


Introduction
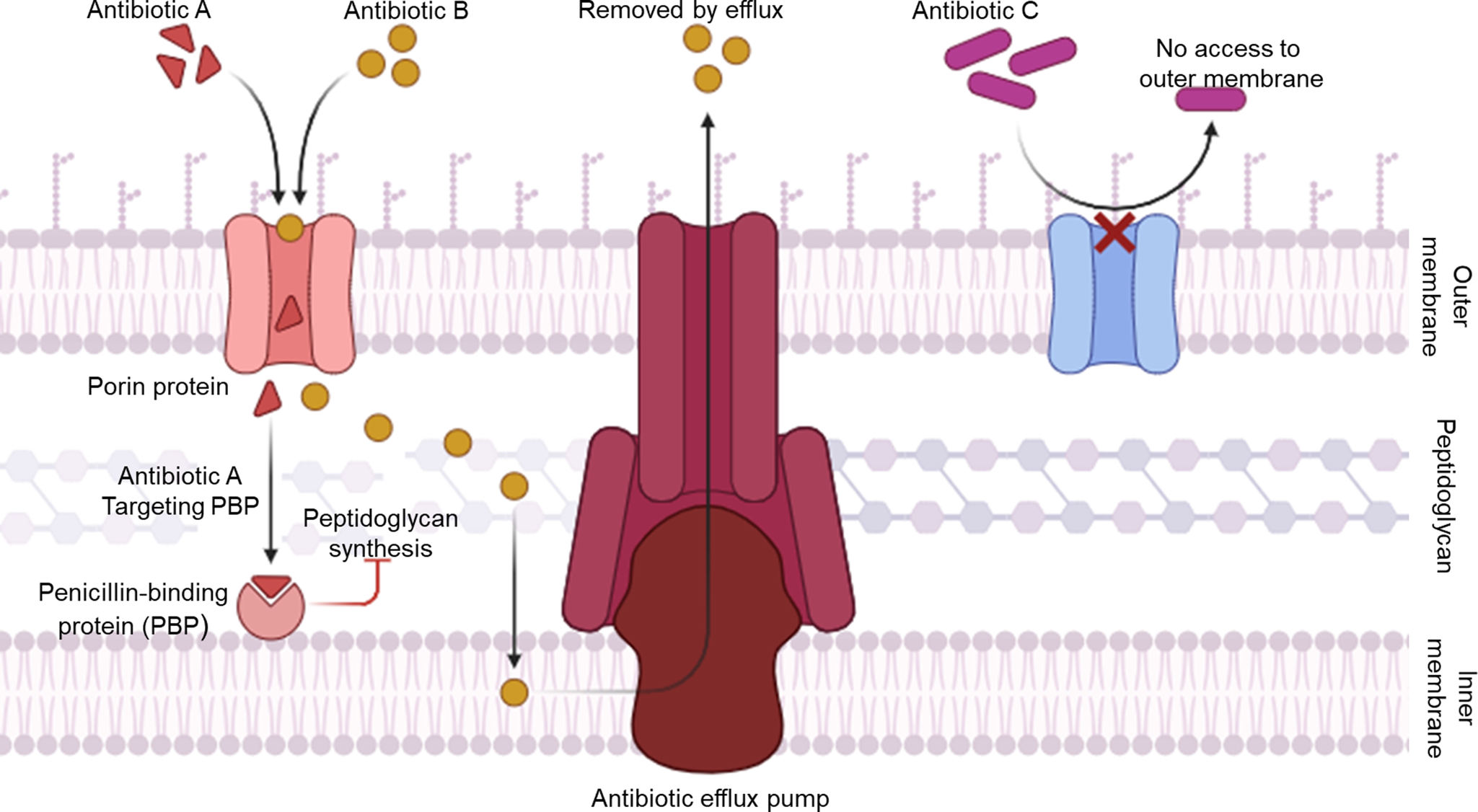
The rise of antibiotic-resistant bacteria poses a major global threat, reducing the effectiveness of current treatments (Ref. Reference Wall1). Mounting evidence indicates that modern medicine is nearing a post-antibiotic era where previously treatable infections may become untreatable (Ref. Reference Irwin2). Indiscriminate antibiotic use in medical, agricultural, and industrial settings has imposed strong selective pressure on human commensal and pathogenic microbes (Ref. Reference Serwecińska3). Amid this growing crisis of antimicrobial resistance, H. pylori stands out as a notably persistent and clinically consequential pathogen (Ref. Reference Tacconelli4). Antibiotic resistance in H. pylori arises primarily from mechanisms that limit drug penetration (e.g., mutations in porin genes such as hopB and hopC that reduce outer membrane permeability) (Ref. Reference De Francesco5), enhance active efflux via overexpression of efflux pumps like those in the RND family (e.g., hefABC) (Ref. Reference Gong6), or modify antibiotic targets within the bacterial cell wall, particularly through multiple point mutations in the pbp1 gene (e.g., Ser414Arg substitution adjacent to the STMK motif) (Ref. Reference Gerrits, Schuijffel, van Zwet, Kuipers, Vandenbroucke-Grauls and Kusters7), and changes in the C-terminal mosaic block of PBP1A, which reduce β-lactam binding affinity while preserving peptidoglycan synthesis (Ref. Reference Dore, Graham and Sepulveda8). As illustrated in Figure 1, these mechanisms collectively enable H. pylori to evade β-lactam antibiotics while maintaining essential cell wall integrity (Ref. Reference Hasanuzzaman, Bang and Gong9). Importantly, H. pylori resistance also prominently features point mutations in ribosomal and DNA gyrase targets that distinguish it from classic Gram-negative models. For clarithromycin (a macrolide), primary resistance stems from high-level point mutations in domain V of the 23S rRNA gene, most commonly A2143G and A2142G. These mutations sterically hinder drug binding to the peptidyl transferase centre, unlike the inducible erm methylase systems dominant in Enterobacteriaceae (Refs Reference Versalovic, Osato, Spakovsky, Dore, Reddy, Stone, Shortridge, Flamm, Tanaka and Graham10, Reference Kavya11). For fluoroquinolones (e.g., levofloxacin), resistance arises via point mutations in the quinolone resistance-determining region (QRDR) of gyrA, particularly N87K and D91N substitutions in helix 4 region. These mutations prevent drug docking to the DNA-gyrase cleavage complex. This mirrors Gram-negative mechanisms but occurs at exceptionally high prevalence (>90% in some regions) due to H. pylori’s single gyrA copy and lack of plasmid-mediated qnr genes (Refs Reference Lee, Thi, Ha, Choi, Kim, Trinh, Anh, Kang, Park, Byun, Lee, Jung, Boonyanugomol, Baik, Kang and Shin12, Reference Salehi, Attaran, Eskini, Esmaeili, Sharifirad, Sadeghi and Mohammadi13). Collectively, these adaptations compromise the effectiveness of conventional antibiotic regimens and underscore the necessity of developing complementary non-antibiotic interventions such as natural and synthetic compounds, probiotics and vaccines to overcome these barriers and achieve sustainable eradication of H. pylori infections.
Mechanisms of antibiotic resistance in Helicobacter pylori. The diagram illustrates three major bacterial defense strategies: (1) decreased permeability where mutations in porin proteins prevent entry of antibiotics (Antibiotic C); (2) active efflux where efflux pumps expel drugs from the cytoplasm (Antibiotic B) and (3) target modification where antibiotics that enter the cell (Antibiotic A) bind to penicillin-binding proteins (PBPs), inhibiting peptidoglycan synthesis, though alterations in PBPs can confer resistance. Together, these mechanisms limit intracellular antibiotic accumulation and effectiveness.

Noted for its persistence in the stomach’s harsh environment, H. pylori is a major cause of chronic gastritis, peptic ulcer disease and gastric cancers, including adenocarcinoma and MALT lymphoma (Ref. Reference Haley and Gaddy14). Its pathogenic success is largely attributed to its adaptive mechanisms that enable survival in acidic and immune-challenging niches (Ref. Reference Fagoonee and Pellicano15). Key among these survival strategies are the production of urease (Ref. Reference Almarmouri, el-Gamal, Haider, Hamad, Qumar, Sebastian, Ghemrawi, Muhammad, Burucoa and Khoder16) which neutralizes gastric acid and facilitates colonization; morphological transformation into coccoid forms, which represent a stress-adaptive state enabling persistence, immune evasion, antibiotic tolerance and potential resuscitation leading to treatment failure and recurrence (Refs Reference Sadeghloo, Fatemi, Raeisi, Azizmohammad Looha and Sadeghi17, Reference Ramezani, Sadeghloo, Azizmohammad Looha and Sadeghi18), and the development of protective biofilms, within which coccoid forms often predominate and confer up to 1000-fold increased tolerance to antibiotics through limited drug penetration, efflux pump overexpression and dormancy (Refs Reference Sadeghloo, Fatemi, Raeisi, Azizmohammad Looha and Sadeghi17, Reference Krzyżek, Grande, Migdał, Paluch and Gościniak19). All of which have recently garnered increased attention due to their contribution to resistance to treatment.
Recent insights indicate the ability of non-antibiotic agents to enhance the therapeutic efficacy of conventional antimicrobial regimens when used in combination (Ref. Reference Laws, Shaaban and Rahman20).
Given these promising developments, a growing number of studies are now focused on identifying alternative or adjunctive strategies that can overcome H. pylori’s resilience. This review synthesizes recent evidence from major databases on non-antibiotic strategies against H. pylori, emphasizing studies from the past decade that explore their mechanisms and potential to overcome antibiotic resistance. By examining recent advances in alternative treatments and their potential to counter antibiotic resistance, this review highlights key directions for future research and clinical application.
Current antibiotic therapies and resistance patterns
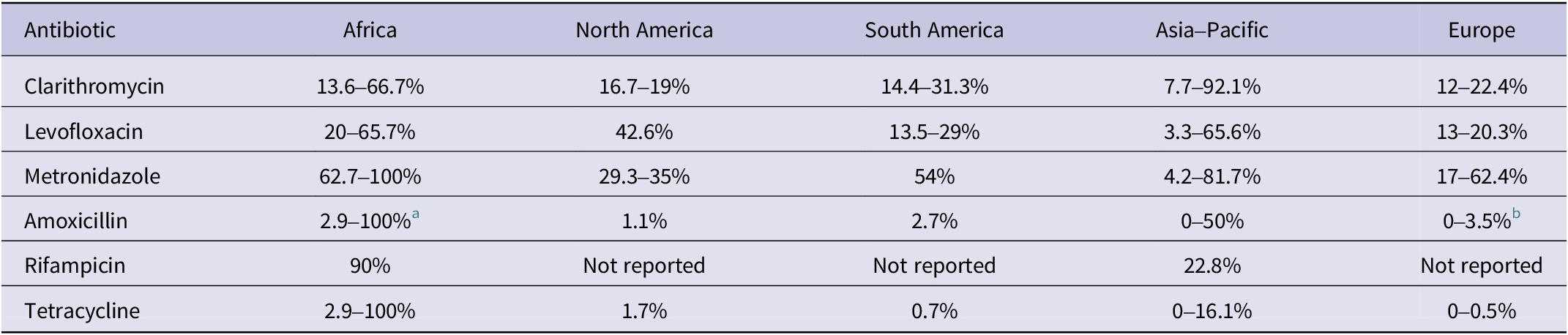
Standard H. pylori eradication therapy typically relies on proton pump inhibitor (PPI)-based regimens. Among these, bismuth quadruple therapy, which combines bismuth, tetracycline, metronidazole and a PPI, is recommended as a first-line option because of its strong efficacy even against resistant strains, although it remains unavailable to large populations in many regions worldwide (Ref. Reference Schulz, Liou, Alboraie, Bornschein, Campos Nunez, Coelho, Quach, Fallone, Chen, Gerhard, Gisbert, Jung, Katelaris, Kim, Lu, Macke, Mahachai, Moss, Remes Troche, Riquelme, Romano, Setshedi, Smith, Suerbaum, Tshibangu-Kabamba, Vilaichone, Yadegar, Yamaoka, Mégraud, el-Omar, Sugano and Malfertheiner21). Alternative regimens include rifabutin triple therapy (PPI, amoxicillin and rifabutin), generally used as a rescue option, and vonoprazan-based dual therapy (vonoprazan and amoxicillin), which improves eradication through stronger acid suppression. Clarithromycin- and levofloxacin-based treatments are now limited to cases guided by antibiotic susceptibility testing (AST) due to declining efficacy. From 2018 to 2023, resistance rates exceeded 15% for clarithromycin in 24 out of 31 countries and for levofloxacin in 18 out of 31 countries, with continental ranges summarized in Table 1 (Ref. Reference Schulz, Liou, Alboraie, Bornschein, Campos Nunez, Coelho, Quach, Fallone, Chen, Gerhard, Gisbert, Jung, Katelaris, Kim, Lu, Macke, Mahachai, Moss, Remes Troche, Riquelme, Romano, Setshedi, Smith, Suerbaum, Tshibangu-Kabamba, Vilaichone, Yadegar, Yamaoka, Mégraud, el-Omar, Sugano and Malfertheiner21). Although WHO removed clarithromycin-resistant H. pylori from its priority list in 2024 (Ref. 22), the decision remains debated as resistance still exceeds 15% in most regions (Ref. Reference Schulz, Liou, Alboraie, Bornschein, Campos Nunez, Coelho, Quach, Fallone, Chen, Gerhard, Gisbert, Jung, Katelaris, Kim, Lu, Macke, Mahachai, Moss, Remes Troche, Riquelme, Romano, Setshedi, Smith, Suerbaum, Tshibangu-Kabamba, Vilaichone, Yadegar, Yamaoka, Mégraud, el-Omar, Sugano and Malfertheiner21). Despite this delisting, the pathogen retains major clinical importance due to its prevalence and links to severe diseases.
Continental ranges of Helicobacter pylori antibiotic resistance rates (2018–2023)

a Rates over 90% have been observed in some countries within this region.
b Remaining under 2% in 14 of 31 evaluated countries globally.
Natural or synthetic compounds against H. pylori
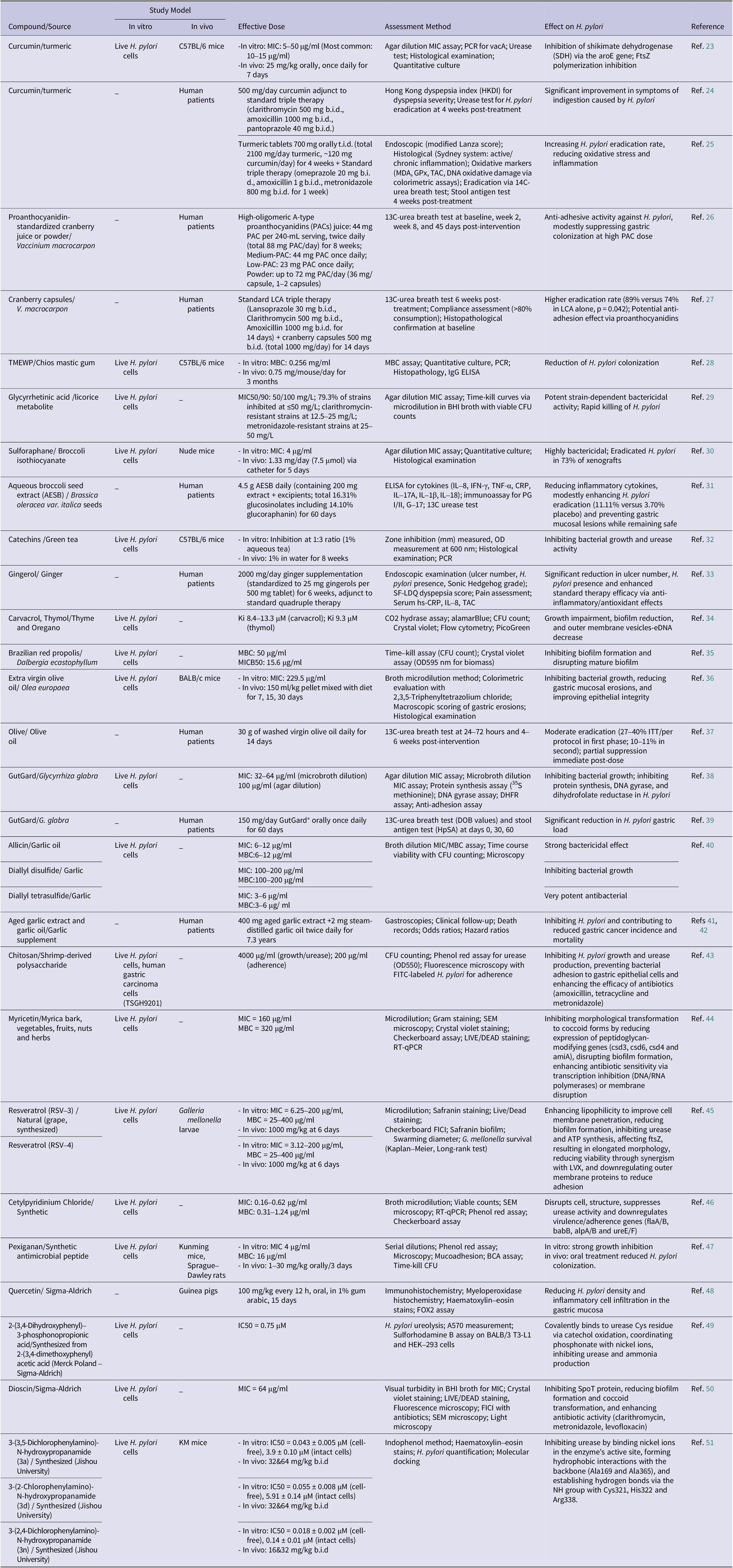
Natural products from medicinal plants, functional foods and dietary sources are increasingly reconsidered as potential anti-H. pylori agents, while several synthetic compounds have also shown promise as novel therapeutic candidates. Emerging evidence supports their integration into combination therapies to enhance eradication outcomes and limit antibiotic resistance. This section summarizes the most relevant natural and synthetic options reported in recent literature (Table 2).
Overview of natural and synthetic compounds with anti-Helicobacter pylori activity

Abbreviations: MIC, Minimum Inhibitory Concentration; PCR, Polymerase Chain Reaction; vacA, Vacuolating Cytotoxin A; SDH, Shikimate Dehydrogenase; aroE, Gene encoding shikimate dehydrogenase; FtsZ, Filamenting Temperature-Sensitive Mutant Z; HKDI, Hong Kong dyspepsia index; b.i.d., Bis in Die (twice daily); PACs, Proanthocyanidins; ITT, Intention-To-Treat; MBC, Minimum Bactericidal Concentration; IgG, Immunoglobulin G; ELISA, Enzyme-Linked Immunosorbent Assay; Ki, Inhibition Constant; CFU, Colony-Forming Unit; OD, Optical Density; BHI, Brain Heart Infusion; AESB, Aqueous broccoli seed extract; CRP, C-reactive protein; PG, Pepsinogen (PG I/II); G-17, Gastrin-17; MDA, Malondialdehyde; GPx, Glutathione peroxidase; TAC, Total anti-oxidant capacity; hs-CRP, High-sensitive C-reactive protein; SF-LDQ, Short-Form Leeds Dyspepsia Questionnaire; eDNA, extracellular DNA; DOB, Delta Over Baseline; HpSA, H. pylori Stool Antigen; FITC, Fluorescein Isothiocyanate; SEM, Scanning Electron Microscopy; RT-qPCR, Real-Time Quantitative Polymerase Chain Reaction; FICI, Fractional Inhibitory Concentration Index; LVX, Levofloxacin; flaA/B, Flagellin A/B; babB, Blood Group Antigen-Binding Adhesin B; alpA/B, Adherence-Associated Lipoprotein A/B; ureE/F, Urease Accessory Proteins E/F; BCA, Bicinchoninic Acid Assay; FOX2, Histochemical assay for oxidative stress; A570, Absorbance at 570 nm; SpoT, Stringent Response Protein; IC50, Half-maximal Inhibitory Concentration; DHFR, Dihydrofolate Reductase.
To provide a rigorous evaluation of these compounds, we applied a hierarchical framework for critical appraisal. Evidence was categorized into three levels: (1) in vitro studies, providing mechanistic insights; (2) Animal models (In vivo), assessing biological plausibility and safety and (3) Human clinical trials, representing the highest level of clinical relevance. Throughout this section, priority is given to human data when available, while acknowledging that many promising compounds currently remain at the preclinical stage.
Brazilian red propolis stands out as a potent natural compound. Santiago et al. (Ref. Reference Liu, Shi, Ren, Ni, Li, Chen, Liu, Yuan, He, Liu, Cao, Yang, Xiao and Zhu51) showed that Brazilian red propolis extract inhibited H. pylori growth (MIC 50–100 μg/ml) with matching bactericidal concentrations. It eliminated bacteria within 24 hours at 50 μg/ml, prevented biofilm formation, and eradicated existing biofilms in vitro. In Wistar rats, oral doses (18.25–300 mg/kg) reduced gastric H. pylori loads and modulated inflammation at higher doses. Extra virgin olive oil (EVOO), rich in phenolic compounds, has also shown promise. Arismendi Sosa et al. (Ref. Reference Arismendi Sosa, Mariani, Vega and Penissi36) found that EVOO, hydroxytyrosol and oleuropein inhibited H. pylori growth in vitro using the broth microdilution method. In vivo, oral administration of EVOO to BALB/c mice infected with H. pylori reduced bacterial colonization and prevented gastric mucosa lesions, highlighting its therapeutic potential. Earlier, Romero et al. (Ref. Reference Santiago, Leandro, Rosa, Silva, Teixeira, Servato, Ambrósio, Veneziani, Aldana-Mejía, Bastos and Martins52) demonstrated that olive oil phenolics, particularly the dialdehydic form of decarboxymethyl ligstroside aglycon, exerted strong bactericidal effects against eight H. pylori strains, including antibiotic-resistant ones, at concentrations as low as 1.3 μg/ml. Complementing these preclinical insights, a pilot clinical study by Castro et al. (Ref. Reference Castro, Romero, de Castro, Vargas, Medina, Millán and Brenes37) evaluated the efficacy of EVOO in eradicating H. pylori in 60 infected adults. Participants received 30 g of washed virgin olive oil daily for 14 days, followed by unwashed oil after a washout period. The urea breath test showed eradication rates of 27% by intention-to-treat and 40% per protocol after the first phase, dropping to 10% and 11% after the second, with some participants withdrawing due to taste or nausea, underscoring moderate clinical effectiveness warranting further optimization. Asha et al. (Ref. Reference Asha, Debraj, Prashanth, Edwin, Srikanth, Muruganantham, Dethe, Anirban, Jaya, Deepak and Agarwal38) investigated the in vitro antibacterial activity of a flavonoid-rich extract from Glycyrrhiza glabra (licorice), commercially known as GutGard®. The extract inhibited H. pylori through multiple mechanisms, including protein synthesis inhibition (evidenced by reduced 35S methionine incorporation), DNA gyrase inhibition (IC50: 4.40 μg/ml) and dihydrofolate reductase inhibition (IC50: 3.33 μg/ml). Glabridin, the primary flavonoid, was particularly effective, though the extract showed limited impact on bacterial adhesion to gastric cells. This suggests licorice could target critical bacterial processes, warranting further clinical investigation. Building on this, Puram et al. (Ref. Reference Puram, Suh, Kim, Bethapudi, Joseph, Agarwal and Kudiganti39) assessed GutGard® (150 mg/day for 60 days) in 107 H. pylori-positive adults. Using the 13C-urea breath test and stool antigen test, the study found 56% negativity in the treatment group versus 4% in the placebo group by stool antigen test, and 48% versus 2% by 13C-UBT, with significant reductions in gastric load and good tolerability, affirming its clinical promise as a standalone management option.
Garlic derivatives have a storied history of antimicrobial use. O’Gara et al. (Ref. Reference O’Gara, Hill and Maslin40) demonstrated in vitro that garlic oil exhibited strong H. pylori inhibition (MIC 8–32 μg/ml and MBC 16–32 μg/ml), outperforming garlic powder (MIC/MBC 250–500 μg/ml). Diallyl tetrasulfide, the most potent garlic constituent (MIC/MBC 3–6 μg/ml), rapidly reduced bacterial motility and induced morphological changes, suggesting potential for intragastric delivery. However, a clinical trial by Zardast et al. (Ref. Reference Romero, Medina, Vargas, Brenes and de Castro53) found that oral garlic (3 g twice daily) did not significantly reduce H. pylori urease activity in patients, indicating limited clinical efficacy. In a follow-up of a randomized placebo-controlled intervention trial, Li et al. (Ref. Reference Li, Zhang, Ma, Li, Zhang, Zhang, Guo, Zhou, Li, Shen, Liu, Han, Blot, Gail, Pan and You41) in 2019 assessed the long-term effects of garlic supplementation (extract and oil for 7.3 years) on gastric cancer prevention in a high-risk population, finding that it significantly reduced gastric cancer mortality (hazard ratio 0.66, 95% CI 0.43–1.00) over 22 years, with effects appearing later in the follow-up period. Another secondary analysis of the same trial by Guo et al. (Ref. Reference Guo, Li, Zhang, Ma, Zhang, Zhang, Zhou, Liu, Han, Li, Pan and You42) in 2020 showed that the protective effect of garlic supplementation on gastric cancer mortality was stronger among non-alcohol drinkers (hazard ratio 0.33, 95% CI 0.15–0.75) and participants with H. pylori infection. Green tea catechins, particularly epigallocatechin-3-gallate (EGCG), have been shown to disrupt H. pylori cell membranes and inhibit urease activity in vitro. In Mongolian gerbil models, EGCG treatment reduced bacterial colonization and gastric inflammation. Similarly, the bioactive components of ginger, including 6-gingerol and 6-shogaol, inhibit H. pylori adhesion to gastric epithelial cells and block Toll-like receptor 4 (TLR4) activation in vitro (Ref. Reference Zardast, Namakin, Esmaelian Kaho and Hashemi54). Extending these observations, Arefpour et al. (Ref. Reference Arefpour, Sadeghi, Zayeri and Hekmatdoost33) carried out a clinical intervention in which peptic ulcer patients received ginger supplementation (2 g/day) in combination with standard quadruple therapy for 6 weeks. Endoscopic analyses revealed notable reductions in ulcer count, H. pylori colonization, Sonic Hedgehog (Shh) expression, and dyspepsia severity compared with the placebo group. Furthermore, serum levels of high-sensitive C-reactive protein (hs-CRP) and IL-8 significantly declined, while total antioxidant capacity increased, supporting the role of ginger’s anti-inflammatory and antioxidant properties in improving gastric mucosal health. Curcumin from turmeric damages H. pylori cell membranes and inhibits urease activity in vitro. A clinical study by Khonche et al. (Ref. Reference Khonche, Biglarian, Panahi, Valizadegan, Soflaei, Ghamarchehreh, Majeed and Sahebkar24) in 2016 evaluated adjunctive curcumin (500 mg/day) with standard triple therapy (clarithromycin, amoxicillin and pantoprazole) for peptic ulcer patients, showing no enhancement in H. pylori eradication rates (73.3% in both groups) but significant improvement in dyspepsia symptoms (Hong Kong dyspepsia index change: −12.90 versus −9.60, p < 0.001) and higher symptom resolution (27.6% versus 6.7%, p = 0.042). In another clinical trial by Judaki et al. (Ref. Reference Judaki25) in 2017, curcumin combined with triple therapy in patients with chronic gastritis associated with H. pylori significantly increased eradication rates (p < 0.05), reduced oxidative damage to DNA (p < 0.05), and decreased inflammation scores compared to triple therapy alone. Cranberry juice, rich in polyphenols, inhibits H. pylori adhesion to gastric mucosa, with human studies showing modest reductions in colonization. Li et al. (Ref. Reference Li, Ma, Guo, Liu, Li, Zhang, Zhang, Zhou, Zhang, Gao, Guo, Ye, Li, You and Pan26) in 2021 tested proanthocyanidin-standardized cranberry juice (44 mg proanthocyanidins per 240 mL, twice daily) for 8 weeks in 522 H. pylori-positive adults, achieving a 20% H. pylori-negative rate compared to lower doses and placebo (p < 0.05), with a positive trend over time and good tolerability. In a subsequent study, Seyyedmajidi et al. (Ref. Reference Seyyedmajidi, Ahmadi, Hajiebrahimi, Seyedmajidi, Rajabikashani, Firoozabadi and Vafaeimanesh27) assessed the efficacy of adding cranberry capsules (500 mg twice daily) to lansoprazole–clarithromycin–amoxicillin triple therapy for 14 days in 200 peptic ulcer patients, observing a significantly higher eradication rate (89% versus 74%, p = 0.042). Broccoli sprouts, containing sulforaphane, induce detoxifying enzymes and reduce H. pylori colonization in animal and human studies, offering a dietary approach to infection management (Ref. Reference Takeuchi, Trang, Morimoto, Nishida, Matsumura and Sugiura55). In a 2-month intervention study, Guo et al. (Ref. Reference Guo, Wang, Mahe, Li, Jiao, Wang, Xie, Liu, Zeng, Hu and Jing31) evaluated the effects of an aqueous broccoli seed extract (AESB) in 110 H. pylori-infected patients without atrophic gastritis. The treatment led to reductions in inflammatory cytokines (IL-8 by 9.02 pg/mL; IFN-γ by 5.08 pg/mL) and in biomarkers such as pepsinogen I (by 24.56 ng/mL). Although the H. pylori eradication rate was modest (11.11% versus 3.70% in the placebo group) and did not reach statistical significance, no treatment-related adverse events were reported.
Myricetin (MYR), a plant-derived polyphenol, exhibits anti-virulence activity against H. pylori. In an in vitro study Krzyżek et al. (Ref. Reference Krzyżek, Migdał, Paluch, Karwańska, Wieliczko and Gościniak44) reported MICs of 160 μg/ml for H. pylori strains J99 and Tx30a, with sub-MICs (1/16–1/4 MIC) slowing the spiral-to-coccoid transformation, maintaining 1.5–4 times more spiral forms after 7–14 days under serum-starvation conditions. MYR at 1/4 MIC reduced biofilm formation by 70% and synergized with amoxicillin, clarithromycin, tetracycline, metronidazole, and levofloxacin, lowering their MICs 4–16-fold, even in clarithromycin- and metronidazole-resistant strains. RT-qPCR showed MYR downregulated morphogenesis genes (csd3, csd6 and amiA) by 4–6-fold, suggesting interference with peptidoglycan remodelling as a key mechanism. Similarly, Grande et al. (Ref. Reference Grande, Carradori, Puca, Vitale, Angeli, Nocentini, Bonardi, Gratteri, Lanuti, Bologna, Simeone, Capasso, de Luca and Supuran56) demonstrated that carvacrol and thymol from thyme and oregano oils selectively inhibit H. pylori α- and β-carbonic anhydrases with inhibition constants in the low micromolar range, while sparing human isoforms. At subinhibitory concentrations, these compounds hindered bacterial growth, reduced biofilm formation and lowered outer membrane vesicle and extracellular DNA release. Computational docking supported their binding to bacterial enzyme sites. Unlike amoxicillin, they were non-toxic to beneficial gut bacteria (Lactobacillus acidophilus, Lactobacillus rhamnosus and Lactobacillus reuteri) at effective concentrations, suggesting potential as adjunct therapies to combat H. pylori antibiotic resistance while preserving gut microflora. These findings highlight MYR’s potential to enhance antibiotic efficacy and reduce virulence.
Resveratrol (RSV), a polyphenol derived from grapes, and its analogues RSV-3 and RSV-4 exhibit potent in vitro anti-H. pylori activity. Di Fermo et al. (Ref. Reference Di Fermo45) reported RSV MICs of 200–800 μg/ml against resistant strains, while RSV-3 and RSV-4, designed for better bioavailability, had MICs of 6.25–200 μg/ml and 3.12–200 μg/ml respectively. These derivatives, combined with levofloxacin (LVX), reduced LVX MICs by up to eightfold in two resistant strains. They also curbed biofilm formation (RSV-4 up to 84.8% at sub-MIC levels) and limited bacterial motility, making them promising for combination therapies.
Cetylpyridinium chloride (CPC), a synthetic quaternary ammonium compound, shows significant in vitro activity. Xun et al. (Ref. Reference Xun, Feng, Li, Yao, Wang, Wei, Jia, Fan, Shi, Lv and Zhang46) reported MICs of 0.16–0.62 μg/ml and MBCs of 0.31–1.24 μg/ml against H. pylori. CPC induced morphological and physiological changes, suppressed virulence and adherence genes (e.g., flaA, flaB and ureE), and inhibited urease activity, positioning it as a promising alternative. Its use in oral antiseptics suggests potential for gastrointestinal applications, though clinical data are lacking.
These findings suggest that natural and synthetic compounds hold potential as alternatives or adjuncts to antibiotic therapies for H. pylori. From a translational perspective, a consistent gap exists between potent in vitro activity and clinical effectiveness for several phytochemicals. For instance, allicin and other organosulfur compounds in garlic exhibit rapid bactericidal effects under laboratory conditions; however, allicin is chemically unstable, rapidly metabolized and poorly bioavailable after oral ingestion, limiting its capacity to reach sustained intragastric bactericidal concentrations in humans (Ref. Reference Deng, Ho, Lan, Xiao and Lu57). Similarly, curcumin demonstrates membrane-disruptive and urease-inhibitory properties in vitro, yet its clinical utility is hindered by extremely low systemic bioavailability, rapid intestinal metabolism, and poor aqueous solubility, necessitating doses far exceeding typical dietary intake to achieve therapeutic plasma or tissue levels (Ref. Reference Bertoncini-Silva, Vlad, Ricciarelli, Giacomo Fassini, Suen and Zingg58). Polyphenols such as resveratrol and EGCG also undergo extensive first-pass metabolism and exhibit limited gastric stability, which may attenuate their antimicrobial potency in vivo despite strong preclinical performance (Ref. Reference Salla, Karaki, el Kaderi, Ayoub, Younes, Abou Chahla, Baksh and el Khatib59). Therefore, while these compounds provide valuable mechanistic insights and may serve as adjunctive agents, their translation into standalone therapies requires optimized delivery systems (e.g., nanoformulations, encapsulation or gastro-retentive platforms) to overcome pharmacokinetic and stability-related barriers.
Modulation of the gut microbiota: the role of specific probiotic strains against H. pylori
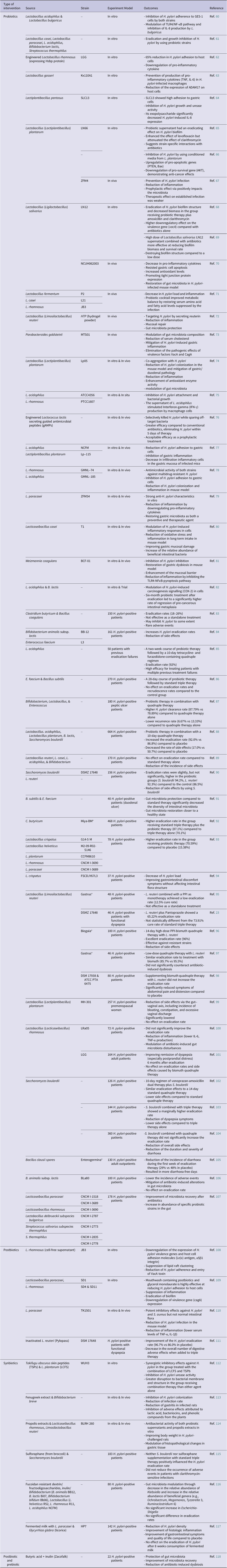
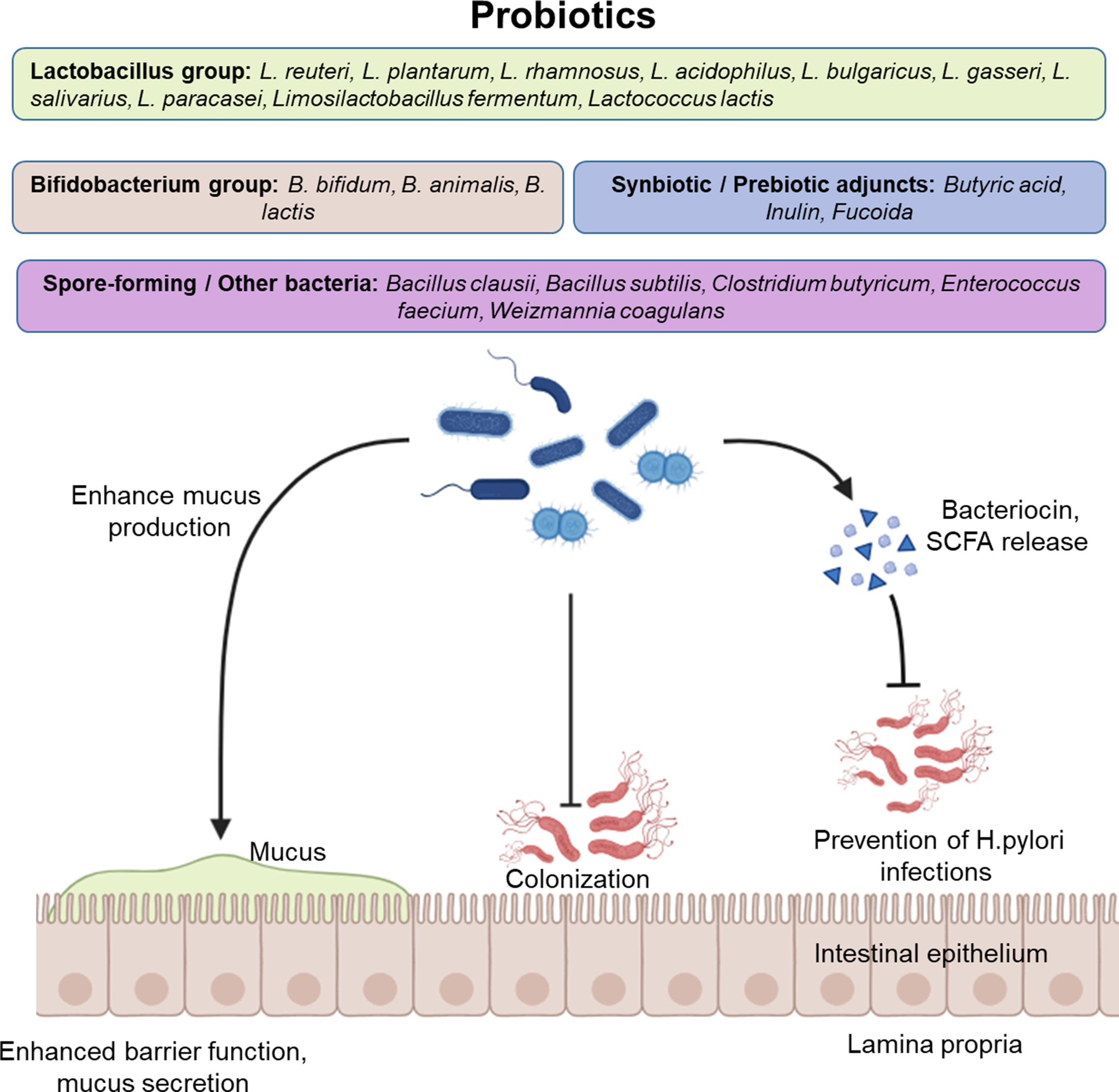
Probiotics have emerged as a promising adjunct in H. pylori management (Table 3 and Figure 2). While they are not effective as standalone treatments, evidence shows that their use alongside standard antibiotic regimens can enhance eradication rates, reduce relapse (Refs Reference Lim, Choi and Chung86, Reference Marinelli, Scalese, Covelli, Ruffa, Bedetti, Bruno and Severi101) and mitigate antibiotic-related side effects (Ref. Reference Han, Li, Nan, Zhou, Li and Li106).
Microbiota-based strategies for Helicobacter pylori eradication and management

Abbreviations: gAMPs: Guided Antimicrobial Peptides; COX-2: Cyclooxygenase-2; IFN-γ: Interferon-gamma; TLR4: Toll-Like Receptor 4; NF-κB: Nuclear Factor kappa B; IL-8: Interleukin-8; IL-6: Interleukin-6; TNF-α: Tumour Necrosis Factor-alpha; PTEN: Phosphatase and Tensin Homolog; Bax: Bcl-2-associated X protein; AKT: Protein Kinase B; ADAM17: A Disintegrin and Metalloproteinase 17; PPI: Proton Pump Inhibitor; Le(x): Lewis x Antigen; α5β1: Alpha-5 Beta-1 Integrin; cagA: Cytotoxin-Associated Gene A; VacA: Vacuolating Cytotoxin A; BCF-01: Bacillus coagulans Form-01; DSMZ: Deutsche Sammlung von Mikroorganismen und Zellkulturen; ATCC: American Type Culture Collection; PTCC: Persian Type Culture Collection; CNCM: Collection Nationale de Cultures de Microorganismes; HTP: Hydrogel Powder; Miya-BM®: C. butyricum Brand; Biogaia®: L. reuteri Brand; Gastrus®: L. reuteri Brand; Enterogermina®: B. clausii Brand; MTS01: Parabacteroides goldsteinii Strain; BLa80: B. animalis subsp. lactis Strain; LN66: Lactiplantibacillus plantarum Strain; Lp05: Lactiplantibacillus plantarum Strain; MH-301: Lactiplantibacillus plantarum Strain; ZFM4: Lactiplantibacillus plantarum Strain; LN12: Ligilactobacillus salivarius Strain; NCUH062003: Ligilactobacillus salivarius Strain; LRa05: Lacticaseibacillus rhamnosus Strain; LGG: L. rhamnosus GG; Kx110A1: L. gasseri Strain; ZFM54: L. paracasei Strain; MN-LF23: Limosilactobacillus fermentum Strain; T1: Lacticaseibacillus casei Strain; SLC13: Lactiplantibacillus pentosus Strain; BLRH 260: Probiotic and Propolis Combination; HP7: L. paracasei and G. glabra Combination; WUH3: L. plantarum Strain; TSPs: Takifugu obscurus Skin Peptides; LCFS: L. plantarum Cell-Free Supernatant; Zacofalk: Butyric Acid and Inulin Combination
Protective mechanisms of probiotic and synbiotic strains against Helicobacter pylori infection. Beneficial bacterial groups – primarily Lactobacillus, Bifidobacterium and spore-forming genera such as Bacillus and Clostridium – colonize the gastric and intestinal mucus layer, releasing bacteriocins and short-chain fatty acids (SCFAs) that inhibit pathogenic colonization and viral infection. Synbiotic and prebiotic adjuncts such as butyric acid, inulin and fucoidan further enhance microbial survival, adhesion and immune modulation, maintaining epithelial integrity and reducing inflammation.

Different approaches have been taken to utilize probiotics in H. pylori treatment. Some studies have used various strains of probiotic bacteria as the primary agent to eradicate H. pylori. Although some studies have found probiotics to be more effective than a placebo (Ref. Reference Wang93) or non-inferior to antibiotic therapy (Ref. Reference Muresan, Pop and Dumitrascu23), in most cases, monotherapy with probiotics was not able to achieve clinical eradication (Refs Reference Zhang, Guo, Li, Chen, Liu, Feng, He, Zhao, Zhang, Chen and Shi83, Reference Dore95). Probiotics did, however, reduce the infection load of H. pylori and alleviate symptoms, and lactic acid bacteria were among the most successful strains (Ref. Reference Hong, Wang, Zhang, Liu and Liu94). Lai et al. (Ref. Reference Lai, Shen, Wang, Ouyang, Zhang, Hu, Zhang, Li, Xu and Zhao72) utilized a novel powder that forms a hydrogel in the digestive tract and were able to increase the survival of L. reuteri and reduce H. pylori-induced inflammation in a C57BL/6 mouse model. Choudhury et al. (Ref. Reference Choudhury, Ortiz, Young, Mahmud, Stoffel, Greathouse and Kearney76) engineered Lactococcus lactis to secrete antimicrobial peptides specifically targeted towards H. pylori and were able to eliminate it in 5 days in a mouse model via oral gavage, while preserving the off-target bacteria of gut microbiota. These findings suggest that while probiotics are still not very formidable in eradicating H. pylori on their own, they can be helpful as an adjunct to antibiotic therapies. In contrast, adjunct probiotics with antibiotics enhance eradication efficacy in clinical studies, especially for well-supported strains. For example, L. reuteri in quadruple/dual regimens achieved 85.7–96% eradication, even against resistant strains, while reducing antibiotic doses (Refs Reference Poonyam, Chotivitayatarakorn and Vilaichone96, Reference Dore, Sau, Niolu, Abbondio, Tanca, Bibbò, Loria, Pes and Uzzau97). Similarly, Saccharomyces boulardii boosted rates marginally (94.2%) and excelled in side-effect reduction (Ref. Reference Naghibzadeh, Salmani, Nomiri and Tavakoli90). Other robust combinations include Clostridium butyricum (87.1% eradication) (Ref. Reference Mukai, Handa, Suyama, Majima and Naito92), multi-strain probiotics (92.0%) (Refs Reference Viazis, Argyriou, Kotzampassi, Christodoulou, Apostolopoulos, Georgopoulos, Liatsos, Giouleme, Koustenis, Veretanos, Stogiannou, Moutzoukis, Poutakidis, Mylonas, Tseti and Mantzaris88, Reference Wang, Wei, Liu, Wang, Zhang and du116) and specifically the blends of Bifidobacterium, Lactobacillus and Enterococcus (87.79%) (Ref. Reference Zhou and Li87).
Another way of utilizing probiotics in H. pylori treatment strategies is along with antibiotic therapy. Studies have been exploring pretreatment or co-treatment of probiotics and antibiotics to increase eradication rates and relapse prevention, especially in antibiotic-resistant strains of H. pylori. Higher efficiency of co-treatment of probiotics and antibiotics was demonstrated across various experimental and clinical settings. In a rat model, Viazis et al. (Ref. Reference Viazis, Argyriou, Kotzampassi, Christodoulou, Apostolopoulos, Georgopoulos, Liatsos, Giouleme, Koustenis, Veretanos, Stogiannou, Moutzoukis, Poutakidis, Mylonas, Tseti and Mantzaris88) showed that a multi-strain probiotic (comprising Lactobacillus, Bifidobacterium and Saccharomyces) significantly enhanced the efficacy of conventional treatments. These findings are further corroborated by numerous human clinical trials, where adding probiotics to triple or quadruple antibiotic regimens led to higher eradication rates than antibiotics alone. Such clinical successes have been reported with C. butyricum (Ref. Reference Mukai, Handa, Suyama, Majima and Naito92) and L. reuteri (Ref. Reference Ivashkin, Maev, Poluektova, Sinitsa, Avalueva, Mnatsakanyan, Simanenkov, Karpeeva, Kopylova, Kuprina, Kucheryavyy, Lapina, Solovyeva, Soom, Cheremushkina, Maevskaya and Maslennikov111).
Formation of complex biofilm is a way by which some strains of H. pylori have become resistant to antibiotic therapies. In an in vitro study, Jin et al. (Ref. Reference Jin and Yang68) demonstrated that adding a cell-free supernatant (CFS) of Lactobacillus salivarius to amoxicillin and clarithromycin can increase the destruction of biofilm structure in a dose-dependent manner (Ref. Reference Jin and Yang69). Ji et al. (Ref. Reference Ji and Yang65) found that CFS of Lactobacillus plantarum increases biofilm degradation when added to levofloxacin but decreases it in combination with clarithromycin, which suggests a strain-specific interaction with different antibiotics.
Side effects are a common issue when using antibiotics and can lead to decreased compliance and treatment failure or cause microbiome disruption-related complications. Probiotics can be used to alleviate these problems. Clinical trials have also shown the benefits of probiotics in managing treatment-related complications; for instance, supplementation with Bacillus clausii (Ref. Reference Plomer, Perez and Greifenberg105) or S. boulardii (Ref. Reference Zhao, Yang, Aruna, Xiao, Song, Huang, Li, Kou, Huang, Ji, Xiong, Peng, Xu and Cheng104) resulted in a significant reduction of the severity and duration of diarrhoea during eradication. Bifidobacterium animalis (Ref. Reference Han, Li, Nan, Zhou, Li and Li106) successfully reduced the incidence of nausea. Abdominal distention and dyspepsia were shown to be decreased by supplementation with L. plantarum (Ref. Reference Han, Liu, Zhang, Zeng, Liu, Gan, Zheng, Liu, Fang and Chen99), L. reuteri (Ref. Reference Moreno Márquez, Fernández Álvarez, Valdés Delgado, Castro Laria, Argüelles Arias, Caunedo Álvarez and Gómez Rodríguez98), and Limosilactobacillus fermentum (Ref. Reference Zhao, Niu, Zhang, Zhao, Zhang, He, Zhang, Mao, Wang, Zhao and Wang119) along with antibiotic regimens. A significant side effect of antibiotics is the disruption of gut microbiota, as beneficial bacteria can be killed in the crossfire between antibiotics and pathogens. Probiotics can restore balance in the microbiota composition. This effect was seen in supplementation with strains found in fermented milk products, Lactobacillus paracasei and L. rhamnosus (Refs Reference Niu, Li, Qian, Liang, Shi and Bu100, Reference FitzGerald, Patel, Eckenberger, Guillemard, Veiga, Schäfer, Walter, Claesson and Derrien107) or a combination of Bacillus subtilis and Enterococcus faecium (Ref. Reference Wu, Wang, Sun, Peng, Lu, Yan, Huang and Yang91). Probiotics use prebiotics as a source of nutrients to confer their health benefits. Abdulkhako et al. (Ref. Reference Abdulkhakov, Markelova, Safina, Siniagina, Khusnutdinova, Abdulkhakov and Grigoryeva118) demonstrated that supplementation with postbiotic butyric acid and prebiotic inulin has a similar effect. Wang et al. (Ref. Reference Wang, Wei, Liu, Wang, Zhang and du116) showed that fucoidan, when used as a prebiotic or in combination with bacteria as a symbiotic, can mitigate dysbiosis in the gut microbiome and prevent the increase of pathogens like Escherichia or Shigella.
H. pylori infection plays a major role in gastric carcinogenesis through inflammation and oxidative stress, and probiotics may counter these mechanisms, with several studies showing inhibition of H. pylori virulence factors (Ref. Reference Teanpaisan and Pahumunto109). in vivo experiments in mice have shown that the Lactobacillus spp. specifically can reduce inflammation and oxidative stress and improve gastric mucosal integrity (Refs Reference Dong74, Reference Yu, Cao, Peng, Wu, Li, Wu, Qing, Zhang, Wang, Huang and Zhao80). Song et al. (Ref. Reference Song, Zhou, Liu, Ge and Li60) utilized human gastric epithelial (GES-1) cells to demonstrate that L. acidophilus and L. bulgaricus effectively suppress IL-8 production by modulating the TLR4/NF-κB pathway. This effect was also observed by Chen et al. (Ref. Reference Chen, Tang, Li, Deng, Yu, Yang, Liu, Cheng, Huang, Guo, Shan, Zhou, Zeng, Bai and Fan81) when using Weizmannia coagulans. Gebremariam et al. (Ref. Reference Gebremariam63) identified a novel anti-inflammatory mechanism whereby Lactobacillus gasseri suppresses the expression of ADAM17, a disintegrin and metalloproteinase 17 also known as TNF-α converting enzyme, resulting in reduced secretion of the pro-inflammatory cytokines TNF and IL-6.
Maleki-Kakelar et al. (Ref. Reference Maleki-Kakelar, Dehghani, Barzegari, Barar, Shirmohamadi, Sadeghi and Omidi66) demonstrated L. plantarum’s anti-cancer effects in gastric cancer cells via PTEN/Bax upregulation and AKT downregulation. Kiani et al. (Ref. Reference Kiani, Mojgani, Kobarfard, Saffarian, Ayatollahi and Khoramjouy114) showed L. rhamnosus and L. reuteri improved histopathology in H. pylori-challenged mice, while Yang et al. (Ref. Reference Yang, Wu, Cheng, Chen, Tseng, Chang and Sheu82) found Bifidobacterium lactis inhibited H. pylori-induced COX-2 signalling and regressed pre-cancerous metaplasia in a trial, supporting probiotics for gastric cancer prevention.
Taken together, current evidence supports a meaningful role for probiotics in the management of H. pylori infection, primarily as adjuncts rather than standalone eradication agents. Although probiotic monotherapy typically yields only moderate eradication rates, reflecting the inherent biological limitations of probiotics in directly eliminating H. pylori compared to antibiotics, their primary clinical significance lies in their ability to enhance antibiotic efficacy. This property may permit the use of lower antibiotic doses or simplified treatment regimens, thereby reducing the risk of resistance development.
H. pylori vaccine development: current candidates, immunological targets and clinical hurdles
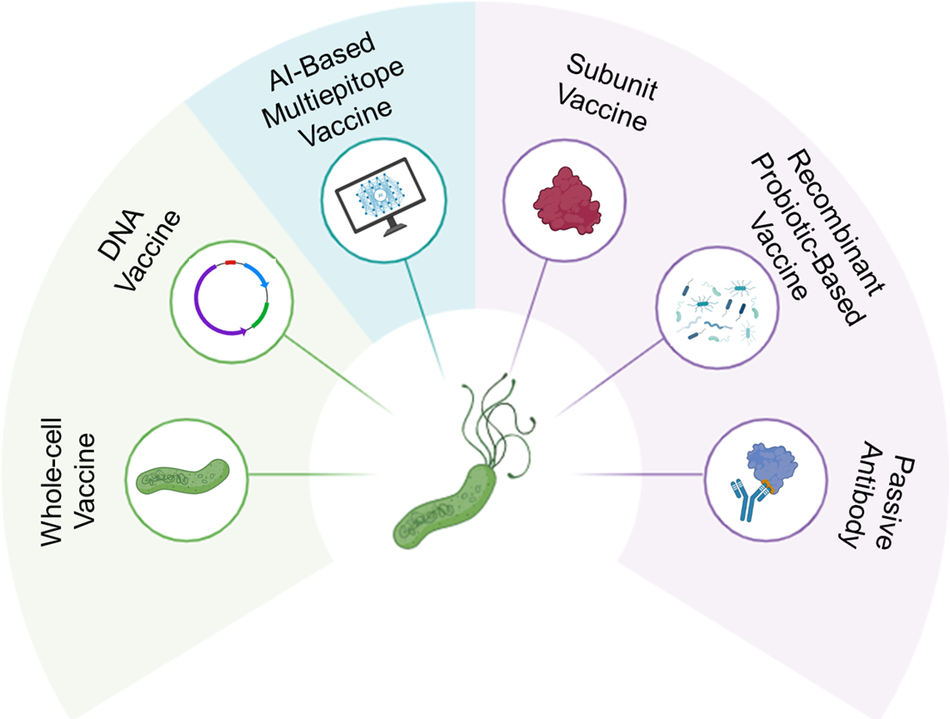
The quest for an effective H. pylori vaccine centre on bacterial components crucial for colonization and pathogenicity (Figure 3). Key antigens such as urease (Ref. Reference Keikha, Eslami, Yousefi, Ghasemian and Karbalaei120), CagA, VacA (Refs Reference Saadat, Higashi, Obuse, Umeda, Murata-Kamiya, Saito, Lu, Ohnishi, Azuma, Suzuki, Ohno and Hatakeyama121, Reference Cover and Blanke122), adhesins (BabA, SabA and HpaA) (Ref. Reference Xu, Soyfoo, Wu and Xu123) and NapA form the basis of current immunological targeting strategies (Ref. Reference Amedei, Cappon, Codolo, Cabrelle, Polenghi, Benagiano, Tasca, Azzurri, D’Elios, del Prete and de Bernard124). Building on these insights, several antibody-based approaches, including live attenuated, subunit, recombinant probiotic and passive antibody vaccines, are being explored to neutralize major virulence factors.
Overview of Helicobacter pylori vaccine and immunotherapy strategies. This schematic illustrates six main approaches. Each targets key virulence factors (e.g., urease, CagA, VacA and adhesins) and represents the evolution from traditional to next-generation, bioinformatically designed and antibody-based strategies for safer and more effective protection.

Live attenuated vaccines
Live attenuated vaccines use non-pathogenic or weakened H. pylori strains to elicit strong immune responses but are unsuitable for immunocompromised individuals (Ref. Reference Gebre, Brito, Tostanoski, Edwards, Carfi and Barouch125). In a preclinical study, Eaton et al. (Ref. Reference Eaton and Krakowka126) observed the production of H. pylori-specific IgG, IgM and IgA antibodies in immunized gnotobiotic piglets following administration of either formalin-killed H. pylori or an oral live attenuated vaccine. However, this immune response did not prevent bacterial infection. Similarly, at the preclinical level, Lee et al. (Ref. Reference Lee and Chen127) used two different adjuvants, non-toxic subunit B (CTB) with or without cholera toxin (CT), together with the whole-cell sonicate of Helicobacter felis. They found that even CTB alone could provoke an immune response and offer protection when challenged with H. felis in mice. In a clinical trial involving infected and uninfected adults, Kotloff et al. (Ref. Reference Kotloff, Sztein, Wasserman, Losonsky, DiLorenzo and Walker128) used a formalin-inactivated, oral H. pylori whole-cell vaccine with or without a mutant Escherichia coli heat-labile toxin (LTR192G) as an adjuvant. Although it did not eradicate H. pylori, it elicited an immune response in uninfected subjects. Summerton et al. (Ref. Reference Summerton, Welch, Bondoc, Yang, Pleune, Ramachandran, Harris, Bland, Jackson, Park, Clements and Nabors129) developed an oral H. pylori vaccine using a formalin-inactivated whole-cell antigen with a mutated E. coli heat-labile toxin adjuvant (dmLT). In mice, dmLT maintained strong immunogenicity while reducing gastric urease activity and bacterial colonization and, in lyophilized form, improved stability and reduced cold-chain dependence. However, early studies showed that, despite robust immune responses, these vaccines did not reliably prevent or eradicate infection, underscoring the need for safer and more effective mucosal adjuvants such as dmLT to unlock their prophylactic potential.
Subunit vaccines
Subunit vaccines use non-living pathogen components (Ref. Reference Hansson, Nygren and Ståhl130), such as inactivated toxins, surface polysaccharides or recombinant proteins, as immunogens (Ref. Reference Moyle and Toth131). They have a more favourable safety profile than live-attenuated vaccines but generally require adjuvants to elicit sufficiently strong immune responses (Ref. Reference D’Amico, Fontana, Cheng and Santos132). Le Guo et al. (Ref. Reference Guo133) describe a multivalent epitope-based vaccine (CWAE) against H. pylori infection in Mongolian gerbils, demonstrating a therapeutic effect through oral immunization. The study uses epitopes from NAP, urease, HSP60 and HpaA. This resulted in higher levels of helper T-cell response, IgG and secretory IgA antibodies against H. pylori, leading to a significant decrease in colony counts in the stomach. Jing Yang et al. (Ref. Reference Yang, Dai, Pan, Wang, Li, Zhu, Li, Shi and Wang134) report protection against infection in BALB/c mice when challenged with H. pylori after oral administration of a multi-epitope vaccine combining CTB-UreI-UreB. Le Guo et al. (Ref. Reference Guo, Yin, Liu, Lv, Li, Duan, Chu, Xi and Xing135) also discuss the immunological features and efficacy of CTB-UE, a multi-epitope vaccine, against H. pylori in the BALB/c mouse model. CTB-UE was able to induce production of specific antibodies against H.pylori virulence factors UreA and UreB at high levels and effectively decrease urease enzymatic activity, leading to a significant decrease in colonization. Hamzehloo et al. (Ref. Reference Hamzehloo, Mosayebi, Khansarinejad, Zolfaghari and Abtahi136) designed a recombinant multi-epitope protein by fusing the immunodominant regions of H. pylori UreB and FlaA antigens. The purified rFlaA-UreB protein, expressed in E. coli, showed strong antigenicity when tested with sera from infected patients, suggesting potential as both a vaccine and diagnostic candidate. This laboratory-based study highlights how combining key antigenic regions can enhance immune recognition and safety, particularly when formulated with mucosal adjuvants such as CTB in future preclinical vaccine models.
A recent study engineered H. pylori outer membrane vesicles (OMVs) as a delivery platform for the UreB, VacA and CagA antigens. This OMV-based recombinant vaccine elicited robust antigen-specific humoral and mucosal responses, induced a mixed Th1/Th2/Th17 profile with predominance of Th17, and significantly protected mice against H. pylori challenge (Ref. Reference Liu, Li, Ma, Lei, Ma, da, Zhou, Tao, Ren, Zeng, Xie, Lin, Jin, Wan, Zhang, Lai, Guo, Li, Shang, Shen, Tao, Gong and Zhang137).
DNA vaccines’ mechanism of action involves the delivery of a plasmid containing the genetic code for specific antigens, enabling endogenous antigen production by the host cells (Ref. Reference Yurina138). These vaccines are regarded as safer alternatives to live-attenuated vaccines (Ref. Reference Pagliari, Dema, Sanchez-Martinez, Montalvo Zurbia-Flores and Rollier139). While they elicit both humoral and cellular immune responses, they are generally characterized by relatively low immunogenicity (Ref. Reference Ingolotti, Kawalekar, Shedlock, Muthumani and Weiner140). Therefore, these vaccines need adjuvants to improve their effectiveness (Ref. Reference Li and Petrovsky141). Using a Flagellin A (FlaA) DNA vaccine, antibodies against FlaA were produced, reducing bacterial adhesion by 70% in murine models. It was also shown that T-cell polarization was towards T1 cells (Ref. Reference Ansari, Tahmasebi-Birgani and Bijanzadeh142). Urease B DNA vaccine was able to stimulate both innate and adaptive immune responses by increasing β-defensins and IL-10 and reducing bacterial load in the gastric mucosa in mice when administered via intra-nasal route (Ref. Reference Hatzifoti, Roussel, Harris, Wren, Morrow and Bajaj-Elliott143). In a phase III randomized placebo-controlled clinical trial conducted in China, uninfected children aged 6 to 15 years received an oral DNA vaccine consisting of the urease B subunit and the heat-labile enterotoxin B subunit of E. coli (Ref. Reference Zeng, Mao, Li, Tong, Wang, Zhang, Guo, Zhao, Li, Wu, Lu, Tan, Liang, Wu, Li, Luo, Zeng, Zhang, Zhang, Guo, Zhu and Zou144). Participants were visited at 4,8,12, 24 and 36 months and breath tests were performed to assess H. pylori infection and positive results were confirmed by ELISA tests on blood samples. A subset of participants donated blood and saliva samples at 6, 12, 24 and 36 months for immunogenicity and antibody persistence measurement. The results showed that vaccination led to IgA production, and in a 3-year follow-up, the vaccination had protective effects without significantly more side effects compared to placebo. A DNA vaccine comprising LeoA, a conserved antigen of H. pylori, and chitosan nanoparticle (CSNP) showed an increase in CD3+ T cells and IgG production, a decrease in bacterial load and gastric injury, and a shift from Th1 to Th2 population in BALB/C mice (Ref. Reference Chaleshtori, Rastegari, Nayeri and Doosti145). Despite their inherently low immunogenicity, preclinical studies reveal DNA vaccines’ ability to elicit Th1 and mucosal immune responses by targeting FlaA and UreB. More importantly, successful and long-lasting protection was demonstrated in the clinical trial. To further enhance the potency of these vaccines, delivery systems such as nanocarriers are being explored and should be the point of focus.
Recombinant probiotic-based vaccines
Recombinant probiotics are promising vaccine carriers that can express pathogenesis-related proteins and stimulate immune responses. Zhang et al. (Ref. Reference Zhang, Ni, Zhang, Luo, Wang, Zhou, Chen, Liu, Qu, Liu and Guo146) engineered a L. lactis (named LL-plSAM-WAE) to express a multiepitope vaccine containing urease, HpaA, HSP60 and NAP. They demonstrated that oral vaccinations of BALB/c mice with LL-plSAM-WAE increased IgG and sIgA against H. pylori and significantly reduced H. pylori colonization. Another study investigated the immunological effect of a recombinant L. lactis (named LL-plSAM-FAdE) that displays a multivalent epitope vaccine antigen (FAdE). The results of this study indicated that LL-plSAM-FAdE effectively activated gastrointestinal mucosal immunity against adhesion factors of H. pylori (including urease, CagL, HpaA and Lpp20) and prevented H. pylori infection in a mouse model. Additionally, this strain may eliminate H. pylori cells that are present in the gastrointestinal tract (Ref. Reference Zhang, Shi, Zhang, Wang, Liu, Li, Wang, Liu and Guo147).
AI-based vaccines
In recent years, integrating bioinformatic methods with immunological insights has helped identify discrete antigens among H. pylori antigens and optimize their selection for vaccine design. In this process, we can expect to achieve improved prophylactic and therapeutic outcomes. Cui et al. (Ref. Reference Cui, Ji, Guan, Su and du148) used online prediction tools ABCpred and IEDB for B-cell and T-cell epitope prediction, respectively, and designed a multiepitope vaccine using highest scoring epitopes in antigenicity and allergenicity from nine candidate H. pylori proteins. These proteins included UreB, SabA, BabA, VacA, CagA, GGT, HtrA, NAP and CagL. In silico immune simulation of the final multiepitope vaccine predicted high activities of Linea B cell, helper T cell, cytotoxic T cell, macrophages, and dendritic cells as well as increased levels of INF-ɣ, IL-2 and IL-10, which indicate a strong immune response to vaccination. Jebali et al. (Ref. Reference Jebali, Esmaeilzadeh, Esmaeilzadeh and Shabani149) took a step further with their vaccine, which is made of five epitopes from five H. pylori target proteins, urease, CagA, HopE, SabA and BabA. After molecular simulation and docking assessments, they constructed the final protein using the solid phase method and enclosed it in lipid nanoparticles. Product properties were affirmed by nanodrop at 280 nm, dynamic light scattering and atomic force microscope. The construct was shown to be non-toxic and non-allergenic while having a 99.05% binding rate with different MHC alleles. Still, safety and immunogenicity of these vaccines are yet to be confirmed, and cellular, animal and human phase assessments are required. This is the barrier separating computational predictions from real-world applications.
Whole-cell vaccines’ inability to provide protection in early efforts shifted the focus towards safer platforms like subunit and DNA vaccines. To address their naturally lower immunogenicity, researchers are exploring the use of bioinformatics to rationally design multi-component vaccines and developing advanced delivery systems, such as nanoparticles, along with engineering sophisticated adjuvants. This modern approach shows great promise as the future of vaccine development for effective immunization against H. pylori.
Passive antibody therapy
Administration of specific antibodies to neutralize pathogenesis-related factors of H. pylori is a promising therapeutic strategy for controlling the infection (Ref. Reference Ardekani, Gargari, Rasooli, Bazl, Mohammadi, Ebrahimizadeh, Bakherad and Zare150). Zhong et al. (Ref. Reference Zhong, Liu, Wang, Hou, Guo and Luo151) recently designed a urease-targeting humanized single-domain antibody (I107W) through in silico studies and developed an engineered E. coli Nissle 1917 (EcN) to express the I107W antibody. This engineered strain significantly alleviated gastric H. pylori colonization and suppressed the expression of pro-inflammatory markers. Moreover, the results demonstrated that I107W enhanced mucosal repair and maintained microbial composition in the H. pylori mice model.
Antibody conjugation can be considered an effective approach to specifically target H. pylori. Wang et al. (Ref. Reference Wang, Song, Gao, Liu, Guan, Mei, Ma and Cui152) developed an antibody-conjugated liposome loaded with indocyanine green (ICG) (HpAb-LiP-ICG). This platform provided a photoacoustic (PA) imaging-guided sonodynamic therapy (SDT) for recognizing and targeting H. pylori infection. PA imaging results revealed the capacity of HpAb-LiP-ICG for H. pylori detection in the stomach. In addition, pathological analysis showed that HpAb-LiP-ICG-mediated SDT specifically targeted H. pylori and had no cytotoxic effect on normal tissues. These findings indicate that this platform provides a promising tool to detect and treat H. pylori in the future.
A source for antibody therapy is egg yolk antibody, which contains immunoglobulin Y (IgY). This antibody source attracted much attention as an antibody therapy for H. pylori infection (Ref. Reference Cheng, Li, Luo, Chi, Zhao, Lin and Xu153). Egg yolk antibody is an immunoglobulin produced by activating avian B lymphocytes against specific pathogens that transfers from serum to yolk, and can be used as passive immunization for the corresponding pathogen (Ref. Reference Pereira, van Tilburg, Florean and Guedes154). Previous studies have demonstrated that IgY can be an effective approach to control H. pylori infection. Attallah et al. (Ref. Reference Attallah, Abbas, Ismail, Abdel-Raouf and el-Dosoky155) developed a yolk antibody (IgY-Hp58) that prevented H. pylori infection and reduced gastritis in the H. pylori mice model. Guo et al. (Ref. Reference Guo, Wu, Guan, Xie, Yang, Li and Zhang156) developed a multivalent anti-H. pylori IgY targeting UreB, BabA2 and FlaA, which demonstrated strong antigen binding, in vitro growth inhibition of H. pylori, and gastric stability when combined with skim milk. In the same study, a 2-week clinical trial in 94 infected patients showed 56% reduction in 13C-urea breath test values, 30.6% eradication rate and 87.3% clinical symptom improvement with no adverse effects.
Taken together, although multiple vaccine platforms have demonstrated robust immunogenicity and reduced colonization in animal models, their clinical translation has remained limited. A recurring challenge is that strong systemic antibody responses do not necessarily confer durable mucosal protection within the highly specialized gastric niche, where sustained secretory IgA responses and appropriately polarized Th1/Th17 immunity are required for bacterial clearance (Refs Reference Sutton and Boag157–Reference Dixon, Hossain, Patel and Algood159). Moreover, H. pylori employs sophisticated immune-evasion mechanisms, including induction of regulatory T cells and persistence within the mucus layer, which can attenuate vaccine-induced protection despite measurable immunogenicity (Ref. Reference Bakiera, Solarz, Kowalczyk, Cichoż-Lach and Korona-Głowniak160). Safety constraints further complicate translation, as potent mucosal adjuvants that enhance protection in preclinical models, such as cholera toxin derivatives, are often unsuitable for widespread human use due to reactogenicity concerns (Ref. Reference Fukuyama, Okada, Yamaguchi, Kiyono, Mori and Yuki161). In addition, antigenic heterogeneity among global H. pylori strains and inter-individual variability in host immune responses reduce the generalizability of promising preclinical findings (Ref. Reference Wylie, Gilbreath, Melton-Celsa and Merrell162). Collectively, these limitations illustrate that immunogenic potency in controlled experimental systems may not guarantee clinical efficacy, emphasizing the need for improved mucosal delivery systems, safer adjuvants and rigorous phase-based validation before vaccine candidates can achieve practical applicability.
Translational perspective and clinical readiness
Non-antibiotic H. pylori therapies differ widely in clinical maturity, from experimental concepts to regimens with human data. This section ranks these options by clinical evidence and feasibility, underscoring the gap between in vitro promise and real-world effectiveness (Table 4).
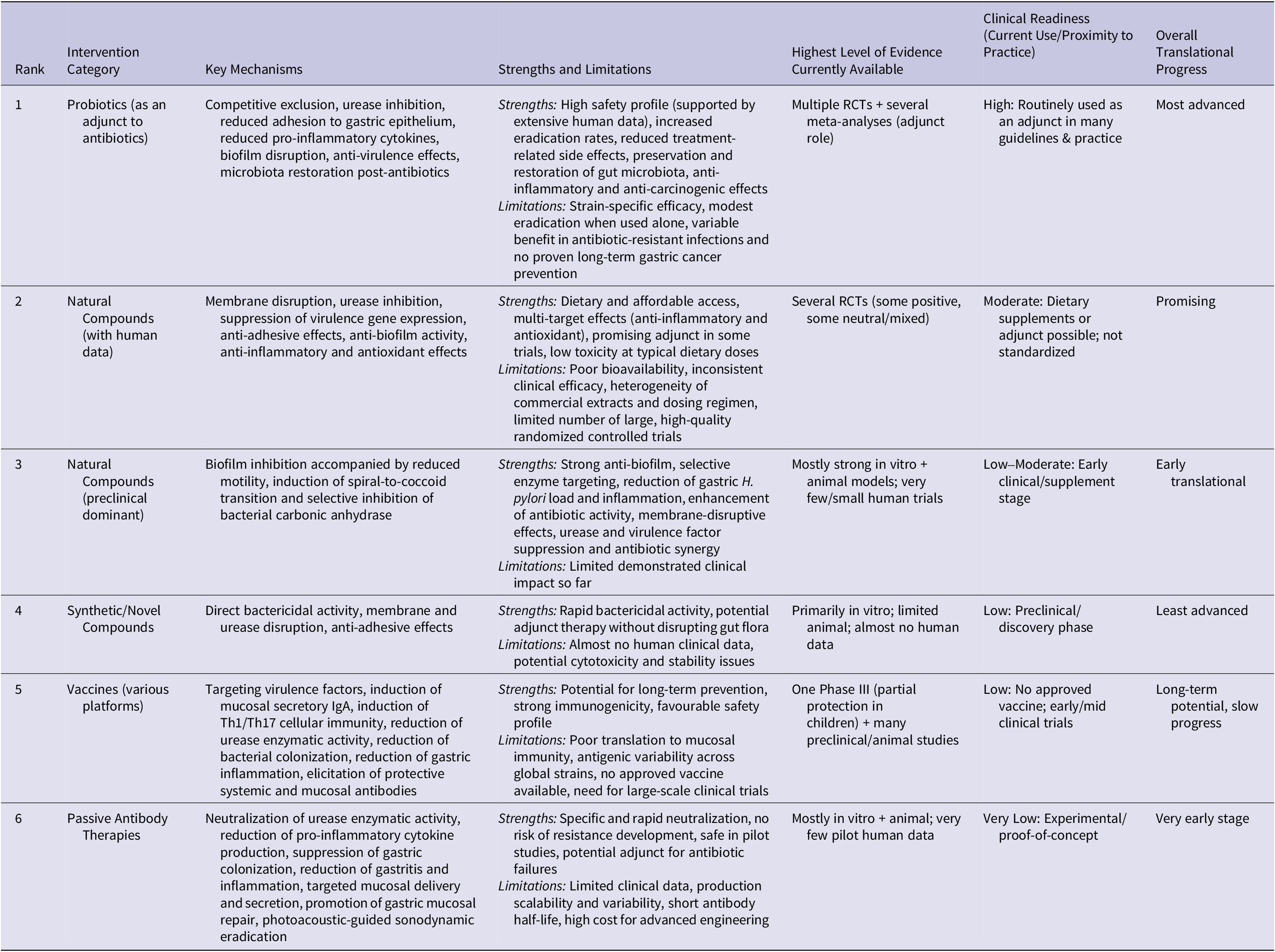
Summary ranking of non-antibiotic interventions against Helicobacter pylori by current level of evidence and clinical readiness

Comparative analysis reveals that antimicrobial peptides and synthetic inhibitors demonstrate strong direct antimicrobial activity, specificity, broad-spectrum effects and immune-modulatory properties in preclinical models, yet their clinical translation is hindered by pharmacokinetic challenges, instability, cytotoxicity and regulatory barriers (Ref. Reference Zheng, Tu, Li, Qu, Li, Peng, Li and Shao163). In contrast, probiotics show moderate direct anti-H. pylori activity and, when used adjunctively, improve eradication rates and reduce side effects in randomized clinical trials. Meta-analyses of standardized randomized controlled trials (RCTs) demonstrate that specific strains enhance standard therapy efficacy and decrease gastrointestinal adverse events, with good reproducibility and safety supported by extensive human use and strain characterization. However, effects remain strain-specific and context-dependent, highlighting the need for standardized strain selection and dosing to optimize clinical benefit (Ref. Reference Yang, Zhou, Han, He, Zhang, Wu and Chen164). Natural compounds and phytochemicals show in vitro anti-virulence, anti-inflammatory and biofilm-disrupting effects with generally good safety, but inconsistent human potency and reproducibility result from variable bioavailability, extraction differences and low achievable gastric concentrations, limiting current translational potential (Ref. Reference Dumra and Ray165). H. pylori vaccines promise long-term specific prevention, but none are in widespread clinical use. Late-stage subunit candidates face major challenges, including antigenic variation, immune evasion, difficulty inducing strong mucosal immunity and the high cost plus regulatory complexity of large multicentre trials (Ref. Reference Liu, Li, Huang and Liu166).
Conclusion
Rising antibiotic resistance in H. pylori demands effective non-antibiotic alternatives. Promising options include natural/synthetic compounds, adjunctive probiotics, vaccines and passive antibody therapies.
Significant biological and translational barriers remain. The immune-tolerant gastric mucosa, dominated by regulatory T cells and suppressed effector responses, impedes protective immunity. Host genetic heterogeneity and variable Th1/Th17 polarization cause inconsistent efficacy across individuals. Historical challenges in mucosal delivery, including mucus penetration, sustained secretory IgA/Th17 induction and adjuvant reactogenicity, continue to limit progress despite promising preclinical results. Taken together, integrated non-antibiotic approaches, especially strain-specific probiotics, should complement standard therapies, while optimized bioactive compounds and future vaccine or antibody platforms represent medium- and long-term priorities towards effective and durable H. pylori control.
Abbreviations
- ADAM17
-
A Disintegrin and Metalloproteinase 17
- AKT
-
Protein Kinase B
- AlpA/B
-
Adhesin-like Proteins A/B
- AST
-
Antibiotic Susceptibility Testing
- ATCC
-
American Type Culture Collection
- BabA
-
Blood Group Antigen-Binding Adhesin
- BabB
-
Blood Group Antigen-Binding Adhesin B
- Bax
-
Bcl-2-associated X protein
- BCA
-
Bicinchoninic Acid
- BHI
-
Brain Heart Infusion
- BQT
-
Bismuth Quadruple immune evasion
- CagA
-
Cytotoxin-Associated Gene A
- CagL
-
Cytotoxin-Associated Gene L
- CFS
-
Cell-Free Supernatant
- CFU
-
Colony-Forming Unit
- CNCM
-
Collection Nationale de Cultures de Microorganismes
- COX-2
-
Cyclooxygenase-2
- CPC
-
Cetylpyridinium Chloride
- CSNP
-
Chitosan Nanoparticle
- CT
-
Cholera Toxin
- CTB
-
Cholera Toxin B-subunit
- CWAE
-
Multivalent Epitope-Based Vaccine (NAP, urease, HSP60, HpaA)
- DHFR
-
Dihydrofolate Reductase
- dmLT
-
Double-Mutant Heat-Labile Toxin
- DSMZ
-
Deutsche Sammlung von Mikroorganismen und Zellkulturen
- EGCG
-
Epigallocatechin-3-Gallate
- ELISA
-
Enzyme-Linked Immunosorbent Assay
- EVOO
-
Extra Virgin Olive Oil
- FAdE
-
Multivalent Epitope Vaccine Antigen
- FICI
-
Fractional Inhibitory Concentration Index
- FlaA
-
Flagellin A
- FlaB
-
Flagellin B
- FOXP2
-
Forkhead Box P2
- FtsZ
-
Filamenting Temperature-Sensitive Mutant Z
- GGT
-
Gamma-Glutamyl Transpeptidase
- H. pylori
-
Helicobacter pylori
- HopE
-
Helicobacter Outer Protein E
- HpaA
-
Helicobacter pylori Adhesin A
- HSP60
-
Heat Shock Protein 60
- HtrA
-
High-Temperature Requirement A
- IC50
-
Half-Maximal Inhibitory Concentration
- ICG
-
Indocyanine Green
- IgY
-
Immunoglobulin Y
- IL-2
-
Interleukin-2
- IL-6
-
Interleukin-6
- IL-8
-
Interleukin-8
- IL-10
-
Interleukin-10
- INF-γ
-
Interferon-gamma
- Ki
-
Inhibition Constant
- Le(x)
-
Lewis x Antigen
- Lpp20
-
Lipoprotein 20
- LTR192G
-
Mutant Escherichia coli Heat-Labile Toxin
- MALT
-
Mucosa-Associated Lymphoid Tissue
- MBC
-
Minimum Bactericidal Concentration
- MIC
-
Minimum Inhibitory Concentration
- MYR
-
Myricetin
- NapA
-
Neutrophil-Activating Protein A
- NF-κB
-
Nuclear Factor kappa-light-chain-enhancer of activated B cells
- OD
-
Optical Density
- OMVs
-
Outer Membrane Vesicles
- PA
-
Photoacoustic
- PBPs
-
Penicillin-Binding Proteins
- PPI
-
Proton Pump Inhibitor
- PTCC
-
Persian Type Culture Collection, randomized controlled trials (RCTs)
- PTEN
-
Phosphatase and Tensin Homolog
- RSV
-
Resveratrol
- RT-qPCR
-
Real-Time Quantitative Polymerase Chain Reaction
- SabA
-
Sialic Acid-Binding Adhesin
- SCFAs
-
Short-Chain Fatty Acids
- SDH
-
Shikimate Dehydrogenase
- SDT
-
Sonodynamic Therapy
- SEM
-
Scanning Electron Microscopy
- TLR4
-
Toll-Like Receptor 4
- TMEWP
-
Chios Mastic Gum
- TNF
-
Tumour Necrosis Factor
- UreA
-
Urease Subunit A
- UreB
-
Urease Subunit B
- UreE
-
Urease Accessory Protein E
- UreF
-
Urease Accessory Protein F
- UreI
-
Urease Inner Membrane Protein
- VacA
-
Vacuolating Cytotoxin A
Acknowledgements
We used ChatGPT-5 to assist with paraphrasing and improving the clarity of the English language in this manuscript; no scientific content was generated by the tool.
Author contribution
A.F.T., A.N., S.N.M. and M.R. contributed to the writing of the manuscript and table design. S.N.M. was responsible for figure design. E.N.M. edited the manuscript. Z.S. conceptualized and designed the study, contributed to manuscript writing and edited the manuscript. H.R. contributed to manuscript writing, table design and edited the manuscript.
Funding statement
The authors have not disclosed any funding.
Competing interests
The authors declare that they have no competing interests.

 Open access
Open access