


Introduction
Agroecological farming systems rely on ecological processes and biodiversity to transition towards sustainable agricultural practices (Altieri, Reference Altieri1999; Foley et al., Reference Foley, DeFries, Asner, Barford, Bonan, Carpenter, Chapin, Coe, Daily, Gibbs, Helkowski, Holloway, Howard, Kucharik, Monfreda, Patz, Prentice, Ramankutty and Snyder2005; Gleismann, Reference Gliessman2016; Isaac et al., Reference Isaac, Sinclair, Laroche, Olivier and Thapa2024). Among the various forms of agroecological practices, natural farming in India is a grassroot farmer-led movement, which aims to meet the dual goals of addressing food security and mitigating environmental challenges (Duddigan et al., Reference Duddigan, Shaw, Sizmur, Gogu, Hussain, Jirra, Kaliki, Sanka, Sohail, Soma, Thallam, Vattikuti and Collins2023; Ravisankar et al., Reference Ravisankar, Praveen Rao, Thakur, Krishnan, Rani, Joshi, Prusty, Shamim, Ansari, Singh, Raghavendra, Swarnam, Panwar, Bhaskar, Kumar and Singh2024). Natural farming involves using on-farm resources to minimize the usage of external chemical inputs. Additionally, natural farming emphasizes biological and ecological functions within agroecosystems using practices such as no-till, mulching, and intercropping, and unique homemade bio-inputs made from on-farm bioresources (Darjee et al., Reference Darjee, Singh, Dhar, Pandey, Dwivedi, Sahu, Rai, Alekhya, Padhan, Ramalingappa and Shrivastava2024; Duddigan et al., Reference Duddigan, Collins, Hussain, Osbahr, Shaw, Sinclair, Sizmur, Thallam and Winowiecki2022).
Agroecological transitions can be defined as a shift towards more biodiverse-mediated models of agricultural systems (Tittonell, Reference Tittonell2020; Wezel et al., Reference Wezel, Gemmill-Herren, Bezner Kerr, Barrios, Rodrigues Gonçalves and Sinclair2020). According to Gliessman (Reference Gliessman2016), this transition can be understood through five progressive levels, which range from improving efficiency of conventional inputs to input substitution using ecological processes, to full redesign of agroecosystems, and ultimately to broader transformations of food systems and governance. Several studies on agroecological transitions have focussed on understanding how soil health (Thapa et al., Reference Thapa, Muthuprakash, Damani, Bell and Isaac2025) and biodiversity (Blaix et al., Reference Blaix, Dumont, Bloor, Zagaria, Fleurance, Joly and Huguenin-Elie2025) outcomes change in response to management (Teixeira et al., Reference Teixeira, Bianchi, Cardoso, Tittonell and Peña-Claros2021). These studies tend to show that soil health and biodiversity increase along an agroecosystem transition.
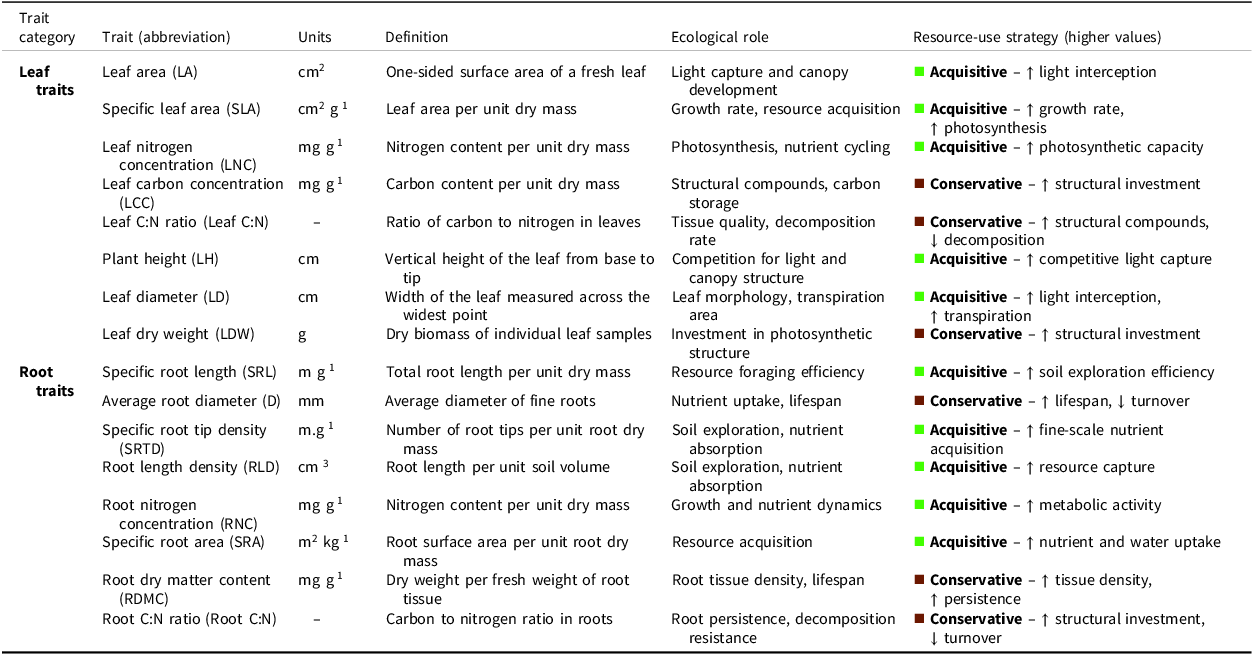
Yet, few studies examine how such a transition impacts crop physiology, and in particular, crop trait expression. Changes in crop trait expression may have consequences for agroecosystem functions. Specifically, crop functional traits, defined as morphological, physiological, and phenological characteristics of plants that influence their fitness, resource-use efficiency, and ecosystems interactions (Díaz et al., Reference Díaz, Kattge, Cornelissen, Wright, Lavorel, Dray, Reu, Kleyer, Wirth, Colin Prentice, Garnier, Bönisch, Westoby, Poorter, Reich, Moles, Dickie, Gillison, Zanne, Chave, Joseph Wright, Sheremet’ev, Jactel, Baraloto, Cerabolini, Pierce, Shipley, Kirkup, Casanoves, Joswig, Günther, Falczuk, Rüger, Mahecha and Gorné2016; Garnier et al., Reference Garnier, Navas and Grigulis2015), will respond to management practices and contribute to key agroecosystem services, including nutrient cycling, abiotic and biotic stress resilience, and crop productivity (Martin and Isaac, Reference Martin and Isaac2015; Pacini et al., Reference Pacini, Bruschi, Ferretti, Santoni, Serafini and Gaifami2023). Studies have shown that trait variation, both within and among crops, can significantly influence agroecosystem functioning (Buchanan et al., Reference Buchanan, Baskerville, Oelbermann, Gordon, Thevathasan and Isaac2020; Gagliardi et al., Reference Gagliardi, Avelino, Beilhe and Isaac2020; Isaac et al., Reference Isaac, Cerda, Rapidel, Martin, Dickinson and Sibelet2018; Sauvadet et al., Reference Sauvadet, Trap, Damour, Plassard, Van den Meersche, Achard, Allinne, Autfray, Bertrand, Blanchart, Deberdt, Enock, Essobo, Freschet, Hedde, de Melo Virginio Filho, Rabary, Rakotoarivelo, Randriamanantsoa, Rhino, Ripoche, Rosalie, Saj, Becquer, Tixier and Harmand2021), while other studies have applied a functional trait framework to better understand the effects of domestication on crops (Nimmo et al., Reference Nimmo, Brar, Martin and Isaac2025). Understanding intraspecific trait variation, and trait relationships, helps shape our capacity to make accurate predictions about the performance and fate of crops to optimize crop attributes in agroecological or natural farming systems (see Table 1).
Leaf and root traits measured across agroecological transitions drawn from established trait-based ecological literature (Wright et al. (Reference Wright, Reich, Westoby, Ackerly, Baruch, Bongers, Cavender-Bares, Chapin, Cornelissen, Diemer, Flexas, Garnier, Groom, Gulias, Hikosaka, Lamont, Lee, Lee, Lusk, Midgley, Navas, Niinemets, Oleksyn, Osada, Poorter, Poot, Prior, Pyankov, Roumet, Thomas, Tjoelker, Veneklaas and Villar2004), Bardgett et al. (Reference Bardgett, Mommer and De Vries2014), Guerrero-Ramírez et al. (Reference Guerrero-Ramírez, Mommer, Freschet, Iversen, McCormack, Kattge, Poorter, van der Plas, Bergmann, Kuyper, York, Bruelheide, Laughlin, Meier, Roumet, Semchenko, Sweeney, van Ruijven, Valverde-Barrantes, Aubin, Catford, Manning, Martin, Milla, Minden, Pausas, Smith, Soudzilovskaia, Ammer, Butterfield, Craine, Cornelissen, de Vries, Isaac, Kramer, König, Lamb, Onipchenko, Peñuelas, Reich, Rillig, Sack, Shipley, Tedersoo, Valladares, van Bodegom, Weigelt, Wright and Weigelt2021), Buchanan (Reference Buchanan2022))

Acquisitive traits refer to functional characteristics associated with rapid resource uptake and use, high metabolic activity, and fast tissue turnover, typically favoured under resource-rich or dynamic environments.
Conservative traits refer to characteristics associated with greater structural investment, slower resource use, longer tissue lifespan, and increased persistence, typically favoured under resource-limited or stressful environments.
Maize (Zea mays L.) is one of the most widely cultivated crops globally (Kiboi et al., Reference Kiboi, Bautze, Matheri, Riar and Fliessbach2025), especially in the context of India and natural farming (Mallareddy et al., Reference Mallareddy, Vinay, Padmaja, Jayasree, Padmaja and Vijayakumar2023). Maize has undergone dramatic changes in shoot and root development, physiology, and trait expression, through selection in increasingly managed agroecosystems (Schmidt et al., Reference Schmidt, Bowles and Gaudin2016). Early landraces were cultivated in diverse, low-input systems yet subsequent shifts towards mechanized tillage, synthetic fertilizer use, and monocultures intensified selection for aboveground traits that maximize yield under high-input environments (Gaudin et al., Reference Gaudin, McClymont and Raizada2011). As a result, these changes may have potentially resulted in unintended changes in root system development, rhizosphere functioning, and plant–microbe interactions (Isaac et al., Reference Isaac, Nimmo, Gaudin, Leptin, Schmidt, Kallenbach, Martin, Entz, Carkner, Rajcan, Boyle and Lu2021).
Maize trait expression is strongly shaped by management context, particularly in relation to nutrient availability and soil structure (Jiang and Whalen, Reference Jiang and Whalen2025). Similar patterns have been documented globally, where maize functional traits respond to soil fertility, resource availability, and environmental gradients across diverse agroecosystems (Li et al., Reference Li, Zeng, Xu, Wang, Wu, Cao, Lan, Liu, Lu and Zhang2015; Lynch, Reference Lynch2013; Vetterlein et al., Reference Vetterlein, Phalempin, Lippold, Schlüter, Schreiter, Ahmed, Carminati, Duddek, Jorda, Bienert, Bienert, Tarkka, Ganther, Oburger, Santangeli, Javaux and Vanderborght2022). Maize grown in conventional agriculture typically exhibits traits such as high specific leaf area (SLA), rapid aboveground biomass accumulation, shallow, dense root systems, and high nutrient uptake efficiency optimized for immediate nutrient and water availability (Lynch, Reference Lynch2013; Jiang and Whalen, Reference Jiang and Whalen2025). These conditions, while maximizing short-term productivity, often result in lower nutrient use efficiency and reduced root plasticity, making maize systems especially vulnerable to nutrient losses and environmental stresses (Robertson and Vitousek, Reference Robertson and Vitousek2009). In contrast, maize grown under organic systems frequently demonstrates higher root: shoot ratios, deeper rooting, increased reliance on mycorrhizal associations, greater root plasticity, and finer root structures, all of which individually and cumulatively reflect selection for enhanced nutrient foraging in nutrient-limited environments (Gaudin et al., Reference Gaudin, Tolhurst, Ker, Janovicek, Tortora, Martin and Deen2015). Overall, one might hypothesize that shifts from synthetic to organic and diversified cropping systems lead to changes in maize trait strategies, favouring plastic, responsive, and synergistic maize phenotypes that are better adapted to complex agroecosystems. However, little is known on trait expression changes over the entire time course of agroecological transitions.
In this study, our main objective is to understand how maize functional trait expression and resource acquisition strategies differ along agroecological transitions. We ask 1) do root and leaf traits change along an agroecological transitions? If so: 2) do root and leaf traits become more acquisitive or conservative as transitions proceed? And 3) do root or leaf traits show increasing variability along agroecological transitions? We hypothesize that crops shift their resource strategies to acquisitive trait expression, as crops encounter organic amendments and higher diversity, and that root traits show high variability along an agroecological transition. By linking trait expression with transition phases, we aim to identify the most responsive functional traits to understand crop strategy sensitivity along agroecological transitions, contributing a novel understanding of crop adaptation and agroecosystem functioning.
Methods
Study sites and characteristics
We conducted our study on 44 farms in September to October 2022 located in Dewas region, western Madhya Pradesh, India. These sites represented maize-growing farms in three sampling clusters/villages. The sites fall within 20°17′ and 23°20′ N latitude and 75°54′ and 77°08′ E longitude that span one agroclimatic zone. The area experiences a subtropical climate, with an average annual rainfall of 1067 mm (Rizwan et al., Reference Rizwan, Kulshreshtha and Dev2016).
The participation of farmers and crop selection were facilitated through collaboration with local NGO partners, Samaj Pragati Sahayog. This organization supports farmers through training, and also and provision of seeds, including maize of the same seed source, which was our selected test crop. Farms were categorized into five distinct transitional phases based on their management practices, history of chemical inputs use and application of bio-inputs (Figure 1). These five phases form a chronosequence of various stages of transition. Phase 1 (n = 9) farms include conventional farms applying pesticides, fertilizers, and other chemical inputs. Phase 2 (n = 9 farms) includes farms, which recently (within the past two years) stopped using chemical pesticides but continue to use other chemical inputs such as fertilizers. Phase 3 (n = 10 farms) includes farms which stopped applying pesticides over three years ago but still use fertilizers. Phase 4 (n = 8 farms) includes farms who recently (within the past four years) stopped using all chemicals, including fertilizers and pesticides, and have started using bio-inputs. Phase 5 (n = 8) includes farms who have not used chemicals and pesticides for over five years but have been consistently adding bio-inputs.
Conceptual representation of agroecological transitions over time and across phases used for analysis. Phase 1 farms depend fully on chemical inputs and are characterized by conservative root traits such as high root dry matter content (RDMC) and root carbon: nitrogen ratios (C:N). Transitional farms (Phases 2–3) show mixed strategies, including increasing specific root length (SRL) and specific root area (SRA). In agroecological farms (Phases 4–5), which do not apply any synthetic inputs, acquisitive and resilient traits, including high SRL and SRA, lower RDMC, and stable SLA. The y-axis denotes an increase in root trait plasticity alongside a reduction in external chemical inputs and an increase in agroecological management inputs over time.

Leaf and root trait sampling
In each of the 44 farms, we selected three representative plots (1 × 1 m in size) for plant and root trait measurements (see Table 1 for traits, units, definition, ecological role, and resource use strategy). Specifically, from each sampling plot, individual plants were selected, and leaf height (LH) and leaf diameter (LD) were measured. Then, the third leaf from top of the plant was collected and a photograph was taken for analysing the LA analysis. We selected the third fully expanded upper leaf to minimize intraplant variation due to leaf age and ontogeny, and sampled across three 1 × 1 m quadrats per farm to capture within-farm variability. While this design does not encompass all possible intraplant and intraplot heterogeneity, it provides a practical and standardized sampling scale for robust comparison across farms under field conditions (Cornelissen et al., Reference Cornelissen, Lavorel, Garnier, Díaz, Buchmann, Gurvich, Reich, ter Steege, Morgan, van der Heijden, Pausas and Poorter2003; Pérez-Harguindeguy et al., Reference Pérez-Harguindeguy, Díaz, Garnier, Lavorel, Poorter, Jaureguiberry, Bret-Harte, Cornwell, Craine, Gurvich, Urcelay, Veneklaas, Reich, Poorter, Wright, Ray, Enrico, Pausas, de Vos, Buchmann, Funes, Quétier, Hodgson, Thompson, Morgan, ter Steege, van der Heijden, Sack, Blonder, Poschlod, Vaieretti, Conti, Staver, Aquino and Cornelissen2013). Leaf area (LA, cm2) was determined using ImageJ V1.53a (Wayne Rasband, National Institute of Health, USA) software ImageJ based on pixel counting.
Fresh leaves were weighed fresh then dried to a constant mass at 65°C for 48 hours then weighed again to obtain leaf dry weight (LDW, g). SLA (cm2 g−1) was calculated as the ratio of LA to dry mass. Dried leaves were then homogenized in a Retsch ball mill (45 s; Retsch, Düsseldorf, Germany), and ∼ 0.1 g of ground material was sealed in foil capsules for leaf LNC and LCC concentration determination (% mass) analysed using a LECO CN628 (LECO Corporation, Minnesota, USA). Leaf C and N data were in turn used to derive leaf C:N ratios.
Following leaf trait collection, volumetric root traits were measured using a soil core. A section of soil was exposed to a depth of 10 cm, approximately 5 cm away from the base of the plant, and a 100 cm3 soil core was hammered horizontally towards the base of the plant. The soil core was filled with soil, plugged, and then emptied into a plastic bag. The roots were separated from the soil by sieving the sample through a 2 mm sieve. The roots were then scanned using a flatbed scanner to analyse root morphological traits such as root length (RL, cm), root area (RA, cm3) and average root diameter (D, cm) all using WinRhizo software (Regents Instruments, Montreal, Canada). Roots fresh weight was taken, then roots were dried in an oven at 65°C for 24 hours and then weighed to obtain dry mass and root dry matter content (RDMC). Root dry mass was used to calculate the specific root length (SRL, cm g−1) and specific root area (SRA, cm2 g−1). The specific root length (SRL, m g−1) was calculated as SRL divided by dried weight (g). Dried roots were then ground with a Retsch ball mill (Retsch, Düsseldorf, Germany) and analysed for total root carbon and total root nitrogen (RNC mg g−1) concentration using approximately 0.1 g on a LECO CN628 (LECO Corporation, Minnesota, USA) used to determine root C:N ratio (Root C:N).
Statistical analysis
All statistical analyses were performed using R Studio v. 4.5.2 (R Foundation for Statistical Computing, Vienna, Austria). Normality of maize leaf and root traits was assessed using Shapiro-Wilk tests, and traits were log-transformed where necessary. Descriptive statistics on the aboveground and belowground functional trait dataset were calculated for each trait at each different transition phase. The dataset included n = 132 observations per root and leaf traits. To evaluate the effects of agroecological transition phases on trait expression, a two-way ANOVA was conducted.
Intraspecific variation in root and leaf traits was quantified using the coefficient of variation (CV) across the full dataset. To assess distributional characteristics, both normal and log-normal models were fitted using maximum likelihood estimation. The best-fitting model was selected based on log-likelihood ratios, and traits best described by log-normal distributions were log-transformed for further analysis.
To explore multivariate trait relationships, we performed a principal component analysis (PCA) using the ‘vegan’ R package (Oksanen et al., Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, Mcglinn, Minchin, Hara, Simpson, Solymos, Stevens and Szoecs2019). To test for differences in multivariate trait composition across agroecological phases, we conducted a permutational multivariate analysis of variance (PerMANOVA) using the ‘adonis2’ function in the ‘vegan’ package, based on Euclidean distance matrices and n = 999 permutations. Group-specific 95% confidence ellipses were overlaid on the PCA biplot to visualize trait dispersion within each phase.
To quantify the functional trait space occupied by maize under different agroecological phases, we also constructed multidimensional hypervolumes using the ‘hypervolume’ R package (Blonder et al., Reference Blonder2018). Here, a functional trait hypervolume is defined as the multidimensional space occupied by a set of functional traits, representing the range and distribution of trait combinations expressed by crops under a given management regime (sensu Blonder et al., Reference Blonder, Lamanna, Violle and Enquist2014; Hutchinson, Reference Hutchinson1957). Hypervolume size reflects trait diversity and functional breadth, overlap reflects shared trait strategies, and centroid distance reflects shifts in dominant trait syndromes. Hypervolumes were generated separately for root and leaf traits using Gaussian kernel density estimation. Prior to analysis, all trait data were z-transformed (mean = 0, standard deviation = 1) to standardize trait scales. Hypervolume metrics, including total volume, Jaccard similarity, centroid distance, and unique volume percentages, were calculated to compare trait space overlap and divergence across transition phases. Simulated trait distributions were generated using the ‘replicate’ function based on trait means and standard deviations to ensure robust estimation of hypervolume metrics. To assess functional divergence across the agroecological transition, we calculated the centroid distance between trait hypervolumes of phase 1 and phase 5. To explore the trajectory of change, we also analysed pairwise distances among all phases.
Results
Intraspecific variation in leaf and root traits across agroecological phases
Variability in leaf traits was generally lower than in root traits (Tables 2 and 3). LH showed the lowest coefficient of variation (CV = 9.46%). In contrast, SLA (CV = 50.4%), LDW (CV = 31.2%), and LD (CV = 30.6%) displayed greater dispersion. Notably, leaf C:N ratio (CV = 27.2%) and LNC (CV = 25.3%) showed moderate variability. Root traits displayed higher intraspecific variability, with the highest CV observed for RDMC (CV = 288%), followed by SRA (CV = 92%) and SRL (CV = 153%). Other root traits, including SRTD (CV = 142%) and root C:N ratios (CV = 75.6%), also showed notable variation, indicating the presence of both acquisitive and conservative root strategies across phases. In contrast, D and RNC exhibited lower variability (CV = 31.2% and 39.4%, respectively), suggesting more conservative expression of these traits.
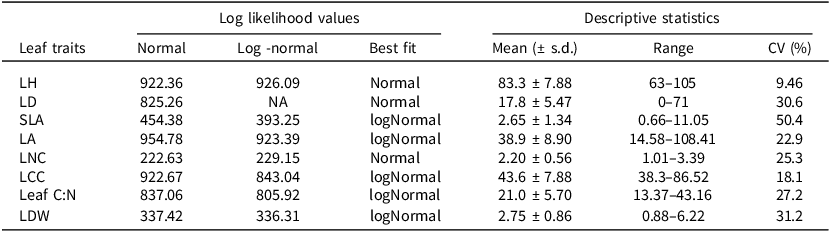
Model fit statistics for normal vs. log-normal distribution comparisons and descriptive statistics for leaf traits. Descriptive statistics are calculated from the original trait data (units as defined in Table 1 above)
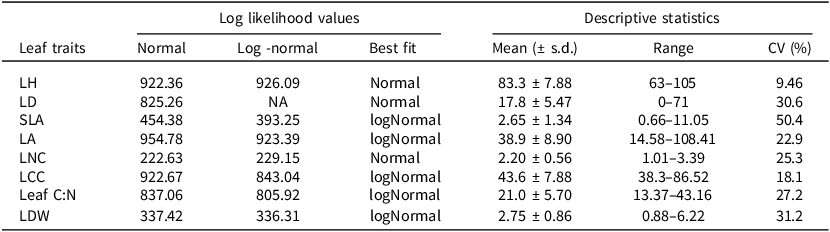
Log-likelihood values were used to compare model fits between normal and log-normal distributions for each leaf trait, with the distribution showing the higher log-likelihood selected as the best fit. Descriptive statistics (mean, range, and coefficient of variation, CV) are reported from the original, untransformed data. Traits identified as log-normally distributed were log-transformed prior to analyses.
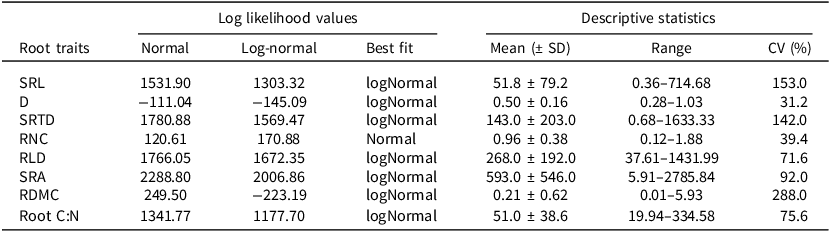
Model fit statistics for normal vs. log-normal distributions and descriptive statistics for root traits. Descriptive statistics are calculated from the original trait data (units as defined in Table 1 above)

Log-likelihood values were used to compare model fits between normal and log-normal distributions for each root trait, with the distribution yielding the higher log-likelihood selected as the best fit. Descriptive statistics (mean, range, and coefficient of variation, CV) are reported based on the original, untransformed trait values. Traits identified as log-normally distributed were log-transformed prior to subsequent analyses.
Root and leaf trait variation under different phases
Several root traits showed significant responses to agroecological transition phase (Table 4), for root D; (F = 5.93, p < 0.001), RNC; (F = 3.64, p = 0.008), RLD; (F = 3.59, p = 0.008), and root C:N ratio (F = 4.22, p = 0.003). In contrast, SRL, SRTD, SRA, and RDMC did not vary significantly across transition phases (p > 0.05). The regions in which sampling was conducted also influenced root trait expression, including for SRL (F = 7.27, p = 0.001), D (F = 20.31, p < 0.001), SRTD (F = 8.54, p < 0.001), RLD (F = 21.58, p < 0.001), SRA (F = 4.63, p < 0.012), and root C:N (F = 5.48, p < 0.01). Notably, significant phases-by-region interactions were found for SRTD (F = 8.54, p < 0.001).
ANOVA results for leaf and root traits across different treatment types and regions, and their interaction effects

Table 4. Long description
The table presents ANOVA results for leaf and root traits across different treatment types and regions and their interaction effects. It includes columns for Trait, Source, Df, Sum Sq, F (p-Value), and Significance. The table is divided into leaf traits and root traits. Each row lists specific traits and their corresponding values for different sources such as Phases, Region, and Phases x Region. Notable trends include significant responses in root traits like root D, RNC, RLD, and root C:N ratio across transition phases, while traits like SRL, SRTD, SRA, and RDMC did not vary significantly. Regional influences are also noted for several traits. Significant phase-by-region interactions are observed for SRTD.
Results of two-way ANOVA testing the effects of agroecological transition phase (Phases), region (District), and their interaction (Phases × Region) on leaf and root functional traits. Traits marked with “Log” were log-transformed prior to analysis to meet assumptions of normality. F-values are reported with associated p-values in parentheses. Significance levels are indicated as: * p < 0.05, ** p < 0.01, *** p < 0.001. Non-significant effects (p ≥ 0.05) are shown without symbols.
For leaf traits, significant transition phase effects were observed for LH (F = 3.76, p < 0.01) and leaf C:N (F = 3.72, p < 0.01). Region had a strong effect on several leaf traits, including LH, SLA, LNC, and leaf C:N (p < 0.001 in all cases). Again here, we detected significant phase-by-region interaction effects for LH, SLA, LNC, LCC, leaf C:N ratios and LDW.
Multivariate trait patterns across various phases
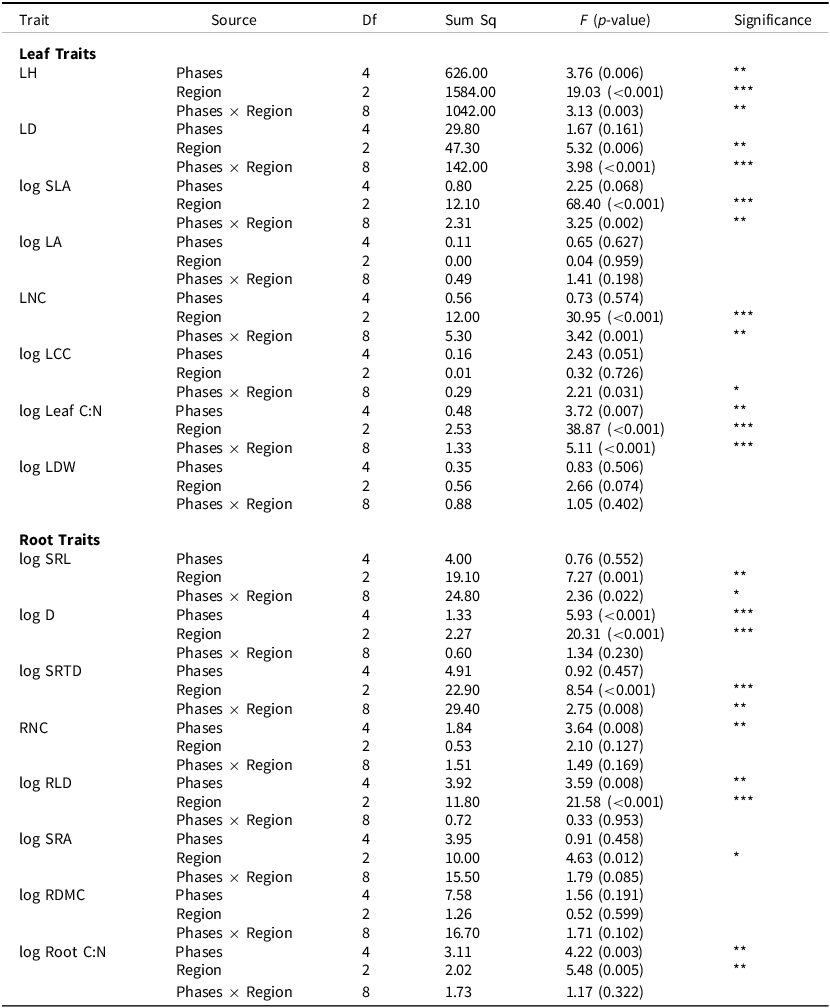
For root traits, the first two dimensions of our PCA explained 31.3% and 22.3% of the variation (Figure 2B), respectively. Traits such as root D, RDMC, and root C:N loaded positively along Dim1, while SRL, SRA, and SRTD loaded negatively, indicating a trait gradient from conservative to acquisitive root strategies. Our PerMANOVA showed a significant effect of transition phase on multivariate trait expression (F = 2.58, p = 0.001), indicating that root functional traits varied along the agroecological gradient (Figure 2B).
Principal component analysis (PCA) biplots of (A) leaf and (B) root functional traits of maize across five agroecological transition phases under natural farming in central India. Vectors represent trait loadings, and ellipses denote 95% confidence intervals for each phase. PERMANOVA indicates significant phase-level differences in root traits.

Figure 2. Long description
Panel A: A principal component analysis biplot displays leaf functional traits of maize across five agroecological transition phases. The x-axis is labeled Dim1 (29.9 percent) and the y-axis is labeled Dim2 (19.3 percent). Vectors represent trait loadings, and ellipses denote 95 percent confidence intervals for each phase. The traits include LNC, LCC, SLA, Leaf C N, LDW, LD, LH, and LA. The PERMANOVA results show F = 1.52 p = 0.05. Panel B: Another principal component analysis biplot shows root functional traits of maize across the same five phases. The x-axis is labeled Dim1 (31.3 percent) and the y-axis is labeled Dim2 (22.3 percent). Vectors represent trait loadings, and ellipses denote 95 percent confidence intervals for each phase. The traits include RNC, SRA, SRL, SBRT, RDMC, Root C N, and RLD. The PERMANOVA results show F = 2.58, p = 0.001.
In contrast, multivariate leaf trait expression did not differ across transition phases (F = 1.52, p = 0.05; Figure 2A). Here, the first two PCA components accounted for 29.9% and 19.3% of the total variation in leaf traits. Trait vectors such as SLA, leaf C:N, and LNC were the primary contributors along PCA axis 1, while LD, LDW, and LH aligned more closely with PCA axis 2. Despite high trait diversity, ellipses representing leaf trait expression at different transition phases showed considerable overlap, indicating less pronounced differentiation in leaf trait expression across phases.
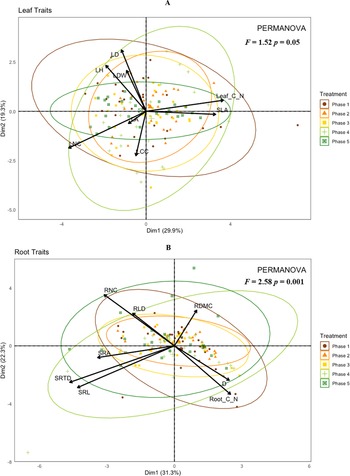
Hypervolume analysis shows that leaf traits (Figure 3A) show a much smaller spread of trait space than roots traits (Figure 3B), indicating that roots show a greater range of functional trait expression belowground. In addition, root trait space shows less crossover in transition phase volumes (Table 5), suggesting increasing differentiation of root trait strategies along the agroecological gradient.
Leaf (A) and root (B) trait hypervolumes across five agroecological phases, based on multiple bivariate functional trait axes (roots: log_SRL, log_D, log_SRTD, RNC, log_Root C:N; leaves: log_SLA, log_LA, log_Leaf_C_N, LNC, log_LDW). Coloured points represent observed trait combinations for each phase, with phase-specific centroids highlighted in white. Data were log-transformed where necessary and standardized to allow comparison of multivariate trait spaces across variables with differing units.

Mean hypervolumes of maize root and leaf trait space across agroecological transition phases

Hypervolumes represent the multidimensional trait space of maize root and leaf traits within each agroecological transition phase. Values are mean estimates from standardized trait data; higher values indicate greater functional trait diversity and dispersion.
Discussion
Maize functional traits vary across an agroecological transition
Our analysis of maize functional traits across five agroecological transition phases shows shifts in both root and leaf traits strategies, as farms transition from chemical-based farming systems to bio-inputs based agroecological management. Our findings support our hypotheses, demonstrating that root traits respond more sensitively to agroecological transitions than leaf traits and that functional niche space shifts along the agroecological management gradient. Morphological root traits, such as SRL and SRA, show an increase in values from phase 1 (conventional) to phases 4 and 5 (agroecological), suggesting a shift towards finer, more absorptive root systems that enhance resource acquisition under low-input organic conditions. High SRL and SRA are indicative of greater root foraging potential, enabling access to nutrient-rich microsites, but they also reflect a strategy associated with rapid turnover and short-lived absorptive roots (Cahill et al., Reference Cahill, McNickle, Haag, Lamb, Nyanumba and St. Clair2010; Lambers et al., Reference Lambers, Raven, Shaver and Smith2008). These trait strategies are advantageous in low-input natural farming systems where nutrient availability is dependent on organic matter mineralization and soil biotic interactions rather than relying on off-farm chemical inputs (Drinkwater and Snapp, Reference Drinkwater and Snapp2007; Heckman et al., Reference Heckman, Weil, Magdoff and Francis2009; Isaac et al., Reference Isaac, Nimmo, Gaudin, Leptin, Schmidt, Kallenbach, Martin, Entz, Carkner, Rajcan, Boyle and Lu2021). Moreover, in organic systems, nutrients are gradually released via microbial mineralization of organic amendments such as composts, manures, and cover crops (Gardner and Drinkwater, Reference Gardner and Drinkwater2009; Isaac et al., Reference Isaac, Nimmo, Gaudin, Leptin, Schmidt, Kallenbach, Martin, Entz, Carkner, Rajcan, Boyle and Lu2021; Yan et al., Reference Yan, Pan, Lavallee and Conant2020), enhancing SOM and soil biology, whereas synthetic fertilizers provide immediate availability but often reduce SOM and microbial biomass (Cayuela et al., Reference Cayuela, Sinicco and Mondini2009; Drinkwater and Snapp, Reference Drinkwater and Snapp2007; Gardner and Drinkwater, Reference Gardner and Drinkwater2009). These contrasting nutrient dynamics observed in organic and chemical-based systems strongly influence crop trait expression. Organic systems favour acquisitive root strategies such as higher SRL, thinner roots, and greater RNC that enhance exudation and SOM mineralization (Isaac et al., Reference Isaac, Nimmo, Gaudin, Leptin, Schmidt, Kallenbach, Martin, Entz, Carkner, Rajcan, Boyle and Lu2021), while chemical systems with readily available nutrients favour more conservative root traits with reduced reliance on microbial interactions.
In contrast, leaf trait expression patterns were less clear. For leaf traits, SLA and LA increased from phases 1 to 5, indicating an adaptation towards higher photosynthetic surface area with lower tissue investment, traits commonly associated with acquisitive resource-use strategies (Wright et al., Reference Wright, Reich, Westoby, Ackerly, Baruch, Bongers, Cavender-Bares, Chapin, Cornelissen, Diemer, Flexas, Garnier, Groom, Gulias, Hikosaka, Lamont, Lee, Lee, Lusk, Midgley, Navas, Niinemets, Oleksyn, Osada, Poorter, Poot, Prior, Pyankov, Roumet, Thomas, Tjoelker, Veneklaas and Villar2004) and identified by farmers as importnat (Isaac et al. Reference Isaac, Cerda, Rapidel, Martin, Dickinson and Sibelet2018). In contrast, traits such as LDW and LD remained stable or decreased slightly, reflecting a shift away from structural investment towards metabolic efficiency.
Our study provides empirical evidence that root traits are more responsive to agroecological transition phases than leaf traits. High intraspecific variation in root traits such as SRL, SRA, and RDMC, particularly in later transition phases, indicates enhanced plasticity and responsiveness to management-induced changes in soil conditions. These trends observed in our study strongly align with prior research studies, which indicate that root systems are key interfaces for sensing soil heterogeneity and shifting resource acquisition strategies under organic and low-input systems (Armstrong et al., Reference Armstrong, Miller, McAlvay, Ritchie and Lepofsky2021; Gaudin et al., Reference Gaudin, Tolhurst, Ker, Janovicek, Tortora, Martin and Deen2015; Lynch, Reference Lynch2013). Such root trait plasticity supports adaptive nutrient acquisition strategies, both through direct foraging and indirect mechanisms such as microbial recruitment, exudation, and enzymatic mobilization (Cantarel et al., Reference Cantarel, Pommier, Desclos-Theveniau, Diquélou, Dumont, Grassein, Kastl, Grigulis, Laîné, Lavorel, Lemauviel-Lavenant, Personeni, Schloter and Poly2015; Isaac et al., Reference Isaac, Nimmo, Gaudin, Leptin, Schmidt, Kallenbach, Martin, Entz, Carkner, Rajcan, Boyle and Lu2021; Meier et al., Reference Meier, Finzi and Phillips2017). These shifts are particularly crucial in natural farming systems that rely on ecological nutrient management rather than external inputs, where plant traits that foster tight nutrient cycling and minimal losses can enhance sustainability (Altieri, Reference Altieri1999; Tittonell, Reference Tittonell2020).
Root trait trade-offs indicate key functions
Root trait expression among the agroecological transition phases reflects functional trade-offs. While traits such as high SRL enhance nutrient exploration, SRL alone does not directly stimulate nutrient mineralization, which depends on rhizosphere processes mediated by root exudation and microbial activity. Mineralization is instead more strongly associated with trait combinations involving greater root biomass, higher root N content, and larger root diameters, which can promote increased exudation and microbial stimulation (Bargaz et al., Reference Bargaz, Noyce, Fulthorpe, Carlsson, Furze, Jensen, Dhiba and Isaac2017; Borden et al., Borden et al., Reference Borden, Anglaaere, Owusu, Martin, Buchanan, Addo-Danso and Isaac2020). Therefore, the wider variation observed in root traits is not only structural but coupled with nutrient acquisition mechanisms and possibly linked to enhanced root exudation and soil organic matter (SOM) mineralization (Guyonnet et al., Reference Guyonnet, Guillemet, Dubost and Haichar2018; Henneron et al., Reference Henneron, Kardol, Wardle, Cros and Fontaine2020). To this end, trait hyper volume differences across agroecological transition categories (Figure 3) are likely better suited to predict or explain trait-based responses to multifaceted belowground growing conditions verses single traits.
Moreover, SRA and SRL are key predictors of root–microbe symbioses, including associations with arbuscular mycorrhizal fungi and nitrogen-fixing microbes, which are critical in organic and low-input systems (Martin and Isaac, Reference Martin and Isaac2018; Wen et al., Reference Wen, Li, Shen, Tang, Xiong, Li, Pang, Ryan, Lambers and Shen2019). Thus, the observed variability reflects underlying ecological strategies, ranging from rapid resource capture to nutrient mobilization and microbial facilitation. Similar patterns have been reported by Lynch (Reference Lynch2013), who emphasized the importance of root architectural plasticity in nutrient acquisition and stress resilience under low-input systems.
Our results show that root D and RNC exhibit moderate variation (CVs = 31.2% and 39.4%, respectively), yet it is possible that even small shifts in these traits may modulate microbial activity in the rhizosphere by altering exudate quantity and quality. Importantly, lower root C:N ratios (mean = 51.0 ± 38.6; CV = 75.6%) may indicate an increase in labile carbon availability for microbial stimulation, a trait particularly relevant for nutrient cycling in organic systems (Herz et al., Reference Herz, Dietz, Gorzolka, Haider, Jandt, Scheel, Bruelheide and Weigelt2018; Jilling et al., Reference Jilling, Keiluweit, Contosta, Frey, Schimel, Schnecker, Smith, Tiemann and Grandy2018).
Trait coordination and trait space along an agroecological transition
Multivariate analyses revealed clear gradients in root traits. Conservative traits such as RDMC and root diameter loaded strongly in early phases, while acquisitive traits such as SRL and SRA were dominant in later phases. Similar trait-based PCA separation patterns have been reported in studies of functional trait change under land-use gradients (Laughlin and Messier, Reference Laughlin and Messier2015). Significant interactions between transition phase and region for several traits also point to the role of environmental filtering and local context in shaping trait expression (Garnier and Navas, Reference Garnier and Navas2011). Such findings highlight the need to account for spatial variation in agroecological systems when interpreting trait dynamics (de Bello et al., Reference de Bello, Vandewalle, Reitalu, Lepš, Prentice, Lavorel and Sykes2013).
Hypervolume analysis revealed meaningful ecological differentiation. Changes in hypervolume size reflect shifts in functional niche space. Expansion indicates increased functional diversity and ecological plasticity, suggesting enhanced adaptive capacity under agroecological management, whereas contraction or displacement signifies trait convergence or altered resource-use strategies. These patterns highlight how management transitions reshape crop ecological strategies (Blonder et al., Reference Blonder, Lamanna, Violle and Enquist2014; Blonder, Reference Blonder2018; Cornwell et al., Reference Cornwell, Schwilk and Ackerly2006). Notably, root trait space expanded markedly by phase 5, suggesting increased trait diversity under agroecological practices. Similarly, leaf hypervolumes peaked in phase 1 and remained larger in phases 4 and 5, possibly reflecting shifts in resource-use strategies. Smaller trait volumes in intermediate phases (2 and 3) and partial recovery in phases 4 and 5 point to a reorganization process where crop physiology adapts to reduced input and biologically enriched soil environments. Pairwise centroid distances, particularly between phase 1 and phase 5 (1.742 for root, 0.518 for leaf), further support functional divergence across the transition. These findings align with prior work (e.g., Blonder et al., Reference Blonder, Lamanna, Violle and Enquist2014; Díaz et al., Reference Díaz, Kattge, Cornelissen, Wright, Lavorel, Dray, Reu, Kleyer, Wirth, Colin Prentice, Garnier, Bönisch, Westoby, Poorter, Reich, Moles, Dickie, Gillison, Zanne, Chave, Joseph Wright, Sheremet’ev, Jactel, Baraloto, Cerabolini, Pierce, Shipley, Kirkup, Casanoves, Joswig, Günther, Falczuk, Rüger, Mahecha and Gorné2016; Prieto et al., Reference Prieto, Roumet, Cardinael, Dupraz, Jourdan, Kim, Maeght, Mao, Pierret, Portillo, Roupsard, Thammahacksa and Stokes2015), emphasizing that trait space volume and centroid shifts can reveal ecological differentiation even when multivariate tests are statistically non-significant. Hence, hypervolume-based approaches offer complementary insights into plant functional trait shifts during agroecological transitions. Similar to our study findings, recent crop-focused studies, for example, Nimmo et al. (Reference Nimmo, Violle, Entz, Rolhauser and Isaac2023), demonstrated that wheat root trait hypervolumes vary strongly with soil management, reflecting plasticity in foraging strategies, while Sauvadet et al. (Reference Sauvadet, Dickinson, Somarriba, Phillips-Mora, Cerda, Martin and Isaac2021) showed that cocoa functional trait space expands under agroecological management, capturing shifts in nutrient acquisition strategies, and Ajal et al. (Reference Ajal, Jäck, Vico and Weih2021) highlighted how intercropping alters trait hypervolumes, indicating complementary resource-use strategies among crop mixtures. Collectively, these studies confirm that hypervolume-based approaches provide complementary insights into plant functional differentiation across management systems and cropping strategies.
Our study provides novel insights into crop functional trait dynamics during agroecological transitions. While trait variation was associated with agroecological transition phases, potential influences of environmental heterogeneity inherent to smallholder farming systems may be present. Our research was conducted across 44 maize farms in central India, and while the sample captures real-world variability within smallholder systems, the findings are context specific and are not directly generalizable to other agroecological regions with differing climatic, edaphic, and socio-economic conditions. Further research across diverse geographic settings is necessary to validate and extend these results.
Conclusion
This study investigates the functional trait expression of maize in the context of agroecological transitions adopting natural farming across 44 farms in Madhya Pradesh, India. These farms represent three distinct farming systems: a) chemical-based (phase 1), b) transitioning (phases 2 and 3), and c) agroecological (phases 4 and 5). They examine a unique lens to explore how management practices influence trait variation and resource acquisition strategies across an agroecological transition gradient with distinct phases. By taking the traits-based approach and assessing traits such as root biomass, specific root length, and total root carbon and nitrogen, this study examines the patterns of functional trait variation in maize cropping systems under different agroecological transition phases. Importantly, the results presented here pertain to early and intermediate stages of agroecological transition focused on on-farm management changes.
The analysis of maize functional traits across distinct five agroecological transition phases shows that there are shifts in both root and leaf traits strategies as farms transition from chemical-based farming systems to bio-inputs based agroecological management.
This study underscores the importance of root traits and hypervolume-based metrics in capturing ecological transitions in agroecosystems. Our results suggest that agroecological management promotes a shift towards acquisitive belowground strategies, particularly in advanced phases of transition. Leaf traits, in contrast, remain more conservative. These findings align with broader theories in trait-based ecology and contribute to growing evidence that functional trait approaches can inform agroecological design and monitoring in diverse cropping systems. Root trait variability observed across the transition phases in maize not only reveals important physiological adjustments but also underlines their ecological importance for resilient, low-input agriculture.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0014479726100337.
Acknowledgements
This work would not have been possible without the selfless help of all the farmers that were integral to the study. We express our deep gratitude and appreciation for their participation. We thank the local NGO – Samaj Pragati Sahayog (SPS) and National Coalition of Natural Farmers, and the community resource personnel from SPS for logistics and support in the field.
Author contributions
AT and MEI designed the study; AT and SM conducted field work; AT wrote initial draft; AT, SM, OD, ARM, MEI edited draft; AT, ARM, MEI analysed data; OD and MEI secured research funds and supervised.
Funding statement
This research paper is funded by the International Doctoral Cluster Program, University of Toronto, an NSERC Discovery Grant to MEI, and the Sustainable Food and Farming Futures cluster at the University of Toronto Scarborough. Research was approved under the Nagoya protocol for microbial analysis on Indian soils.
Competing interests
The authors declare that they have no competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.





 Open access
Open access