


Sustainable dairy production encompasses economic, environmental and social dimensions, as well as animal welfare and consumer values. European consumers not only value the quality of animal-derived foods but increasingly emphasize animal welfare considerations (Lovarelli et al., Reference Lovarelli, Bacenetti and Guarino2020). One animal welfare indicator that has attracted considerable public attention is the lifespan of dairy cows. The productive lifespan of high-yielding dairy cows is often substantially shorter than their natural life expectancy of approximately 20 years (De Vries and Marcondes, Reference De Vries and Marcondes2020). In Sweden, the average lifespan of a dairy cow is approximately 5 years (61.4 months), corresponding to a productive life of about 2.4 lactations. This estimate reflects the period from first calving to culling, assuming an average age at first calving of 27 months (Växa, 2024). Approximately 75% of Swedish dairy cows reach their second lactation, 50% reach the third and only 30% reach the fourth lactation (NAV, 2010).
The primary reasons for culling include infertility, mastitis and lameness (Langford and Stott, Reference Langford and Stott2012). The incidence of mastitis generally increases with lactation number (Verbeke et al., Reference Verbeke, Piepers, Supré and De Vliegher2014). In Sweden, the incidence of veterinary-treated clinical mastitis increases from 7.1 cases per 100 cows in primiparous cows to 23.1 cases per 100 cows in cows in their sixth or higher lactation (Växa, 2024). However, some studies have reported comparable mastitis incidence between primiparous cows and substantially older cows (McDougall et al., Reference McDougall, Arthur, Bryan, Vermunt and Weir2007).
Early culling negatively affects farm profitability, as milk yield typically increases until approximately the fifth lactation before declining thereafter (Smith et al., Reference Smith, Strawderman, Schukken, Wells, Pradhan, Espejo, Whitlock, Van Kessel, Smith and Wolfgang2010; Langford and Stott, Reference Langford and Stott2012). Clasen et al. (Reference Clasen, Fikse, Ramin and Lindberg2024) demonstrated that improved management strategies could extend the average productive lifespan of a dairy herd by 1 year, simultaneously reducing methane emissions and improving farm profitability. Similarly, De Vries and Marcondes (Reference De Vries and Marcondes2020) reported that increased cow longevity reduced phosphorus losses and methane emissions per unit of milk and meat produced. However, extending productive lifespan requires that milk quality and technological properties are maintained and not adversely affected.
In addition to infection status, lactation number is one of the most influential factors affecting somatic cell count (SCC). Numerous studies have reported higher average SCC in milk from older cows compared with younger cows (Salsberg et al., Reference Salsberg, Meek and Martin1984; Alhussien and Dang, Reference Alhussien and Dang2018b). Although SCC naturally increases with age and advancing stages of lactation, elevated SCC may also be observed in otherwise healthy cows (Walstra et al., Reference Walstra, Wouters and Geurts2006). SCC is widely used as an indicator of mastitis prevalence within dairy herds, raw milk quality and overall hygienic conditions of milk production (Petzer et al., Reference Petzer, Karzis, Donkin, Webb and Etter2017). European Union legislation mandates regular SCC monitoring as part of raw milk quality control programmes, and dairies commonly apply financial incentives for low-SCC milk due to the association between high SCC and impaired technological properties and reduced shelf life of dairy products (Barbano et al., Reference Barbano, Rasmussen and Lynch1991).
Elevated SCC, typically defined as >300,000 cells/ml, is associated with increased activity of the indigenous proteolytic enzyme plasmin (PL) (Politis et al., Reference Politis, Lachance, Block and Turner1989). PL is generated through the activation of its zymogen plasminogen (PG) by urokinase-type activators associated with somatic cells (Politis, Reference Politis1996). The PL-PG system plays a central role in milk protein degradation, particularly of caseins, which are critical for cheese yield and overall dairy product quality. Consequently, PL and PG concentrations and activities are key milk quality parameters when considering strategies to extend the productive lifespan of dairy cows.
In our previous study (Johansson et al., Reference Johansson, Lindberg and Lundh2024), pooled milk samples from younger cows (lactations 1 and 2) and older cows (lactations ≥3) were compared. Milk from older cows exhibited higher PL activity, whereas milk from younger cows showed higher PG-derived activity and greater total proteolysis. Building on these findings, the present study analysed individual cow milk samples to investigate associations between lactation number and milk composition, SCC and protein profile. Emphasis was placed on the proteolytic activity of PL and PG in relation to lactation number. Milk samples were collected from Swedish Red Breed (SRB) and Swedish Holstein (SH) cows, and milk quality traits associated with proteolysis were evaluated.
We hypothesized that increasing lactation number is associated with systematic changes in milk quality traits, including SCC, casein composition and the activity of the PL-PG system, resulting in altered proteolytic activity in milk from older cows compared with younger cows. The variables included in the statistical model were PL, PG, the PG-to-PL ratio (PG/PL), total casein, total β-casein, β-casein A1, β-casein A2, β-casein B and SCC.
Materials and methods
Collection of milk samples from participating herds
Farms participating in the study were recruited in collaboration with the Arla Foods Member Service Division. For practical reasons, only herds using milking parlours or tie-stall systems were invited to participate. The invited herds were located in the Swedish counties of Uppland, Södermanland, Västmanland and Västerbotten and included cows of the two most common dairy breeds in Sweden: SH and SRB. Although the study aimed to collect similar numbers of milk samples from both breeds, this was not fully achieved. Of the 12 voluntary farms, eight herds comprised only SRB cows, two herds comprised only SH cows and one herd included cows of both breeds.
Milk samples were collected from individual cows on these farms between September 22 and November 10, 2020. Within participating herds, cow selection was based on breed, stage of lactation and lactation number. In total, milk samples were obtained from 109 dairy cows, of which 20 were SH and 89 were SRB. All cows were in mid- to late lactation, defined as at least 8 weeks after the most recent calving and no more than 12 weeks before the next planned calving. All milk samples were collected during evening milking.
Fresh milk samples were transported at +4°C to the Department of Molecular Sciences, Swedish University of Agricultural Sciences (SLU), Uppsala, for analysis of gross milk composition, milk protein profile and proteolytic activity, with a focus on PL activity. Subsequently, milk samples were categorized into three groups according to lactation number: lactations 1–2 (N = 56), lactations 3–4 (N = 32) and lactations ≥5 (N = 21). Within these groups, the numbers of SRB cows were 46, 24 and 19, respectively, while the corresponding numbers of SH cows were 10, 8 and 2.
Milk sample preparation
Whole fresh milk was used for the analysis of pH and gross milk composition. For analyses of PL-PG activity and milk protein profile, milk was defatted by centrifugation at 1,864 × g for 10 min at 4°C (Sorvall Super T21, Sorvall Products L.P., Newton, CT, USA). The defatted milk samples were stored at −20°C until analysis.
Analysis of milk pH and gross composition
Milk pH was measured using a pH meter (Seven Compact S210, Mettler-Toledo, Stockholm, Sweden) after equilibration of fresh milk samples at room temperature for 1 h. Analysis of gross milk composition was performed at the Department of Applied Animal Science and Welfare, SLU, Uppsala. Total fat, protein, lactose, total solids, saturated fatty acids, unsaturated fatty acids, monounsaturated fatty acids, polyunsaturated fatty acids, myristic acid (C14:0), palmitic acid (C16:0), stearic acid (C18:0) and oleic acid (C18:1c9) were determined by mid-infrared spectroscopy (Fourier Transform Infrared, FOSS, Hilleröd, Denmark). SCC was determined by flow cytometry using a Delta Instruments CombiScope™ FTIR 600HP (PerkinElmer, Springfield, IL, USA).
PL and PG-derived activity
PL and PG-derived activities in milk samples were measured according to Korycka-Dahl et al. (Reference Korycka-Dahl, Ribadeau-Dumas, Chene and Marta1983), as modified by De Vries et al. (Reference De Vries, Brandt, Lundh, Holtenius, Hettinga and Johansson2016). After thawing, defatted milk samples were analysed in duplicate using 96-well microplates (Sarstedt, Nümbrecht, Germany). Briefly, PL and PG were dissociated from casein micelles by incubating defatted milk with ε-amino-n-caproic acid, followed by ultracentrifugation at 100,000 × g for 1 h at +4°C (Optima MAX-XP, Beckman Coulter, Inc., Bromma, Sweden).
PL activity was determined in the resulting milk serum using a chromogenic substrate (2.5 mg/ml), pyro-Glu-Phe-Lys-p-nitroanilide hydrochloride (Biophen CS-41(03), Hyphen BioMed, Neuville-sur-Oise, France). To determine total proteolytic activity, PG, the inactive precursor, was activated to PL by the addition of urokinase (49.5 Plough units). Both PL activity and total activity (PL + PG-derived activity) were measured at 37°C using a multimode microplate reader (POLARstar Omega, BMG Labtech, Ortenberg, Germany). Absorbance at 405 nm was recorded every 3 min for 120 min, and enzyme activity was expressed as the change in absorbance per unit time (ΔA405/Δt). Data were evaluated using Omega Data Analysis software (version 5.50 R4).
PG-derived activity was calculated as the difference between total activity and PL activity. The PG/PL was calculated by dividing PG-derived activity by PL activity.
Milk protein profiles
Milk protein separation was performed using a 7100 capillary electrophoresis (CE) system (Agilent Technologies Co., Santa Clara, CA, USA) equipped with an uncoated fused-silica capillary, as described by Johansson et al. (Reference Johansson, Åkerstedt, Li, Zamaratskaia and Sternesjö Lundh2013). Briefly, separations were conducted after the addition of 0.017 M d,l-dithiothreitol to the sample buffer on the day of sample preparation to reduce disulphide bonds in milk proteins. Milk (300 µl) was mixed with sample buffer (700 µl), incubated at room temperature for 1 h and subsequently defatted by centrifugation at 10,000 × g for 10 min (Hitachi-Himac CT 15R, Tokyo, Japan).
Protein detection was performed by UV–visible absorbance at 214 nm. Relative concentrations of individual milk proteins were calculated based on peak area and expressed as a percentage of the total integrated peak area in the electropherogram using Agilent 7100 CE software (Rev. C01.08 [210]).
Statistical analyses
Milk quality traits were analysed using linear mixed models. Because the distributions of the response variables were right-skewed, each variable was transformed using a base-10 logarithm prior to analysis. The models were fitted in SAS 9.4 (SAS Institute Inc., Cary, NC USA) using the PROC MIXED procedure. For each milk quality trait, the following model structure was used: fixed effects of breed (SH, SRB) and lactations (1–2, 3–4 and ≥5) and random effect of farm (1–12), to account for farm-level clustering of cows. The effect of breed was removed from the final analysis, because it did not contribute significantly to the model. The final model was, therefore, specified as:
 \begin{equation*}{\text{y}_{\text{ij}}}\, = \,{\text{lo}}{{\text{g}}_{10}}\,\left( {{\text{X}_{\text{ij}}}} \right) = \mu + {\text{lactatio}}{{\text{n}}_\text{i}}\, + {\text{far}}{{\text{m}}_\text{j}}\, + {\varepsilon _{\text{ij}}},\end{equation*}
\begin{equation*}{\text{y}_{\text{ij}}}\, = \,{\text{lo}}{{\text{g}}_{10}}\,\left( {{\text{X}_{\text{ij}}}} \right) = \mu + {\text{lactatio}}{{\text{n}}_\text{i}}\, + {\text{far}}{{\text{m}}_\text{j}}\, + {\varepsilon _{\text{ij}}},\end{equation*}where Xij is the untransformed milk quality trait, and yij is its log10-transformed value. μ is the general mean and εij is the random residual. Degrees of freedom were calculated using the Kenward–Roger method. Least squares means (LSMeans) for lactation were extracted on the log10-scale together with their 95% confidence intervals. Pairwise comparisons among lactation levels were adjusted using Tukey’s method. To aid interpretation, LSMeans and confidence limits were back-transformed to the original measurement scale. Boxplots illustrating the distributions of PL activity, PG activity and the PG/PL across lactation groups and breeds were generated using PROC SGPLOT. Associations between milk protein fractions, SCC and components of the PL system were assessed using Pearson’s correlation coefficients in PROC CORR. The correlations were calculated using pairwise deletion of missing values; therefore, the number of observations contributing to each correlation varied (N = 44–109). All variables were log10-transformed prior to analysis. A principal component analysis (PCA) (Wold et al., Reference Wold, Esbensen and Geladi1987) was performed among samples with complete protein fraction data (N = 33; 14 in lactations 1–2, N = 13 in lactations 3–4 and N = 6 in lactations ≥5) in PROC PRINCOMP to summarize covariation among milk quality traits and to identify variables associated with differences between lactation groups.
Results
Milk quality traits in relation to lactation number
Milk quality traits (Table 1) were compared among lactation groups (lactations 1–2, lactations 3–4 and lactations ≥5). Significant differences were observed for several traits. SCC was 52% lower in young cows (lactations 1–2) compared with cows in their fifth or higher lactation. The relative proportion of total casein was 4% higher in milk from cows in lactations 1–2 than in milk from cows in lactations ≥5. In contrast, total whey protein, α-lactalbumin and β-casein B were lower in milk from cows in lactations 1–2 compared with cows in lactations 3–4. The relative proportion of αs2-casein was highest in milk from cows in lactations ≥5, exceeding that observed in milk from cows in lactations 1–2 and 3–4. Total β-casein decreased progressively with an increasing number of lactations and was highest in lactations 1–2, intermediate in lactations 3–4 and lowest in lactations ≥5. Similarly, the relative concentration of κ-casein was higher in lactations ≥5 compared with lactations 1–2 and 3–4. PL activity was lower in lactations 1–2 than in lactations 3–4 and ≥5, whereas the PG/PL was higher in lactations 1–2 compared with the older lactation groups.
Comparison of milk quality traits of milk samples from cows grouped according to their number of lactations, i.e., lactations 1–2, lactations 3–4 and lactations ≥5

SFA, saturated fatty acid; UFA, unsaturated fatty acid; MUFA, monounsaturated fatty acid; PUFA, polyunsaturated fatty acid; SCC, somatic cell count.
Data were log10-transformed and are presented as back-transformed geometric means with their 95% confidence intervals (CIs) and P-value.
Means within a row with different superscripts (a–c) differ (P < 0.05, in bold text).
1 Lactations 1–2: n = 51; lactations 3–4: n = 29; lactations ≥5: n = 19.
2 Individual proteins expressed as % of total integrated area in the chromatograms.
Variation in milk quality traits associated with PL and PG-derived activity
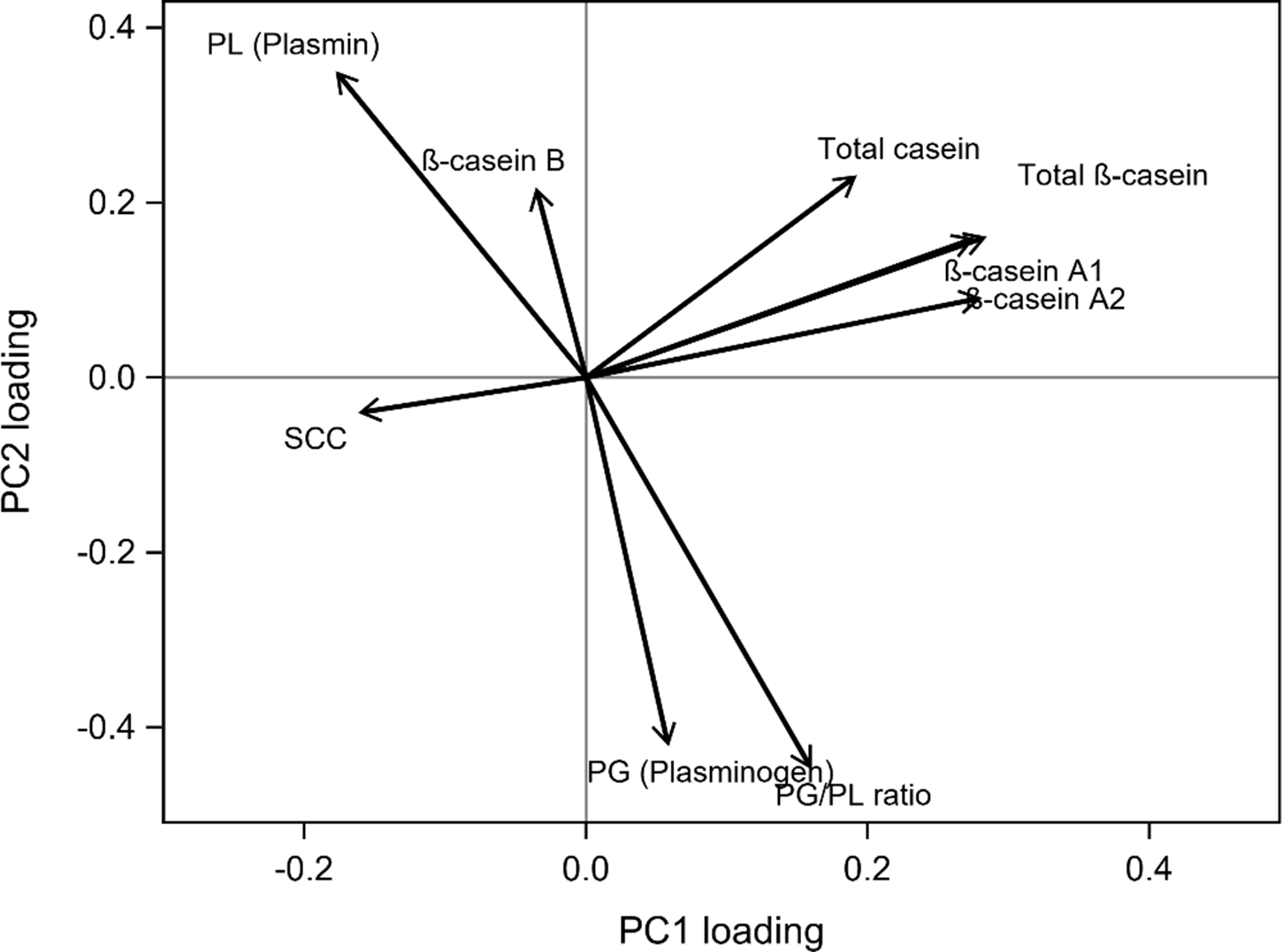
The explorative PCA revealed that the first principal component (PC1) explained 44% of the total variance, while the second principal component (PC2) explained an additional 24%. The analysis was conducted on complete observations only, resulting in a reduced sample size (N = 33). Despite this, the PCA captured a large proportion of the total variance, with PC1 and PC2 together explaining 68% of the variation, suggesting that the major sources of variation were robustly represented (Fig. 1). The variables considered for the model were PL, PG, PG/PL, total casein, total β-casein, β-casein A1 and A2, β-casein B and SCC. Each dot in the score plot represents a milk sample, with the colour indicating the lactation number of the corresponding cow. The PCA score plot revealed a moderate separation of samples by lactation group, with cows in lactations 1–2 and ≥5 showing the most distinct distributions along PC1. The intermediate group (lactations 3–4) overlapped with both groups, consistent with its transitional biological status. The PCA loading plot showed that PC1 was primarily driven by coordinated variation in casein-related traits, with total casein and β-casein fractions loading positively and opposing variables related to proteolytic activity, indicating a dominant gradient between casein integrity and proteolysis (Fig. 2).
Principal component analysis (PCA) score plot of milk samples based on log10-transformed milk protein fractions, somatic cell count and components of the plasmin system. Individual observations are shown as symbols according to lactation number, circles represent lactations 1–2, squares represent lactations 3–4 and triangles represent lactations ≥5. N = 33, i.e., samples with complete protein fraction data. Ellipses represent 95% confidence regions around group means. The separation occurs primarily along PC1, which explains 44.3% of the total variance. PC2 explains an additional 24.1%.

Loading plot from the principal component analysis (PCA) showing the contribution of milk quality traits to the first two principal components (PC1 and PC2). Arrows represent variable loadings, where arrow length indicates the strength of each variable’s contribution and arrow direction indicates its association with a given component. PC1 explains 44.3% of the total variance and is primarily driven by coordinated variation in casein-related traits, such as total casein and β-casein fractions, contrasted by variables associated with proteolytic activity. PC2 explains 24.1% of the total variance, reflecting secondary variation associated with the balance between plasmin activity and plasminogen-related measures (N = 33). SCC, somatic cell count.

Correlations between milk quality traits
Among all cows, strong positive correlations were observed among β-casein fractions (Table 2). In particular, β-casein A1 and A2 (r = 0.80, P < 0.001), and total β-casein and total casein content were strongly associated (r = 0.81, P < 0.001). PL activity was strongly and negatively correlated with PG/PL (r = −0.68, P < 0.001), whereas PG concentration was strongly positively correlated with PG/PL (r = 0.76, P < 0.001), in agreement with expectations. A significant, although weak, negative correlation was observed between SCC and total β-casein (r = −0.23, P < 0.05), indicating a reduction in intact casein with increasing somatic cell levels.
Pearson correlation among milk quality traits for milk samples from all cows and grouped according to number of lactations, irrespective of breed

PL, plasmin; PG, plasminogen; PG/PL, plasminogen-to-plasmin ratio; SCC, somatic cell count; Tot, total; Tot cas, total casein; NS, not significant; N, number of individual cows.
Only milk quality traits relevant for the proteolytic activity in milk (plasmin, plasminogen, plasminogen-to-plasmin ratio, total casein, total β-casein, β-casein A1, β-casein A2, β-casein B and SCC) were included in the analysis. Only significant results are shown.
All variables were log10-transformed prior to analysis. Correlations were calculated separately within lactation groups. Statistical significance was assessed at P < 0.05 (***P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05).
In cows within lactations 1–2, β-casein fractions were strongly inter-correlated, with a particularly strong association between β-casein A1 and A2 (r = 0.79, P < 0.001). Total β-casein (r = 0.73, P < 0.001) and β-casein A2 (r = 0.42, P < 0.01) were positively correlated with total casein content. No significant correlations were observed between PL, PG or SCC and casein fractions, suggesting limited proteolytic influence in young cows. The expected inverse relationship between PL and PG/PL was already evident (r = −0.66, P < 0.001), indicating functional regulation within the PL system.
In cows in lactations 3–4, correlations between casein fractions and markers of proteolysis became more pronounced. Total β-casein was negatively correlated with SCC (r = −0.47; P < 0.05), indicating increasing degradation or loss of casein with elevated cell counts. A very strong negative correlation was observed between PL and PG/PL (r = −0.88, P < 0.001), while PG remained positively associated with PG/PL (r = 0.54, P = 0.01).
In older cows (lactations ≥5), correlations indicated age-related changes in milk protein integrity. β-Casein A2 showed a positive correlation with total casein content (r = 0.49, P < 0.05), suggesting altered casein composition rather than simple loss of protein.
PL system components remained strongly interrelated. PG and PG/PL were very strongly positively correlated (r = 0.86, P < 0.001), while PL was negatively correlated with PG/PL (r = −0.46, P < 0.05). No significant correlations were observed between SCC and PL, indicating that proteolytic changes in older cows were not solely driven by inflammatory status.
Pairwise correlations among milk quality traits are summarized in heat maps for all cows and stratified by lactation group (Fig. 3a–d), illustrating both the strength and direction of associations.
(a–d) Heat maps of pairwise Pearson correlation coefficients (r) among selected milk quality traits within lactation groups. Panels show (a) lactations 1–2, (b) lactations 3–4, (c) lactations ≥5 and (d) all cows. Colours indicate correlation direction and magnitude (red = positive, blue = negative), and values within cells represent r. Correlations were calculated using pairwise deletion of missing values. Due to missing values, the number of observations contributing to individual correlations varied across variable pairs (N = 44–109).

Overall, the correlation analysis revealed a consistent internal structure among milk casein fractions across parities, while associations between caseins, SCC and the PL system became increasingly pronounced with advancing lactation, indicating age-related alterations in milk proteolysis.
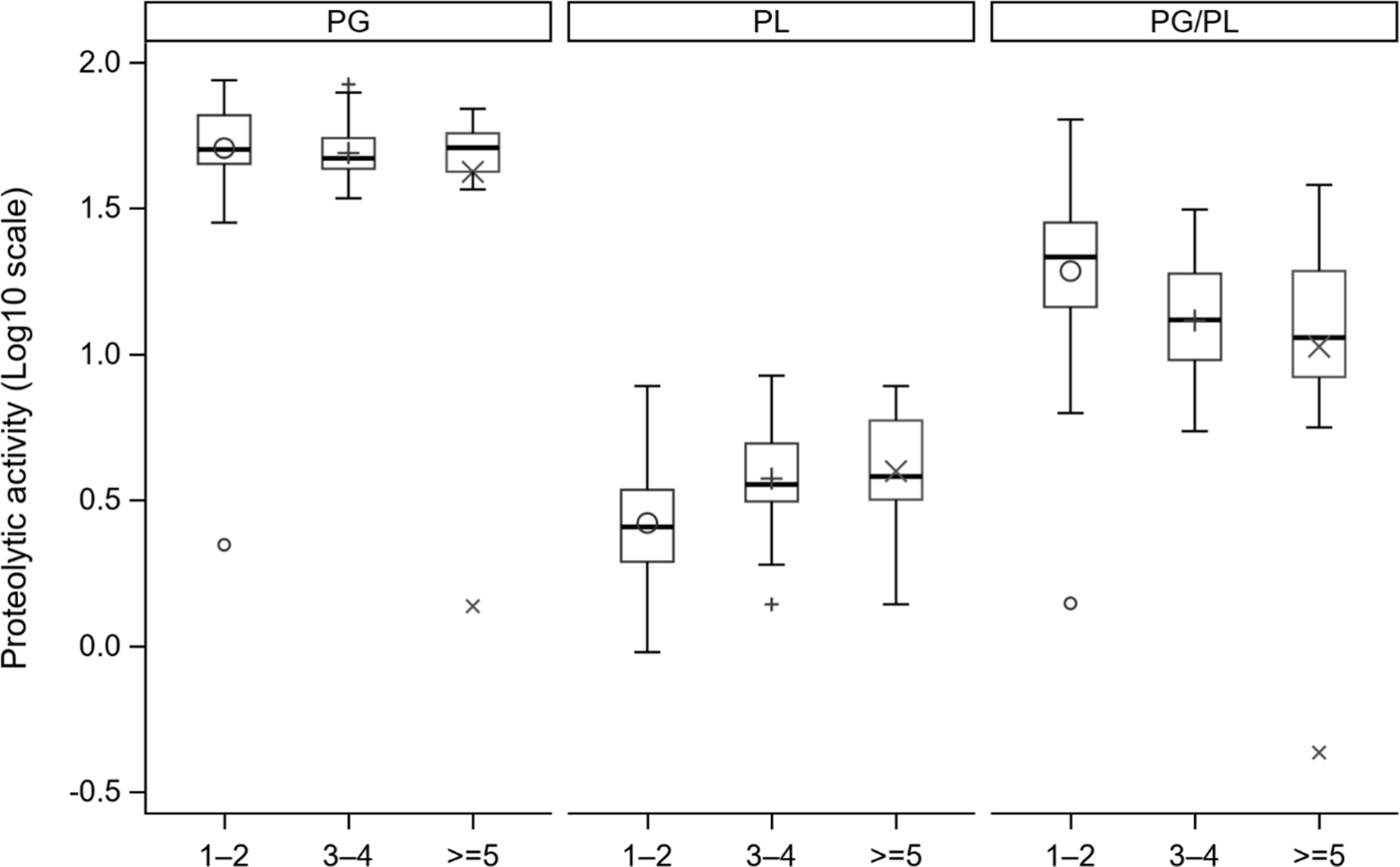
The distribution of proteolytic activity differed across lactation groups, shown in boxplots (Fig. 4). PL activity showed an increasing median value with increasing number of lactations. In contrast, PG levels exhibited a more stable distribution across parities, with only minor shifts in median values.
Boxplots showing the distribution of plasminogen (PG) and plasmin (PL) activity (log10-transformed) in milk samples from cows of different lactations: 1–2, 3–4 and ≥5. Boxes represent the interquartile range with the median indicated by the horizontal line. The mean is indicated by symbols according to lactation number: circles for lactations 1–2, plus signs for lactations 3–4 and × for lactations ≥5. Whiskers represent the minimum and maximum values. Symbols located outside whiskers are outliers.

Discussion
This study investigated differences in gross milk composition, milk protein profile, SCC and PL-related proteolytic activity in milk from individual dairy cows differing in lactation number. Particular emphasis was placed on the PL-PG system and its associations with SCC and milk protein composition.
Although the average SCC was below 300,000 cells/ml in all lactation groups, cows in lactations ≥5 exhibited significantly higher SCC than cows in their first or second lactation. Lower SCC in younger cows has previously been associated with lower milk yield and reduced mammary epithelial cell turnover (Saravanan et al., Reference Saravanan, Das, De and Panneerselvam2015; Gonçalves et al., Reference Gonçalves, Cue, Botaro, Horst, Valloto and Santos2018). In agreement with the present findings, numerous studies have reported a positive association between lactation number and SCC (Blackburn, Reference Blackburn1966; Ng-Kwai-Hang et al., Reference Ng-Kwai-Hang, Hayes, Moxley and Monardes1984; Alhussien and Dang, Reference Alhussien and Dang2018b; Stocco et al., Reference Stocco, Cipolat-Gotet, Stefanon, Zecconi, Francescutti, Mountricha and Summer2023). Because SCC is widely used as an indirect indicator of mammary gland health, this relationship has often been interpreted as reflecting poorer udder health in older cows.
However, earlier studies suggest that elevated SCC in older cows may not necessarily indicate ongoing infection. Sheldrake et al. (Reference Sheldrake, Hoare and McGregor1983) reported that lactation number per se did not affect SCC when infection status at the quarter level was considered and concluded that higher SCC in older cows likely reflected the cumulative effects of previous intramammary infections. Similarly, Natzke et al. (Reference Natzke, Everett and Postle1972) demonstrated that cows remaining uninfected for extended periods could maintain low SCC even at advanced ages. In the present study, cows in lactations 1–2 had lower SCC than older cows, consistent with the findings of Alhussien and Dang (Reference Alhussien and Dang2018a), who observed stable SCC up to the fourth lactation followed by a significant increase in cows with more than four parities. However, as the differential SCC was not performed in this study, we could not determine whether the elevated SCC observed in older cows was driven by changes in specific leukocyte or epithelial cell fractions, which may influence the interpretation of age-related differences.
PL is the major indigenous protease in bovine milk and plays a dual role in dairy processing, with both beneficial and detrimental effects depending on product type and processing conditions (France et al., Reference France, O'Mahony, Kelly, Kelly and Larsen2021). The present results revealed clear differences in milk quality traits associated with PL and PG activity across lactation groups. Milk from cows in lactations 1–2 exhibited lower PL activity, lower SCC and lower relative concentrations of β-casein B but a higher PG/PL and higher relative total casein content compared with milk from older cows.
Both PL and PG activity levels observed in this study were lower than those reported in previous investigations of pooled milk samples from young and older cows (Johansson et al., Reference Johansson, Lindberg and Lundh2024) and from dairy silo milk (Karlsson et al., Reference Karlsson, Langton, Innings, Wikström and Lundh2017). This discrepancy may be explained by the compositional averaging inherent in bulk or silo milk, which represents milk from numerous cows across a wide range of lactation numbers and physiological states. In individual cows, PG migrates from blood into the mammary gland, where it can be activated to PL in the presence of appropriate activators, e.g., cell-derived urokinase-type PG activator (Korycka-Dahl et al., Reference Korycka-Dahl, Ribadeau-Dumas, Chene and Marta1983). Consequently, lower PL activity in milk from cows in lactations 1–2 likely reflects both lower SCC and reduced PG activation. However, a limitation of this study is the lack of precise days in milk data and the restriction to cows classified as mid- to late lactation. This reduces generalizability and does not allow assessment of early-lactation dynamics, where PL activity is known to be more variable (Nicholas et al., Reference Nicholas, Auldist, Molan, Stelwagen and Prosser2002).
Previous studies have similarly reported lower PL activity in early lactations compared with later parities (Politis, Reference Politis1996). Politis (Reference Politis1996) demonstrated that milk from the first three lactations exhibited significantly lower PL activity than milk from higher lactations and attributed this variation in part to differences in SCC. Given that PL preferentially hydrolyses caseins, particularly β-casein (Ismail and Nielsen, Reference Ismail and Nielsen2010), the higher relative β-casein content observed in milk from cows in lactations 1–2 is consistent with their lower PL activity.
Increased PL activity has previously been associated with advancing lactation number and SCC levels exceeding 300,000 cells/ml (Politis et al., Reference Politis, Lachance, Block and Turner1989; Bastian et al., Reference Bastian, Brown and Ernstrom1991; Mazal et al., Reference Mazal, Vianna, Santos and Gigante2007). The biological role of PL in milk is complex, as its activity may reflect mammary epithelial turnover, involution and tissue remodelling (Politis, Reference Politis1996). Two principal mechanisms contribute to increased PL activity in milk: increased transfer of PL from blood and enhanced activation of PG to PL within the mammary gland (Politis et al., Reference Politis, Lachance, Block and Turner1989). The PG/PL provides insight into the latter mechanism.
In the present study, the PG/PL was highest in milk from cows in lactations 1–2 and decreased with increasing lactation number, indicating greater activation of PG to PL in older cows. This interpretation is supported by the strong negative correlation between PG/PL and PL activity across all lactation groups. Elevated PL activity in milk from older cows may, therefore, be associated with increased mammary epithelial cell turnover, as suggested by De Vries et al. (Reference De Vries, Brandt, Lundh, Holtenius, Hettinga and Johansson2016), and aligns with the mechanistic lactation model proposed by Dijkstra et al. (Reference Dijkstra, Lopez, Bannink, Dhanoa, Kebreab, Odongo, Fathi Nasri, Behera, Hernandez-Ferrer and France2010), which predicts higher secretory cell death rates in multiparous cows.
The partial overlap observed between lactation groups in the PCA indicates gradual, rather than discrete, changes in milk composition with increasing lactation number. Nevertheless, the consistent separation of group centroids and the structure of the confidence ellipses suggest that the observed patterns reflect biologically meaningful variation associated with parity.
Conclusions
PL activity was lower in milk from cows in lactations 1 and 2 than in milk from older cows. Differences between parities were also observed for SCC, PG activation and milk protein composition. Although extending cow longevity beyond current averages may lead to moderately increased PL activity in milk, this is unlikely to have practical significance in herds with good udder health. Elevated PL activity may be of relevance for ultra-high-temperature processed products, whereas effects on fresh dairy products and cheese manufacture are expected to be limited. Furthermore, bulk milk at the farm level represents a mixture of milk from cows in multiple lactations, thereby attenuating potential effects of higher PL activity in milk from older individuals. Overall, the results indicate that extending the productive lifespan of dairy cows in healthy herds does not compromise milk quality.
Acknowledgements
The authors thank all dairy farmers who participated in this study. We also acknowledge Emma Gratte and Lena Loftäng from the Arla Foods Member Service Division for their assistance in recruiting farms for participation. This study was financially supported by the Swedish Farmers’ Foundation for Agricultural Research (Stockholm, Sweden; grant number R-18-26-131).
Use of artificial intelligence (AI) tools
During the preparation of this manuscript, OpenAI ChatGPT (version 5.2) was used for language and grammar editing. The authors reviewed and revised the manuscript and take full responsibility for the content of the publication.





 Open access
Open access