Introduction
Synthesised pheromones, and related insect attractants that attract conspecifics over long distances, are used to monitor and manage many pest species (Witzgall et al. Reference Witzgall, Kirsh and Cork2010) and in programmes to conserve rare and endangered species (e.g., Larsson Reference Larsson2016). Because sampling multiple species simultaneously may be desirable, there is growing interest in developing multi-species lures that attract a number of species to a single trap. Such multi-species lures have been tested for scolytine bark beetles (Rassati et al. Reference Rassati, Toffolo, Roques, Battisti and Faccoli2014), lepidopterans (Brockerhoff et al. Reference Brockerhoff, Suckling, Roques, Jactel, Branco and Twidle2013), mirid bugs (Yasuda and Higuchi Reference Yasuda and Higuchi2012), and cerambycid beetles (Fan et al. Reference Fan, Denux, Courtin, Bernard, Javal and Millar2019), among others.
Multi-species lures can greatly simplify trapping programmes by reducing the number of traps needed and therefore the costs and effort required to deploy and service them (Brockerhoff et al. Reference Brockerhoff, Suckling, Roques, Jactel, Branco and Twidle2013). However, care must be taken in developing such blends because of the potential for antagonism among compounds in a blend that reduces attraction of target species to their pheromones. Antagonism is more likely when pheromones of closely related species are combined because of similarities in their pheromone chemistries (e.g., Brockerhoff et al. Reference Brockerhoff, Suckling, Roques, Jactel, Branco and Twidle2013; Byers et al. Reference Byers, Birgersson and Francke2013; Silva et al. Reference Silva, Zou, Bento, Hanks and Millar2017). Therefore, the most effective multi-species lures are likely to be composed of the pheromones of more distantly related taxa, such as blends of pheromones of beetles and hemipterans (Chase et al. Reference Chase, Stinger, Butler, Liebhold, Miller, Shearer and Brockerhoff2018).
Here, we explore potential interference among pheromones of beetles of two families, Cerambycidae and Elateridae (Coleoptera). A possible scenario for a combined cerambycid–elaterid lure would be in detecting incursions of invasive species from either family. For example, North America has been invaded by a number of economically important click beetles, particularly in the genus Agriotes (Elateridae) (Rashed and van Herk Reference Rashed and van Herk2024), and by several harmful species of the Cerambycidae, such as Asian longhorned borer and Tetropium spp. (Eyre and Haack Reference Eyre, Haack and Wang2017), with other species likely to be accidentally introduced, such as Aromia bungii (Faldermann) and Xylotrechus chinensis (Chevrolat) (both Cerambycidae), which are pests of tree fruits and vineyards, respectively (Sarto i Monteys et al. Reference Sarto i Monteys, Ribes and Savin2021; Urano et al. Reference Urano, Taki and Shoda-Kagaya2022).
Mate location in cerambycids in the largest subfamilies Cerambycinae and Lamiinae is mediated by aggregation–sex pheromones produced by males that attract both sexes (Millar and Hanks Reference Millar, Hanks and Wang2017). The larvae of most cerambycid species are endophytic, feeding within woody tissues of plants that range from living to dead and decomposing (Hanks Reference Hanks1999). In contrast, most elaterids use female-produced sex pheromones that attract only males (Tóth Reference Tóth2013; Rashed and van Herk Reference Rashed and van Herk2024). Larvae of most elaterid species are subterranean and can damage crop plants (Traugott et al. Reference Traugott, Benefer, Blackshaw, van Herk and Vernon2015; Rashed and van Herk Reference Rashed and van Herk2024). Larvae of forest-inhabiting elaterids often feed beneath the bark of trees, where they are predators of other subcortical insects, including cerambycids (e.g., Linsley Reference Linsley1959; Ware and Stephen Reference Ware and Stephen2006; Zhang et al. Reference Zhang, Yang, Huang and Fan2008).
Our experimental objective was to test a generic cerambycid pheromone blend, a generic elaterid pheromone blend, and a combination of the two to determine whether attraction of the target elaterid species was affected by the presence of cerambycid pheromones. Trap capture data were not analysed for cerambycid beetles because their numbers were insufficient for statistical analysis.
The blend of cerambycid aggregation–sex pheromones used in the present experiment was developed by Hanks et al. (Reference Hanks, Millar, Mongold-Diers, Wong, Meier and Reagel2012) and was composed of six pheromones shared by a number of species in the Cerambycinae (racemic 3-hydroxyhexan-2-one and 2-methylbutan-1-ol, syn-2,3-hexanediol) and Lamiinae ([E]-6,10-dimethyl-5,9-undecadien-2-ol, the corresponding acetate, and 2-[undecyloxy]ethanol). This blend, and similar combinations of cerambycid pheromones, have proven effective in attracting numerous cerambycid species in North America (e.g., Hanks and Millar Reference Hanks and Millar2013; Hanks et al. Reference Hanks, Mongold-Diers, Atkinson, Fierke, Ginzel and Graham2018) and elsewhere in the world (e.g., Roques et al. Reference Roques, Ren, Rassati, Shi, Akulov and Audsley2023).
The blend of elaterid attractants was developed by Rusen et al. (Reference Rusen, Millar, McElrath and Hanks2026) and included synthesised sex pheromones or sex attractants of nine species of elaterids (Coleoptera: Elateridae) native to the midwestern United States of America. Of eight species captured in numbers sufficient for statistical analysis, half were as strongly attracted to the blend as they were to their synthetic attractant; two of the remaining species showed lesser but still significant attraction to the blend, whereas the blend was not attractive to two species, suggesting antagonism among the components. Seven of the attracted species of the Elateridae were targeted with the same blend in the present study, including Elater abruptus Say, Melanotus ignobilis Melsheimer, Melanotus sagittarius (LeConte), Melanotus similis (Kirby), and Parallelostethus attenuatus (Say) of the subfamily Elaterinae, and the congeners Gambrinus griseus (Palisot de Beauvois) and G. rudis Brown of the Dendrometrinae.
syn- and anti-2,3-Hexanediols were synthesised as described in Lacey et al. (Reference Lacey, Ginzel, Millar and Hanks2004, Reference Lacey, Moreira, Millar, Ray and Hanks2007). Other cerambycid compounds were purchased from commercial sources, including racemic 3-hydroxyhexan-2-one, racemic (E)-6,10-dimethyl-5,9-undecadien-2-ol (known as fuscumol), the corresponding (E)-6,10-dimethyl-5,9-undecadien-2-yl acetate (fuscumol acetate), and 2-(undecyloxy)ethanol (monochamol; from Bedoukian Research, Inc., Danbury, Connecticut, United States of America), and racemic 2-methylbutan-1-ol (Aldrich Chemical, Milwaukee, Wisconsin, United States of America).
Attractants in the elaterid blend were synthesised as follows: tetradecyl acetate (as in Williams et al. Reference Williams, Serrano, Johnson and Millar2019), 1,8-octanediol dihexanoate (Millar et al. Reference Millar, Williams, Halloran, Serrano, Grommes and Huseth2022), geranyl butyrate (Serrano et al. Reference Serrano, Landolt, Reyes Corral and Millar2022), 5-methylhexyl (Z)-4-decenoate, 11-dodecenyl butyrate, and limoniic acid (Millar et al. Reference Millar, Williams, Serrano, McElrath, Zou and Grommes-Yeager2024), and butyl (Z)-5-dodecenoate and (E)-13-hexadecenyl acetate (Hanks et al. Reference Hanks, Davis, McElrath, Zou and Millar2025).
The two study sites for the field experiment in east–central Illinois, United States of America, are approximately 90 km apart: Forest Glen Preserve (Vermilion County; 40.0152° N, −87.5677° W; property of the Vermilion County Conservation District; http://www.vccd.org/) and Robert Allerton Park (Piatt County; 39.996° N, –88.651° W; property of the University of Illinois). Both sites are wooded with second-growth and old-growth hardwood forests dominated by oaks, Quercus spp. (Fagaceae), hickories, Carya spp. (Juglandaceae), and maples, Acer spp. (Sapindaceae).
Attraction of elaterids to experimental treatments was assessed with black cross-vane panel traps (AlphaScents, Portland, Oregon, United States of America) coated with the fluoropolymer dispersion Fluon® PTFE (AGC Chemicals Americas, Inc., Exton, Pennsylvania, United States of America). Traps were hung from inverted L-shaped frames constructed from polyvinyl chloride irrigation pipe at a height of approximately 0.5 m. Captured beetles were preserved by partially filling trap basins with saturated aqueous NaCl solution. Pheromone lures were polyethylene press-seal baggies (Bagette model 14770, 5.1 × 7.6 cm, 0.05 mm thick, Cousin Corp., Largo, Florida, United States of America) that contained a cotton dental roll (1 × 4 cm, Patterson Dental Supply, Mendota Heights, Minnesota, United States of America). Lures containing attractants of cerambycids were loaded with 25 mg of achiral monochamol and 50 mg of the remaining racemic compounds (i.e., 25 mg of each enantiomer) in 1 mL of hexane. Lures for elaterids contained 2 μL of each of the eight attractant compounds in 100 μL of hexane. In the combined cerambycid-plus-elaterid blend treatments, traps were baited with one lure of each blend. Control traps were baited with sachets loaded with 100 μL of hexane.
One transect of traps, with one trap per treatment (traps located ∼10 m apart), was set up at each study site on 25 May 2025 at the Allerton site and on 29 May 2025 at the Forest Glen site, and the experiment was ended on 1 and 4 August 2025, respectively, encompassing the seasonal activity periods of the targeted elaterid species (see Hanks et al. Reference Hanks, Davis, McElrath, Zou and Millar2025). Traps were serviced every 2–3 days, at which time, treatments were rotated down transects to allow for location effects. Elaterids were identified with reference to voucher specimens verified by a taxonomist (see Rusen et al. Reference Rusen, Millar, McElrath and Hanks2026). Representative voucher specimens were deposited with the collection of the Illinois Natural History Survey, Champaign, Illinois, United States of America.
Trap capture data were analysed separately for the seven elaterid species. Captured beetles were assumed to be males, based on our earlier experiments with the same species (e.g., Millar et al. Reference Millar, Williams, Halloran, Serrano, Grommes and Huseth2022; Millar et al. Reference Millar, Williams, Serrano, McElrath, Zou and Grommes-Yeager2024; Hanks et al. Reference Hanks, Davis, McElrath, Zou and Millar2025). Replicates were defined by the number of transects and collection date (i.e., replication over space and time). Differences between treatment means were tested with the nonparametric Friedman’s test (PROC FREQ, option CMH; SAS Institute 2011). Replicates with no elaterid beetles in any trap within a given transect of traps were dropped from analyses to allow for a lack of activity. Pairs of treatment means were compared by converting the raw data to ranks (Zar Reference Zar1999) and by using the nonparametric Ryan–Einot–Gabriel–Welsch Q (REGWQ) multiple comparison test (SAS Institute 2011).
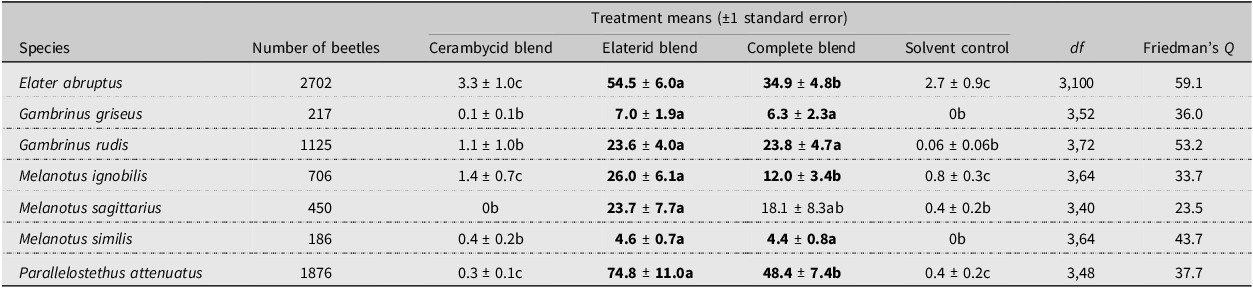
As expected, none of the seven elaterid species was significantly attracted to traps baited only with the cerambycid pheromone blend (Table 1). There were no significant differences in attraction of three of the species (G. griseus, G. rudis, and M. similis) to the elaterid blend versus the combined blend of cerambycid-plus-elaterid attractants (Table 1). Of the remaining species, the combined blend attracted fewer males of E. abruptus, M. ignobilis, and P. attenuatus than did the elaterid blend alone, suggesting antagonism by components of the cerambycid blend. Nevertheless, the combined blend attracted significant numbers of those species relative to the controls. For the seventh species, M. sagittarius, males were significantly attracted by the elaterid blend compared to the control, but the mean for the combined blend was intermediate.
Results of field experiment targeting seven species of elaterid beetles. Means within species with different letters are significantly different (REGWQ test, P < 0.05), and treatment means in bold are significantly different from that of the control. P-values for all analyses were less than 0.0001
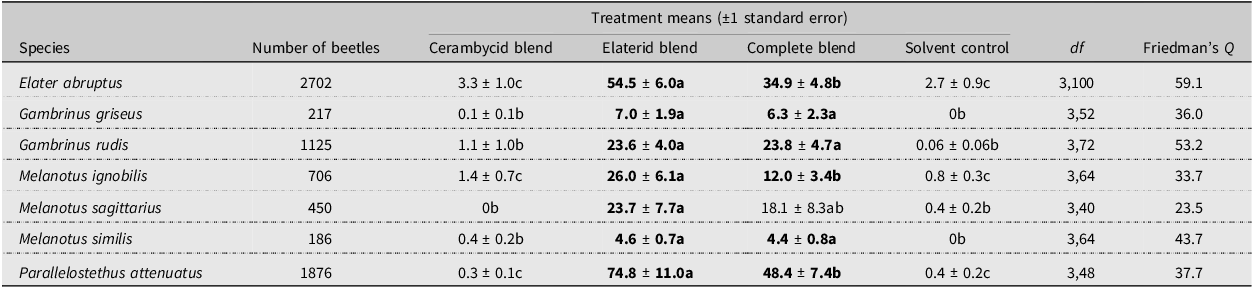
The variable influence of the blend of cerambycid pheromones on attraction of elaterids to the elaterid blend may be related to the nature of their larval hosts. That is, the larvae of the elaterid species that were antagonised by the cerambycid pheromone blend develop in woody tissues of plants, as do larvae of most cerambycid species (Švácha and Lawrence Reference Švácha, Lawrence, Leschen and Beutel2014). Specifically, larvae of E. abruptus, M. ignobilis, and P. attenuatus feed within dead woody plants (Jewitt Reference Jewitt1946; Kirk Reference Kirk1922; Mathison Reference Mathison2021), and fewer adults of these species were attracted to the elaterid blend when it was combined with the cerambycid blend. Adults of M. sagittarius also may have been inhibited by the complete blend, and larvae of this species are also likely to feed subcortically (Kirk Reference Kirk1922).
The antagonistic effect of the cerambycid pheromones on attraction of these species to their pheromones suggests that there is some selective advantage in detecting and avoiding cerambycids. Such could be the case if cerambycid larvae dominate in aggressive interactions with subcortical elaterids. Larvae of elaterids in that niche may be predaceous, and in fact, one of our study species, E. abruptus, may prey upon saproxylic larvae of a scarab species (Steffek et al. Reference Steffek, Grommes, Hanks and Mitchell2014). However, cerambycid larvae also can be facultative predators, just as they consume conspecifics while burrowing through wood (e.g., Togashi Reference Togashi1986; Victorsson and Wikars Reference Victorsson and Wikars1996; Hanks et al. Reference Hanks, Paine and Millar2005). Thus, the ability of male elaterids to detect aggregation–sex pheromones of cerambycids may be beneficial if it reduces the chances they would be attracted by conspecific females to hosts already occupied by ovipositing cerambycid females, preventing their descendants from being eaten by cerambycid larvae. In contrast, larvae of G. griseus, G. rudis, and M. similis, whose adults were not influenced by cerambycid pheromones, are believed to feed on roots of deciduous agricultural plants (Fattig Reference Fattig1951; Riley and Keaster Reference Riley and Keaster1979; Mathison Reference Mathison2021; Smith et al. Reference Smith, van Herk, Schwinghamer, Baute, Limay-Ross and Frewin2024), where there is no potential for predation by cerambycid larvae.
Although our study was designed to also test the effects of elaterid pheromones on attraction of cerambycids to their pheromones, those plans were stymied by the negligible catches of cerambycids. The cause of these low numbers of cerambycids is unknown, but the low numbers follow a decline in trap captures in the area of our studies in recent years (unpublished data). Professor Marlin Rice (emeritus, Iowa State University, Ames, Iowa, United States of America) has confirmed similar declining numbers of cerambycids during pheromone and light trapping in the American Midwest (Marlin Rice, personal communication).
In summary, our data indicate some degree of disruption of elaterid semiochemical communication by cerambycid pheromones, but not to the extent that combined lures could not be used to monitor the elaterid species described here.
Acknowledgements
The authors thank Rachel Rusen, Elijah Davis, and Coen Carroll for technical support and the University of Illinois Division of Parks and Natural Spaces and Vermilion County Conservation District for access to field sites.
Funding statement
The authors appreciate funding support from the United States Department of Agriculture, Agricultural Research Service, and State Potato Partnership Funds, projects 2092-22000-022-00D and 6080-22000-030-00D.
Competing interests
The authors declare that they have no competing interests.