


Introduction
Donkeys ( Equus asinus ) are slaughtered across the world for a variety of reasons, including their meat and by-products, particularly their skins, which are used to make ‘eijao’, a product used in Traditional Chinese Medicine (Gameiro et al. Reference Gameiro, Rezende and Zanella2021; Norris et al. Reference Norris, Little, Ryding and Raw2021). Despite increasing rates of donkey slaughter (Bennett & Pfuderer Reference Bennett and Pfuderer2019; Gameiro et al. Reference Gameiro, Rezende and Zanella2021), very little research has been conducted into the methods used or the factors affecting their effectiveness and the impact on welfare (Fletcher et al. Reference Fletcher, Limon, Whatford, Grist, Knowles and Gibson2022, Reference Fletcher, Limon, Agongo, Akunzule, Essel, Padalino, Grist and Gibson2024).
Penetrating Captive Bolt (PCB) is one of the most common methods used for the stunning of livestock prior to slaughter (Oliveira et al. Reference Oliveira, Dalla Costa, Gibson, Dalla Costa, Coldebella and Gregory2018). Unconsciousness is induced through the combination of the transference of the kinetic energy of the moving metal bolt to the cranium and brain, and the direct physical trauma to critical brain structures (Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Baier & Willson Reference Baier and Willson2020). The extent of focal and diffuse damage to brain structures depends largely on placement and orientation of the bolt into the brain (Terlouw et al. Reference Terlouw, Bourguet and Deiss2016). The highly referred to positioning for PCB slaughter of horses is 10 mm above the intersection of two imaginary lines drawn from the centre middle of the ears and the inside of the eye on the opposite side (Humane Slaughter Association 2016). No such guidance exists for donkeys, however anecdotally most operators tend to use the same position (K Fletcher, personal communication and prior field observation 2023, 2024).
Efficacy and the welfare of animals during PCB stunning has been examined via a variety of indices, including behaviour observations (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Terlouw Reference Terlouw2020; Fletcher et al. Reference Fletcher, Limon, Agongo, Akunzule, Essel, Padalino, Grist and Gibson2024), changes in the electroencephalogram (EEG) (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Gibson et al. Reference Gibson, Oliveira, Dalla Costa and Gregory2019; Dalla Costa et al. Reference Dalla Costa, Gibson, Oliveira, Gregory, Coldebella, Faucitano, Ludtke, Peréirã Buss and Dalla Costa2020), evoked potentials (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015) and trauma to associated brain structures (Dalla Costa et al. Reference Dalla Costa, Gibson, Oliveira, Gregory, Coldebella, Faucitano, Ludtke, Peréirã Buss and Dalla Costa2020). Unlike behavioural and brainstem indices of brain activity, the EEG represents the direct functional activity of the brain and is considered a more reliable indicator of when undoubted unconsciousness occurs (Gibson et al. Reference Gibson, Oliveira, Dalla Costa and Gregory2019).
EEG provides the real-time functional activity of the brain and can be used to complement indirect behavioural indicators after stunning at slaughter (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Gibson et al. Reference Gibson, Oliveira, Dalla Costa and Gregory2019; Dalla Costa et al. Reference Dalla Costa, Gibson, Oliveira, Gregory, Coldebella, Faucitano, Ludtke, Peréirã Buss and Dalla Costa2020; Kumar et al. Reference Kumar, Abubakar, Imlan, Ahmed, Goh, Kaka, Idrus and Sazili2023). As the level of unconsciousness deepens, there is a reduction in total power (Ptot) EEG (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015) and/or a frequency shift to a predominance of low frequency activity. A conscious animal showing ‘normal-like’ (or ‘baseline’) EEG will typically display a combination of low, mid and high frequency activity, with alpha (8 to 12 Hz) waves indicating relaxation and beta (12–30 Hz) indicating a stressed or fearful state (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Kumar et al. Reference Kumar, Abubakar, Imlan, Ahmed, Goh, Kaka, Idrus and Sazili2023). In TBI-induced unconsciousness, animals generally display predominantly high-amplitude, low-frequency ‘transitional’ delta (0.5 to 4 Hz) and theta (4 to 8 Hz) with an absence of alpha or beta activity. These can be recoverable, but an animal is considered to have irrecoverable unconsciousness, or brain death, when an isoelectric waveform is present (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Grandin Reference Grandin, Grandin and Cockram2020; Kumar et al. Reference Kumar, Abubakar, Imlan, Ahmed, Goh, Kaka, Idrus and Sazili2023).
Donkeys are considered stoic and subtle in their behavioural display (Ashley et al. Reference Ashley, Waterman-Pearson and Whay2005), which could lead to misunderstanding or misinterpretation when being assessed during the slaughter process. Therefore, in a research context, the combination of EEG and behavioural indices could provide a more reliable, robust and accurate assessment of welfare and stun effectiveness compared to assessment of behaviour alone. EEG has been successfully used to measure and validate stunning efficiency in other animals at slaughter (Zulkifli et al. Reference Zulkifli, Goh, Norbaiyah, Sazili, Lotfi, Soleimani and Small2014; Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2016; Gibson et al. Reference Gibson, Oliveira, Dalla Costa and Gregory2019). However, to date, there have been no studies that have used EEG activity to evaluate the loss of consciousness of donkeys at slaughter.
The aim of this study was to examine the EEG responses of commercially slaughtered donkeys to PCB stunning and determine how these responses were related to assessed behavioural/brainstem reflexes.
Materials and methods
The study was carried out over six days in April 2024, during routine commercial stunning and slaughter of donkeys (n = 13) at a Mexican abattoir. This project had ethical approval from the Royal Veterinary College Clinical Research Ethical Review Board (URN 2024 2268-3).
Donkeys arrived at the abattoir either on the same day or the day before slaughter. Information about the sex, age, background/origin of the animals and the transport length was not available due to restricted information at the abattoir. The donkeys were slaughtered in the same slaughter line and stun box as the horses and cattle. All animals entered a standard single-file straight raceway and were then individually led, through a manually opened gate, into a cattle stun box. Animals were stunned by multiple commercial operators; it was not possible to measure operator effect due to sensitivity within the abattoir. Convenience sampling was used based on order of animals throughout the day and was dependent upon permission/consent granted by the abattoir and/or suppliers. Prior to, and during, stunning, donkeys were physically restrained with a head/neck rope held by an operator to allow for placement of EEG electrodes for pre-treatment recording. Instrumentation took between 10–20 s, followed by a minimum of 30 s of pre-treatment EEG recorded. Traces were monitored to ensure at least 20 s of continuous stable EEG recorded prior to the shot. After pre-treatment recording, donkeys were shot in a frontal position with a Jarvis USSS-1 penetrating pneumatic stunner (USSS-1, JARVIS® Jarvis Products Corporation; Middletown, CT, USA). Penetrating captive-bolt airline pressure and bolt velocity could not be recorded as there were no in-line meters/regulators. The compressor tank air pressure was set to 120 psi (827 kPa). One minute after the shot, donkeys were released, via a second manually operated gate, into the bleeding area. Prior to bleeding, each donkey was ear-tagged to allow head identification, post mortem.
Behavioural assessment
Immediately after shooting, each donkey was assessed for signs of effectiveness of stunning by three members of the research team from different positions (side of the stun box, front of the stun box, and in the roll-out area). All three observers remained consistent in their individual positions, assessing animals at different points with constant communication between the team to confirm observations. In particular, the presence or absence of immediate collapse, rhythmic breathing, palpebral and corneal reflex, spontaneous eye blinking, eyeball rotation, nystagmus, gasping, response to stimulation of the septum (with forceps), tonic and clonic convulsions were evaluated after the PCB shot until shackling and hoisting for exsanguination (Gibson et al. Reference Gibson, Bedford, Chancellor and Limon2015; Fletcher et al. Reference Fletcher, Limon, Agongo, Akunzule, Essel, Padalino, Grist and Gibson2024) (Table 1).
Brainstem and behavioural signs of consciousness assessed immediately post pneumatic penetrating captive bolt (PCB) shot in donkeys (n = 13) in a Mexican abattoir (adapted from Gibson et al. Reference Gibson, Bedford, Chancellor and Limon2015)

Table 1. Long description
From the top row downward, the left column lists behaviours and cranial or spinal responses, while the right column provides descriptions. The entries are: Lack of immediate collapse, described as the animal failing to collapse immediately after the shot; Righting reflex, making a coordinated effort to stand or lift head; Vocalisations, vocalising independently from exhalation; Rhythmic respiration, ribcage moving in and out rhythmically; Gasping, spasmodic sharp intake of breath with mouth open; Corneal reflex, involuntary blink reflex when cornea is stimulated; Palpebral reflex, involuntary blink reflex when the medial canthus is stimulated; Spontaneous blinking, opening or closing eyelid without stimulation; Nystagmus, rapid involuntary movements of the eye; Eyeball rotation, eyes rotated, not central, sclera visible; Response to nostril pinch, pinching nasal septum followed by pain reaction or withdrawal; Involuntary muscle spasms, absence of tonus in body or excessive muscle activity.
Donkeys were classified as incompletely concussed if they failed to collapse or rhythmic breathing was present and/or if at least two of the following parameters were present: positive corneal reflex; positive palpebral reflex; eyeball rotation; and nystagmus (Gibson et al. Reference Gibson, Bedford, Chancellor and Limon2015). Behaviours and brainstem indices were sampled continuously from immediately after the shot (including the EEG recording period, EEG electrode removal and roll-out), with observations continuing until animals were shackled and hoisted prior to exsanguination.
Observations were recorded using a Dictaphone (Olympus VN-713PC, Olympus, Hachioji-shi, Tokyo, Japan) and headset (Sennheiser PC2, Sennheiser electronic GmbH & Co, Wedemark, Germany).
Electroencephalographic recording and assessment
Electroencephalographic data were recorded using three 27-gauge stainless steel subdermal electrodes (Neuroline Subdermal, Ambu Inc, Glen Burnie, MD, USA), placed as a three-electrode montage in the skin with the: active (non-inverting) right of midline in-line with the back of the eyes; reference (inverting), over the right caudal aspect of the frontal bone (top of head) just rostral of the front of the ears; and ground electrode caudal to the poll on the right. EEG signals were amplified and filtered with an analog filter (Dual Bio Amp, ADInstruments Ltd, Sydney, Australia) with low- and high-pass filters of 100 and 0.1 Hz, respectively. The signals were digitalised (1 kHz) with a 4/35 PowerLab (ADInstruments Ltd, Sydney, Australia) digital-to-analogue converter and recorded on an Apple MacBook Air (Apple Inc Cupertino, CA, USA) for off-line analysis. Interelectrode impedance was tested and ranged between 1.0 and 1.9 kΩ (MkIII Checktrode, UFI, Morro Bay, CA, USA). Each animal acted as their own control with comparisons made between pre- (PRE; 20 s) and post-treatment (POST; 30 s) EEG waveforms. After the completion of EEG recording, the electrodes were removed and the animals ejected from the stunning box, shackled and hoisted and then bled in accordance with routine procedure of the abattoir.
EEG epochs contaminated by artefacts such as over- and under-scale (DC drift), large single spikes, or electromyography were manually rejected from analysis using Chart 8.1.24 (ADInstruments Ltd). All waveforms were digitally filtered with a passband of 1 to 30 Hz, and traces were inspected visually and compared with baseline using the classification systems developed by Gibson et al. (Reference Gibson, Johnson, Murrell, Mitchinson, Stafford and Mellor2009a,Reference Gibson, Johnson, Murrell, Hulls, Mitchinson, Stafford, Johnstone and Mellorb, Reference Gibson, Oliveira, Dalla Costa and Gregory2019). They were classified into one of five categories: (1) movement artefact; (2) normal-like EEG; (3) transitional EEG; (4) high amplitude low frequency (HALF) EEG; and (5) isoelectric EEG. Briefly, normal-like EEG represents an activity that is similar in amplitude and frequency to the baseline period. Transitional EEG was classified as suppressed activity of having either an amplitude of less than half of that of the pre-treatment EEG and/or depressed high-frequency activity. HALF EEG was classified as waveforms of predominantly high amplitude, low frequency activity based on morphology. Isoelectric EEG was classified as a trace with an amplitude of < 1/8 (12.25%) of that of normal-like pre-stunning EEG with little or no low-frequency components.
The EEG power spectra of uncontaminated epochs were analysed. Fast Fourier transformation with a Welch window was applied to non-overlapping 1-s epochs (PRE and every second POST), generating sequential power spectra with 1-Hz frequency bins. Spectral data contaminated by movement artefacts were excluded based on the subjective analysis.
EEG spectral data were calculated and are displayed as percentage changes in total power (Ptot), delta (0.5 to 4 Hz), theta (4 to 8 Hz), alpha (8 to 12 Hz), and beta (12 to 30 Hz) power from PRE values.
Shot position assessment
Donkeys’ heads were removed (the majority at the atlanto-occiptial joint but some were removed between the atlas [C1] and axis cervical vertebra [C2]) immediately post-bleeding and prior to skinning, and morphometric measurements taken post-skinning. Head length from the top of the poll to the tip of the nasal plane was measured, along with the width of the head from the widest point of each eye and the distance from shot position to the tip of the nasal plane. For each animals, the shot entry position on the cranium was determined as the deviation (lateral and rostro-caudal), from 10 mm above the intersection of lines (determined with placement of durapore tape [3M Company Maplewood, MN, USA]) from the middle of each eye to the base of the opposite ear (Humane Slaughter Association 2016). This position was recorded on transparent acetates placed over the head that was previously skinned.
Statistical analysis
Data were recorded and entered into Microsoft Excel® 2016 (Microsoft Corporation, Redmond, USA). SPSS (IBM® SPSS Statistics 28.0.0.0, 2022) and R version 4.3.1. (R Core Team 2023) within the RStudio environment were used for statistical analyses. Descriptive behavioural data were reported as proportion (percentage of animals showing behaviour). Distribution of numeric data were tested with the Shapiro-Wilk test. Normally distributed (EEG and shot deviation) data were analysed with paired t-test or repeated measures ANOVA to determine associations between variable factors (e.g. shot position/deviation) and outcome (EEG or behavioural signs); while non-normally distributed data were analysed with Mann-Whitney U or Friedmann tests, where appropriate. Sagittal and lateral deviations were reported as mean (± SD) and P ≤ 0.05 used as the indicator of significance, with trends indicated where P < 1.0.
Results
Electroencephalography (EEG)
Immediately after application of PCB all donkeys had periods of movement artefact in the EEG (Figure 1). The duration varied between donkeys with a mean (± SD) of 1.8 (± 1.1) s; range: 1–5 s. Seven donkeys were classed as completely concussed based on EEG data, in these animals movement artefact was followed by periods of HALF (8.0 [± 9.1]; range: 6–12 s) or transitional (mean duration 18.8 [± 9.1]; range: 11–23 s) EEG activity, before becoming isoelectric in five of the donkeys. The EEG of donkeys D9 and D11 remained transitional during the entire POST 30-s data recording period. No animal received a repeat shot attempt.
Characteristics of electroencephalogram (EEG) waveforms in individual donkeys (time-point 0) shot with Penetrating Captive Bolt (PCB). White bars represent movement artefact; light grey transitional EEG; dark grey isoelectric EEG; cross-hatched high amplitude low frequency (HALF), and black bars representing normal-like active EEG. Numbers represent duration of the EEG states (s).

Figure 1 Long description
Horizontally stacked barchart showing characteristics of electroencephalogram (EEG) waveforms in individual donkeys (time- point 0) shot with Penetrating Captive Bolt (PCB). White bars represent movement artefact; light grey transitional EEG; dark grey isoelectric EEG; cross- hatched high amplitude low frequency (HALF), and black bars representing normal-like active EEG. Numbers represent duration of the EEG states (s).
Six donkeys (46%; 6/13) had periods of apparently normal-like EEG activity after application of PCB, this period was confined to the initial 10 s post-shooting, before reverting to either HALF or transitional activity. The mean duration of normal-like EEG activity was 4.8 (± 2.6); range: 1–9 s. In four (66%; 4/6) of these animals, the EEG eventually became isoelectric before the end of the data recording period. The EEG of donkeys D1 and D12 was transitional by the end of the 30-s data recording period.
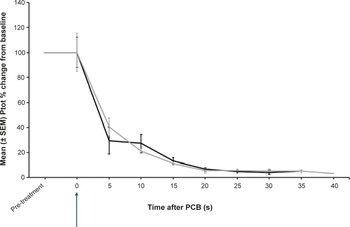
After PCB, there was a significant decrease in Ptot, as a percentage change from pre-treatment values (P = 0.002) (Figure 2). There was no significant difference in Ptot after 5 s between donkeys that were assessed as incomplete or completely concussed (P = 1). In completely concussed donkeys, power in the beta frequency band significantly decreased from pre-treatment values within 5 s of PCB (P = 0.01) and remained depressed for the rest of the recording period. For frequency bands delta, theta and alpha there was a non-significant trend towards a decrease (P = 0.625) 5 s after PCB. Theta, alpha and beta power remained elevated in the initial 5–10 s after the PCB shot in animals that were incompletely concussed (Figure 3). However, this was momentary before reducing to similar levels as completely concussed animals, with no significant difference between the two groups (completely and incompletely concussed; all P-values were P > 0.05).
Mean (± SEM) total power (Ptot) of the electroencephalogram (EEG) of donkeys (n = 13) before and after being shot (time-point 0, blue arrow) with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir. The grey line represents donkeys (n = 6) that had periods of incomplete concussion and the black line those that were completely concussed (n = 7). Note this excludes periods of movement artefact for incomplete concussion (time-points 25 s, 35s; n = 5 and 40 s; n = 2) and complete concussion (time-points 5 s; n = 5 and 35; n = 3).

Figure 2 Long description
Line graph showing mean (± SEM) total power (Ptot) of the electroencephalogram (EEG) of donkeys (n = 13) before and after being shot (time- point 0, blue arrow) with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir (n = 13). The grey line represents donkeys (n = 6) that had periods of incomplete concussion (n = 6) and the black line those that were completely concussed (n = 7). Note this excludes periods of movement artefact for incomplete concussion (time-points 25 s, 35s; n = 5 and 40 s; n = 2) and complete concussion (time-points 5 s; n = 5 and 35; n = 3).
Mean (± SEM) power of (a) delta, (b) theta, (c) alpha and (d) beta frequency bands of the electroencephalogram (EEG) of donkeys (n = 13) before and after being shot (time-point 0, blue arrows) with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir. The grey line represents donkeys that had periods of incomplete concussion (n = 6) and the black line those that were completely concussed (n = 7). Note this excludes periods of movement artefact for incomplete concussion (time-points 25 s, 35s; n = 5 and 40 s; n = 2) and complete concussion (time-points 5 s; n = 5 and 35; n = 3).

Figure 3 Long description
Line graph showing mean (± SEM) power of delta (A), theta (B), alpha (C) and beta (D) frequency bands of the electroencephalogram (EEG) of donkeys (n = 13) before and after being shot (time-point 0, blue arrows) with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir (n = 13). The grey line represents donkeys that had periods of incomplete concussion (n = 6) and the black line those that were completely concussed (n = 7). Note this excludes periods of movement artefact for incomplete concussion (time--points 25 s, 35s; n = 5 and 40 s; n = 2) and complete concussion (time-points 5 s; n = 5 and 35; n = 3).
Shot position
The mean (± SD) deviation from the suggested shot position was 30.8 (± 13.1) mm rostral-caudal and 10.4 (± 6.5) mm lateral (Figure 4), with 92% (12/13) of shots caudal of this position. Mann-Whitney U tests found a significant relationship between the presence of normal-like EEG activity post-shot and rostral-caudal deviation (P = 0.02) but not lateral deviation (P = 0.73). Similarly, there was a significant relationship between behavioural/reflex signs of incomplete concussion and rostral-caudal deviation (P = 0.02) but not lateral (P = 0.22).
Shot entrance site relative to the Humane Slaughter Association (HSA 2016) suggested position for donkeys (0 mm) shot with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir (n = 13) based on the operator’s perspective (+ is left and caudal of the animal’s midline). Solid blue circles represent donkeys that were completely concussed based on electroencephalogram (EEG) data, while red crosses represent those incompletely concussed based on EEG.

Figure 4 Long description
Scatterplot showing shot entrance site relative to the Humane Slaughter Association (HSA 2016) suggested position for donkeys (0 mm) shot with a pneumatically powered Penetrating Captive Bolt (PCB) at a Mexican abattoir (n = 13) based on the operator’s perspective (+ is left and caudal of the animal’s midline). Solid blue circles represent donkeys that were completely concussed based on electroencephalogram (EEG) data, while red crosses represent those incompletely concussed based on EEG.
Behavioural/brainstem signs
One donkey (D6) was classified as incompletely concussed, showing both rhythmic respiration and spontaneous blinking, but was not re-shot (Table 2).
Behavioural and cranial/spinal responses (%) of donkeys (n = 13) shot with pneumatic penetrating captive bolt (PCB) in a Mexican abattoir

Table 2. Long description
Table outlining behavioural and cranial/spinal responses of donkeys (%) shot with pneumatic penetrating captive bolt (PCB) (n=13) in a Mexican abattoir.
* Some responses could not be recorded from all animals due to accessibility issues.
D1 had a shallow depth of concussion based on behavioural/brainstem signs, displaying both a palpebral reflex and spontaneous blinking. This donkey also displayed normal-like EEG activity for 9 s post-PCB but was not re-shot. Meanwhile, D5 showed normal-like EEG activity post-shot, was unable to have ocular reflexes checked due to accessibility issues but did show muscle spasms. Similarly, the ocular reflex could not be assessed for donkey D6, but this animal did have a period of normal-like EEG post-PCB and displayed both rhythmic respiration and spontaneous blinking. Donkey D8 had a short period of normal-like EEG activity (4 s) after PCB and was also observed to show gasping post-shot and briefly head shake laterally. Meanwhile, donkeys 12 and 13 had 1- and 4-s periods of normal-like EEG activity, respectively, but presented with no behavioural signs of incomplete concussion. Donkey D10 was the only completely concussed animal (based on EEG data), that did not display any behavioural/reflexes, except for gasping, which was agonal in nature.
Discussion
This study is the first to examine the EEG responses of donkeys to PCB stunning and explore how these responses related to behavioural signs indicative of brainstem activity. Assessment of EEG is generally considered the most objective, reliable way to assess unconsciousness and predict brain injury prognosis, compared with behavioural indicators (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Wang et al. Reference Wang, Huang, Ma, Zhao, Liu and Xu2021). EEG can complement and validate behavioural indicators after stunning to demonstrate the level of consciousness an animal is experiencing, whether they are showing ‘normal-like’ or baseline EEG, indicating consciousness, low-frequency ‘transitional’ EEG, indicating potentially recoverable unconsciousness, or isoelectric EEG, where consciousness is considered unrecoverable and the animal is brain dead (Verhoeven et al. Reference Verhoeven, Gerritzen, Hellebrekers and Kemp2015; Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Gibson et al. Reference Gibson, Oliveira, Dalla Costa and Gregory2019; Grandin Reference Grandin, Grandin and Cockram2020; Kumar et al. Reference Kumar, Abubakar, Imlan, Ahmed, Goh, Kaka, Idrus and Sazili2023).
In the study, no animals were determined to have maintained consciousness for the full duration of the recording period. However, six exhibited transient ‘normal-like’ EEG patterns for up to 9 s post-stunning, which subsequently evolved into transitional or high-amplitude low-frequency (HALF) waveforms. Notably, four animals did not reach an isoelectric EEG state, an indicator of irreversible brain dysfunction during the observation period. This raises concern that these animals could have potentially been at risk of regaining some form of consciousness prior to exsanguination. However, this could not be examined due to the recording period being limited to 30 s post-shot due to the operational requirements of the abattoir and to prevent suffering from recovery by ensuring prompt exsanguination. Together, these findings highlight the potential risks associated with PCB stunning of donkeys, particularly regarding the duration of time during which they might maintain consciousness, and the importance of the reduction of the stun-to-stick interval in donkeys to mitigate the return of consciousness in potentially reversibly stunned animals. Furthermore, this study sheds light on the fact that behavioural signs alone can mean that abattoir operators could fail to recognise the maintenance or recurrence of consciousness in some animals prior to exsanguination.
The Humane Slaughter Association (HSA 2016) suggests that exsanguination should occur a maximum of 60 s post-stun. However, the practicality of this being applied under commercial slaughter conditions, to allow for the time required for animals to be mechanically hoisted post-stun, prior to being bled, requires further research.
In this study, there was a high proportion (46%) of donkeys that had periods of normal-like EEG indicating incomplete concussion. Potential causes for this high failure rate could have been the following:
-
(1) Shot position and angle of the shot
Whilst all operators attempted to shoot frontally rather than a poll shot, donkeys tended to be shot caudally of the suggested position, with this deviation associated with the presence of normal-like EEG activity post-stun but not with behavioural signs of ineffective stunning. A caudal shot, combined with a shallow angle due to the fixed-angle of the pneumatic PCB stunner, particularly where donkeys moved or lowered their heads, is more likely to have missed the ascending reticular activating system (ARAS), which regulates cortical activity and conscious awareness. The ARAS includes the thalamus, midbrain and/or rostral pons, with these therefore the target structures for PCB stunning to achieve irrecoverable unconsciousness (Wedekind et al. Reference Wedekind, Hesselmann, Lippert-Grüner and Ebel2009; Terlouw Reference Terlouw2020; Edlow et al Reference Edlow, Claassen, Schiff and Greer2021; Fletcher et al. Reference Fletcher, Benedetti, Limon, Grist, Padalino, Hernández Gil and Gibson2025). A caudal shot could potentially cause merely superficial damage to the cerebellum, which controls posture and movement, resulting in the animal maintaining or recovering consciousness, but being unable to right itself (Grist et al. Reference Grist, Knowles and Wotton2019; Terlouw Reference Terlouw2020; Večerek et al. Reference Vecerek, Kamenik, Voslarova, Vecerkova, Machovcova, Volfova and Konvalinova2020; Gibson et al. Reference Gibson, Ridler, Limon, Lamb, Williams and Gregory2025). There is also a risk of the shot hitting the spinal cord, causing paralysis but not irrecoverable unconsciousness (Terlouw Reference Terlouw2020). However, this hypothesis could not be verified, as brain pathology was not performed on these donkeys.
-
(2) Restraint
In this study, the donkeys were restrained by a neck rope held tightly by personnel, enabling more precise application of the EEG electrodes and consequently a more precise shot to the suggested position on the head. However, head restraint is not typically used for equids being slaughtered by PCB (Fletcher et al. Reference Fletcher, Limon, Agongo, Akunzule, Essel, Padalino, Grist and Gibson2024) and was not usual practice for this abattoir, potentially impacting shot position and causing additional stress to the donkey prior to stunning. Restraint and placement of the PCB were complicated by the design of the stunning box, which was initially designed for cattle, and operators were required to lean down into the box to reach the donkeys, who were much smaller in stature than slaughter-weight cattle.
-
(3) Airline pressure and velocity
Stunning effectiveness could also have been influenced by airline pressure and velocity, although it was not possible to assess and control this. Abattoir operators should check the device regularly to ensure that it is working correctly and capable of reaching an adequate penetration depth to achieve irreversible unconsciousness (Gibson et al. Reference Gibson, Mason, Spence, Barker and Gregory2014; Kamenik et al. Reference Kamenik, Paral, Pyszko and Voslarova2019; Baier & Willson Reference Baier and Willson2020; Grist et al. Reference Grist, Bock, Knowles and Wotton2020).
-
(4) Level of operator experience and training
These could also have influenced stunning effectiveness however were unable to be verified due to a lack of information on training levels of expertise.
-
(5) Lack of specific guidance for stunning position for donkeys
Use of the same reference points suggested for horses is commonplace and was the case in this study. However, even if similar positioning could be hypothesised, this does not account for differences in skull morphology between these two species (Merkies et al. Reference Merkies, Paraschou and McGreevy2020). This could have partially influenced the number of ineffectively stunned animals. For this reason, empirical anatomical studies on appropriate shooting and stunning position for donkeys would be valuable further research.
Out of the six animals that showed periods of ‘normal like’ EEG, only one based on behavioural/brainstem signs, was classified as incompletely concussed, four showed behavioural signs and reflexes potentially indicating signs of a shallow depth of concussion, while the other two did not show any signs. Brain pathology could have confirmed whether these two animals which were not showing signs, had any marked trauma to key brain regions. This highlights how reliance on behavioural signs alone may not be reflective of the actual experience of the animal, potentially resulting in the underestimation of the animals that are still conscious after stunning. Whilst animals should always be routinely monitored between stunning and bleeding, an absence of behavioural signs does not absolutely ensure unconsciousness. This is particularly relevant where animals have incurred a shot into the spinal cord, absence of behavioural signs can mean paralysis rather than unconsciousness (Kumru et al. Reference Kumru, Vidal, Kofler, Portell and Valls-Solé2010; Terlouw et al. Reference Terlouw, Ducreux and Bourguet2021; Fletcher et al. Reference Fletcher, Benedetti, Limon, Grist, Padalino, Hernández Gil and Gibson2025). However, despite most shots being caudal of the suggested position, potentially into the occipital area, there was no attempt in the present study to shoot animals into the poll (behind the ears) rather than frontally (rostral of the ears), which would have increased the risk of such spinal cord paralysis.
Two animals displayed gasping, although for D10, this is likely to have been agonal in nature due to cerebral hypoxia, and the absence of EEG and behavioural signs of incomplete concussion. Gasping has been reported as a parameter of risk of consciousness or return to consciousness, however it can also be observed in unconscious states (Terlouw et al. Reference Terlouw, Ducreux and Bourguet2021). Gasping is considered the final respiratory effort to sustain life (Poets et al. Reference Poets, Meny, Chobanian and Bonofiglo1999). It is generated by an intrinsic medullary mechanism that is recruited when there is a dysfunction in the pons (St John Reference St John2009). Gasping corresponds to intermittent, inspiratory movements with an open mouth which can be induced by ischaemia or hypoxia (St John Reference St John2009). Gasping is very different from rhythmic breathing, and it is often accompanied by guttural sounds (Terlouw et al. Reference Terlouw, Bourguet and Deiss2016). Blackmore and Newhook (Reference Blackmore and Newhook1982) reported a PCB-shot calf displaying non-rhythmic gasping after having an isoelectric EEG for 60 s. Together, this suggests that although gasping can be a sign of the early stages of return of consciousness, it should be interpreted with caution and not in isolation from other indices (Terlouw et al. Reference Terlouw, Ducreux and Bourguet2021). It is therefore vital that multiple indices are used to determine shot effectiveness, rather than focusing on a single measure. Tools such as EEG are impractical for use in routine commercial slaughter situations, with abattoir operatives therefore needing to rely on behavioural and brainstem indices as important routine measures, despite their limitations, but still exercising caution even where no responses are observed (Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Terlouw Reference Terlouw2020; Fletcher et al. Reference Fletcher, Benedetti, Limon, Grist, Padalino, Hernández Gil and Gibson2025).
The single animal that appeared to show more overt rhythmic respiration, also showed spontaneous blinking and a period of normal-like EEG. If just rhythmic respiration had been seen in the absence of other indices, this may not necessarily indicate consciousness, but suggests that, particularly alongside the presence of eye reflexes, the animal might be more likely to recover consciousness through oxygenated blood continuing to reach the brain (Terlouw et al. Reference Terlouw, Bourguet and Deiss2016; Borzuta et al. Reference Borzuta, Lisiak, Janiszewski and Grześkowiak2019).
Study limitations
Although this study provided the first insights into stunning effectiveness in donkeys, the results must be interpreted with caution as there are certain limitations. Firstly, the sample size of 13 donkeys was small, but unavoidable due to the complexity of abattoir-based EEG research. Secondly, brain pathology of the slaughtered donkeys was not conducted, macro- or microscopically, to determine the precise location of the shot alongside the level and extent of brain trauma, with stunning position measured on skinned heads. This was unavoidable due to the field-based working conditions. Thirdly, animals were not monitored between hoisting and exsanguination, which was not possible due to the design of the abattoir and the risk it presented to researcher safety. The study was also observational in nature, with no interference by the research team on commercial decision making, including shot effectiveness and re-stunning.
In some studies, local anaesthesia is used to desensitise the skin prior to EEG electrode placement (Gibson et al. Reference Gibson, Rebelo, Gowers and Chancellor2018; Rucinque et al. Reference Rucinque, Velarde, Xercavins, Varvaró-Porter, Gibson, Michel and Contreras-Jodar2024). However, in this study it was not possible to use either topical ELMA cream or injected local anaesthetic due to the additional time and stress further handling would cause, and importantly because these were food-producing animals. EMLA cream is not licensed for food-producing animals, and if prescribed would have required withdrawal period (28 days in the UK).
Whilst EEG assessment post-stun briefly delayed exsanguination by a few seconds, abattoir personnel should be routinely monitoring animals from stun to stick, repeating the shot if there is any doubt of effectiveness, and generally minimising delays between these stages to ensure rapid exsanguination. This should then minimise the risk of animals recovering during this period. Finally, the presence of researchers could have influenced both operator and animal behaviour. However, the research team were cognisant to not interfere with the operation of the abattoir, and these risks tend to be unavoidable in commercial conditions.
Animal welfare implications and Conclusion
This study provides the first insight into electroencephalographic (EEG) responses of donkeys following penetrating captive-bolt (PCB) stunning. While PCB stunning generally induced insensibility, the presence of transient normal-like EEG patterns and the absence of isoelectric signals in some animals raise critical animal welfare concerns. These findings suggest a risk of delayed or incomplete loss of consciousness, or recovery of consciousness prior to exsanguination, particularly when shot placement deviates from the optimal trajectory or when stun-to-stick intervals are prolonged. The study emphasises the need for species-specific guidelines for donkeys, improvements in abattoir design, and stricter monitoring protocols to ensure effective stunning and minimise the risk of recovery, pain or distress during slaughter.
Acknowledgements
This study was funded by World Horse Welfare. The authors wish to thank all the staff at the abattoir, Dr Macario Sanne and Dr Sergio Main for their help and support.
Competing interests
None. The views presented in this paper are exclusively those of the authors and are not intended to represent the position of EFSA.






 Open access
Open access