floral assemblage collected from the Millerton Formation includes unaltered or partially carbonized wood, seeds, fruits, and flowers that represent more than fifty species (Weaver, Reference Weaver1949; Johnson, Reference Johnson1962), and at least two dozen species characteristic of modern closed-canopy pine forests (Axelrod, Reference Axelrod, Lipps and Moores1971). Richards and Thurber (Reference Richards and Thurber1966) constrained the age of the Millerton Formation on the east coast of Tomales Bay to older than 50,000 ybp based on carbon-14 and thorium-230–uranium-234 dating of mollusk shells.
Olema Creek Formation: The Pleistocene Olema Creek Formation is stratigraphically above the Millerton Formation and preserves peat, woody debris, upright tree trunks, and aquatic and riparian plant material within GOGA (Galloway, Reference Galloway1977; Grove et al., Reference Grove, Colson, Binkin., Dull and Garrison1995; Henkel et al., Reference Henkel, Elder, Santucci and Clites2015). See the Point Reyes National Seashore section for further discussion on the paleobotany of this formation.
Pinnacles National Monument
Pinnacles National Monument, located between the San Benito and Salinas valleys in California, was established to protect its unique landscape formed by rock spires, crags, and pinnacles of volcanic breccia. Because the geology of the unit is predominantly composed of metamorphic and plutonic rocks, past and future collection of paleobotanical material is limited. The only paleobotanical material reported from the park to date are clasts in dacite breccia that resemble fossil wood. These clasts were found in an unnamed Oligocene–Miocene volcanic unit (MIOLpd of Barthelmes, Reference Barthelmes2022).
3.2.5 Mojave Desert Inventory and Monitoring Network
Death Valley National Park
Death Valley National Park (DEVA) occupies more than 3 million acres in eastern California and western Nevada and is the largest unit managed by the NPS in the contiguous USA. The park preserves a remarkably complete geologic record of two time periods: the Neoproterozoic to the middle Permian (ca. 790–270 Ma) and the Late Eocene (ca. 40 Ma) to present (Tweet et al., Reference Tweet, Santucci and Connors2016). The Cenozoic strata of DEVA are intermixed with igneous flows and volcanic ash falls and were mainly deposited in small, extensional basins that form the characteristic basin and range topography of the region today. Minor Cenozoic paleobotanical resources occur in the Upper Eocene–Lower Oligocene Titus Canyon Formation and the Upper Miocene to Pliocene Furnace Creek and Copper Canyon formations.
Titus Canyon Formation: The Upper Eocene–Lower Oligocene Titus Canyon Formation is exposed in the Grapevine and Funeral mountains in the eastern region of the park. Petrified wood and a partial pinecone identified as Pinus crossii have been recovered from Titus Canyon sediments within DEVA (Axelrod, Reference Axelrod1986). The age of these sediments is constrained to 34.7–30.4 Ma by radiometric dating of tuffs from the base of the unit and the overlying formation (Snow and Lux, Reference Snow and Lux1999; Lander et al., Reference Lander, Hemming, Nyborg and 108Miller2022).
Furnace Creek and Copper Canyon formations: The Furnace Creek and Copper Canyon formations comprise volcanic basalts and fluvial and lacustrine sediments deposited from the Late Miocene into the Pliocene (ca. 5.87–3.15 Ma). These strata preserve leaves of palms, Lyonothamnus (a genus in the Rosaceae now restricted to the Channel Islands), indeterminate grass-like plants, reed casts, charophytes, and a coprolite containing gymnosperm and angiosperm pollen (Spurr, Reference Spurr1903; Axelrod, Reference Axelrod and Death Valley1940; Nyborg and Santucci, Reference Nyborg and Santucci1999; Tweet et al., Reference Tweet, Santucci and Connors2016).
Grand Canyon–Parashant National Monument and Lake Mead National Recreation Area
Grand Canyon–Parashant National Monument (PARA) and Lake Mead National Recreation Area (LAKE) are geographically adjacent and share the same stratigraphy. The oldest National Recreation Area in the country, LAKE encompasses approximately 1.5 million acres in southeastern Nevada and northwestern Arizona. To protect a remote area of northwestern Arizona with exceptional Paleozoic paleontological resources, PARA was established in part from land formerly included within LAKE (Tweet et al., Reference Tweet, Flora, Weeks, McIntyre and Santucci2021). Minor occurrences of paleobotanical resources are known from each NPS unit and are derived from several different geologic formations.
Rainbow Gardens Formation: The Oligocene–Miocene Rainbow Gardens Formation (ca. 25–17 Ma) crops out extensively in the northern portion of LAKE and at Tassi Springs in PARA (Lamb et al., Reference Lamb, Beard and Dragos2018; Tweet et al., Reference Tweet, Flora, Weeks, McIntyre and Santucci2021). This formation is composed of fluvial siliciclastic and paludal and lacustrine carbonate rocks and preserves plant stems, root traces, and unclassified “tubular” plant casts within the parks (Longwell, Reference Longwell1936; Beard, Reference Beard1996; Howard et al., Reference Howard, Hook, Phelps and Block2003).
Other Reports: Lake Mead hosts some paleobotanical resources that are not present in PARA. These include a silicified palm stem recovered from unidentified Paleogene sediments, a floral assemblage from the Upper Miocene (ca. 12–6.2 Ma) Hualapai Limestone, and sub-fossil and fossil wood from the Plio–Pleistocene Bullhead Alluvium (Howard et al., Reference Howard, House, Dorsey and Pearthree2015; Bonde et al., Reference Bonde, Santucci, Tweet, Eichenberg and Moore2018). The latter unit was formerly considered a member of the Muddy Creek Formation but now constitutes its own formation (Bohannon, Reference Bohannon1984). Near the Temple Bar area of LAKE, the Hualapai Limestone has produced impressions of stems, roots, and leaves, some of which resemble grass or reed-like vegetation, and a possible Equisetum stem (Blair and Armstrong, Reference Blair and Armstrong1979).
Mojave National Preserve
Mojave National Preserve (MOJA), located in southern California about 50 km (30 mi) southeast of DEVA, was established to preserve approximately 1.6 million acres of the Mojave Desert. The Cenozoic strata exposed in MOJA are of Neogene age and comprise alluvial fan, eolian, fluvial, and lacustrine sediments interfingered with volcanic flows and eruptions (Tweet et al., Reference Tweet, Santucci and Connors2016; Johnson et al., Reference Johnson, Andeskie, Tweet and Santucci2023). Paleobotanical material has been recovered from a few sites within the park, most notably from lacustrine sediments of Wild Horse Mesa and Hackberry Wash that are informally named the Winkler formation (Reynolds et al., Reference Reynolds, Hunt and Albright1995). Sediments of the Winkler Formation are between two volcanic tuffs dated at 18.5 Ma and 17.7 Ma and preserve logs and wood of S. langsdorfii as well as other unidentified conifer wood and needles (Hazzard, Reference Hazzard1954; Reynolds et al., Reference Reynolds, Hunt and Albright1995; Santucci et al., Reference Santucci, Koch, Kenworthy, Santucci, Koch and Kenworthy2004). These floral remains are preserved alongside coeval ostracods, flamingo-like tracks, and a mammalian fauna that includes the rhinoceros Menoceras (Reynolds et al., Reference Reynolds, Hunt and Albright1995).
Other paleobotanical records from MOJA include reports of petrified “reeds” or reed casts from Hackberry Wash (Strong Reference Strong1975), palm wood and other unclassified wood from Hackberry Mountain (Strong, Reference Strong1975), unclassified petrified wood from the Providence Mountains, and Sequoia wood recorded from Barber Canyon and the mouth of Beecher Canyon (Reynolds et al., Reference Reynolds, Hunt and Albright1995; Tweet et al., Reference Tweet, Santucci and Connors2016). Improved documentation of the stratigraphic context of these occurrences of petrified wood will clarify whether they are derived from one or more geologic units and may aid in the protection of this resource.
3.2.6 Mediterranean Coast Inventory and Monitoring Network
Channel Islands National Park
Channel Islands National Park (CHIS) preserves the unique ecological, archeological, and geological resources of five of the eight Channel Islands that comprise the Channel Island Archipelago off the coast of southern California. The islands included in CHIS are the four northern islands – San Miguel, Santa Rosa, Santa Cruz, and Anacapa – as well as the northernmost island of the Southern Channel Islands, Santa Barbara. Major paleontological discoveries from CHIS are invertebrate faunas and remains of the pygmy mammoth (Mammuthus exilis), but the islands also preserve a wide range of paleobotanical resources. The resources include a possible Cercidiphyllum seed from the Eocene Cozy Dell Formation, root casts in the Pleistocene Potato Harbor Formation, a Pleistocene macroflora on Santa Cruz Island, seeds and caliche root and tree casts from Pleistocene deposits on San Miguel Island, and wood of Cupressus goveniana, Pinus and Prunus from Quaternary sediments on Santa Rosa Island (Tweet et al., Reference Tweet, Santucci and Connors2012b, Reference Tweet, Santucci, Convery, Hoffman and Kirn2020).
San Miguel Island Caliche Deposits: Pleistocene caliche (pedogenic calcium carbonate) deposits are present on several of the Channel Islands and are particularly well developed on San Miguel Island, where carbonate cement formed molds and casts on plant roots and trunks. These caliche deposits appear as pseudo-fossil forests and are thought to have formed during episodes in which windblown sand buried the standing vegetation. The original components of the standing vegetation are uncertain, but some workers have suggested that caliche concretions formed around roots of Rhus integrifolia and that the buried vegetation was shrubland (Cockerell, Reference Cockerell1938; Johnson, Reference Johnson1977). In addition to the calcified stumps and roots, plant macrofossils are also preserved as charcoalified stumps; cone and leaf impressions in calcareous tufa in late Quaternary sediments on San Miguel Island include Pinus and Ribes (Johnson, Reference Johnson1977).
Santa Cruz Island: A rich assemblage of seeds, cones, needles, leaf fragments, stem fragments, wood, and logs has been recovered from a clay, sand, and gravel deposit in the valley of Willow Creek in Cañada de los Sauces del Oeste on the southwestern part of the island (Chaney and Mason, Reference Chaney and Mason1930; Tweet et al., Reference Tweet, Santucci, Convery, Hoffman and Kirn2020). Radiocarbon dates of Douglas fir and charcoal found in this deposit constrain the age to 39,010–11,760 ybp (Fergusson and Libby, Reference Fergusson and Libby1963). The assemblage includes mosses, gymnosperms, and woody and herbaceous angiosperms. Mosses are represented by three species; gymnosperms are represented by wood, foliage, and cones of C. goveniana, Pseudotsuga menziesii, and two species of Pinus; angiosperms include seeds of the genera Arctostaphylos, Arceuthobium, Ceanothus, Cornus, Garrya, and Myrica (Chaney and Mason, Reference Chaney and Mason1930; Johnson, Reference Johnson1977).
Santa Monica Mountains National Recreation Area
Santa Monica Mountains National Recreation Area (SAMO) protects ca. 150,000 acres of the Santa Monica Mountain range on the coast of southern California. The recreation area borders Los Angeles to the southeast and is the largest urban national park in the USA. The coast, canyons, and mountains of the park contain a Mediterranean-climate ecosystem (chapparal) and extensive archeological and paleontological remains. The Neogene fossiliferous rocks of SAMO (Figure 6) were deposited in coastal floodplains and nearshore marine and submarine environments, and the abundance and quality of paleobotanical material preserved in these rocks varies considerably.
Unidentified fragments, wood, and root traces: Several formations within SAMO contain paleobotanical resources that are too fragmentary, poorly preserved, or biologically uninformative to be placed in a detailed ecological or evolutionary context. The Paleocene Simi Conglomerate comprises sandstones and conglomerate deposited in fluvial, nearshore marine, and submarine environments and is known to contain plant fragments at Runyon and Solstice canyons (Colburn et al., Reference Colburn, Jakobsen, Novak, Filewicz and Squires1988) as well as fossil wood (Parker, Reference Parker, Squires and Filewicz1983). The Sespe Formation is a predominantly terrestrial unit that was affected by a marine transgression toward the end of its deposition (Tweet et al., Reference Tweet, Santucci and Connors2012b). The formation includes root casts in its Piuma Member (Lander, Reference Lander2011) and mammals that range in age from Uintan (46.2–42.0 Ma) to Hemingfordian (20.6–16.3 Ma). The Middle Miocene Calabasas Formation, also known as the Upper Topanga Formation, is an offshore marine unit that contains locally abundant plant casts in the northern Santa Monica Mountains (Yerkes and Campbell, Reference Yerkes and Campbell1979).
Vaqueros Formation: The Vaqueros Formation represents sediments deposited in deltaic and marine environments during a period of sea level rise in the Late Oligocene. The unit unconformably overlies the terrestrial Sespe Formation in its western exposures and interfingers with fluvial deposits of the Sespe Formation in its eastern exposures. Fossil palm fronds (Sabalites), fossil log compressions, mollusks, and cartilaginous fish tooth fragments are preserved in a road cut along Piuma Road within SAMO that exposes gray marine sandstone of the Carbon Canyon Tongue of the Vaqueros Formation, which occurs between the lower and upper Piuma Members of the Sespe Formation. The age of these materials is constrained by 40Ar/39Ar dates of tuff in the upper strata of the lower Piuma Member (27.2 ± 0.2 Ma) and the lower strata of the upper Piuma Member (21.1 ± 0.2 Ma) (Lander Reference Lander2011).
Topanga Canyon Formation: The Topanga Canyon Formation is divided into four members that span the Early to Middle Miocene (ca. 23.0–15.5 Ma), and collectively their sediments are interpreted to represent two episodes of transgression separated by a prograding delta (Tweet et al., Reference Tweet, Santucci and Connors2012b). Within SAMO, paleobotanical material has been reported from the formation’s basal, deep-marine Encinal Member, and from the Fernwood Member, a terrestrial unit that lies between the nearshore Saddle Peak and Cold Creek members. The Encinal Member contains plant and wood fragments (Flack, Reference Flack1993), and the Fernwood Member contains leaves of Aralia and petrified wood of Platanoxylon (Lander, Reference Lander2011).
Conejo Volcanics: The Conejo Volcanics are comprised of a sequence of pillow lavas, pillow breccias, laharic breccias, and breccia conglomerates that represent a transition between submarine and subaerial volcanic deposition (Stadum and Wiegand, Reference Stadum and Weigand1999). Sparse vascular plant fossils have been recovered from lower strata of this unit exposed within SAMO (Stanton and Alderson, Reference Stanton and Alderson2013), and a notable assemblage of fossil wood was discovered in a tuff breccia in the unit’s upper strata exposed near Westlake Village, adjacent to SAMO (Stadum and Wiegand, Reference Stadum and Weigand1999). The assemblage includes several well-preserved specimens, and analysis of thin sections revealed the presence of the conifers P. menziesii, Calocedrus decurrens, and Pinus ponderosa, as well as angiosperm wood of the Fagaceae, Rhamnaceae, and Rosaceae (Stadum and Wiegand, Reference Stadum and Weigand1999).
Modelo Formation: The Modelo Formation is composed of shale, sandstone, conglomeratic sandstone, and volcanic ash deposited in a marine environment during the Middle to Late Miocene (Tweet et al., Reference Tweet, Santucci and Connors2012b). The formation preserves abundant foraminifera that constrain its age to ca. 15–5.2 Ma (Clark, Reference Clark, Squires and Filewicz1983; Rumelhart and Ingersoll, Reference Rumelhart and Ingersoll1997). Paleobotanical resources of the Modelo Formation within SAMO includes leaves of Platanaus dissecta recovered from a foraminiferal platy shale layer in the unit’s lower strata (Hoots, Reference Hoots1931), small pieces of petrified wood (Soper, Reference Soper1938), and algae from Sepulveda Canyon (Parker and Dawson, Reference Parker and Dawson1965). The algae collection includes holotype specimens of brown and red algae (Tweet et al., Reference Tweet, Santucci and Connors2012b; Supplementary Table 7).
Unnamed Pleistocene sediments: Pleistocene pinecones of Pinus radiata and Pinus remorata and an Arctostaphylos seed have been recovered from the mouth of Potrero Canyon in SAMO (Axelrod, Reference Axelrod and Philbrick1967, Reference Axelrod1990). Axelrod (Reference Axelrod and Philbrick1967) included the cones found in Potrero Canyon Park in a study that traced the evolutionary history of modern Californian closed-cone pines back to the Miocene. Today, P. radiata persists only as five disjunct populations on the California coast and has become critically endangered due to habitat fragmentation, land conversion, and native pathogens (Millar, Reference Millar1998).
3.2.7 Pacific Island Inventory and Monitoring Network
Puʻuhonua o Hōnaunau National Historical Park
Puʻuhonua o Hōnaunau National Historical Park (PUHO) was established to preserve 182 acres on the shoreline of the Mauna Loa volcano, on the western coast of the island of Hawai’i, which contain several significant archeological sites. Lava flows in PUHO have produced tree molds similar to those preserved in Hawai’i Volcanoes National Park. Nearly three dozen molds have been identified and are dated to about 1,000 ybp (Woodcock and Kalodimos, Reference Woodcock and Kalodimos2003, Reference Woodcock and Kalodimos2005). Although detail on many of these molds was lost due to “squeeze-up” formations, in which lava breached the mold from below and infilled portions of the hollow molds, they appear to represent Prichardia, a native palm that still grows in the park today (Hunt et al., Reference Hunt, Santucci and Kenworthy2007).
Hawai’i Volcanoes National Park
Hawai’i Volcanoes National Park, on the island of Hawai’i, includes the peaks of Kīlauea and Mauna Loa, two of the world’s most active volcanoes. In 1962, an eruption of lava through the floor of Aloi Crater temporarily filled the crater with a lava lake that reached depths of up to 50 feet and engulfed a stand of Metrosideros collina var. polymorpha growing on the crater floor. A subsequent eruption lower down on the mountain caused the lava lake to recede into a vent, leaving tree molds and casts in lava pillars, alongside the remains of trees that had not completely burned. The casts often preserve features such as leaf attachment and bark imprints (Hamilton, Reference Hamilton1964; Hunt et al., Reference Hunt, Santucci and Kenworthy2007).
3.3 Intermountain Region
The Intermountain Region includes 62 NPS units in Arizona, Colorado, Montana, New Mexico, Oklahoma, Texas, Utah, and Wyoming. This region is further divided into seven inventory and monitoring networks (Figure 10): the Chihuahuan Desert Network, Greater Yellowstone Network, the Northern Colorado Plateau Network, the Rocky Mountain Network, the Southern Colorado Plateau Network, the Sonoran Desert Network, and the Southern Plains Network. Paleobotanical material has been discovered in 15 NPS units that fall within this region and along two National Heritage Trails that pass through the region.
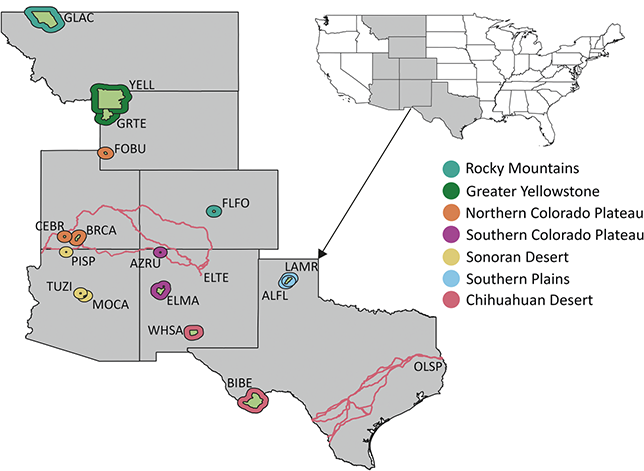
Figure 10 Map of NPS units with Cenozoic paleobotanical resources in the Intermountain Region, colored by inventory and monitoring network. See Figure 1 legend for details on the park symbology and acronyms.
The Intermountain Region is particularly rich in Eocene paleobotanical resources, which are conserved in eight NPS units (Figures 11 and 12). Among these resources are the abundantly fossiliferous shales of Florissant Fossil Beds National Monument and Fossil Butte National Monument that preserve some of the best-known Eocene plant assemblages worldwide, and the iconic fossil forests of Yellowstone National Park that document an interesting mode of fossil preservation. The long history of collection and study of fossil plants from Florissant and Fossil Butte has enabled detailed reconstruction of their fossil species (e.g., Manchester and Crane, Reference Manchester and Crane1987; McMurran and Manchester, Reference McMurran and Manchester2010), ecological communities (e.g., Meyer, Reference Meyer2003; Grande, Reference Grande2013), plant–insect interactions (e.g., Smith, Reference Smith2008), and surrounding landscape (e.g., Meyer, Reference Meyer1986). Importantly, these floras also serve as reference points for the identification of fossil taxa and interpretation of fossil plant communities preserved in other Paleogene sediments of the western USA and feature prominently in some syntheses of the vegetational history of North America (e.g., Wing, Reference Wing1987, Reference Wing, Janis, Scott and Jacobs1998; Graham Reference Graham1999; DeVore and Pigg, Reference DeVore and Pigg2010; Pigg and DeVore, Reference Pigg and DeVore2010). The fossil plants of Yellowstone also have a long history of collection and similar potential to serve as a key reference point in the Eocene history of the Rocky Mountain Region but are not nearly as well-studied as the assemblages from Florissant or Fossil Butte (Lesquereux, Reference Lesquereux and Hayden1872; Knowlton, Reference Knowlton1899; Dorf, Reference Dorf1964). The flora of the Lamar River and Sepulcher Formations dates from the Early Eocene Climatic Optimum and is coeval with several other Eocene floras of Wyoming (Wing, Reference Wing1987) but has not been revised since its initial description (Knowlton, Reference Knowlton1899; Supplementary Table 8).

Figure 11 Stratigraphic chart of Paleogene plant-fossil–bearing formations in the Intermountain Region.
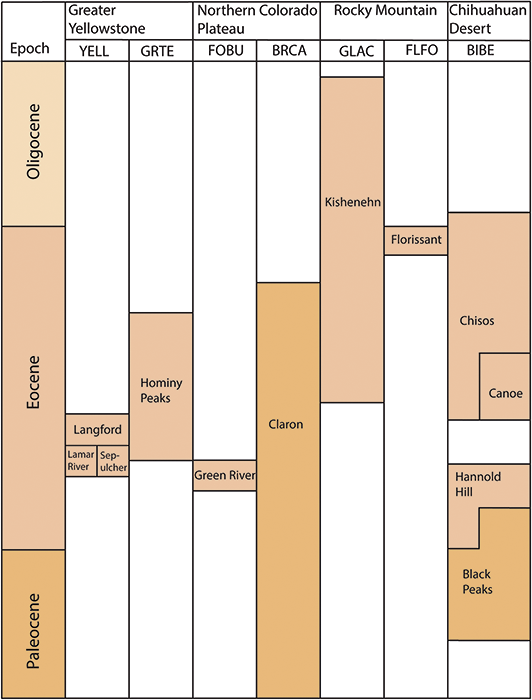
Figure 12 Summary of the temporal distribution and significance of Cenozoic paleobotanical resources in the Intermountain Region. See Figure 2 legend for explanation of significance ratings.
3.3.1 Rocky Mountain Inventory and Monitoring Network
Glacier National Park
Glacier National Park (GLAC) encompasses over a million acres in far northern Montana and shares its northern boundary with the US–Canada border. The Kishenehn basin, a structural and depositional basin spanning approximately 150 km (93 mi) along the Flathead Fault between northern Montana and southern British Columbia, is a prominent geologic feature within GLAC and preserves diverse floral and faunal remains in the Middle Eocene Kishenehn Formation.
Kishenehn Formation: Fossiliferous exposures of the Middle Eocene Kishenehn Formation are known from two regions of the Kishenehn basin: the North Fork of the Flathead River Region in southern British Columbia, Canada, and the Middle Fork of the Flathead River in northern Montana. In the Middle Fork Region, which includes the southern boundary of GLAC, the Kishenehn Formation is divided into the lower Coal Creek Member and the upper Pinchot Conglomerate. The Coal Creek Member is estimated to be 46.2 ± 0.4 Ma based on 40Ar/39Ar analysis and preserves a diverse flora of more than 108 plant taxa (Smith et al., Reference Smith, Greenwalt and Manchester2023) alongside a fauna that includes insects (16 orders and dozens of families; Greenwalt et al., Reference Greenwalt, Rose and Siljestrom2014), mammals (26 species; Dawson and Constenius, Reference Dawson and Constenius2018), mollusks (76 species; Pierce and Constenius, Reference Pierce and Constenius2001, Reference Pierce and Constenius2014), and fish (Constenius et al., Reference Constenius, Dawson, Pierce, Walter, Wilson, French and Grabb1989).
Plant fossils of the Kishenehn Formation are found as compressions and impressions of leaves, cones, fruits, and seeds in lacustrine oil shales and siltstones. According to Smith et al. (Reference Smith, Greenwalt and Manchester2023), the cones, fruits, and seeds include representatives of at least 14 families: Cupressaceae, Pinaceae, Betulaceae, Brassicaceae, Cercidiphyllaceae, Eucommiaceae, Juglandaceae, Oleaceae, Platanaceae, Rutaceae, Salicaceae, Sapindaceae, Simaroubaceae, and Ulmaceae (Figure 13). Vegetative material includes stems of Equisetum and at least 27 types of leaves. The most common of these leaf types are Alnus, Macginitiea wyomingensis, Trochodendroides, and Metasequoia. The flora also includes several aquatic plants: Lemna, Wolffiella, Potamogeton, and Ceratophyllum.

Figure 13 Fossil plants from the Intermountain Region. All scale bars = 1 cm. (a–c) Fossil leaves and reproductive structures collected from Big Bend National Park. (d) Leaf of Diospyros lamarensis collected from the banks of the Lamar River in Yellowstone National Park. (e) Holotype specimen of Devallia montana collected from Amethyst Mountain in Yellowstone National Park. (f) Paleobowdichia lamarensis collected from Yellowstone National Park. (g) Unidentified angiosperm leaves collected from the Claron Formation in Bryce Canyon National Park. (h) Fagopsis longifolia from Florissant Fossil Beds. (i) Plagiopodopsis cockerelliae from Florissant Fossil Beds. (j) Winged seed of Carpolithes lunatus, an extinct member of the Cercidiphyllaceae, recovered from the Kishenehn Formation in Glacier National Park. (k) Fruit of Thlaspi, an extant genus in the Brassicaceae, recovered from Glacier National Park.
The Kishenehn flora is significant for its geographic placement relative to other Eocene floras, its rich diversity of vegetative and reproductive material, and its cooccurrence with a diverse faunal assemblage. The sites within GLAC are the northernmost Middle Eocene paleobotanical localities in the US Rocky Mountain Region, and highly diverse. The flora contains many taxa that today grow naturally only in Asia (e.g., Ailanthus, Koelreuteria, Boniodendron, Dipteronia, Engelhardia [Palaeocarya], and Cercidiphyllaceae). The flora also contains fossils that represent the earliest known occurrences of the genera Lemna, Thalictrum, Myriophyllum, Thlaspi, and Chenopodium, and the first reported fossil occurrences of Wolffiella and Agrimonia. Many taxa have unclear affinities to any particular plant family, including two new genera and species established by Smith et al. (Reference Smith, Greenwalt and Manchester2023). Furthermore, there are paleobotanically productive horizons of the Kishenehn Formation within GLAC that are yet to be explored. In addition to the highly fossiliferous shales, Kishenehn sandstones are known to contain at least one taxon, Macginitiea.
Florissant Fossil Beds National Monument
The paleobotanical resources now protected at Florissant Fossil National Monument (FLFO) were first discovered by European scientists in the early 1870s, long before it was authorized as a National Monument in 1969. The monument was created in opposition to housing development plans (Leopold and Meyer, Reference Leopold and Meyer2012), with the intent to continuously monitor the fossil resources, develop interpretive programs, and aid scientists in the study and explication of its natural history. Consequently, the fossil resources of FLFO are among the best known and most secure of any NPS unit, as can be evidenced by the Florissant fossil database (https://florissantdatabase.colorado.edu/) and the mountain of scientific journal articles and popular scientific literature pertaining to the Florissant Lake Beds that have been published since its incorporation (Meyer et al., Reference Meyer, Wasson and Frakes2008).
Florissant Formation: All plant macrofossils recovered from FLFO come from the Florissant Formation. The Florissant Formation contacts the Wall Mountain Tuff at its base and lava flows of the Thirtynine Mile Volcanic field at the top (Evanoff et al., Reference Evanoff, McIntosh and Murphey2001). This formation comprises shales, mudstones, and conglomerate that were deposited in the Florissant paleovalley. The valley was episodically dammed by volcaniclastic debris, giving rise to the formation’s highly fossiliferous lacustrine deposits known as the Florissant Lake Beds (Evanoff et al., Reference Evanoff, McIntosh and Murphey2001). Outcrops of these ancient lakebeds extend over nearly 9,600 acres in central Colorado at elevations ranging from 2,500 to 2,700 m (8,200–9,000 ft) (McLeroy and Anderson, Reference McLeroy and Anderson1966). The National Monument consists of approximately 5,998 acres, protecting the southern arm of the Florissant Lake Beds.
Radiometric dates of sanidine crystals recovered from within the Florissant Formation and the underlying Wall Mountain Tuff and biostratigraphic age estimates indicate that the formation is latest Eocene in age (MacGinitie, Reference MacGinitie1953; Epis and Chapin, Reference Epis and Chapin1975; Evanoff et al., Reference Evanoff, McIntosh and Murphey2001). Samples from the three highest stratigraphic levels within the Florissant Formation produced a mean sanidine 40Ar/39Ar single-crystal age of 34.07 ± 0.10 Ma, and samples taken from the underlying Wall Mountain Tuff produced an age estimate of 36.07 Ma (Evanoff et al., Reference Evanoff, McIntosh and Murphey2001).
Plant fossils are preserved as impressions in each of the shale units, and in situ fossil stumps are present at the base of the lower mudstone. These specimens are recorded in the FLFO database and were recovered from more than 50 different localities. Unfortunately, the great majority of specimens are only attributed to “Florissant” and not any particular quarry. The lack of precise field data for most specimens hinders comparative studies among localities of the lake beds and determination of which fossil specimens were initially collected within park boundaries. MacGinitie (Reference MacGinitie1953) mapped three historical localities – the Denver Museum of Natural History locality (collected in 1915 or 1916), the “Princeton” locality (collected in 1880), and Scudder’s initial locality (collected in 1879) – as well as the localities University of Colorado 3731–3733, and tabulated abundances of specimens he recovered from four different quarries; he collected these specimens while conducting a revision of the Florissant flora (table 2 in MacGinitie, Reference MacGinitie1953). Each of these localities, except for the Denver Natural History Museum locality, appear to fall within FLFO.
The plant macrofossils of Florissant have been studied by numerous researchers over the past 150 years (e.g., Lesquereux, Reference Lesquereux1878; Cockerell, Reference Cockerell1908; Knowlton, Reference Knowlton1916; MacGinitie, Reference MacGinitie1953; Manchester, Reference Manchester2001; see Meyer, Reference Meyer2003, for an overview). The first comprehensive revision of the fossil flora was published by MacGinitie (Reference MacGinitie1953). He recognized 115 species of bryophytes, horsetails, ferns, gymnosperms, and angiosperms in 84 genera and 44 families, and an additional 29 taxa incertae sedis, which included extinct species of uncertain taxonomic affinity and specimens too poorly preserved for accurate taxonomic assessment (MacGinitie, Reference MacGinitie1953). In an update to MacGinitie’s monograph, which includes revisions of previously published species as well as 18 recently discovered taxa, Manchester (Reference Manchester2001) reported 84 genera and 96 species of leaves, 70 genera and 73 species of reproductive structures (flowers, fruits, seeds), and five genera and six species of fossil wood. Thus, the flora contains as many as 169 species if it is assumed that there is no taxonomic overlap between leaves, wood, and reproductive structures assigned to the same genus but not yet found in organic connection. This approximation is roughly equal to the number of pollen types recognized in the flora (Leopold and Clay-Poole, Reference Leopold and Clay-Poole2001; Wingate and Nichols, Reference Wingate and Nichols2001; Bouchal et al., Reference Bouchal, Zetter and Denk2016). Manchester (Reference Manchester2001) also prepared a pared-down list of reliably identified seed plants that includes 46 genera in 28 families of angiosperms and six genera in three families of gymnosperms. Many of the most spectacular specimens are illustrated in full color in the book Fossils of Florissant (Meyer, Reference Meyer2003). A database with color images of type specimens of Florissant fossils acquired by Herb Meyer is accessible online (https://florissantdatabase.colorado.edu).
Among angiosperms, the families with the greatest megafossil diversity at Florissant are the Fabaceae (3–9 genera, 5–11 spp.), Fagaceae (2 genera, 10 spp.), Rosaceae (9 genera, 11 spp.), Sapindaceae (3–8 genera, 7–11 spp.), and Ulmaceae (3 genera, 5 spp.). Fabaceae, Ulmaceae, and Sapindaceae are the only families to be represented by fossil leaves, wood, and reproductive structures (MacGinitie, Reference MacGinitie1953; Manchester, Reference Manchester2001; Wheeler, Reference Wheeler2001). The most diverse genera are Quercus (9 spp.), Rhus (3 spp.), Acer (3 spp.), Crataegus (3 spp.), and Salix (3 spp.). The most abundantly represented in terms of specimens are the extinct genera Cedrelospermum (formerly assigned to Zelkova by MacGinitie, Reference MacGinitie1953) and Fagopsis, an extinct oak relative. Although not as abundant or diverse as the dicotyledonous component of the flora, several monocotyledonous taxa have been described from Florissant based on leaves, stems, and reproductive material (Manchester, Reference Manchester2001). The extant monocot genera Potamogeton, Stipa, and Dioscorea have been reliably identified, and Typha and Smilax have putative records (MacGinitie, Reference MacGinitie1953; Manchester, Reference Manchester2001). A single fan-palm leaf specimen was recovered (pl. 3, figure 1 in Manchester, Reference Manchester2001) but lacks sufficient detail to assign to a particular extant genus and was therefore assigned to the fossil genus Palmacites.
Among gymnosperms, the most diverse families are Pinaceae and Cupressaceae. Pinaceae are represented by seeds, cones, three-needled fascicles, and five-needled fascicles attributed to the extant genera Abies, Picea, and Pinus; Cupressaceae are represented by wood, seed cones, pollen cones, and branchlets assigned to the extant genera Chamaecyparis and Sequoia (MacGinitie, Reference MacGinitie1953; Manchester, Reference Manchester2001; Meyer et al., Reference Meyer, Wasson and Frakes2008).
Non-seed plants also form a minor component of the flora. All fern taxa historically described from Florissant are now treated as a single species, Dryopteris guyottii (Lesquereux, Reference Lesquereux1883; Cockerell, Reference Cockerell1908; MacGinitie, Reference MacGinitie1953). Additional records of non-seed plants include the horsetail Equisetum florissantense and two species of Plagiopodopsis (Figure 13), an extinct genus in the bryophyte family Grimmiaceae (Cockerell, Reference Cockerell1915; Britton and Hollick, Reference Britton and Hollick1907, Reference Britton and Hollick1915; Steere, Reference Steere1947; MacGinitie, Reference MacGinitie1953).
In addition to the megafossil leaves, flowers, and fruits preserved as impressions in the shale, the Florissant Formation is also famous for petrified wood, including the iconic huge stumps of Sequoia near the Park visitor center. Angiosperm wood is also present but occurs in smaller pieces, many of which have probably been lost to years of private collecting prior to establishment of the park. Several wood types have been described (Wheeler, Reference Wheeler2001; Wheeler and Meyer, Reference Wheeler and Meyer2012).
Detailed investigations of Florissant pollen and spores, both by light microscopy (Leopold and Clay-Poole, Reference Leopold and Clay-Poole2001; Wingate and Nichols, Reference Wingate and Nichols2001), and by combined light and electron microscopy (Bouchal et al., Reference Bouchal, Zetter and Denk2016), provide more evidence in support of taxa known from megafossils, plus information on the presence of additional taxa not known from megafossils, such as Cathaya (Pinaceae), Onagraceae, Tetracentron (Trochodendraceae), and Diospyros (Ebenaceae).
The recovery of well-preserved reproductive fossils and revision of material originally assigned to extant genera has resulted in the recognition of a greater number of extinct genera at Florissant and undermines the assumption that Paleogene fossil leaves can be placed unequivocally in extant genera (Manchester and Crane, Reference Manchester and Crane1983, Reference Manchester and Crane1987; Manchester, Reference Manchester1989a, Reference Manchester1992, Reference Manchester2001; Manchester and Donoghue, Reference Manchester and Donoghue1995; Wang and Manchester, Reference Wang and Manchester2000). Seven extinct genera of angiosperms have been described from the Florissant flora: Fagopsis (Fagaceae; Figure 13), Cedrelospermum (Ulmaceae), Asterocarpinus (Betulaceae), Diplodipelta (Caprifoliaceae), Florissantia (Malvaceae), Chaneya (aff. Simaroubaceae), and Palaeopotamogeton (aff. Potamogetonaceae).
3.3.2 Greater Yellowstone Inventory and Monitoring Network
Yellowstone National Park
Yellowstone National Park (YELL), incorporated in 1872 as the first National Park in the nation, encompasses 2.2 million acres in northwestern Wyoming. The geology of this area includes a remarkably complete and fossiliferous stratigraphic record, preserving plants, vertebrates, invertebrates, and trace fossils in dozens of geologic units that span 540 million years. Significant paleobotanical remains are preserved in three formations of the Absaroka Volcanic Supergroup, which encompasses all Eocene volcanic and volcaniclastic rocks in the vicinity of Yellowstone (Smedes and Prostka, Reference Smedes and Prostka1972), and in Quaternary lake sediments.
Lamar River Formation: The Lamar River Formation represents volcaniclastic Lower Eocene (ca. 51–49 Ma) sediments in the Lamar River Valley (Fritz, Reference Fritz1982). This unit contains two of Yellowstone’s best known fossil forests at Amethyst Mountain and Specimen Ridge (Knowlton, Reference Knowlton1899; Dorf, Reference Dorf1964; Fritz, Reference Fritz1982; Wheeler, Reference Wheeler2024), as well as numerous horizons that have yielded fossil leaves (Figure 13; Lesquereux, Reference Lesquereux and Hayden1872; Knowlton, Reference Knowlton1899; Dorf, Reference Dorf1964).
The fossil stumps and logs preserved at Specimen Ridge were initially interpreted as representing as many as 27 discrete fossil forests buried in situ by episodic deposition of volcanic debris (Dorf, Reference Dorf1964). Subsequently, observations of high-energy mudflows transporting stumps from high to low elevations in modern volcanic regions, stratigraphic cooccurrence of fossil taxa whose modern relatives have nonoverlapping climatic tolerances, and fossil woods with and without seasonal growth rings inspired extensive discussion of the extent of taphonomic mixing that occurred during deposition and preservation of the Specimen Ridge flora (Dorf, Reference Dorf1960, Reference Dorf1964; Fritz, Reference Fritz1980, Reference Fritz1982, Reference Fritz1986; Yuretich, Reference Yuretich1984; Wing, Reference Wing1987; Wiemann et al., Reference Wiemann, Manchester and Wheeler1999).
The material that buried these forests consists of conglomerates, breccias, and tuffaceous siltstones and sandstones; fossil wood is associated with the former two lithologies, as well as paleosols, and fossil leaves are found as compressions and impressions in the tuffaceous material (Knowlton, Reference Knowlton1899; Dorf, Reference Dorf1964; Fritz, Reference Fritz1982). The wood assemblage at Specimen Ridge is numerically dominated by the conifer families Cupressaceae and Pinaceae but also includes several angiosperm families, notably Anacardiaceae, Betulaceae, Cornaceae, Fagaceae, Juglandaceae, Lauraceae, and Ulmaceae (Knowlton, Reference Knowlton1899; Beyer, Reference Beyer1954; Wheeler et al., Reference Wheeler, Scott and Barghoorn1977, Reference Wheeler, Scott and Barghoorn1978; Wheeler, Reference Wheeler2024).
In contrast to the wood assemblages, angiosperms dominate the leaf floras described by Knowlton (Reference Knowlton1899), both numerically and in diversity. Knowlton described more than 139 species of fossil leaves from the leaf flora, though this is likely an overestimation of the number of species represented by the fossils. Two of the authors of this contribution (Huegele and Manchester) examined Knowlton’s original published diagrams and photographs of his original specimens housed at the Smithsonian National Museum of Natural History in Washington, DC, and observed that, although many taxa seem correctly identified, many others need revision or synonymy (see Supplementary Table 8: Preliminary Review of Leaf Taxa). The impression flora contains horsetails (Equisetum stems) and fern foliage (Figure 13) reviewed by Collinson (Reference Collinson2001, Reference Collinson2002), several conifers, and numerous angiosperm plant families, including all of the families represented by wood in addition to Fabaceae (Knowlton, Reference Knowlton1899; Dorf, Reference Dorf1964; Herendeen et al., Reference Herendeen, Cardoso, Herrera and Wing2022). It is also possible that Musaceae, Ebenaceae (Figure 13), Cercidiphyllaceae, Sabiaceae, Betulaceae, Cornaceae, Moraceae, Lauraceae, and Magnoliaceae are represented in the leaf flora, but detailed study will be needed to confirm their presence (Figure 13; Supplementary Table 8). Some potentially notable taxa among the leaf flora include tropical elements like Musaphyllum (foliage of Musaceae) and Artocarpus, whose presence, if confirmed, would imply warm and wet environmental conditions based on the current distributions of their living relatives. The need for systematic revision of this flora has been alluded to by previous authors (Andrews and Lenz, Reference Andrews and Lenz1946) and should be considered a resource management priority for paleontology in YELL.
The fossil forests of Amethyst Mountain were first described more than 100 years ago (Holmes, Reference Holmes1879; Platen, Reference Platen1908). As at Specimen Ridge, petrified wood is preserved in a sequence of vertically stacked layers that represent forests preserved in distinct depositional episodes (Wheeler et al., Reference Wheeler, Scott and Barghoorn1977, Reference Wheeler, Scott and Barghoorn1978; Fritz, Reference Fritz1980, Reference Fritz1982; Wheeler, Reference Wheeler2024). A detailed study of 64 stumps and logs from a single lithological unit in this sequence revealed that upright stumps, stumps that were upright but offset from vertical, and horizontal logs occurred together in a single layer, and this cooccurrence likely indicates a mixture of in situ and transported material (Fritz and Fisk, Reference Fritz and Fisk1978). Preliminary identifications of the wood from this layer provide similar results to detailed systematic studies from Specimen Ridge. Many of the same families are present, and angiosperm wood types represent a large portion of taxa but a small percentage of individuals (Wheeler et al., Reference Wheeler, Scott and Barghoorn1977, Reference Wheeler, Scott and Barghoorn1978; Fritz and Fisk, Reference Fritz and Fisk1978; Wheeler, Reference Wheeler2024). Exposures of the Lamar River Formation at Amethyst Mountain also preserve fossil leaves described by Lesquereux (Reference Lesquereux and Hayden1872) and Knowlton (Reference Knowlton1899).
Sepulcher Formation: The Sepulcher Formation is a lateral equivalent of the Lamar Ridge Formation exposed in the northwest region of the park and in the geographically adjacent Gallatin National Forest (Fritz, Reference Fritz1982). Fossil forests like those preserved in the Lamar River Formation at Specimen Ridge and Amethyst Mountain have been identified within YELL at Specimen Creek and Bighorn Peak (Andrews, Reference Andrews1939; Andrews and Lenz, Reference Andrews and Lenz1946; Wheeler et al., Reference Wheeler, Scott and Barghoorn1977, Reference Wheeler, Scott and Barghoorn1978; Chadwick and Yamamoto, Reference Chadwick and Yamamoto1984; Wheeler, Reference Wheeler2024). Nine wood-bearing stratigraphic levels were identified at Specimen Creek, and each preserves a mixture of upright stumps and horizontal logs (Chadwick and Yamamoto, Reference Chadwick and Yamamoto1984). Taxonomic groups present at Specimen Creek but not known from the wood floras of the Lamar River Formation include Ebenaceae, Hamamelidaceae, and Salicaceae (Chadwick and Yamamoto, Reference Chadwick and Yamamoto1984). The Sepulcher Formation also includes some of the source localities for the leaf and fruit fossils described by Knowlton (Reference Knowlton1899).
Langford Formation: The Middle Eocene (ca. 46–47 Ma) Langford Formation is extensively exposed in the Lamar River Valley, where it unconformably overlies the Lamar River Formation (Fritz, Reference Fritz1994; Santucci, Reference Santucci1998). A paleobotanically fossiliferous horizon of the Langford Formation was discovered in a road cut between Lake Butte and Sylvan Pass during construction of the East Entrance Road (Fritz, Reference Fritz1994; Santucci, Reference Santucci1998). This horizon produced many large leaves of an extinct genus of sycamore, Macginitiea (Fritz, Reference Fritz1994), which are now held in the Yellowstone Park collection.
Quaternary sediments: Yellowstone National Park includes several sections that preserve late Pleistocene (ca. 140,000 ybp to present day) palynomorphs and plant macrofossils (Baker, Reference Baker1976, Reference Baker1986; Baker and Richmond, Reference Baker and Richmond1978). The southeast arm of Yellowstone Lake has been particularly productive, yielding palynomorphs in lake cores and plant macrofossils in stream cuts along tributaries to Yellowstone Lake (Baker, Reference Baker1970, Reference Baker1986). The palynomorph assemblages are numerically dominated by Pinus and Picea, but also include other conifers, Abies and Pseudotsuga, and the dicot trees Alnus, Betula, Quercus, and Salix. Macrofossil assemblages record many of these same taxa, in addition to herbs such as Potentilla, Gaultheria, and Selaginella. The combined sequence of pollen and macrofossils throughout the Sangamonian and Wisconsin glacial–interglacial cycles suggests that vegetation transitioned from dense, coniferous forests to open, parkland, coniferous forests or tundra during cool periods (Baker, Reference Baker1986).
Grand Teton National Park
Grand Teton National Park (GRTE) preserves part of the Teton Range of northwestern Wyoming. Rocks exposed within the park range from Cambrian to Quaternary and contain minor Cenozoic paleobotanical material in the middle Eocene Hominy Peaks Formation and the Pliocene Teewinot Formation.
Hominy Peaks Formation: The Hominy Peaks Formation is made up of volcaniclastic rocks and encompasses all Eocene exposures within the Teton Range (Leopold et al., Reference Leopold, Liu, Love and Love2007). Exposures of the formation near the northwest corner of GRTE preserve fragments of carbonized plant remains, mats of contorted plant debris, and fossil pollen at several stratigraphic horizons. Palynomorphs from the Hominy Peaks Formation include eight types of ferns, eight types of gymnosperms, 26 types of eudicots, and three types of monocots. Biostratigraphic correlation of this assemblage was used to establish a Middle Eocene age, and its taxonomic composition resembles that of the Middle Eocene Kisinger Lakes flora of northwestern Wyoming (MacGinitie, Reference MacGinitie1974; Leopold et al., Reference Leopold, Liu, Love and Love2007). East of GRTE, fossil-leaf–bearing horizons have been identified in the underlying Pinyon Conglomerate at Whetstone Mountain and in unnamed Eocene rocks exposed at Togwotee Pass (Tweet et al., Reference Tweet, Santucci and Connors2013)
Neogene Resources: Minor occurrences of paleobotanical material have been reported from several Neogene units within GRTE. Pliocene palynomorphs have been recovered from the Shooting Iron and Teewinot formations (Leopold et al., Reference Leopold, Liu, Love and Love2007), and coniferous wood and a possible pinecone fragment were identified from the Teewinot Formation (Tweet et al., Reference Tweet, Santucci and Connors2013). In Quaternary sediments, Santucci and Wall (Reference Santucci and Wall1999) reported a piece of ca. 27 kya carbonized wood found in a laminated silt near Pilgrim Creek, and Holocene charcoal recovered from the area around Jackson Hole (including within GRTE) and palynomorphs recovered from Hedrick Pond in GRTE provide a record of recent fire and vegetation history (Pierce and Good, Reference Pierce and Good1992; Jacobs and Whitlock, Reference Jacobs and Whitlock2006, Reference Jacobs and Whitlock2008).
3.3.3 Northern Colorado Plateau Inventory and Monitoring Network
Fossil Butte National Monument
Fossil Butte National Monument (FOBU), just west of the town of Kemmerer in southwestern Wyoming, has attracted interest since the late nineteenth century because of the wealth of fossil fish and other vertebrate remains preserved in fissile lacustrine shales of the Green River Formation (Fossil Butte Member). Green River Formation fossils continue to be popular for sales to museums, interior decorators, and fossil enthusiasts. Although amateur collecting of these fossils is now prohibited on Bureau of Land Management and National Monument land, major discoveries continue to be made in privately owned or leased fish quarries outside of the monument. Fossil plant remains are recovered mainly as a byproduct of fish excavation, but the iconic large palm fronds are highly sought-after and expensive. Other plant remains are diverse, but well-preserved specimens are relatively rare.
Green River Formation: The prolific shales occur in the Fossil Butte Member of the Green River Formation and date to about 52 Ma (Grande, Reference Grande2013) based on radiometric analyses. This member is older than the Parachute Creek Member of the same formation, which is well known for fossil plants recovered from numerous sites in eastern Utah near Bonanza and western Colorado north of Rifle and in the area of Douglas Pass (MacGinitie, Reference MacGinitie1969).
The plant fossils recovered from FOBU are impressions in shale recovered by splitting with thin, wide chisels. The flora of Fossil Butte is too diverse to list exhaustively, encompassing more than 250 leaf types and 100 fruit types (Tweet et al., Reference Tweet, Santucci, Connors and Kenworthy2012c). Here we call attention to some of the notable remains collected from the monument. The leaves include aquatic plants such as Salvinia, Ceratophyllum, and Nelumbo), as well as a wide range of terrestrial plants. Distinctive winged fruits of Ailanthus are among the more common plant remains. Platycarya, a genus of the walnut family now confined to eastern Asia, is commonly found, represented by leaflets and catkins (Wing and Hickey, Reference Wing and Hickey1984). The extinct elm-related plant Cedrelospermum is represented both by leaves and winged fruits. However, many of the other constituents have not been satisfactorily identified and some probably represent as-yet-undescribed extinct genera. The extinct rabbit ear-winged fruit, Lagokarpos, is relatively common in these beds and is shared with other Eocene lacustrine strata in North America, Germany, and Tibet (McMurran and Manchester, Reference McMurran and Manchester2010; Tang et al., Reference Tang, Liu and Wu2019). A fortunate aspect of these shales is that they can be readily collected in large slabs or sheets, so there is a greater chance of recovering more complete and informative specimens. In rare instances, leaves and fruits are attached to the same twig, providing direct proof that these organs represent the same species of plant. Several plant taxa were described from this flora by Brown (Reference Brown1937), but the identities of some, such as those he attributed to Ficus, remain uncertain. The most comprehensive treatment of the flora, illustrating the best examples of many different morphotypes, was published in color by Lance Grande in his book the Lost World of Fossil Lake (Grande, Reference Grande2013).
Cedar Breaks National Monument
Cedar Breaks National Monument (CEBR) is located in southwest Utah, approximately 90 km (56 mi) west of Bryce Canyon National Park, and was established to preserve the Cedar Breaks amphitheater (Tweet et al., Reference Tweet, Santucci, Connors and Kenworthy2012c). The monument has similar geology to BRCA (see below) and also preserves endocarps belonging to either Celtis or Prunus in the Paleocene–Middle Eocene Claron Formation (Schneider, Reference Schneider1967). Other paleobotanical material recovered from CEBR includes pollen, conifer needles, and wood of Picea found in a core from Alpine Pond that spans the last 3,000 years (Anderson et al., Reference Anderson, Hasbargen, Koehler and Feiler1999). Other Quaternary sites close to the boundaries of the park preserve pollen, conifer needles, and other plant macrofossils from the Late Pleistocene (Mulvey et al., Reference Mulvey, Currey and Lindsay1984; Anderson et al., Reference Anderson, Hasbargen, Koehler and Feiler1999; Madsen et al., Reference Madsen, Sarna-Wojcicki and Thompson2002).
Bryce Canyon National Park
Bryce Canyon National Park (BRCA) preserves approximately 35,835 acres in southwest Utah and is known for its scenic geologic features, including pinnacles, spires, and horseshoe-shaped amphitheaters. Paleobotanical resources contained within BRCA are limited to some discoveries within the Paleocene–Middle Eocene Claron Formation (Tweet et al., Reference Tweet, Santucci, Connors and Kenworthy2012c). Seeds belonging to either Celtis or Prunus have been reported from the Claron Formation, and the source layer has been correlated with a fossiliferous layer of the Claron Formation in Cedar Breaks National Monument, roughly 90 km (about 56 mi) west of BRCA (Schneider, Reference Schneider1967). More recently, Celtis endocarps and leaf impressions (Figure 13) were collected and identified in 2013 (Tut Tran, NPS, pers. comm., 2023). Leaf impressions have also been reported (Gregory, Reference Gregory1950), but none were identified or collected. Other paleobotanical material from the Claron Formation of BRCA includes molds of angiosperm seeds and fossilized wood in the park collections (Tut Tran, NPS, pers. comm., 2023), and root traces preserved in the formation’s upper member (Taylor, Reference Taylor1993; Tweet et al., Reference Tweet, Santucci, Connors and Kenworthy2012c). Age constraints on the Claron Formation are provided by a Middle Eocene rodent jaw collected from its upper member, and radiometric dating of volcaniclastic strata in the overlying Brian Head Formation to 36 Ma (Biek et al., Reference Biek, Anderson, Rowley and Maldonado2011; Eaton et al., Reference Eaton, Tibert and Biek2011).
3.3.4 Southern Colorado Plateau Inventory and Monitoring Network
Aztec Ruins National Monument
Aztec Ruins National Monument (AZRU) preserves the ruins of a twelfth-century Pueblo Indian community in northwestern New Mexico. The bedrock of this unit is composed of the fossiliferous Nacimiento Formation, but the unit only occupies a small area (318 acres) with few geologic exposures (KellerLynn, Reference KellerLynn2016). Paleobotanical material found within AZRU is sparse and includes unidentified petrified wood, a possible leaf, and a possible seed held in the park collections. Petrified wood modified by humans is found in artifact assemblages and reworked petrified wood is found in Quaternary terrace deposits (Tweet et al., Reference Tweet, Santucci, Kenworthy and Mims2009; KellerLynn, Reference KellerLynn2016). The original source of this wood is unknown, but the Paleocene Nacimiento Formation, which contains petrified wood and fossil leaves outside of AZRU and is mapped nearby, is a likely candidate.
El Malpais National Monument
El Malpais National Monument (ELMA) in western New Mexico is a minor source of paleobotanical resources. Tree molds preserved in lava flows can be seen in the Bandera flows (11,000 ybp) at Ke Cave Trading Post (Laughlin and Perry, Reference Laughlin and Perry1997). Charcoal and “baked” roots from the Lava Crater (17,000 ybp), Bandera, and McCartys flows (3,000 ybp) have been radiocarbon dated (Laughlin et al., Reference Laughlin, Charles, Reid and White1993, Reference Laughlin, Poths, Healey, Reneau and Woldegabriel1994; Mabery, Reference Mabery1997). Quaternary-aged sub-fossil wood was also found and studied in ELMA for climate data (Grissino Mayer and Swetnam, Reference Grissino Mayer and Swetnam2000; Poore et al., Reference Poore, Pavich and Grissino-Mayer2005).
3.3.5 Sonoran Desert Inventory and Monitoring Network
Tuzigoot National Monument
Tuzigoot National Monument (TUZI) protects the remains of a pueblo built by the Sinagua people between about 1100 and 1450 and a surrounding area of about 800 acres. It is located about 30 km (19 mi) northwest of Montezuma Castle and has similar geology to that unit (see below). Undescribed plant fossils from TUZI have been found at a locality established by the Museum of Northern Arizona (MNA 330–0) in Pliocene strata of the Verde Formation (Tweet et al., Reference Tweet, Santucci and Kenworthy2008).
Montezuma Castle National Monument
Montezuma Castle National Monument in central Arizona preserves cliff dwellings built by the Sinagua people in the twelfth and thirteenth centuries and a large permanent spring, Montezuma Well. Cenozoic paleobotanical material has been found from a few Neogene and Quaternary sites in the Montezuma Well unit. The Miocene–Pliocene Verde Formation preserves plant fossils at a locality established by the Museum of Northern Arizona (MNA 295–0) near Montezuma Well and in the limestone walls of the well (Lange, Reference Lange1957; Tweet et al., Reference Tweet, Santucci and Kenworthy2008). The fossils in the well walls are reported as “reed-like” impressions and molds of plant debris (Lange, Reference Lange1957). Pleistocene plant impressions and palynofloras have been identified in travertine deposits dated to between 400,000 and 700,000 ybp nearby the well. The macrofloral impressions include leaves of trees and riparian shrubs and a pinecone similar to extant P. ponderosa (Hevly et al., Reference Hevly, Nations and Zabo1992). Cores taken from the permanent spring at the bottom of Montezuma Well have yielded pollen and plant macrofossils, which have been used to reconstruct the site’s Quaternary environmental history (Hevly, Reference Hevly1974; Davis and Shafer, Reference Davis and Shafer1992, Reference Davis and Shafer2006; Hevly et al., Reference Hevly, Nations and Zabo1992; KellerLynn, Reference KellerLynn2019).
3.3.6 Southern Plains Inventory and Monitoring Network
Alibates Flint Quarries National Monument and Lake Meredith National Recreation Area
Alibates Flint Quarries National Monument and Lake Meredith National Recreation Area (LAMR) in western Texas are adjoining regions managed jointly by the NPS that share the same geology. The most notable paleobotanical material associated with the two units comes from the Miocene–Pliocene Ogallala Formation. Wilson (Reference Wilson1988) reported root casts, silicified grass anthoecia, Celtis endocarps, and a Clarendonian (13.6–10.3 Ma) vertebrate assemblage from the informal “LX” member of the Ogallala Formation. However, it is unclear what specimens were found within which NPS unit (Hunt and Santucci, Reference Hunt and Santucci2001). Other paleobotanical resources include plant debris in an ash bed known as the Lava Creek ash (about 640 kya) in LAMR’s South Canyon, and likely reworked petrified wood alongside proboscidean remains in Pleistocene gravels (Hunt and Santucci, Reference Hunt and Santucci2001; Tweet et al., Reference Tweet, Santucci and Connors2015).
3.3.7 Chihuahuan Desert Inventory and Monitoring Network
Old Spanish National Historical Trail
The Old Spanish National Historic Trail (OLSP) protects a network of Indigenous trails, early trade and exploration routes, and horse and mule paths that span approximately 4,300 km (2,700 mi) and cross California, Nevada, Arizona, Utah, Colorado, and New Mexico. Petrified wood was found along OLSP in Cañon Largo, New Mexico, during the 1859 Macomb Expedition, a government-funded expedition with the goal of developing a route to transport military supplies between Santa Fe, New Mexico, and southern Utah (Madsen, Reference Madsen2010). The wood is unidentified but is likely derived from the Paleocene Nacimiento Formation or the Eocene San Jose Formation. The potential for recovery of Cenozoic paleobotanical material along much of OLSP is limited (the youngest bedrock predates the Cenozoic), with a few exceptions. The trail passes through or nearby several other NPS-managed areas known to contain Cenozoic paleobotanical resources, including MOJA, LAKE, and AZRU.
White Sands National Park
White Sands National Park (WHSA) in south-central New Mexico preserves one of the world’s largest gypsum and sand dune fields. Cenozoic plant macrofossil remains from WHSA are present in Quaternary sediments and include external molds of cacti and other plants that became encrusted with or replaced by gypsum crystals (Vandiver, Reference Vandiver1936; Santucci et al., Reference Santucci, Kenworthy and Visaggi2007), and layers of Ruppia cirrhosa (ditchgrass) endocarps (Bennett et al., Reference Bennett, Bustos and Pigati2021a). The ditchgrass endocarps are preserved at several stratigraphic horizons within a section that also includes palynomorphs and tracks of humans, proboscideans, and canids. Radiocarbon dating of the endocarps and palynomorphs constrain the age of seven stratigraphic horizons preserving human tracks to approximately 23–21 kya within the Last Glacial Maximum (Bennett et al., Reference Bennett, Bustos and Pigati2021a, Reference Bennett, Bustos and Pigati2021b; Pigati et al., Reference Pigati, Springer and Honke2023). These tracks provide a temporal extension of the coexistence of early human inhabitants of North America and Pleistocene megafauna.
Big Bend National Park
Big Bend National Park (BIBE) in southwestern Texas conserves over a million acres of Chihuahuan Desert and is very well known for its Cretaceous and Early Cenozoic assemblages of fossil plants and vertebrates. The Cenozoic paleobotanical resources of BIBE include petrified wood, leaves, and root traces that occur in four formations that span the Early Paleocene to Early Oligocene. Geographically, these resources are concentrated in the Tornillo Flats of the northeast-central section of the park where the Tornillo Group, defined by the Javelina, Black Peaks, and Hannold Hill formations, crops out extensively.
Black Peaks Formation: The Blacks Peaks Formation is stratigraphically positioned between the Javelina and Hannold Hill formations, and collectively these units constitute the Tornillo Group, a succession of Upper Cretaceous, Paleocene, and Lower Eocene fluvial strata that preserve abundant floral and faunal remains (Lehman et al., Reference Lehman, Wick, Beatty, Straight and Wagner2018). Two intervals in the Black Peaks Formation contain well-preserved petrified wood. The “log-jam” level is a thin but widespread stream-channel sandstone that preserves many thousands of logs and is dated to the Early Paleocene (Torrejonian; 63.3–60.2 Ma) based on mammal biostratigraphy (Wheeler and Lehman, Reference Wheeler and Lehman2009). The wood recovered from this horizon includes three angiosperm taxa, Paraphyllanthoxylon abbottii (affinity to Burseraceae), cf. Plataninium haydenii (affinity to Platanaceae), and cf. Cunonioxylon sensu Gottwald (affinity to Cunoniaceae), but most specimens (all but two) studied belong to the taxon P. abbottii (Wheeler, Reference Wheeler1991; Wheeler and Lehman, Reference Wheeler and Lehman2009). Two types of araucarioid wood (BP 8 and 9 of Wheeler and Lehman, Reference Wheeler and Lehman2005) have been identified from higher levels in the formation, above an erosional surface overlain by Eocene and younger strata. Exact locality information for these specimens is on file at the North Carolina Museum of Natural Sciences (Wheeler and Lehman, Reference Wheeler and Lehman2009).
Hannold Hill Formation: The overlying Hannold Hill Formation is exposed in the Tornillo Flats Region of BIBE and comprises mudstones, sandstones, and conglomerate deposited in an inland floodplain setting (Wick and Corrick, Reference Wick and Corrick2015). Some petrified wood is found as rounded or reworked clasts within a sandstone conglomerate (Lehman et al., Reference Lehman, Wick, Beatty, Straight and Wagner2018). Early Eocene (Wasatchian; 55.4–50.3 Ma) vertebrate assemblages have been recovered from sandstones in the upper part of the Black Peaks Formation and lower part of the Hannold Hill Formation (Lehman et al., Reference Lehman, Wick, Beatty, Straight and Wagner2018).
Canoe Formation: The Middle Eocene Canoe Formation unconformably overlies the Hannold Hill Formation and comprises conglomerate, sandstones, siltstones, and mudstones deposited in braided stream and floodplain settings (Wick and Corrick, Reference Wick and Corrick2015; Wick, Reference Wick2021). Paleobotanical resources of the Canoe Formation include leaves, reproductive structures, and transported, in situ, and reworked fossil wood (Figure 13). The Big Yellow Sandstone Member contains Eocene conifer and angiosperm wood, abraded fragments of wood believed to have been exhumed from underlying Cretaceous strata during Eocene entrenchment, and, in the McKinley Hills area of BIBE, a small forest of 92 tree stumps (Wick, Reference Wick2021). These stumps have been described as poorly preserved and the samples taken were not viable for analysis or identification, but the site is extensive and has potential for further study (Tom Lehman, pers. comm., 2023). Limonite-encrusted logs are also found within the park (Lehman et al., Reference Lehman, Wick, Beatty, Straight and Wagner2018). Fossil leaves and reproductive structures are preserved in siltstone and lignite layers originally attributed to the Javelina Formation (Lawson, Reference Lawson1972), but subsequent stratigraphic study indicates that the strata and fossils represent the Canoe Formation (Turner et al., Reference Turner, Berry and Page2011; Wick, Reference Wick2021). Based on the fossil leaf impressions, Lawson (Reference Lawson1972) identified six taxa: Ampelopis acerifolia, Ficus cf. F. tennesseensis, Laurus socialis, P. raynoldsi, Saccoloma gardneri, and Typha.
Chisos Formation: Overlying the Canoe Formation, the Chisos Formation shows evidence of past volcanism and includes igneous intrusions, lavas, ash-falls, and fluvial volcaniclastic sediments deposited between ca. 42 and 32 Ma (Wick and Corrick, Reference Wick and Corrick2015). The Chisos Formation has been reported to contain petrified wood, although occurrences are rare, and no specimens have been identified (Maxwell et al., Reference Maxwell, Lonsdale, Hazzard and Wilson1967). There is also a record of root traces being common in certain volcanic areas (Wilson and Runkel, Reference Wilson and Runkel1989).
El Camino Real de los Tejas National Historical Trail
El Camino Real de los Tejas National Historical Trail (ELTE) designates approximately 4,000 km (2,500 mi) of Indigenous trails and trade routes that connected Mexico City and Los Adaes (the capital of Spanish Texas prior to 1770), and were heavily used by Spanish colonists during their occupation of land that is today Texas and northwest Louisiana (Figure 10). Cenozoic paleobotanical resources are present along certain parts of the trail. Stone City Bluff is a fossil site on ELTE that has produced unidentified plant debris (Stenzel et al., Reference Stenzel, Krause and Twining1957; Hodgkinson, Reference Hodgkinson1974). A Quaternary-aged mammoth site on the trail also includes plant debris and pollen (Carpenter et al., Reference Carpenter, Bousman and Potapova2013). In some parts of Texas and Louisiana, the trail roughly follows sediments belonging to the Paleocene and Eocene Wilcox and Claiborne groups that have produced diverse fossil floras in Texas and Tennessee, respectively. Exposures of the Wilcox Group within a mile of the trail in this region have yielded abundant fossil leaves (Deussen, Reference Deussen1924). There is also potential to discover fossil wood from exposures of the Cenozoic Catahoula and Yegua formations located along the trail in south-central Texas.
3.4 Midwest Region
The Midwest Region includes 53 NPS units in Illinois, Indiana, Iowa, Kansas, Michigan, Minnesota, Missouri, Nebraska, North Dakota, Ohio, South Dakota, and Wisconsin (Figure 14). These units are divided into three inventory and monitoring networks: the Great Lakes Network, the Heartland Network, and the Northern Great Plains Network (NGPN). Collectively, they preserve paleobotanical resources in 11 different NPS units and 18 different formations that span the Paleocene to Holocene.
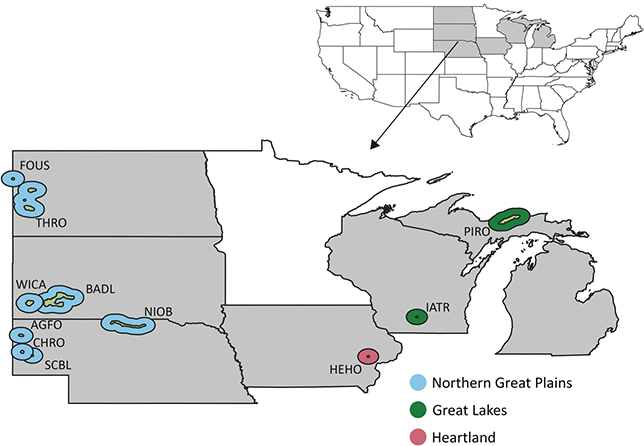
Figure 14 Map of NPS units with Cenozoic paleobotanical resources in the Midwest NPS, colored by inventory and monitoring network. See Figure 1 legend for details on the park symbology and acronyms.
The NPS units of the Midwest Region preserve significant paleobotanical resources in the Paleocene petrified forests of Theodore Roosevelt National Park and within Late Eocene–Late Miocene deposits of South Dakota and Nebraska units that are thought to document the early development of the North American grassland ecosystems that characterize the natural vegetation of much of the Midwest today (Figures 15 and 16; Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991; Strömberg and McInerney, Reference Strömberg and McInerney2011). The assemblage of in situ stumps, fossil leaves, and reproductive structures preserved in the Upper Paleocene Sentinel Butte Formation at the Petrified Forest Plateau of Theodore Roosevelt National Park offers insight into the floristic composition and paleoecology of mid-latitude Paleocene forests and a point of comparison with the well-documented localities of the Sentinel Butte Formation outside of the park (Crane et al., Reference Crane, Manchester and Dilcher1990; Manchester et al., Reference Manchester, Pigg and Crane2004) and with the Middle and Late Paleocene assemblages of the Fort Union Group in Wyoming and Montana (Wing et al., Reference Wing, Alroy and Hickey1995; Wilf, Reference Wilf2000; Peppe, Reference Peppe2010), among other North American Paleocene floras (see Wing, Reference Wing, Janis, Scott and Jacobs1998; Graham, Reference Graham1999; and Pigg and DeVore, Reference Pigg and DeVore2010 for reviews). Collectively these Paleocene floras document the forested ecosystems that emerged several million years after the K–Pg mass extinction and provide a regional context for interpreting episodes of pulsed taxonomic turnover at the Paleocene–Eocene boundary documented from individual basins (Wilf, Reference Wilf2000; Wing et al., Reference Wing, Harrington and Smith2005).
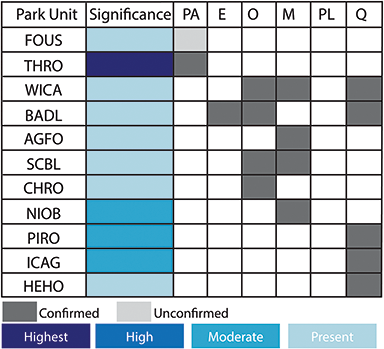
Figure 15 Summary of the temporal distribution and significance of Cenozoic paleobotanical resources in the Midwest Region. See Figure 2 legend for explanation of significance ratings
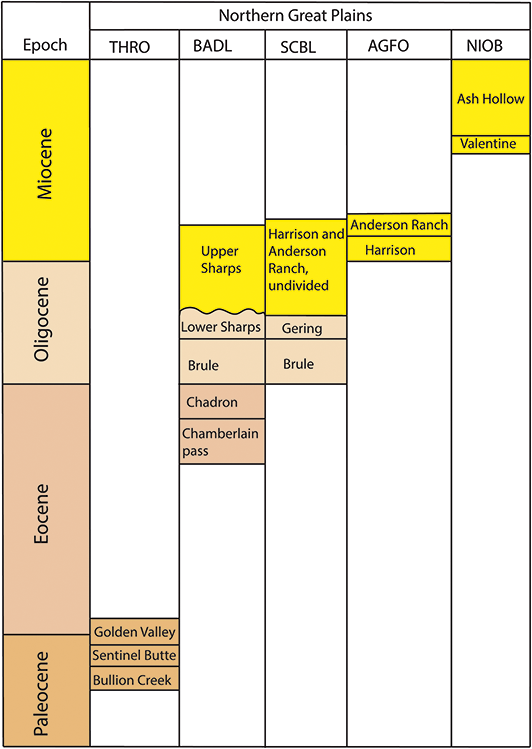
Figure 16 Stratigraphic chart of Tertiary plant-fossil–bearing formations in the Midwest Region.
The history of North American grasslands has been inferred using four lines of (paleo)botanical evidence: macrofossils, microfossils (pollen and phytoliths), paleosols, and molecular phylogenies (Graham, Reference Graham1999; Edwards et al., Reference Edwards, Osborne and Strömberg2010; Strömberg, Reference Strömberg2011). Three of these types of evidence (macrofossils, microfossils, and paleosols) are preserved within or adjacent to NPS units of the Midwest Region. Paleosols described from Lower Oligocene strata of the White River Group in Badlands National Park (Retallack, Reference Retallack1983) and Lower Miocene strata of the Arikaree Group in Agate Fossil Beds National Monument (Retallack, Reference Retallack1997), phytoliths collected from exposures of Brule Formation and undivided Miocene Monroe Creek and Harrison Formations with correlative strata in Scotts Bluff National Monument (Strömberg, Reference Strömberg2004), and grass macrofossils collected from the Ash Hollow Formation with correlative strata in Niobrara National Scenic River (Thomasson, Reference Thomasson1980) support three different scenarios regarding the age of open grassland ecosystems in the Great Plains (reviewed in Strömberg, Reference Strömberg2011).
3.4.1 Northern Great Plains Inventory and Monitoring Network
Fort Union Trading Post National Historic Site
The Fort Union Trading Post National Historic Site (FOUS) is located on the North Dakota–Montana state line south of Williston, North Dakota. Within FOUS, small pieces of petrified wood are commonly found amongst gravels that have been reworked by fluvial and glacial transport and are likely derived from the Paleocene Fort Union Group (Tweet et al., Reference Tweet, Santucci and Kenworthy2011; Tweet, Reference Tweet2016).
Theodore Roosevelt National Park
Theodore Roosevelt National Park (THRO) preserves about 70,000 acres of badlands in North Dakota. The park contains thick exposures of Paleocene rocks that represent three different formations well-known for their preservation of fossil wood, leaves, and reproductive structures at localities outside the park (Hickey, Reference Hickey1977; Crane et al., Reference Crane, Manchester and Dilcher1990; Erickson, Reference Erickson1999). Recently, park monitoring surveys have discovered similar material within THRO (Salcido et al., Reference Salcido, Wilson and Tweet2022) and suggest potential for further paleobotanical investigation.
Bullion Creek Formation: The Bullion Creek Formation records fluvial, lacustrine, and estuarine deposition from ca. 61 to 59 Ma (Hoganson and Murphy, Reference Hoganson and Murphy2003; Belt et al., Reference Belt, Hartman and Diemer2004; Anderson et al., Reference Anderson, Hartman and Wesselingh2006). Bullion Creek deposits in western North Dakota have a characteristic sequence of gray claystone and siltstone, lignite, yellow siltstones and sandstones, and sandstone (Salcido et al., Reference Salcido, Wilson and Tweet2022). These lithologies are interpreted as representing a succession of floodplain, swamp, crevasse splay, and point bar deposits. The formation preserves paleobotanical material, including leaves of Cercidiphyllum and unidentified petrified wood and seeds (Jacob, Reference Jacob1973; KellerLynn, Reference KellerLynn2007; Tweet et al., Reference Tweet, Santucci and Kenworthy2011). Outside of THRO, at Wannagan Creek Quarry, the Bullion Creek Formation preserves a diverse floral assemblage of more than 100 plant taxa and associated arthropod trace fossils based on leaf and fruit impressions in lignitic shale (Erickson, Reference Erickson1999; Haire et al., Reference Haire, Hanks and Tremain2001).
Sentinel Butte Formation: The lithology of the Upper Paleocene Sentinel Butte Formation is similar to the underlying Bullion Creek Formation, but the Sentinel Butte Formation can be distinguished by its weathered color (gray vs. yellow) and abundance of petrified wood (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). One particularly notable fossil locality within THRO is the Petrified Forest Plateau in the western section of the South Unit; this locality preserves fossil leaves and tree trunks (Figure 17), some of which are in their original growth position (Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991).

Figure 17 Fossil plants from the Midwest Region. (a) Unidentified fossil leaf collected from the Sentinel Butte Formation in the North Unit of Theodore Roosevelt National Park. Photos by Charles Salcido. Scale bar = 1 cm. (b) In situ fossil log in the Sentinel Butte Formation of Theodore Roosevelt National Park. (c) Petrified wood found along the banks of the Niobrara River in Niobrara National Scenic River.
The Petrified Forest Plateau preserves fossil trees in two organic-rich horizons, an upper lignite and a lower silicified layer, separated by 37 cm (15 in) of iron oxide–stained mudstones (Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991). Most wood appears to be cupressaceous (formerly described as taxodiaceous), according to Fastovsky and McSweeney (Reference Fastovsky and McSweeney1991, citing the help of Steve Manchester). Other wood includes Dacrydium and other podocarpaceous wood (unpublished reports by Coffin, Reference Coffin1984, and unpublished reports by his students, M. Arct and J. H Coulton) and Sciadopitys (Sciadopityaceae, formerly classified as Pinaceae; unpublished reports by B. F. Kukachka). Rare angiosperm wood has also been recovered, including a sample with similarities to Cercidiphyllum (Cercidiphyllaceae; Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991, with the help of S. Manchester). The Petrified Forest Plateau also preserves leaves of Cercidiphyllum and Platanus and Equisetum stems in lenticular deposits of fine-grained sandstone (Fastovsky and McSweeney, Reference Fastovsky and McSweeney1991). Collectively, the presence of thick organic horizons in poorly developed paleosols, abundant upright tree trunks with buttressed bases and taxonomic affinity to Cupressaceae, and fragmentary mollusk and gastropod fossils suggest that these deposits accumulated in a lowland swampy area of a floodplain, potentially analogous to modern bald cypress swamps.
The Sentinel Butte Formation has produced additional paleobotanical material from other localities within and near the park. The South Unit Loop Survey (Salcido et al., Reference Salcido, Wilson and Tweet2022) reported the fruits of Jenkinsella, the revised name of Cercidiphyllaceae fruits formerly known as Nyssidium (Golovneva and Alekseev, Reference Golovneva and Alekseev2017), and Porosia (fam. indet.), and Jain and Hall (Reference Jain and Hall1969) described the holotype of the fern megaspore Azolla stanleyi (Salviniaceae) from a horizon in the South Unit of THRO. Adjacent to the South Unit of THRO in Medora, North Dakota, silicified peat deposits preserve fossil leaves and wood of Ginkgo, Metasequoia, other conifers, and angiosperms (Ting, Reference Ting1972; Harr and Ting, Reference Harr, Ting and Romans1977), and quarries at the Medora Public Dig Site within the park preserve plant debris in carbonaceous mudstone.
Outside of THRO, the flora of the Sentinel Butte Formation is known from the Almont locality in Morton County and the Beicegel Creek locality in McKenzie County, North Dakota (Crane et al., Reference Crane, Manchester and Dilcher1990; Manchester et al., Reference Manchester, Pigg and Crane2004; Pigg and DeVore, Reference Pigg and DeVore2010). The Almont site preserves a diverse megaflora of 35–40 plant taxa based on leaf, fruit, and flower impressions and a palynoflora of more than 50 taxa (Crane et al., Reference Crane, Manchester and Dilcher1990; Pigg et al., Reference Pigg, Manchester and DeVore2008; Zetter et al., Reference Zetter, Farabee and Pigg2011). The Beicegel Creek site preserves many of the same taxa as Almont but in a slightly higher energy depositional environment and includes fossil fruits with organic preservation amenable to study through the cellulose acetate peel technique (Pigg et al., Reference Pigg, DeVore and Taylor2023).
Golden Valley Formation: The Golden Valley Formation conformably overlies the Sentinel Butte Formation and is divided into the lower Bear Den Member and the upper Camels Butte Member by laterally equivalent beds referred to as either the Taylor Bed or Alamo Bluff Lignite (Hickey, Reference Hickey1977; Tweet et al., Reference Tweet, Santucci and Kenworthy2011). These beds are used as markers for the Paleocene–Eocene boundary, ca. 56 Ma (Cohen et al., Reference Cohen, Finney, Gibbard and Fan2013). Within THRO, the Golden Valley Formation is only represented by erosional remnants of the Taylor Bed, a strongly resistant layer of silicified siltstone and freshwater limestone; the underlying Bear Den Member, composed mainly of siltstone and claystone, is thought to have eroded away (Hoganson and Campbell, Reference Hoganson and Campbell1997; Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The Taylor Bed has produced plant fragments (unidentifiable roots, stems, and twigs; Hoganson and Campbell, Reference Hoganson and Campbell1997; KellerLynn, Reference KellerLynn2007), and root and stem casts (Biek and Gonzalez, Reference Biek and Gonzalez2001). Outside of THRO, the contemporaneous Alamo Bluff Lignite and slightly older Bear Den Member have yielded palynomorphs and a moderately diverse macrofossil assemblage of Equisetum, ferns, conifers, and monocot and dicot angiosperms (Hickey, Reference Hickey1977).
Unnamed gravel deposits: Unnamed terrace gravel deposits containing petrified wood are present in the North and South Units of THRO. The age of these deposits is uncertain but is broadly considered to represent Neogene deposition prior to Pleistocene glaciations (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). It is also uncertain whether the petrified wood found in these deposits is reworked from older units (Salcido et al., Reference Salcido, Wilson and Tweet2022).
Wind Cave National Park
Wind Cave National Park (WICA) in southwestern South Dakota comprises about 34,000 acres of land surrounding Wind Cave, one of the most extensive cave systems in the world with more than 260 km (160 mi) of surveyed passages (Herring et al., Reference Herring, Tweet and Santucci2023). The stratigraphy and paleobotany of Cenozoic strata within WICA is similar to that of other parks with the NGPN, and the park’s primary Cenozoic paleobotanical resources are rhizoliths and Celtis endocarps from the Eocene–Oligocene White River Group. Such material has been recovered alongside invertebrate and vertebrate assemblages from several localities in the northeastern part of WICA, the Centennial Site and the Klukas Sites (Martin, Reference Martin1984, Reference Martin1988; Brown, Reference Brown2014). Quaternary sediments of the Red Valley in southwestern WICA have produced phytoliths and charcoal (Fredlund and Tieszen, Reference Fredlund and Tieszen1997).
Badlands National Park
Badlands National Park (BADL) in central South Dakota preserves approximately 240,000 acres and contains rich fossil assemblages in its Upper Cretaceous, Eocene, and Oligocene strata. Fossiliferous paleosols, sandstones, siltstones, and mudstones deposited from the Middle Eocene into the Early Oligocene belong to the White River Group. Although best known for their abundant vertebrate fossils, these strata also preserve significant paleobotanical remains in the form of wood, endocarps, and root traces (Tweet et al., Reference Tweet, Santucci and Kenworthy2011).
Chamberlain Pass Formation: The basal formation of the White River Group, the Chamberlain Pass Formation, is a thin (ca. 11 m) unit thought to have been deposited in the Middle to Late Eocene based on dates from the top of the overlying Chadron Formation of 33.9 ± 0.13 Ma (Prothero and Whittlesey, Reference Prothero and Whittlesey1995). The formation comprises conglomerate, sandstone channel deposits, overbank mudstones, and floodplain paleosols (Terry and Evans, Reference Terry and Evans1994). Paleosols of the Chamberlain Pass Formation are reported to contain root traces inside and outside BADL (Retallack, Reference Retallack1983; Tweet et al., Reference Tweet, Santucci and Kenworthy2011), and large pieces of fossil wood tentatively identified as Cupressaceae have been found in a basal conglomerate of the formation east of the park (Clark, Reference Clark1967).
Chadron Formation: The overlying Chadron Formation is interpreted to represent stream, pond, riverbank, and floodplain deposits that document a transition from a humid or subhumid woodland environment to drier and cooler grasslands during the latest Eocene (ca. 37–34 Ma) (Retallack, Reference Retallack1983; Prothero and Emry, Reference Prothero, Emry and Woodburne2004). Paleobotanically productive strata in this formation are the Gleska and Ohaka Series paleosols and a level at the base of the formation that produced fossil wood. Root traces occur in both the Gleska and Ohaka series paleosols, and the Gleska series is also a known source of Celtis endocarps (Retallack, Reference Retallack1983). The woody material collected from the base of the formation is a rooted stump with potential affinity to the Dipterocarpaceae, a tropical plant family with a modern center of diversity in southeast Asia (Shuler and Santucci, Reference Shuler and Santucci1986). Outside of BADL, the Chadron Formation contains freshwater limestone deposits with charophytes and aquatic plants and vertebrate coprolites with unidentified plant remains (Evans, Reference Evans1998, Reference Evans1999; Sandler et al., Reference Sandler, Chin and Rogers1998).
Brule Formation: The youngest member of the White River Group is the Lower Oligocene (ca. 33–30 Ma) Brule Formation (Prothero and Emry, Reference Prothero, Emry and Woodburne2004). The Brule Formation is divided into two members. The lower Scenic Member is characterized by well-developed and highly fossiliferous pinkish and green-gray paleosols and lesser quantities of silt, sand, and volcanic ash; the upper Poleslide Member is characterized by light-gray to gray-brown paleosols, eolian volcanic sediments, and highly fossiliferous sandstones (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The paleosols of each member contain root traces and Celtis endocarps and are interpreted as documenting a continued transition from a warm temperate and subhumid climate to a cool temperate and semiarid climate (Retallack, Reference Retallack1983, Reference Retallack1984). The Scenic Member of the Brule Formation also preserves unidentified tree stumps and bits of plants in coprolites (DiBenedetto, Reference DiBenedetto1998; Wrenn and DiBenedetto, Reference Wrenn and DiBenedetto2002).
Sharps Formation: The Upper Oligocene Sharps Formation overlies the White River Group and also comprises floodplain deposits and extensive paleosols (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The formation is divided by an unconformity into a lower (ca. 29.9–28.9 Ma) and an upper (ca. 28.4–20.0 Ma) section. This formation contains several series of paleosols that have been used to infer a cool semiarid climate and steppe/prairie vegetation (Retallack, Reference Retallack1981). The only paleobotanical material to be reported from the Sharps Formation within BADL are root traces (Retallack, Reference Retallack1983).
Quaternary sediments: Rawling and Fredlund (Reference Rawling and Fredlund1999) mention undescribed phytoliths collected from soils buried under eolian dunes at Sheep Mountain Table in BADL. Quaternary floodplain, river terrace, eolian, landslide, and alluvial deposits are present elsewhere in the park and may contain similar remains.
Agate Fossil Beds National Monument
Agate Fossil Beds National Monument (AGFO), located in northwestern Nebraska, is known for its rich vertebrate remains within fluvial Miocene units (Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020). These units also preserve evidence of ancient plant communities in root traces (rhizoliths) and plant fragments recovered from the Harrison and Anderson Ranch formations.
Harrison Formation: The Lower Miocene Harrison Formation is part of the Arikaree Group, a sedimentary package characterized by eolian, volcaniclastic, and fluvial sediments deposited during the Late Oligocene and Early Miocene (Hunt, Reference Hunt1990; Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020). Paleosols of the Harrison Formation preserve evidence of ancient vegetation in root traces and mollic epipedons, surface horizons of small, rounded clods of clayey and organic-rich soil that are produced by the fine roots and abundant earthworms characteristic of grassland ecosystems (Retallack, Reference Retallack1997; Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020). Some root traces are associated with plant fragments and occur within the corkscrew-shaped burrows of an extinct beaver called Palaeocastor (Martin and Bennett, Reference 113Martin and Bennett1977). These large burrows (210–275 cm [83–108 in] in length), known as Daemonelix, were initially interpreted as freshwater sponges or a novel type of extinct plant (Martin and Bennett, Reference 113Martin and Bennett1977). The age of the Harrison Formation is constrained to 22.9 ± 0.08 Ma based on 40Ar/39Ar dates of the Agate Ash, a thinly laminated white tuff that has been diagenetically altered to smectite (Izett and Obradovich, Reference Izett and Obradovich2001; Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020).
Anderson Ranch Formation: The Anderson Ranch Formation is fossiliferous throughout AGFO and contains two particularly well-known bonebeds at Carnegie Hill and University Hill. Each of these localities contains a succession of fluvial, lacustrine, eolian/volcaniclastic, and shallow freshwater deposits (Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020). Rhizoliths are present in silty sandstone deposits containing eolian volcaniclastic loess (Hunt, Reference Hunt1990). The conformable contact of the Anderson Ranch Formation with the underlying Harrison Formation is relatively close to the Agate Ash, suggesting that sediments of the Anderson Ranch Formation were also deposited during the Early Miocene, not much later than 22 Ma (Kottkamp et al., Reference Kottkamp, Santucci, Tweet, De Smet and Starck2020).
Scotts Bluff National Monument
Scotts Bluff National Monument (SCBL) in western Nebraska was established to preserve Scotts Bluff, a landmark for the California, Oregon, and Mormon trails, and the surrounding ca. 3,000 acres (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). Paleobotanical material found within the monument is mainly confined to root traces within paleosols, but equivalent units to those inside the park contain root traces, plant fragments, phytoliths, Celtis endocarps, and pollen.
Brule Formation: The Lower Oligocene Brule Formation is the uppermost unit of the White River Group. Exposures of the Brule Formation within SCBL are divided into two members, the Orella Member and the Whitney Member, and these members are correlative with the Scenic and Poleslide members of the Brule Formation recognized in BADL (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). Each member is known to contain paleobotanical resources near SCBL, but material collected from these units within SCBL is poorly documented. The Orella Member is the basal unit of the Brule Formation at SCBL and is composed of brown, gray, green, and pink volcaniclastic mudstone and siltstone (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). Outside of the park, the Orella Member has produced phytoliths (Strömberg, Reference Strömberg2004), root traces (LaGarry et al., Reference LaGarry, Terry and Hunt1998), casts of tree limbs, and other plant fragments (Nixon and LaGarry-Guyon, Reference Nixon and LaGarry-Guyon1993). The age of these resources is constrained to 34–31.85 Ma based on correlations of the underlying Chadron Formation and an ash layer within the overlying Whitney Member (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The Whitney Member has also produced paleobotanical material outside of SCBL, including plant fragments, root traces, phytoliths (Meehan, Reference Meehan1994; Grandstaff and Terry, Reference Grandstaff and Terry2009), and Celtis endocarps (Ash Hollow State Park; Diffendal et al., Reference Diffendal, Pabian and Thomasson1996). Based on the age of the base of the unconformably overlying Gering Formation, these materials were deposited between 31.85 and 28.31 Ma (Swinehart and Diffendal, Reference Swinehart and Diffendal1997).
Arikaree Group: Within SCBL, the Arikaree Group is represented by the Upper Oligocene Gering Formation, and the undivided Upper Oligocene to Lower Miocene Monroe Creek and Harrison formations. The Gering Formation is correlative with the Upper Sharps Formation exposed in BADL, and it also preserves root traces (Loope et al., Reference Loope, Mason, Bao, Kettler and Zanner2005). Deposition of the Gering Formation is thought to have occurred in a relatively short period of time based on an age of 28.1 ± 0.03 Ma for the Twin Sisters Pumice Conglomerate low in the unit, and an age of 27.29 ± 0.08 Ma for an ash bed in the overlying Monroe Creek Formation (Tedford et al., Reference Tedford, Swinehart, Swisher, Prothero and Emry1996; Bailey, Reference Bailey2004). The undivided rocks of the Monroe Creek Formation and Harrison Formation are brown and gray silty sandstones representing deposition in braided streams, ephemeral shallow lakes, and interchannel plains blanketed by windblown sediments. Within SCBL, these rocks contain zones of root traces and bioturbation (Graham, Reference Graham2009; Tweet et al., Reference Tweet, Santucci and Kenworthy2011), and phytoliths and root traces have been found in these formations outside of the park (Strömberg, Reference Strömberg2002, Reference Strömberg2004).
Chimney Rock National Monument
The Chimney Rock National Historical (CHRO) Site in southwestern Nebraska preserves a prominent landmark (Chimney Rock) for the California, Mormon, and Oregon trails (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The site, an NPS-affiliate owned by the Nebraska State Historical Society, is not officially part of the NGPN but has similar geology to SCBL located just 30 km (19 mi) to the northwest. The only paleobotanical materials preserved at CHRO are root casts in the Whitney Member of the Oligocene Brule Formation (Meehan, Reference Meehan1994). Outside of CHRO, the Whitney Member preserves plant fragments, root traces, phytoliths (Meehan, Reference Meehan1994; Grandstaff and Terry, Reference Grandstaff and Terry2009), and Celtis endocarps (Ash Hollow State Park; Diffendal et al., Reference Diffendal, Pabian and Thomasson1996).
Niobrara National Scenic River
Niobrara National Scenic River (NIOB) protects an area of about 23,000 acres along a 122 km (76 mi) stretch of the Niobrara River in north-central Nebraska. This NPS unit is rich in paleontological resources and preserves an extensive sequence from the past 15 million years (Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The Ogallala Group, comprised by the Valentine and Ash Hollow formations in NIOB, represents the Miocene portion of this sequence and preserves a variety of plant fossils.
Valentine Formation: The Valentine Formation underlies the Ash Hollow Formation, and its age is constrained to 13.6–12.18 Ma by the Hurlburt Ash of the basal Cornell Dam Member of the Valentine Formation and the Swallow Ash of the basal Cap Rock Member of the Ash Hollow Formation (Woodburne, Reference Woodburne2007). Within NIOB, the Cornell Dam Member has produced fossil reproductive structures of Berriochloa and Celtis alongside abundant mammal and herpetological remains at the Norden Bridge and Egelhoff quarries (Voorhies and Corner, Reference Voorhies and Corner1993; Gabel et al., Reference Gabel, Backlund and Haffner1998); wood is also present at Norden Bridge and other NIOB sites in the Valentine Formation (Voorhies, Reference Voorhies1990). Fossil wood (Figure 17) has been reported from the overlying Crookston Bridge Member of the Valentine Formation (G. Warrick, NIOB, written comm. to Tweet and Santucci, August 2021). One well-preserved piece of wood has vessels and aliform parenchyma; This combination of traits suggests an affinity with either the legume (Fabaceae) or mulberry (Moraceae) family. Other logs and wood fragments have been observed eroding from sandy channel deposits around the level of the Niobrara River. Outside of NIOB, the Kilgore flora contains a rich assemblage of similarly aged leaf and fruit fossils dominated by the leaves and associated fruitlets of Platanus vitifolia (MacGinitie, Reference MacGinitie1962).
Ash Hollow Formation: The Ash Hollow Formation documents deposition spanning at least 12.18–6.62 Ma based on ashes dated from the lower and upper parts of the unit (Tucker and Voorhies, Reference Tucker and Voorhies2005; Tweet et al., Reference Tweet, Santucci and Kenworthy2011). The only paleobotanical material recognized from the Ash Hollow Formation within NIOB are root casts in the Cap Rock Member, but the formation contains diverse plant records outside of the park. Three-dimensional, occasionally silicified fruits and seeds of Poaceae, Cyperaceae, Juglandaceae, and Ulmaceae have been recovered from several horizons of the Ash Hollow Formation during field collections and by floating sediments in water (Thomasson, Reference Thomasson1980, Reference Thomasson1987; Gabel et al., Reference Gabel, Backlund and Haffner1998). One particularly notable finding from the Cap Rock Member outside of NIOB is the preservation of Berriochloa anthoecia within the oral cavity and rib cage of articulated Teleoceras specimens (Voorhies and Thomasson, Reference Voorhies and Thomasson1979). This record provides insight into the diet of the extinct mammal and trophic interactions in the Miocene grassland of the Great Plains.
3.4.2 Great Lakes Inventory and Monitoring Network
Ice Age National Scenic Trail and Ice Age Scientific Reserve
Ice Age National Scenic Trails traces the southern boundary of the last glacial maximum for 2,000 km (about 1,200 mi) between Taylors Falls and Sturgeon Bay, Wisconsin. Ice Age National Scientific Reserve is an NPS-affiliate comprising nine Ice Age related areas managed by the state of Wisconsin. Paleobotanical resources within Ice Age National Scenic Trail areas include Late Pleistocene and Holocene palynomorphs from Devil’s Lake State Park (Attig et al., Reference Attig, Clayton, Lange and Maher1990), Butler Lake (Long et al., Reference Long, Power and McDonald2011), and the Two Creek Forest Bed in Two Creeks Buried Forest State Natural Area. The Two Creek Forest Bed is a 0.5–5 cm (0.2–2 in) thick layer of organic debris and macroscopic plant parts dated to 11,850 ybp (Culberson, Reference Culberson1955; Broecker and Farrand, Reference Broecker and Farrand.1963). The plant and insect communities of the Two Creek Forest Bed include 19 species of bryophytes, among them two (Ditrichum flexicaule and Calliergoa turgescens) that no longer occur in Wisconsin; wood, twigs, and cones of Picea mariana; and coleopteran, dipteran, and hymenopteran insects (Wilson, Reference Wilson1932, Reference Wilson1936; Culberson, Reference Culberson1955; Morgan and Morgan, Reference Morgan and Morgan1979).
Pictured Rocks National Lakeshore
Pictured Rocks National Lakeshore (PIRO) protects 730,000 acres on the southern shore of Lake Superior in Michigan’s Upper Peninsula. Proterozoic and Paleozoic sandstones comprise much of the bedrock within the lakeshore, but Holocene paleobotanical material is preserved in glaciofluvial deposits and ancient soils buried by the Great Sable Sand Dunes (Anderton and Loope, Reference Anderton and Loope1995), and in sediment cores drawn from Great Sable Lake and Beaver Lake (Fisher and Whitman, Reference Fisher and Whitman1999; Loope et al., Reference Loope, Fisher, Jol, Anderton and Blewett2004). The macropaleobotanical materials recovered from these localities are wood fragments that have been radiocarbon dated to establish a chronology for organic horizons within sediment cores (Anderton and Loope, Reference Anderton and Loope1995; Fisher and Whitman, Reference Fisher and Whitman1999; Loope et al., Reference Loope, Fisher, Jol, Anderton and Blewett2004). In sediment cores from Great Sable Lake, Loope et al. (Reference Loope, Fisher, Jol, Anderton and Blewett2004) identified a buried northern white cedar (Thuja occidentalis) forest from ca. 2990 ybp. Outside of PIRO, fossil pollen from the early Holocene (9500–6000 ybp) has been recovered from Buck Lake sediments and includes Picea, Pinus, Betula, Acer, Quercus, Ulmus, other deciduous trees, and prairie forbs (Davis et al., Reference Davis, Douglas and Calcote2000).
3.4.3 Heartland Inventory and Monitoring Network
Herbert Hoover National Historic Site
Herbert Hoover National Historic Site (HEHO) in eastern Iowa encompasses about 200 acres of surficial, unconsolidated Quaternary sediments (Graham, Reference Graham2011). Known Cenozoic paleobotanical resources in HEHO are limited; the only plant fossil reported from HEHO is a piece of Pleistocene petrified wood that was found 8 m (26 ft) below the surface in a well dug in 1983 (Graham, Reference Graham2011, Reference Graham2017).
3.5 Southeast, National Capital, and Northeast Regions
The Southeast, National Capital, and Northeast regions encompass the remainder of NPS units that contain Cenozoic paleobotanical resources (Figure 18). Cenozoic paleobotanical material is comparatively sparse in this part of the USA but has been found in 15 NPS-administered units that fall into six inventory and monitoring networks: the Gulf Coast Network, National Capital Region Network, Mid-Atlantic Network, Eastern Rivers and Mountains Network, Northern Coast and Barrier Network, and the Northeast Temperate Network.
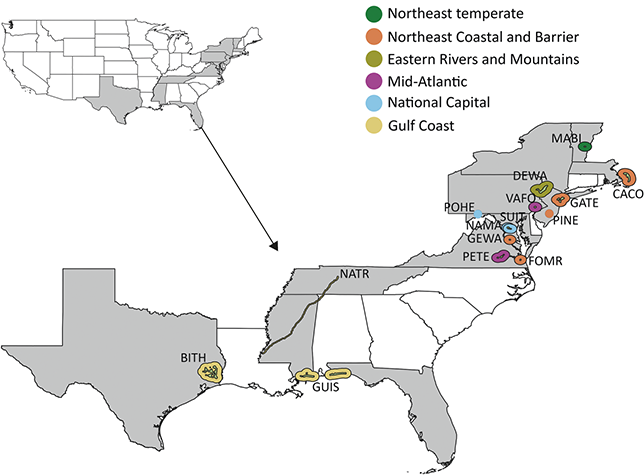
Figure 18 Map of NPS units with Cenozoic paleobotanical resources in the Southeast, National Capital, and Northeast regions, colored by inventory and monitoring network. See Figure 1 legend for details on the park symbology and acronyms.
Paleobotanical resources from the NPS units in eastern USA mainly occur in surficial Quaternary deposits or in the subsurface; however, numerous significant paleofloras are known from outside NPS areas. Paleogene floras of the region are preserved in sediments of the Wilcox and Claiborne Groups in the Mississippian Embayment (Berry, Reference Berry1916a; Dilcher, Reference Dilcher1971; Crepet and Nixon, Reference Crepet and Nixon1989), and a smaller number of Neogene floras are scattered across the eastern USA in Tennessee (Gray fossil site; Hermsen, Reference Hermsen2021, Reference Hermsen2023), Alabama (Citronelle flora; Stults and Axsmith, Reference Stults and Axsmith2015), Maryland (Brandywine flora; McCartan et al., Reference McCartan, Tiffney and Wolfe1990), and Vermont (Brandon Lignite; Tiffney, Reference 125Tiffney1994). Within NPS units, some notable Quaternary assemblages include the Pleistocene seed and fruit floras found alongside abundant vertebrate remains in the Cumberland and Port Kennedy bone caves, which are located along the Potomac National Heritage Trail and in Valley Forge National Historic Park, and the remains of a Pleistocene bald cypress (Taxodium) swamp discovered during excavations at Washington Circle, in the National Mall, Washington, DC (Figure 19). Notable subsurface discoveries from NPS units are the palynofloras of the Mays Landing borehole from affiliated Pinelands National Preserve in New Jersey that provide data on the composition of Oligocene and Miocene floras of the northeast (Owens et al., Reference Owens, Bybell and Paulachok1988; Graham, Reference Graham1999) and the palynofloras of Duck Pond from Cape Cod National Seashore that record changing tree distributions during the Holocene (Winkler, Reference Winkler1985).
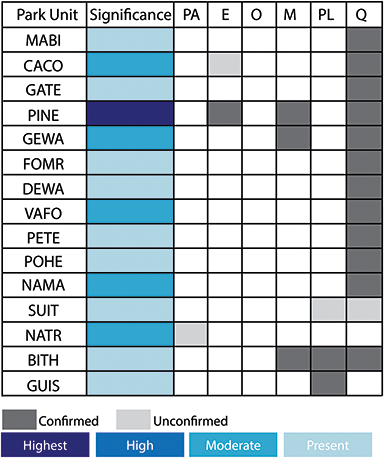
Figure 19 Summary of the temporal distribution and significance of Cenozoic paleobotanical resources in the Southeast, National Capital, and Northeast regions. See Figure 2 legend for explanation of significance ratings.
3.5.1 Northeast Temperate Inventory and Monitoring Network
Marsh-Billings-Rockefeller National Historic Park
Marsh-Billings-Rockefeller National Historic Park (MABI) in eastern Vermont commemorates an area significant to the lives of several conservationists: George Perkins Marsh, Frederick Billings, Mary French Rockefeller, and Laurance S. Rockefeller. The only Cenozoic paleobotanical materials known from within MABI are two pieces of marl from The Pogue, a former wetland that is now a manmade pond (Tweet et al., Reference Tweet, Santucci and Kenworthy2010; Thornberry-Ehrlich, Reference Thornberry-Ehrlich2011). These pieces of marl are dated to the Holocene and preserve veined leaf impressions, twigs, and grass stems (Tweet et al., Reference Tweet, Santucci and Kenworthy2010; Thornberry-Ehrlich, Reference Thornberry-Ehrlich2011). The specimens are held in the park’s collections.
3.5.2 Northeast Coastal and Barrier Inventory and Monitoring Network
Cape Cod National Seashore
Cape Cod National Seashore (CACO) in Massachusetts preserves about 65 km (40 mi) of sandy beaches, marshes, and ponds along the Atlantic coast. Paleobotanical material has been recovered from CACO in four different contexts: microfossils in subsurface deposits retrieved from deep wells or boreholes; reworked petrified wood in surficial outwash deposits; Pleistocene petrified wood; and microfossils in surficial lake or pond sediments retrieved by shallow cores (Tweet et al., Reference Tweet, Santucci and Connors2014). The material found in deep wells from CACO are palynomorphs, lignite coal, and carbonized wood interpreted as Eocene in age (Zeigler et al., Reference Zeigler, Tuttle, Tasha and Giese1964), and the fossiliferous sediments may have been initially deposited elsewhere and subsequently transported by glaciers (Uchupi et al., Reference Uchupi, Giese, Aubery and Kim1996). Dark gray silicified wood fragments that are Cretaceous or Early Cenozoic in age are present in the glacial drift of western Cape Cod and have traced to a bedrock source near Middleboro, Massachusetts (Oldale, Reference Oldale2001). Several samples of Pleistocene petrified wood were found in the Wellfleet Quadrangle, within CACO and dated to over 25,000 radiocarbon ybp (Oldale, Reference Oldale1968). Quaternary pollen cores from lakes and ponds within CACO are the park’s most significant paleobotanical material and record the regional vegetation over approximately the past 11,500 years (Tweet et al., Reference Tweet, Santucci and Connors2014). One site in Wellfleet, Duck Pond, has an especially complete record of the past 12,000 years, and its deposits include identifiable moss macrofossils (Winkler, Reference Winkler1985).
Gateway National Recreation Area
The Gateway National Recreation Area (GATE) consists of three geographically isolated units in New York (NY) and New Jersey (NJ): Jamaica Bay Unit (NY), Sandy Hook Unit (NJ), and the Staten Island Unit (NY). Pre-Quaternary deposits are not exposed in any of these units, and there is a hiatus between the Upper Cretaceous and Pleistocene in the bedrock (Tweet et al., Reference Tweet, Santucci and Connors2014), limiting the potential of GATE to furnish Cenozoic plant macrofossils.
Paleobotanical resources discovered within or near GATE are specimens of plant debris found in reworked nodules that have washed ashore on beaches of the Sandy Hook and Jamaica Bay units (Chamberlain et al., Reference Chamberlain, Stoffer and Messina2006) and lignite, wood, rootlets, and reed fragments recovered from boreholes at the Sandy Hook and Jamaica Bay units (Tweet et al., Reference Tweet, Santucci and Connors2014). Potential sources of fossiliferous nodules are the Quaternary Cape May (300–75 kya) and Gardiner Clay (125–75 kya) formations and the Cenomanian–Turonian Magothy and Raritan formations (Chamberlain et al., Reference Chamberlain, Stoffer and Messina2006; Tweet et al., Reference Tweet, Santucci and Connors2014).
Boreholes from the Sandy Hook Unit extend through Quaternary and Upper Cretaceous deposits (Kümmel and Poland, Reference Kümmel and Poland1910; Minard, Reference Minard1969). Some of the Cretaceous deposits have been attributed to the Raritan Formation, which has yielded exceptional plant macrofossils and mesofossils outside of GATE (Berry, Reference Berry1911; Crepet et al., Reference Crepet, Nixon and Gandolfo2001). Higher in the stratigraphic column, Quaternary pollen records from GATE are relevant to reconstructions of forest succession in New York and New England during the Holocene (Tweet et al., Reference Tweet, Santucci and Connors2014).
Pinelands National Reserve
Pinelands National Reserve (PINE) is an NPS-affiliated unit located in the New Jersey Coastal Plain. As with other units of the eastern states, most paleobotanical resources and all pre-Quaternary resources associated with the Pinelands have been derived from boreholes. Boreholes taken from several localities within PINE (Owens et al., Reference Owens, Bybell and Paulachok1988; Miller et al., Reference Miller, Sugarman and van Fossen1994, Reference Miller, Sugarman and Browning1998, Reference Miller, Sugarman and Browning1999; Browning et al., Reference Browning, Sugarman, Miller, Miller, Sugarman and Browning2011) provide a detailed account of the region’s subsurface stratigraphy and include paleobotanical materials such as pollen, wood, and phytoliths from formations that span the Middle Eocene to Late Miocene. The palynofloras of the Mays Landing borehole (Owens et al., Reference Owens, Bybell and Paulachok1988) are one of few available lines of evidence for reconstructing the Paleogene and Neogene vegetational history of the northeastern USA (Graham, Reference Graham1999). The Pinelands are also a source of Quaternary palynological resources, and pollen samples have been used to study the dynamics of forest succession during the Pleistocene and Holocene (Potzger, Reference Potzger1945; Florer, Reference Florer1972).
Paleogene units: The oldest Cenozoic paleobotanical material discovered within PINE are wood fragments and burrowed pyritized wood from the Middle–Late Eocene Shark River Formation; these were retrieved from corehole ACGS-4 along Great Egg Harbor River and the Double Trouble core site located in the northern part of PINE (Owens et al., Reference Owens, Bybell and Paulachok1988; Browning et al., Reference Browning, Sugarman, Miller, Miller, Sugarman and Browning2011). The Shark River Formation represents lower shoreface and marine shelf deposits. Borehole ACGS-4 also produced wood and pollen from three Oligocene units representing nearshore and marine shelf deposits (Owens et al., Reference Owens, Bybell and Paulachok1988).
Cohansey Sand and Kirkwood Formation (Miocene): The Cohansey Sand was deposited during the Middle to Late Miocene in paludal and lagoon environments, and an organic-rich layer present in the lagoon deposits has yielded wood fragments and pollen in boreholes taken along Great Egg Harbor River (ACGS-4 corehole) and Bass River (Owens et al., Reference Owens, Bybell and Paulachok1988; Miller et al., Reference Miller, Sugarman and Browning1998). The Kirkwood Formation represents neritic and prodeltaic deposits from the Early to Middle Miocene, and sections from corehole ACGS-4 and a core taken in Ancora, New Jersey, contain wood, pollen, and phytoliths (Owens et al., Reference Owens, Bybell and Paulachok1988; Miller et al., Reference Miller, Sugarman and Browning1999).
George Washington Birthplace National Monument
George Washington Birthplace National Monument (GEWA) is located in eastern Virginia along the Potomac River and preserves the area around the former plantation owned by the Washington family. Plant macrofossils have been recovered in GEWA from the Lower to Middle Miocene Calvert Formation and just outside of GEWA from the Pleistocene Tabb Formation. These units are exposed, respectively, on riverbanks of the Potomac and one of its tributaries, Popes Creek.
Calvert Formation: The Lower to Middle Miocene Calvert Formation represents shallow marine deposits and is known to contain diverse macrofossil types such as wood, leaves, invertebrate shell fragments, fish bones, scales, teeth, and mammal bones (Shattuck, Reference Shattuck1904; Vogt and Eshelman, Reference Vogt and Eshelman1987; Tweet et al., Reference Tweet, Santucci and Connors2014; Tweet and Santucci, Reference Tweet and Santucci2017). The exposures within GEWA are a source of shark teeth and pieces of carbonized wood, presumably derived from waterlogged trunks swept out to sea. Significant plant macrofossil localities of the Calvert Formation occur in Richmond, Virginia, and the District of Columbia, and are interpreted as representing the landward extent of the formation (Berry, Reference Berry1916b).
Tabb Formation: The Pleistocene Tabb Formation consists of river and coastal plain deposits and is subdivided into the Sedgefield, Lynnfield, and Poquoson members, which collectively span from 130 to 33 kya (Mixon et al., Reference Mixon, Berquist, Newell and Johnson1989; Scott et al., Reference Scott, Swift, Whittecar and Brook2010). An assemblage of fossil fruits, seeds, and tendrils was recovered from deposits of the Tabb Formation on the bank of Popes Creek outside of GEWA, a tributary of the Potomac River that forms the park’s southern boundary (Berry, Reference Berry1938; Tweet et al., Reference Tweet, Santucci and Connors2014). Taxa described from this assemblage include aquatic monocotyledons (Naias and Zannichellia palustris) and herbaceous, woody, and vining dicotyledons (Quercus, Phytolacca, Vitis) (Berry, Reference Berry1938). Plant fossil localities of the Tabb Formation have also been recovered to the west and south of GEWA on the banks of the Rappahannock River (Berry, Reference Berry1906, Reference Berry1909). At these localities, plant macrofossils – mainly seeds – were recovered from a peat layer overlying in situ cypress stumps and likely come from the Sedgefield Member of the Tabb Formation (Mixon et al., Reference Mixon, Berquist, Newell and Johnson1989).
Fort Monroe National Monument
Fort Monroe National Monument (FOMR) commemorates a historic military base in Hampton, Virginia, on the southern tip of the Virginia peninsula. From an artesian well in FOMR, Fontaine (Reference Fontaine1882) described a 1 m (3 ft) stratum of coarse, yellowish-green sand and indurated sandstone containing coal and plant fragments intervening between Miocene marine deposits above and below.
3.5.3 Eastern Rivers and Mountains Inventory and Monitoring Network
Delaware Water Gap National Recreation Area
Delaware Water Gap National Recreation Area (DEWA) preserves about 70,000 acres of forest and waterway along the Delaware River in northwestern New Jersey and southwestern Pennsylvania. The only paleobotanical discovery within Cenozoic sediments of DEWA is sub-fossil wood recovered at a depth of 6.4 m (21 ft) in a pit dug during the construction of the Bushkill access area in Bushkill, Pennsylvania. The log yielded a radiocarbon date of 4105 ± 90 ybp (ver Straeten et al., Reference ver Straeten, Monteverde, Inners, Inners and Fleeger2001).
3.5.4 Mid-Atlantic Inventory and Monitoring Network
Valley Forge National Historic Park
Valley Forge National Historic Park in eastern Pennsylvania preserves 3,500 acres surrounding an encampment site of the Continental Army during the Revolutionary War. The area surrounding the former encampment includes the Kennedy Bone Cave, a sinkhole deposit that was discovered in the 1870s during limestone quarrying near the former town of Port Kennedy, Pennsylvania. The cave is thought to have formed as an underground cavern from dissolution of limestone, and it eventually became a sinkhole when the roof collapsed. During the middle Pleistocene, amphibians, reptiles, birds, mammals, and plants fell into the hole and became buried by debris. Plant remains include twigs, leaves, fruits, and seeds and are referrable to extant species, including Quercus alba, Corylus americana, and Pinus rigida (Mercer, Reference Mercer1899).
Petersburg National Battlefield
Petersburg National Battlefield in Petersburg, Virginia, commemorates the site of a prolonged siege between the Confederate and Union armies during the American Civil War. The unit comprises a small area with few geologic exposures, but a twig was reported in an oxbow lake clay deposit from the center of the Petersburg Crater site (Hal Pranger, Geologic Resources Division, pers. comm.).
3.5.5 National Capital Region Inventory and Monitoring Network
Potomac Heritage National Scenic Trail
Potomac Heritage Trail (POHE) encompasses about 1,100 km (684 miles) of existing and planned trails that traverse Pennsylvania, Maryland, Virginia, and Washington, DC. The Cumberland Bone Cave in Cumberland, Maryland, is a sinkhole deposit that preserves a middle Pleistocene flora and fauna and is located along the Great Allegheny Passage trail of the POHE network (Hodnett et al., Reference Hodnett, Eshelman, Gardner and Santucci2023a). The cave was collected periodically between 1912 and 2012 and contains snails, fish, amphibians, reptiles, birds, mammals, and plants. Plant remains include indeterminate seeds and grass stems and fruits of Celtis occidentalis and Quercus.
National Mall and Memorial Parks
The National Mall and Memorial Parks (NAMA) in Washington, DC, includes fourteen NPS units that preserve 1,000 acres of parkland and numerous significant natural and cultural resources within the nation’s capital (Figure 18). Pleistocene fossil plant material has been recovered from two units of NAMA, the Washington Monument and Washington Circle (Darton, Reference Darton1950; Knox, Reference Knox1966). The fossil plant material from the Washington Monument was recovered from three of fifteen test boreholes that were made to evaluate the prospect of placing concrete piers down to bedrock under the foundation of the monument. These cores revealed that approximately 18 m (60 ft) of sand, gravel, and blue clay overly bedrock within the grounds of the Washington Monument and yielded partially petrified wood and petrified wood in sand (Darton, Reference Darton1950).
Various excavations in the District of Columbia between 16th Street NW and Washington Circle have revealed deposits of a Pleistocene bald cypress swamp (Berry, Reference Berry1933; Knox, Reference Knox1966). These deposits contain in situ cypress stumps and diverse fruits, seeds, leaves, and pollen (Berry, Reference Berry1933; Knox, Reference Knox1966). Berry (Reference Berry1933) described 37 different kinds of plants, which include bald cypress (Taxodium distichum), monocots, and woody and herbaceous dicots. Knox (Reference Knox1966) mapped the lateral extent of the swamp and collected pollen samples from several levels of the swamp deposits. The dominant components of the palynoflora shift from spruce to pine to deciduous angiosperms moving up in the profile, and Knox (Reference Knox1966) suggested that these shifts reflect a warming trend that is concordant with the swamp having existed during the most recent (Sangamon) interglacial, approximately 100,000 ybp.
Suitland National Parkway
Suitland National Parkway (SUIT) is part of National Capital Parks East (NACE), a management unit that encompasses 14 park sites, parkways, and statuary preserving significant natural and cultural heritage and covering about 8,000 acres between Capitol Hill and the Maryland suburbs (Nelson et al., Reference Nelson, Santucci and Tweet2019). Underlying this network are three significant fossiliferous rock units: the Potomac Group (Upper Cretaceous), the Pamunkey Group (Paleocene–Eocene), and the Chesapeake Group (Lower–Middle Miocene). Cenozoic plant macrofossil occurrences in this network are limited to undiagnostic wood fragments recovered from within SUIT during the construction of the Branch Avenue Metro Station (Kenworthy and Santucci, Reference Kenworthy and Santucci2004). However, plant fossil localities in close proximity to NACE units have been recorded in the Calvert Formation at Good Hope Hill, Washington, DC, and in “Upland gravels” (map unit Tu) in Brandywine, Maryland (Berry, Reference Berry1916b; Carr, Reference Carr1950; McCartan et al., Reference McCartan, Tiffney and Wolfe1990). The Calvert Formation consists mainly of shallow marine sediments deposited during the Early and Middle Miocene (Vogt and Eshelman, Reference Vogt and Eshelman1987; Tweet et al., Reference Tweet, Santucci and Connors2014). It is present within several NACE units and is a major formation within SUIT, where it has yielded cetacean and other mammal fossils (Nelson et al., Reference Nelson, Santucci and Tweet2019).
The “Upland gravels” are an informal unit consisting of poorly sorted sand, cobbles, pebbles, and gravel laid down by the ancestral Potomac River during the Late Miocene and Pliocene (McCartan et al., Reference McCartan, Tiffney and Wolfe1990; Nelson et al., Reference Nelson, Santucci and Tweet2019), and the deposits are present with six NACE units. This unit is the source of the Brandywine flora, one of few Neogene sites in eastern North America that has produced well-preserved macrofossils, including leaves, seeds, fruit, and pollen that represent 49 different taxa, and one of the last occurrence of Pterocarya (Juglandaceae) before it went extinct in North America (McCartan et al., Reference McCartan, Tiffney and Wolfe1990; Stults et al., Reference Stults, Wagner-Cremer and Axsmith2011, Reference Stults, Tiffney and Axsmith2022).
3.5.6 Gulf Coast Inventory and Monitoring Network
Natchez Trace Parkway
Natchez Trace Parkway (NATR) is a 715 km (444 mi) recreational road that stretches between Natchez, Mississippi, and Nashville, Tennessee. It does not contain confirmed records of Cenozoic paleobotanical material; however, leaf impressions and fossil wood have been found from a locality known as the Jeff Busby site in the Little Mountain Recreation Area, near Mathiston, Mississippi (Black, 1962; Kenworthy et al., Reference Kenworthy, Santucci and Visaggi2007). Based on geological maps, this material was likely collected from the Paleocene Tuscahoma Formation, but the locality has not been relocated since it was first found and it is not clear if the fossils came from within or just outside NATR (Hodnett et al., Reference Hodnett, Boensch, Smith, Tweet and Santucci2023b). There are other Cenozoic formations present within NATR that have yielded paleobotanical fossils outside the park boundary. These formations include the Zilpha Clay, Porters Creek Clay, Kosciusko Sand, and Vicksburg Group (Kenworthy et al., Reference Kenworthy, Santucci and Visaggi2007).
Outside of NATR, the Tuscahoma formation has produced Paleocene and Eocene palynomorphs from cores in Mississippi and Alabama (Harrington, Reference Harrington2008) and an Early Eocene macroflora from the Red Hot Truck Stop locality in Meridian, Mississippi (Danehy et al., Reference Danehy, Wilf and Little2007). This formation is thought to include the Paleocene–Eocene boundary (Harrington et al., Reference Harrington, Kemp and Koch2004).
Big Thicket National Preserve
Big Thicket National Preserve (BITH) in east Texas was the first preserve to be incorporated in the NPS, in 1974. The preserve protects water corridors and a variety of plant communities, such as floodplain forest and swamps, savannah, and upland forests, that comprise approximately 97,000 acres. Within BITH, geologic strata are mainly exposed in erosive contexts such as riverbanks. They were deposited between the Miocene and the Holocene (Kenworthy et al., Reference Kenworthy, Santucci and Visaggi2007). Three formations within BITH, the Miocene Fleming Formation, the Pliocene–Pleistocene Willis Formation, and the Pleistocene Beaumont Formation, have produced petrified wood, but in some instances, this wood may be reworked from older sources exposed north of the park, such as the Catahoula Formation (Singleton, Reference Singleton2008; Thornberry-Ehrlich, Reference Thornberry-Ehrlich2018).
The Willis Formation is the most significant source of petrified wood within BITH. Fossil localities from this formation along the Trinity and Neches rivers yield petrified wood from basal gravels and from Pleistocene fluvial strata (Kenworthy et al., Reference Kenworthy, Santucci and Visaggi2007). The wood specimens from localities in the park are unstudied, but areas adjacent to BITH with Pleistocene outcrops of the Willis Formation have produced fossil wood belonging to Quercus and Ulmus (Singleton, Reference Singleton2008; Thornberry-Ehrlich, Reference Thornberry-Ehrlich2018). In the northern section of BITH, a piece of petrified palm wood was reported, but the record lacks stratigraphic context (Watson, Reference Watson2006).
Gulf Islands National Seashore
Gulf Islands National Seashore (GUIS) protects 258 km (160 mi) of coastline, primarily barrier islands off the mainland of Florida and Mississippi. The only paleobotanical record from GUIS is pollen and pulverized plant material recovered from Pliocene sediments in a core taken from Horn Island in Mississippi (Gohn et al., Reference Gohn, Brewster-Wingard and Cronin1996; Clinton et al., Reference Clinton, Visaggi, Tweet, Santucci and Irick2023).
4 Conclusion
In our inventory report, we have demonstrated that the fossil plant resources contained in the NPS are: (1) geographically widespread, occurring in 74 NPS units distributed through all seven NPS regions and 25 states; (2) temporally widespread, occurring in 13 Paleocene formations, 27 Eocene formations, 19 Oligocene formations, 27 Miocene formations, 12 Pliocene formations, and 50 Quaternary units; (3) taxonomically diverse, including bryophytes, ferns, gymnosperms, and at least 40 families of angiosperms; (4) globally significant, yielding 393 type specimens and numerous biostratigraphically and biogeographically notable occurrences; and (5) in need of further curation, monitoring, and research.
Cenozoic fossil plants of the NPS are a significant scientific, educational, and cultural resource that have great potential to deepen global understanding of the history of plants, terrestrial ecosystems, and climate, and to connect and educate public audiences about their natural heritage. Cenozoic NPS paleobotanical resources are also heterogeneous, and vary considerably in their quality and abundance, extent of previous collection and study, and degree of awareness among scientific and public audiences. On one end of this spectrum are paleobotanical Lagerstätten such as the lacustrine shales of Florissant Fossil Beds and Fossil Butte National Monuments, and the fossil forests of Yellowstone National Park that have sustained over a century of paleobiological research and hold prominent profiles in the public’s perception of paleobotany; even among these sites, a consummate baseline understanding of taxonomy has not been achieved (e.g., Supplementary Table 8). Then there are NPS units like Aniakchak National Monument and Preserve, Denali, Katmai, and Wrangell–St. Elias National Parks and Preserves that have produced extensive paleobotanical collections, including type specimens, but require additional geochronologic, sedimentological, and stratigraphic context as well as further taxonomic study to be fully integrated into our understanding of the floristic, vegetational, and paleoclimatic history of North America. Conversely, there are other NPS units such as Theodore Roosevelt and Glacier National Parks whose paleobotanical resources are well-situated in stratigraphic context but have not yet, or only recently (e.g., Smith et al., Reference Smith, Greenwalt and Manchester2023) been extensively collected or studied taxonomically. Elsewhere on this spectrum are the late Neogene and Pleistocene resources of Golden Gate and Santa Monica National Recreation Areas that represent small, well-studied collections that are poorly known to the public despite being well-suited to science communication efforts. The records of these two parks document the recent evolutionary history of extant species (e.g., P. radiata) and ecosystems (e.g., closed-cone pine forests) with possible implications for land management and conservation (Axelrod, Reference Axelrod and Philbrick1967; Millar, Reference Millar1998). Finally, there are the many instances where NPS units contain minor paleobotanical resources (e.g., occasional petrified wood) that should continue to be monitored but are unlikely to provide major contributions to paleobotany.
The successful management of NPS fossil plant resources to fulfill their potential for scientific research and science communication will require an appropriate pairing between management intensity and the scope, significance, and distribution of fossil plants within NPS units. The baseline inventory presented here offers a starting point for the development of monitoring, curation, and research strategies that meet the unique needs of each of these 74 NPS units with Cenozoic paleobotanical resources. After fostering over 150 years of paleobotanical exploration, lands protected by the National Park Service continue to yield important fossil discoveries and will certainly play an important role in the future of North American paleobotany.
Acknowledgments
This inventory of paleobotanical resources from National Park Service areas would not have been possible without the previous research undertaken and articles published by scores of paleontologists, all of whom we have identified and cited in this publication. This project was supported through the National Park Service–Paleontological Society partnership known as the Paleontology in the Parks Fellowship Program. We would like to specifically recognize the leadership within the Paleontological Society for their support of this project and the fellowship, including Rowan Lockwood, College of William & Mary and past president of the Paleontological Society, and Brenda Hunda, Curator of Invertebrate Paleontology, Cincinnati Museum Center, Geier Collections & Research Center. We also appreciate the support of National Park Service staff within the Geologic Resources Division and those working at various parks and monuments. Finally, we extend our thanks to Dale Greenwalt and MacKenzie Smith for providing photos of fossils from Glacier National Park used in this publication.

Series Editor-in-Chief
Brenda R. Hunda
Cincinnati Museum Center
About the Series
The Elements of Paleontology series is a publishing collaboration between the Paleontological Society and Cambridge University Press. The series covers the full spectrum of topics in paleontology and paleobiology, and related topics in the Earth and life sciences of interest to students and researchers of paleontology.
The Paleontological Society is an international nonprofit organization devoted exclusively to the science of paleontology: invertebrate and vertebrate paleontology, micropaleontology, and paleobotany. The Society’s mission is to advance the study of the fossil record through scientific research, education, and advocacy. Its vision is to be a leading global advocate for understanding life’s history and evolution. The Society has several membership categories, including regular, amateur/avocational, student, and retired. Members, representing some 40 countries, include professional paleontologists, academicians, science editors, Earth science teachers, museum specialists, undergraduate and graduate students, postdoctoral scholars, and amateur/avocational paleontologists.