Introduction
Aedes albopictus (Diptera: Culicidae), commonly referred to as the ‘Asian tiger mosquito’, is characterised by its robust invasive capabilities and extensive distribution (Bonizzoni et al., Reference Bonizzoni, Gasperi, Chen and James2013; Su et al., Reference Su, Guo, Deng, Xu, Zhou, Zhou, Li, Zhong, Kong, Wang, Liu, Wu, Yan and Chen2019). It serves as a vector for various mosquito-borne diseases such as dengue fever and Zika virus, which pose significant threats to human health, resulting in substantial economic losses worldwide and presenting formidable challenges to global public health governance. Given that vaccines against the diseases transmitted by Ae. albopictus are still under development (Khetarpal and Khanna, Reference Khetarpal and Khanna2016), current preventive and control measures primarily involve environmental management, chemical control, and biological control. Chemical control predominantly relies on the use of insecticides to eliminate mosquitoes, with deltamethrin being a commonly used pyrethroid insecticide (Dusfour et al., Reference Dusfour, Vontas, David, Weetman, Fonseca, Corbel, Raghavendra, Coulibaly, Martins, Kasai and Chandre2019; Piedra et al., Reference Piedra, Rodriguez, Lopez, Ruiz, Martinez, Garcia, Rey and Bisset2023). Over the past few decades, deltamethrin insecticides have played a crucial role in combating mosquito-borne diseases like dengue fever (Smith et al., Reference Smith, Kasai and Scott2016; Bo et al., Reference Bo, Yu, Xi and Tang2018). However, excessive and prolonged usage of deltamethrin has resulted in its residues persisting in the environment and impacting the survival of Ae. albopictus (Hou et al., Reference Hou, Liu, Wang, Wu, Li and Gong2020; Sraku et al., Reference Sraku, Owusu-Asenso, Abdulai, Addo, Mensah, Owusu, Akuamoah-Boateng, Mohammed Sabtiu, Boadu, Danquah, Kwarteng, Appiah-Kwarteng, Forson, Attah and Afrane2025).
Deltamethrin belongs to the type II pyrethroid insecticide, which is classified as the most potent among synthetic pyrethroid insecticides. It possesses several advantages, including high efficacy, quick degradation in water, broad-spectrum activity, and cost-effectiveness. Consequently, it has gained extensive utilisation in mosquito control and agricultural production (Lu et al., Reference Lu, Sun, Ares, Anadón, Martínez, Martínez-Larrañaga, Yuan, Wang and Martínez2019; Qi et al., Reference Qi, Lin, Gao, Wu, Zhang, Wei, Xiao, Wang, Peng, Clark, Park and Sun2022). However, prolonged usage of deltamethrin insecticide has led to varying degrees of resistance in Ae. albopictus, posing new challenges for vector control. Additionally, the overuse of pesticides has resulted in a considerable accumulation of their residues in water bodies (Wolfram et al., Reference Wolfram, Stehle, Bub, Petschick and Schulz2018; Wu et al., Reference Wu, Gao, Xie, Wu, Song, Yuan, Wu and Ou2022; Gu et al., Reference Gu, Chen, Jiang, Liao, Long and Xie2024; Shi et al., Reference Shi, Zhang, Chen, Mao, Feng, Yang, Zhao, Wu and Chen2024). Deltamethrin can enter water bodies through surface runoff. It has low water solubility, dissipates rapidly from the aqueous phase, and is typically present in stream water for only a few hours (Liess et al., Reference Liess, Schulz, Liess, Rother and Kreuzig1999). However, deltamethrin can adsorb to surfaces such as river sediments, microbial biofilm, food, and habitats, prolonging its environmental persistence (Gan et al., Reference Gan, Lee, Liu, Haver and Kabashima2005; Rasmussen et al., Reference Rasmussen, Reiber, Holmstrup and Liess2017). A study carried out at a municipal wastewater treatment plant in Tabriz, a city in northwestern Iran, demonstrated that the concentration of deltamethrin residues in water bodies can reach as high as 787 ng/L (Firouzsalari et al., Reference Firouzsalari, Shakerkhatibi, Pourakbar, Yadeghari, Safari and Sarbakhsh2019). The current reporting limits for deltamethrin and permethrin are higher than the minimum aquatic benchmark; therefore, the true number of benchmark exceedances could be higher than reported (Budd et al., Reference Budd, Wang, Ensminger and Phillips2020). A study by Betz-Koch et al. (Reference Betz-Koch, Grittner, Krauss, Listmann, Oehlmann and Oetken2024) using artificial indoor streams demonstrated that deltamethrin at a concentration of 64 ng/L caused lethal and adverse sublethal effects on aquatic communities. Rasmussen et al. (Reference Rasmussen, Reiber, Holmstrup and Liess2017) showed that in water bodies containing 1 ng/L deltamethrin, combined exposure through water and food significantly reduced the emergence of the caddisfly Anabolia nervosa by 60% and significantly postponed emergence timing. Furthermore, deltamethrin residues can be transmitted and bioaccumulated through the food chain across trophic levels (He et al., Reference He, Ni, Jin and Fu2020), ultimately entering the human body and posing a potential threat to human health (Marettova et al., Reference Marettova, Maretta and Legáth2017; Shi et al., Reference Shi, Zhang, Chen, Mao, Feng, Yang, Zhao, Wu and Chen2024). Consequently, there is an increasing focus on employing deltamethrin judiciously in both environmental protection and mosquito control.
The life cycle of Ae. albopictus, an insect that undergoes complete metamorphosis, comprises four stages: egg, larva, pupa, and adult. The first three stages occur in water while the adult mosquito resides on land. Due to their aquatic habitat and feeding behaviour involving algae, protozoa, and yeast consumption, Ae. albopictus larvae tend to accumulate deltamethrin residues from their surrounding water (Qu et al., Reference Qu, Shi, Chen, Liu, Yao, Li, Tan and Xie2024). Sublethal exposure to insecticides is known to induce physiological stress and fitness costs in mosquito populations, which may have implications for population dynamics and vector control (Hou et al., Reference Hou, Liu, Wang, Wu, Li and Gong2020; Sraku et al., Reference Sraku, Owusu-Asenso, Abdulai, Addo, Mensah, Owusu, Akuamoah-Boateng, Mohammed Sabtiu, Boadu, Danquah, Kwarteng, Appiah-Kwarteng, Forson, Attah and Afrane2025). Based on previous laboratory studies (unpublished), a susceptible strain of Ae. albopictus developed moderate to high resistance after seven generations of deltamethrin selection, characterised by a significant increase in LC50 and the emergence of metabolic resistance (CYP6 upregulation by two- to threefold and increased GST activity) and potential cuticular resistance (Lac2 elevation). However, the specific effects of multigenerational exposure to environmentally relevant residual concentrations of deltamethrin on the key life history traits of Ae. albopictus – particularly growth, development, and fecundity – remain inadequately characterised. Therefore, in this study, we employed varying concentrations of deltamethrin solution to induce long-term poisoning in Ae. albopictus and simulate the residue levels found in diverse aquatic environments. Subsequently, we investigated the impact of deltamethrin on the growth, development, and fecundity of Ae. albopictus. We aim to elucidate how multigenerational exposure to deltamethrin influences the population dynamics of Ae. albopictus, thereby providing insights for rationalising its usage.
Material and methods
Mosquitoes
The laboratory colony of Ae. albopictus mosquitoes were sourced from Sun Yat-sen University (Guangzhou, Guangdong Province, China; approximately 23.13° N, 113.26° E) and reared in a fully automated ACG II artificial climate chamber (QiuShi, Zhejiang, China), with an ambient temperature of 27 ± 1 °C, relative humidity of 70–85%, and a photoperiod of 14L: 10D. The colony was maintained in our laboratory for more than 10 generations prior to the experiments to establish a stable, laboratory-adapted population. Cat food and yeast powder were ground into a mixed powder at a 2:1 ratio for feeding one to two instar larvae. The three to four instar larvae were fed cat chow powder, and post-eclosion adults were fed 10% sugar water. Mice were used as blood samples for female mosquitoes to induce egg-laying, following the method of Zhang et al. (Reference Zhang, Ding, Zhou, Tang, Chen, Chen, Wen and Wang2023).
Ae. albopictus deltamethrin stress treatment
Following the WHO standard larval bioassay method (WHO, 2003), this experiment investigated the chronic behavioural responses of Ae. albopictus larvae exposed to sublethal concentrations of deltamethrin. The active ingredient of the deltamethrin stock solution (Bayer, Germany; Lot number: XK13-003-00459) was 25 g/L. Stock solutions were serially diluted with ddH₂O to prepare a range of test concentrations of 0.0001, 0.0005, 0.001, 0.005, and 0.1 mg/L. For each concentration, 200 mL of the test solution was dispensed into a 250-mL plastic cup. A control group containing 200 mL of ddH₂O without insecticide was also included. Twenty-five fourth-instar larvae, reared under identical conditions, were then introduced into each cup for a 24-h exposure period. Larval mortality was assessed by recording the number of dead individuals in each cup. Larvae were considered dead if they were unresponsive to mechanical stimulation or were exhibiting a moribund state. Each concentration was tested in triplicate, with fourth-instar Ae. albopictus larvae reared in double-distilled water (ddH2O) serving as the control group. The toxicity equation is expressed as y = 3.656 + 1.383 lg x. The calculated 24-h LC50 value, determined through Probit regression analysis, is found to be 0.002275 mg/L, with a 95% confidence interval of 0.001476 < LC50 < 0.003457. Based on the LC50 value and the concentration of deltamethrin detected in the environment (Firouzsalari et al., Reference Firouzsalari, Shakerkhatibi, Pourakbar, Yadeghari, Safari and Sarbakhsh2019; Gu et al., Reference Gu, Chen, Jiang, Liao, Long and Xie2024; Shi et al., Reference Shi, Zhang, Chen, Mao, Feng, Yang, Zhao, Wu and Chen2024), concentrations of 0.0001, 0.0005, and 0.001 mg/L of deltamethrin were selected for long-term exposure. The stock solutions were diluted to achieve final concentrations of 0.0001, 0.0005, and 0.001 mg/L using ddH2O. Following hatching in clean water, first-instar larvae were collected using a syringe and filtering spoon, then placed into white ceramic pots (diameter: 25 cm) containing deltamethrin solution at the respective concentrations. Each pot contained more than 500 larvae. Adult mosquitoes were maintained in cages at an approximate ratio of 1 female:3 males for mating and blood-feeding. The aquatic environment containing larvae and pupae was refreshed daily with deltamethrin solution at the appropriate concentration. Eggs laid by adult mosquitoes were collected and incubated to produce subsequent generations for further deltamethrin treatments, continuing until the fourth generation of mosquitoes was achieved. There was approximately 1 week between each generation of mosquitoes. It should be noted that the LC₅₀ was determined using fourth-instar larvae, but the multigenerational exposure began at the first instar. First-instar larvae may be more sensitive to deltamethrin.
Detection of related gene expression
Mosquitoes were collected using a mosquito trap and killed by freezing at –80°C. Females were sorted on ice and placed into tubes (12 individuals per tube, 1 day post-eclosion). A total of 39 tubes were prepared. Each tube represented one biological replicate (n = 3 per group per generation for the treated groups; n = 3 for the control group). All samples were stored at −80°C. RNA was extracted using the Trizol method; the quality and concentration of RNA were assessed by agarose gel electrophoresis with an electrophoresis system (Bio-Rad, California, USA). cDNA was synthesised for real-time fluorescent quantitative PCR (Bio-Rad, California, USA). The qPCR mixture (10 µL) contained 5 µL of TB Green® Premix Ex Taq™, 3.2 µL of ddH2O, 0.4 µL each of upstream and downstream primers (Table 1) and 1 µL cDNA. The qPCR procedure was as follows: 10 min at 94 °C; 30 s at 94 °C, 30 s at 59 °C, 45 s at 72 °C for a total of 39 cycles, and then 10 min at 72 °C. The actin gene was used as an internal control, and the 2−△△CT method was used to calculate the relative expression of vitellogenin-A1 (Vg-A) and vitellogenin-C (Vg-C) genes in female Ae. albopictus treated with different concentrations and at different generations.
Primers for qRT-PCR

Table 1 Long description
The table lists three primer sets used for quantitative reverse transcription PCR, giving each primer name, its forward and reverse nucleotide sequences, and the corresponding NCBI accession. Vg-A uses forward TCTCGCTTTGGGTAAGTGTTCA and reverse GGCATCGTAGCCGTTCTTGTAG, linked to AY691316.1. Vg-C uses forward ACCAATGGACTGGCACCTACA and reverse CGGCGTATTCTGGCTGCTT, linked to AY691317.1. Actin uses forward GCTACGTCGCCCTGGACTT and reverse AGGAACGACGGCTGGAAGA, linked to DQ657949.1. The entries provide sequence identifiers for locating the referenced targets in NCBI, but the table does not include amplicon size, annealing conditions, or validation metrics, so performance comparisons cannot be inferred.
Measurement of physiological indicators
Fifty first-instar larvae were placed in white ceramic pots containing corresponding concentrations of deltamethrin solution. Larvae were fed daily with brewer’s yeast following the method of Delatte et al. (Reference Delatte, Gimonneau, Triboire and Fontenille2009) with modifications. Development and survival were monitored daily until all individuals either reached the fourth instar or died. Larval molting was confirmed by the presence of exuviae or distinct morphological changes. Developmental duration (days from first instar to fourth instar) and pupation rate (%) = (number of pupae/initial number of larvae) × 100 were recorded for each generation and each concentration. For pupal development, 20 pupae from each group were collected and placed into mosquito cages. Eclosion rate (%) = (number of emerged adults/total number of pupae) × 100 was calculated based on daily observation of adult emergence, following the method of Yee et al. (Reference Yee, Juliano and Vamosi2012). For fecundity assessment, upon emergence, females and males were kept together for 3–5 days to allow mating. Females were then offered a single blood meal. Three days post-blood meal, each female was transferred to an individual oviposition container with one male, lined with damp filter paper and provided with 10% sucrose solution. Eggs were collected and counted daily for up to 14 days post-blood meal, and the number of eggs laid per female was recorded. To determine the hatching rate, 50 mosquito eggs were collected and placed in a white ceramic bowl with ultra-pure tap water, and the number of hatchlings was recorded every day, and the observation ended when no more eggs hatched for 3 consecutive days.
Statistical analysis
Statistical analysis was conducted using SPSS Statistics 20.0, and graphing was performed using GraphPad Prism Version 9.5 software. To analyse the effects of multigenerational deltamethrin exposure on Ae. albopictus, a two-way ANOVA was performed. Two-way ANOVA models allow accounting for more than one known source of variation. Prior to performing two-way ANOVA, the assumptions of normality and homogeneity of variances were assessed. Normality was evaluated using the Shapiro–Wilk test, and homogeneity of variances was evaluated using Levene’s test. The results indicated that the assumptions were met for all dependent variables (p > 0.05), justifying the use of parametric two-way ANOVA. In this study, ‘concentration’ and ‘generation’ were treated as independent variables, with life-history traits and related gene expression serving as dependent variables. Simple main effects were analysed using simple Analysis of Variance (ANOVA) and Tukey’s Honestly Significant Difference tests.
Results
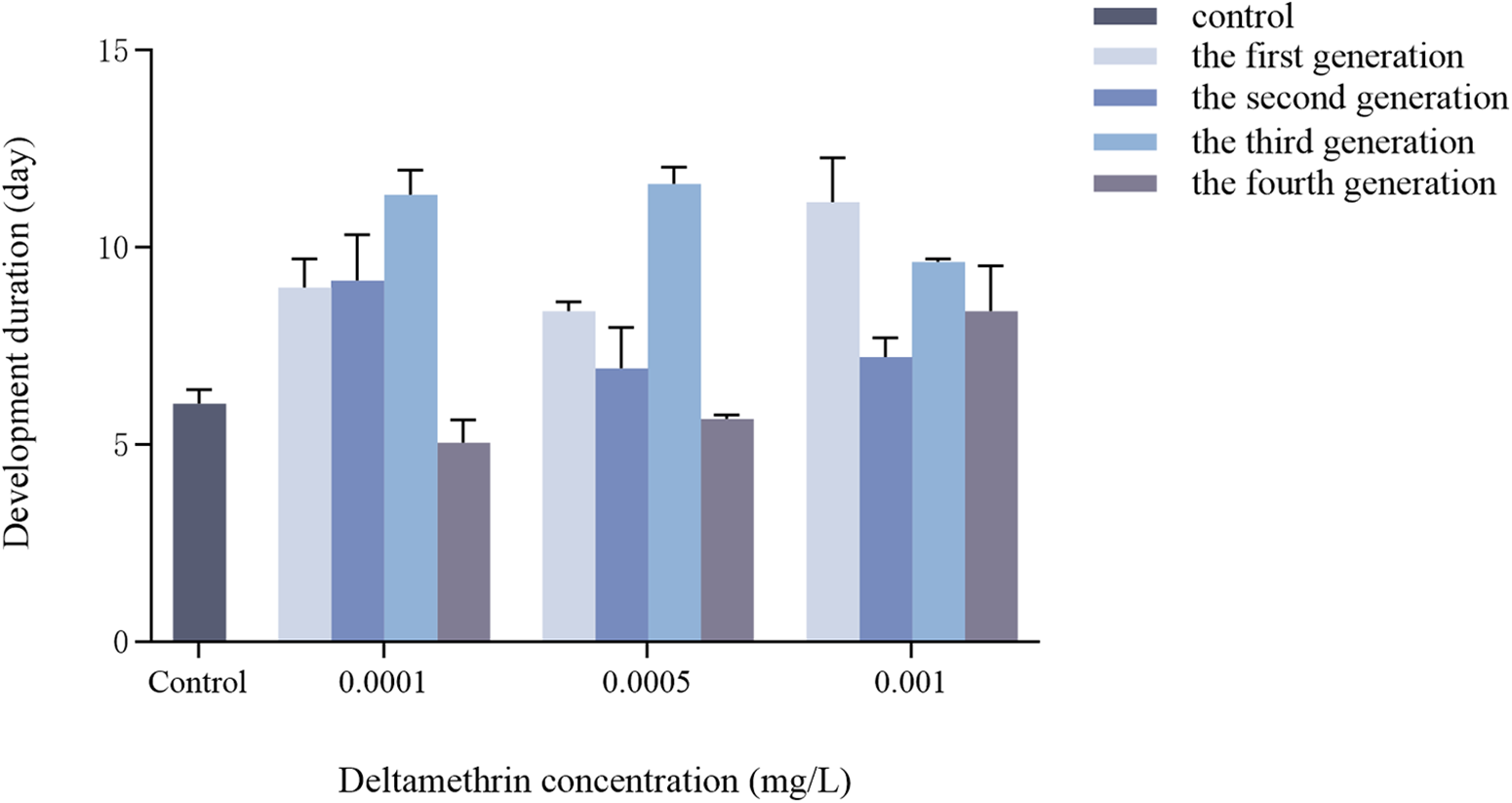
Effects of multigenerational deltamethrin exposure on the developmental duration of Ae. albopictus
There was a significant interaction between concentration and generation on the developmental duration from second-instar larvae to pupae of Ae. albopictus (F = 6.340, df = 9, p < 0.001) (fig. 1). Concentration had a highly significant effect on developmental duration (F = 18.441, df = 3, p < 0.001), and generation also had a highly significant effect on developmental duration (F = 21.948, df = 3, p < 0.001).
Transgenerational changes in the developmental duration of Ae. albopictus were investigated under different concentrations of deltamethrin stress. For each generation (F1–F4), larvae were continuously exposed to deltamethrin from the first instar until adult emergence. All measured individuals from each generation were directly exposed to the treatment. The control group was reared in parallel under identical conditions without insecticide exposure. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 1 Long description
A grouped bar graph with a legend listing: control, the first generation, the second generation, the third generation, the fourth generation. Horizontal axis label: Deltamethrin concentration left parenthesis milligram slash L right parenthesis. Categories shown: Control, 0.0001, 0.0005, 0.001. Vertical axis label: Developmental duration left parenthesis day right parenthesis. Scale shown from 0 to 15. Control category: one bar for control at about 5. 0.0001 category: control about 9; the first generation about 9; the second generation about 11; the third generation about 3. 0.0005 category: control about 8; the first generation about 6; the second generation about 11; the third generation about 4. 0.001 category: control about 11; the first generation about 7; the second generation about 9; the third generation about 8. Several bars include vertical error bars.
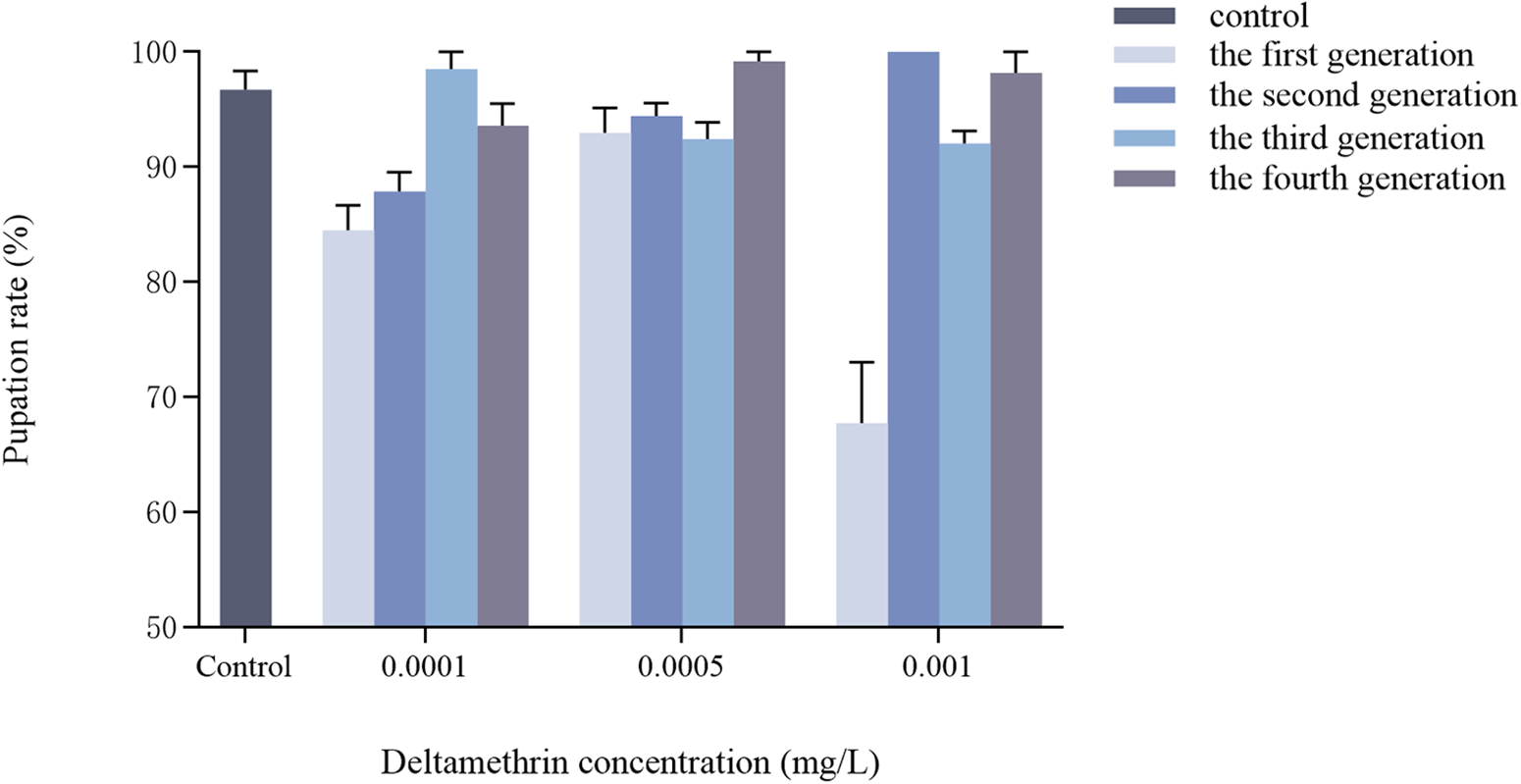
Effects of multigenerational deltamethrin exposure on the pupation rate of Ae. albopictus
No significant interaction was observed between concentration and generation on the pupation rate of Ae. albopictus (F = 0.061, df = 9, p = 1.000) (fig. 2). Concentration had a highly significant effect on pupation rate (F = 728.183, df = 3, p < 0.001), while generation had no significant effect (F = 0.099, df = 3, p = 0.960).
Transgenerational changes in pupation rate of Ae. albopictus were investigated under different concentrations of deltamethrin stress. The double-distilled water was used as a blank control. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 2 Long description
The bar graph includes a legend with five entries: control, the first generation, the second generation, the third generation, the fourth generation. The x axis label is Deltamethrin concentration left parenthesis milligram slash L right parenthesis. The x axis categories are Control, 0.0001, 0.0005, 0.001. The y axis label is Pupation rate left parenthesis percent right parenthesis. The y axis range is 50 to 100. At Control, one bar is near 97. At 0.0001, four bars are near 85, 89, 99 and 95. At 0.0005, four bars are near 95, 94, 93 and 99. At 0.001, four bars are near 68, 100, 92 and 98. Several bars include vertical error bars.
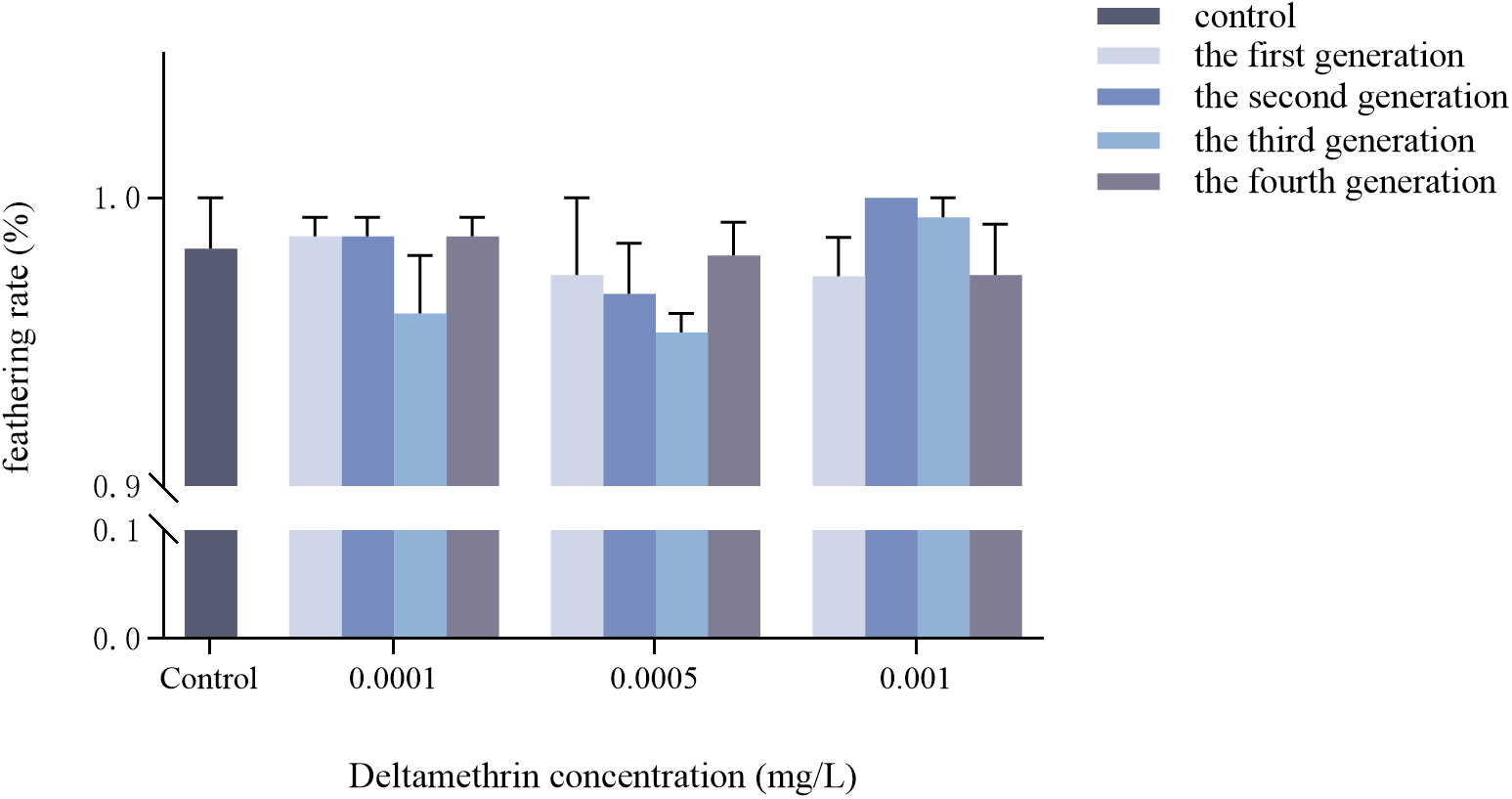
Effects of multigenerational deltamethrin exposure on the eclosion rate of Ae. albopictus
There was no significant interaction between concentration and generation on the eclosion rate of Ae. albopictus (F = 0.610, df = 9, p = 0.779) (fig. 3). Neither concentration (F = 0.992, df = 3, p = 0.409) nor generation (F = 0.440, df = 3, p = 0.726) had a significant effect on eclosion rate.
Transgenerational changes in the eclosion rate of Ae. albopictus were investigated under different concentrations of deltamethrin stress. The double-distilled water was used as a blank control. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 3 Long description
The bar graph shows hatching rates of Ae. albopictus across different deltamethrin concentrations and generations. The horizontal axis is labeled Deltamethrin concentration left parenthesis milligrams slash L right parenthesis, with categories Control, 0.0001, 0.0005 and 0.001. The vertical axis is labeled hatching rate left parenthesis percent right parenthesis, ranging from 0.9 to 1.0. Bars are vertical and grouped by concentration, with multiple bars per group representing different generations. The legend indicates control, the first generation, the second generation, the third generation and the fourth generation, each represented by different shades. At Control, the hatching rate is approximately 1.0 percent. At 0.0001 milligrams slash L, the first generation shows a hatching rate slightly below 1.0 percent, while the fourth generation is slightly above 0.9 percent. At 0.0005 milligrams slash L, the hatching rates are similar across generations, all close to 1.0 percent. At 0.001 milligrams slash L, the hatching rates remain consistent, with slight variations among generations. Error bars indicate variability. Overall, hatching rates are stable across concentrations, with minor differences between generations.
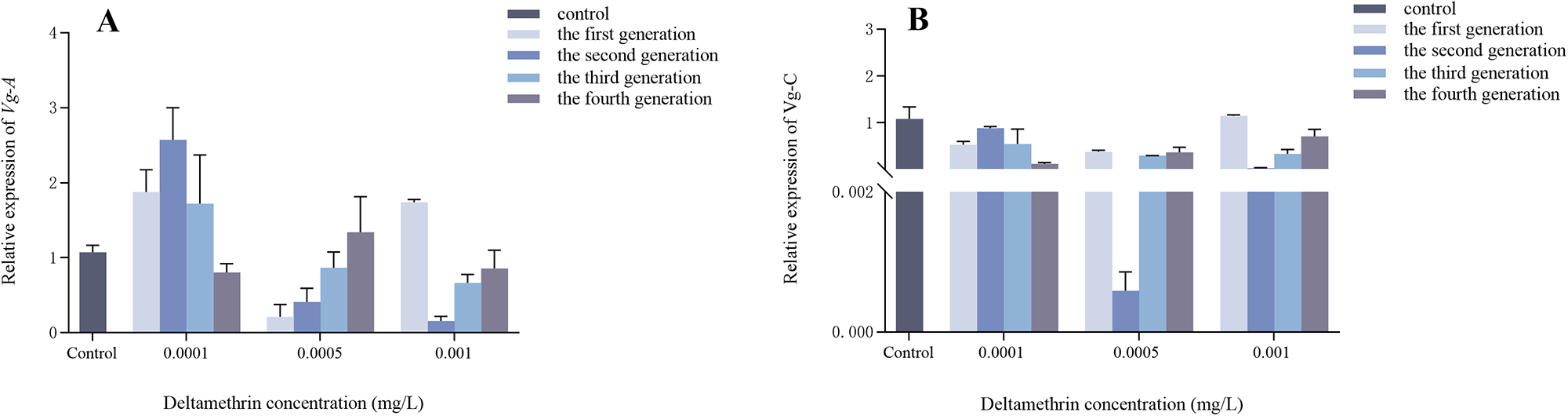
Effects of multigenerational deltamethrin exposure on Vg expression in Ae. albopictus
As shown in fig. 4A, a significant interaction was found between concentration and generation on Vg-A gene expression in female Ae. albopictus (F = 5.524, df = 9, p < 0.001). Concentration had a highly significant effect on Vg-A expression (F = 11.705, df = 3, p < 0.001), whereas generation had no significant effect (F = 0.461, df = 3, p = 0.712).
Generational changes in Vg expression were assessed in Ae. albopictus following a 24-h treatment with varying concentrations of deltamethrin, while double-distilled water was used as the control group. qRT-PCR was employed to measure the relative expression levels of Vg. (A) Vg-A. (B) Vg-C. Relative expression levels were calculated in comparison with control group, which was ascribed an arbitrary value of 1. Three biological replicates and three technical replicates were set up. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 4 Long description
The image consists of two bar graphs labeled A and B. Graph A displays the relative expression levels of Vg-A gene across different generations and deltamethrin concentrations. The x-axis is labeled 'Deltamethrin concentration (mg/L)' with categories: Control, 0.001, 0.005 and 0.01. The y-axis is labeled 'Relative expression of Vg-A'. Bars represent control, first generation, second generation, third generation and fourth generation. Key values include higher expression at 0.01 mg/L for the fourth generation. Graph B shows the relative expression levels of Vg-C gene under similar conditions. The x-axis is labeled 'Deltamethrin concentration (mg/L)' with the same categories. The y-axis is labeled 'Relative expression of Vg-C'. Bars represent control, first generation, second generation, third generation and fourth generation. Expression levels are relatively stable across concentrations, with slight variations. Both graphs use vertical bars and include a legend for generational differentiation.
A significant interaction was detected between concentration and generation on Vg-C gene expression in female Ae. albopictus (F = 2.419, df = 9, p = 0.032) (fig. 4B). However, neither concentration (F = 2.076, df = 3, p = 0.123) nor generation (F = 0.671, df = 3, p = 0.576) had a significant effect on Vg-C expression.
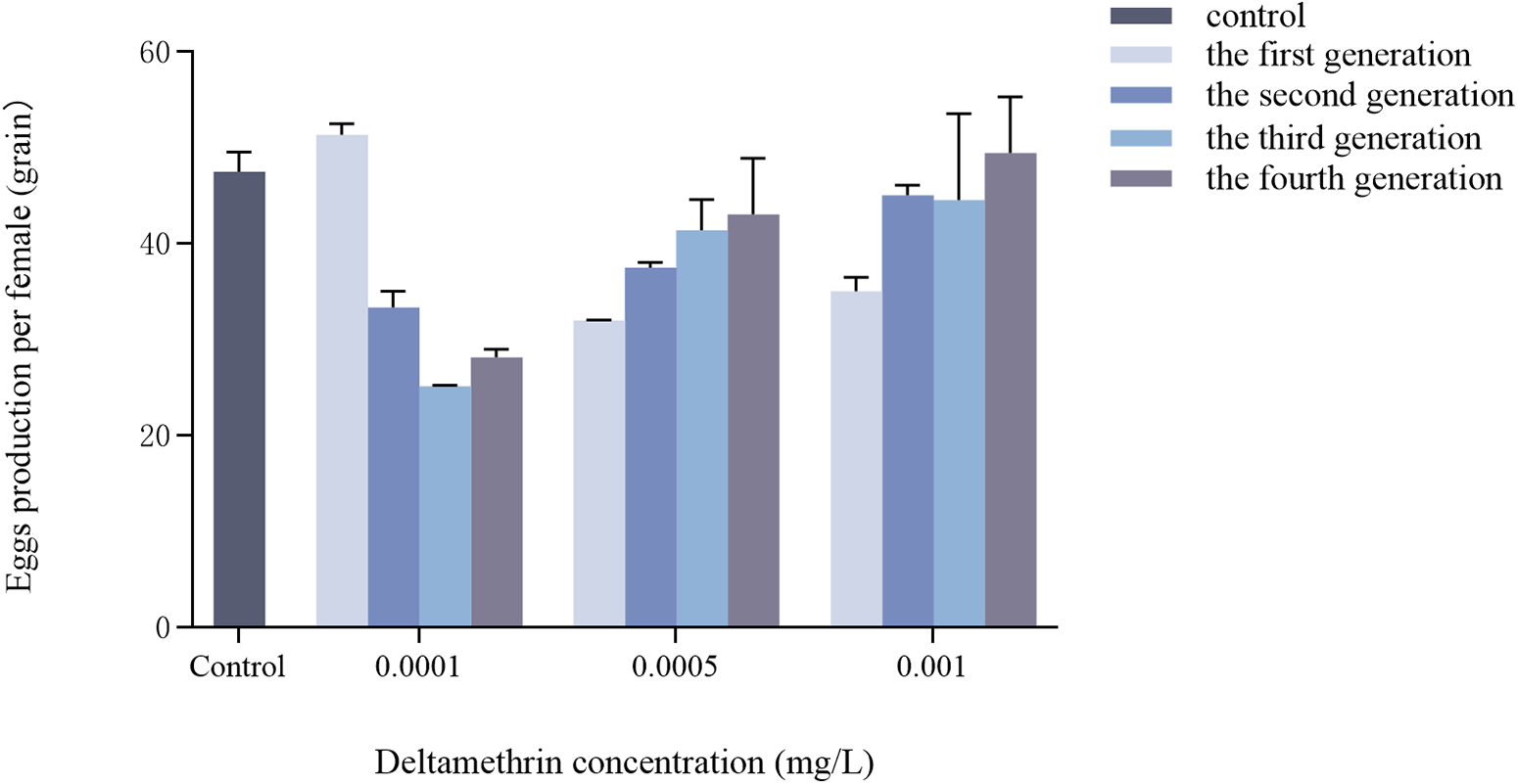
Effects of multigenerational deltamethrin exposure on the single-female egg production of Ae. albopictus
No significant interaction was observed between concentration and generation on the number of eggs laid per female Ae. albopictus (F = 2.079, df = 9, p = 0.062) (fig. 5). Concentration had a significant effect on fecundity (F = 3.059, df = 3, p = 0.042), while generation had no significant effect (F = 0.230, df = 3, p = 0.875).
Transgenerational changes in single-female egg production of Ae. albopictus were investigated under different concentrations of deltamethrin stress. The double-distilled water was used as a blank control. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 5 Long description
A grouped vertical bar graph with error bars. The y axis label is Egg production per female left parenthesis egg right parenthesis. The y axis scale shows 0, 20, 40, 60. The x axis label is Deltamethrin concentration left parenthesis milligram slash L right parenthesis. The x axis categories are Control, 0.0001, 0.0005, 0.001. A legend lists five groups: control, the first generation, the second generation, the third generation, the fourth generation. At Control, five bars are shown with heights between about 25 and 50 eggs. At 0.0001, five bars are shown with heights between about 25 and 35 eggs. At 0.0005, five bars are shown with heights between about 30 and 45 eggs. At 0.001, five bars are shown with heights between about 35 and 50 eggs.
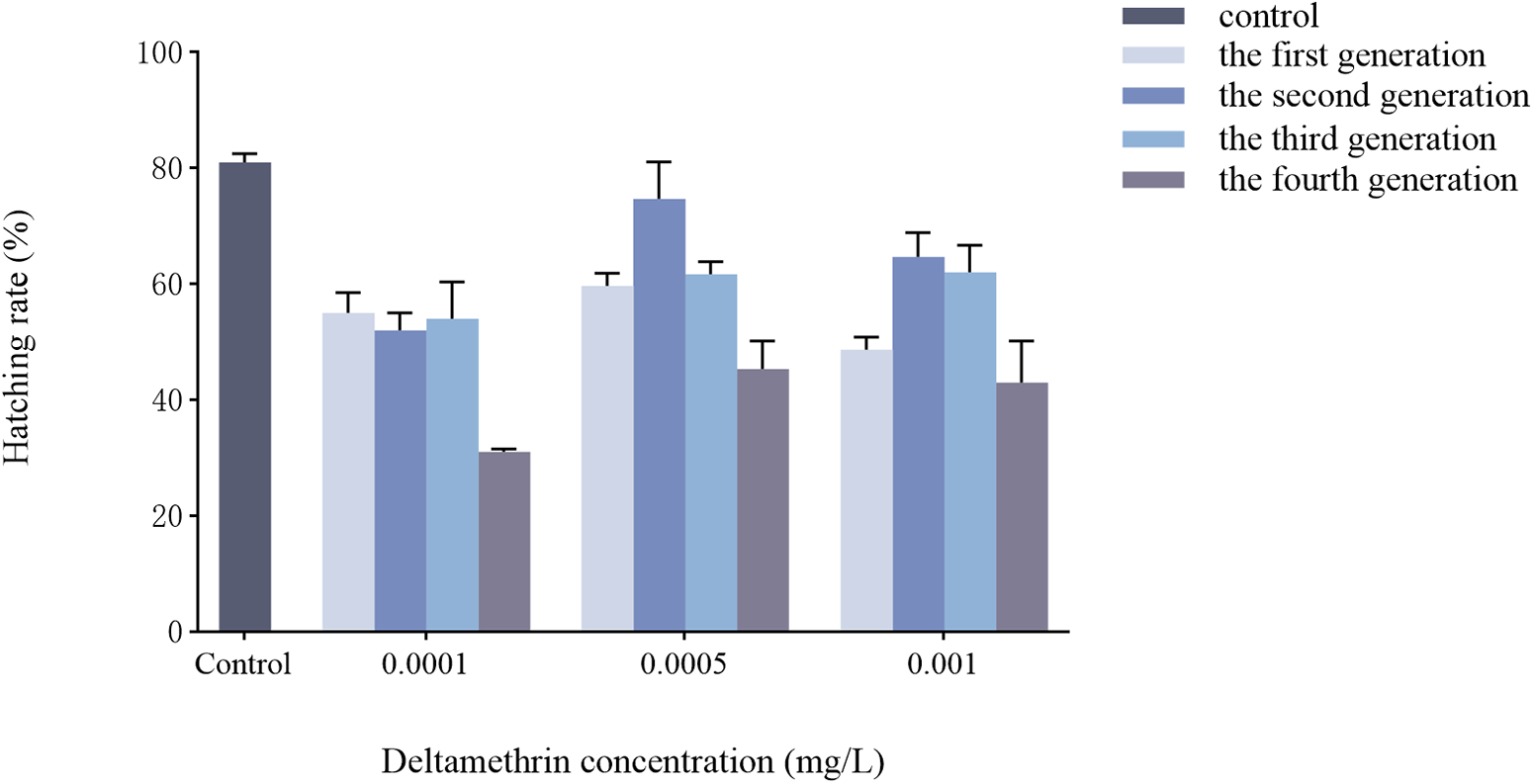
Effects of multigenerational deltamethrin exposure on the hatchability of Ae. albopictus
As shown in fig. 6, a significant interaction was found between concentration and generation on the hatching rate of Ae. albopictus eggs (F = 4.618, df = 9, p = 0.001). Both concentration (F = 27.239, df = 3, p < 0.001) and generation (F = 23.933, df = 3, p < 0.001) had highly significant effects on hatching rate.
Transgenerational changes in hatching rate of Ae. albopictus were investigated under different concentrations of deltamethrin stress. The double-distilled water was used as a blank control. Data are presented as mean ± SEM (n = 3 biological replicates per group).

Figure 6 Long description
The bar graph compares hatching rates of Ae. albopictus eggs across four generations under varying deltamethrin concentrations. The x-axis represents deltamethrin concentration in milligrams per liter, with categories: Control, 0.0001, 0.0005 and 0.001. The y-axis shows hatching rate percentage, ranging from 0 to 100 percent. Each concentration category includes bars for control, first generation, second generation, third generation and fourth generation, differentiated by shading. Error bars indicate variability. At 0.0001 milligrams per liter, the first generation shows the highest hatching rate, while the fourth generation is lowest. At 0.0005 milligrams per liter, the second generation has the highest rate. At 0.001 milligrams per liter, the control group maintains the highest rate. Overall, hatching rates decrease with higher concentrations and later generations tend to have lower rates.
Discussion
In this study, all three deltamethrin treatments significantly prolonged the larval developmental duration of Ae. albopictus compared to the control group reared in ddH2O (fig. 1). However, this effect did not follow a simple linear pattern. At the lowest concentration (0.0001 mg/L), developmental duration initially increased but then decreased relative to the control, whereas at higher concentrations (0.0005 and 0.001 mg/L), a fluctuating increasing trend across generations was observed. Our findings are consistent with previous studies demonstrating that sublethal doses of pyrethroids can disrupt normal insect development (Kumar and Chapman, Reference Kumar and Chapman1984; Krueger et al., Reference Krueger, Hanford, Weissling, Vélez and Anderson2021). For instance, Zuo et al. (Reference Zuo, Wang, Lin, Li, Peng, Piñero and Chen2016) reported that high-potency cypermethrin significantly extended the total developmental time of Rhopalosiphum padi. However, while these studies primarily focused on single-generation exposure, our results further reveal that such developmental disruptions can persist and fluctuate across multiple generations. This pattern indicates that the effects of long-term exposure to low concentrations of deltamethrin on developmental timing are not necessarily constant but may vary as exposure continues across generations.
Similar multigenerational or sustained effects have been documented in other aquatic invertebrates. Njattuvetty et al. (Reference Njattuvetty Chandran, Fojtova, Blahova, Rozmankova and Blaha2018) found that exposing Chironomus riparius – a dipteran species sharing an aquatic larval stage with Ae. albopictus – to sublethal concentrations of the neonicotinoid insecticide imidacloprid for 10 days significantly reduced larval growth. This parallel is particularly informative, as the shared life-history traits between chironomids and mosquitoes strengthen the inference that sublethal insecticide exposure can broadly impact developmental parameters in aquatic Diptera. Additionally, exposure to bisphenol compounds has been shown to delay reproductive initiation in Diaphanosoma celebensis and reduce molting frequency in Daphnia magna (Chen et al., Reference Chen, Li, Li, Yuan, Li and Liu2021; Cho et al., Reference Cho, Jeong and Lee2022), suggesting that disruption of endocrine or metabolic pathways may represent a common mechanism underlying such persistent developmental effects across taxa.
Regarding pupation and eclosion rates (figs 2 and 3), there was no significant interaction between concentration and generation on them. These results differ from previous studies on sublethal doses of pyrethroids, which have been shown to significantly reduce insect pupation and eclosion rates (Alvarez-Gonzalez et al., Reference Alvarez-Gonzalez, Briceño, Ponce-Garcia, Villanueva‐Segura, Davila‐Barboza, Lopez‐Monroy, Gutierrez‐Rodriguez, Contreras‐Perera, Rodriguez‐Sanchez and Flores2017; Osoro et al., Reference Osoro, Machani, Ochomo, Wanjala, Omukunda, Munga, Githeko, Yan and Afrane2021); Deltamethrin deposited in the environment inhibits the eclosion of Chironomus tentans (Maul et al., Reference Maul, Brennan, Harwood and Lydy2008). The relatively low concentrations used in this study may explain the lack of effect. Recently, Hu et al. (Reference Hu, Zhang, Tang, Liu, Xia, Wang, Li, Gao, Li, Lv and Yang2024) reported that pupation and eclosion rates did not change significantly in a λ-cyhalothrin-resistant strain of Cydia pomonella, consistent with our findings. This stability may reflect physiological stress responses to prolonged insecticide exposure, indicating that key developmental processes were maintained under sublethal stress.
Considering the larval developmental duration, eclosion rate, and pupation rate collectively, it was observed that the low concentration of deltamethrin treatment in this experiment exerted a more pronounced impact on the developmental duration of Ae. albopictus. However, its effect on the eclosion rate and pupation rate of Ae. albopictus was not statistically significant, possibly due to the relatively low concentration employed in this study during the larval stage. This lower level of epidermal hardening and weaker osmotic resistance may have contributed to the reduced effect observed in this treatment group. During the pupal and nymphal stages, insects undergo gradual epidermal sclerotisation. The body surface serves as the first barrier to insecticide penetration (Togawa et al., Reference Togawa, Augustine Dunn, Emmons and Willis2007), and increased sclerotisation has been associated with reduced cuticular penetration of pyrethroids (Balabanidou et al., Reference Balabanidou, Grigoraki and Vontas2018). Whether such mechanisms operate in Ae. albopictus under sublethal deltamethrin exposure, and whether they allow for metabolic self-regulation, requires further investigation. Furthermore, it has been demonstrated that the surface area to volume ratio significantly influences organism sensitivity. It is plausible that smaller organisms with larger surface area to volume ratios may experience an accelerated accumulation rate of toxic substances in water. Simultaneously, their lower fat to protein ratio, enhanced toxin build-up in organs, an underdeveloped antioxidant system, and a less developed physical structure could contribute to increased susceptibility. These factors potentially explain why the low concentration of deltamethrin had a more pronounced impact on the developmental stages of Ae. albopictus larvae compared to fourth instar larvae and pupae (Cadmus et al., Reference Cadmus, Kotalik, Jefferson, Wheeler, McMahon and Clements2019).
Vitellogenin (Vg) is a glycosphingolipid protein specifically found in the hemolymph of sexually mature oviparous females and serves as the precursor for yolk protein (YP), which is essential for egg yolk synthesis. Vg exhibits a strong association with insect reproductive capacity and is thought to play an important role during embryonic development in oviparous animals. The extent of its utilisation has been reported to be closely linked to insect egg production (Tufail and Takeda, Reference Tufail and Takeda2008; Reading and Sullivan, Reference Reading and Sullivan2011; Amano et al., Reference Amano, Uno, Koyama, Hiramatsu, Todo and Hara2019). It should be noted, however, that the associations between Vg expression and fecundity observed in the present study are correlative rather than causal. The relative expression trends of Vg-A and Vg-C encoding YPs in Ae. albopictus were consistent under three concentration gradients (fig. 4A, B). Under the 0.0001 mg/L treatment, Vg expression showed an initial increase followed by a decrease across generations, while single-female egg production gradually declined (figs 4A, B and 5). In the first two generations, the trends in Vg expression and egg production were not consistently aligned, suggesting that transcriptional changes in Vg do not always directly translate into reproductive output. This may reflect post-transcriptional regulation, delayed effects on ovarian development, or other physiological constraints. In later generations, both Vg expression and egg production decreased, suggesting a possible multigenerational physiological effect of deltamethrin on reproductive physiology. At 0.0005 mg/L, both Vg genes exhibited a generation-by-generation increasing trend, which paralleled the trend in egg production. At 0.001 mg/L, Vg expression showed an initial decrease followed by an increase, whereas egg production gradually declined across generations. These concentration-dependent and generation-dependent patterns suggest a complex relationship between Vg transcription and reproductive output under sublethal insecticide stress. However, several limitations should be considered when interpreting these results. First, Vg expression was measured at a single adult time point (1 day post-eclosion), which may not capture dynamic changes throughout the vitellogenic cycle. Second, we did not examine ovarian morphology or oocyte developmental stages, which would provide direct evidence of reproductive impairment. Therefore, the observed associations between Vg expression and fecundity remain correlative rather than causal. Study by Hahn et al. (Reference Hahn, Schenk and Schulz2002) on the aquatic midge Chironomus riparius have shown that environmental chemicals can disrupt vitellogenesis and YP content, demonstrating that the relationship between xenobiotic exposure and reproductive processes in aquatic insects is complex and likely involves multiple hormonal factors (Valle, Reference Valle1993; Dong et al., Reference Dong, Ye, Guo and Hu2009). The expression of Vg genes is also closely associated with Vg receptor dynamics. Further studies incorporating time-series sampling, ovarian histology, and investigation of hormonal regulation and receptor mechanisms are needed to elucidate the specific pathways by which deltamethrin affects Ae. albopictus reproduction.
Regarding single-female egg production (fig. 5), this study revealed that deltamethrin exposure exerted concentration-dependent effects on the fecundity of Ae. albopictus. At the lowest concentration (0.0001 mg/L), single-female egg production progressively decreased across successive generations. In contrast, at higher concentrations (0.0005 and 0.001 mg/L), egg production showed a gradual increase over generations, although it remained below or comparable to control levels in each generation. Notably, the interaction between concentration and generation on egg production was not statistically significant, suggesting that the main effect of concentration may be the primary driver of the observed variation in fecundity. These findings are consistent with a growing body of evidence demonstrating that pyrethroid insecticides can impair insect reproductive output. Early studies documented reduced oviposition in Aedes aegypti following pyrethroid exposure (Liu et al., Reference Liu, Todd and Gerberg1986), and subsequent research has extended this observation to other species: sublethal concentrations of high-potency cypermethrin significantly reduce egg production in Drosophila suzukii (Shaw, Reference Shaw2019). Similarly, exposure to sublethal concentrations of deltamethrin reduces egg production in immature Chironomus columbiensis (Montaño-Campaz et al., Reference Montaño-Campaz, Dias, Bacca, Toro-Restrepo and Oliveira2022). The convergence of findings across multiple taxa suggests that pyrethroid-induced reproductive suppression may be a generalised phenomenon.
However, the concentration–dependent response pattern observed in this study, specifically a decline in fecundity at the lowest concentration and partial recovery at higher concentrations across generations, suggests a potential non-linear dose–response relationship. This pattern may reflect the induction of physiological compensatory mechanisms at higher exposure levels, or alternatively, the modulation of low-concentration cumulative effects by alternative regulatory pathways under increased chemical stress. Notably, previous studies have demonstrated that pyrethroid insecticides may also exert a stimulatory effect on the oviposition activity of insects. This is consistent with our observation of increased egg production across multiple generations at concentrations of 0.0005 and 0.001 mg/L deltamethrin. Bunn et al. (Reference Bunn, Balanay and Richards2019) reported that sublethal doses of deltamethrin enhanced the egg-laying ability of Nilaparvata lugens. One possible mechanism underlying this stimulatory effect involves male reproductive systems: deltamethrin treatment significantly elevated levels of reproduction-associated male accessory glands proteins in males of N. lugens, which were then transferred to females through mating, ultimately leading to increased female egg production (Ge et al., Reference Ge, Wang, Zhao, Wu and Huang2010). These findings suggest that reproductive outcomes following sublethal insecticide exposure may reflect a net balance between direct toxic effects on females and indirect stimulation via male-mediated mechanisms.
The hatching rate of Ae. albopictus was significantly reduced in the 0.0001, 0.0005, and 0.001 mg/L deltamethrin treatments compared to the control group treated with distilled water (fig. 6). However, the response patterns differed by concentration: at the lowest concentration (0.0001 mg/L), hatching rates exhibited a fluctuating decline across generations, whereas at higher concentrations (0.0005 and 0.001 mg/L), an initial increase followed by a decline was observed. These concentration-dependent patterns suggest that sublethal deltamethrin exposure disrupts reproductive success, with the direction and magnitude of effects modulated by exposure level and generational timing. Daaboub (Reference Daaboub2018) demonstrated that high deltamethrin-resistant populations of Anopheles labranchiae had significantly lower hatching rates than sensitive populations. Poumachu et al. (Reference Poumachu, Lontsi-Demano, Zeb, Djoufounna, Djiappi-Tchamen, Hashem, Alajmi, Mochie, Awono-Ambene, Antonio-Nkondjio, Tchuinkam and Abd-Allah2025), on the other hand, found that deltamethrin treatment significantly reduced the hatching rate of Culex quinquefasciatus eggs. These results are consistent with previous findings indicating that sublethal doses of pyrethroids can considerably reduce insect hatching rates (Crawley et al., Reference Crawley, Gordon, Kowles, Potter and Haynes2017). This implies that the toxicity of deltamethrin exerts an inhibitory influence on the hatching process of Ae. albopictus eggs, potentially by disrupting female mosquito oviposition behaviour, resulting in inviable or empty eggs and a reduced rate of normal egg hatching. Additionally, the gradual accumulation of deltamethrin within Ae. albopictus may enhance its reproductive system toxicity, ultimately leading to a sublethal effect.
In summary, our study demonstrates that even a single generation of exposure to low concentrations of deltamethrin imposes physiological stress on Ae. albopictus, However, prolonged exposure to deltamethrin and its accumulation in successive generations can still cause harm to Ae. albopictus. These findings align with growing evidence that sublethal insecticide exposure can reduce population fitness and drive physiological or life-history adjustments (Gressel, Reference Gressel2011; Bantz et al., Reference Bantz, Camon, Froger, Goven and Raymond2018). Individuals surviving such exposure may contribute to population recovery through reproduction, potentially sustaining population density and complicating vector control efforts. Consequently, greater attention must be paid to the management of environmental insecticide residues – particularly in aquatic habitats – and the application rates of pyrethroids should be carefully regulated to minimise accumulation. It should be noted that the laboratory strain used in this study is insecticide-susceptible, whereas field populations are often pre-exposed to various agrochemicals and may possess enhanced stress response mechanisms. Furthermore, although our continuous exposure design provides controlled conditions, it does not fully replicate the fluctuating deltamethrin concentrations found in natural aquatic environments due to degradation and adsorption. Of further note, target site mutations remain unexamined and warrant further investigation. Finally, it is important to acknowledge the low replication level in this study (n = 3 per group per generation); therefore, future studies with larger sample sizes are needed to confirm and extend our findings. Despite these limitations, our results provide a physiological framework for understanding how sublethal deltamethrin exposure can induce transgenerational fitness costs in Ae. albopictus. Future work should focus on quantifying insecticide pollution levels in field habitats, assessing the corresponding physiological responses in local mosquito populations, and evaluating life-history trade-offs under realistic exposure scenarios. Such integrated approaches will improve our understanding of sublethal insecticide effects on mosquito population dynamics and support the development of sustainable vector management strategies.
Author contributions
JY.S.: Methodology, Investigation, Visualization, Writing – original draft. C.Z.: Methodology, Writing – original draft, Writing – review and editing. RY.L.: Investigation, Writing – review and editing. JY.X.: Writing – original draft. JC.Y.: Visualization. B.T.: Conceptualization, Funding acquisition. SG.W.: Conceptualization, Funding acquisition.
Financial support
This work was supported by the National Natural Science Foundation of China [Grant Numbers 32272608 and 31672081].
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethics approval and consent to participate
All animal procedures have been approved by the Experimental Animal Ethics Committee (AEWC) of Hangzhou Normal University (Approval No. HSD-20240719-02).
Data availability
Data will be made available on request.