



Impact Statements
Walter G. (Walt) Whitford was a remarkable ecologist who loved to study termites that live in the soils of drylands. Australia has an abundance of termites in its dry regions, so Walt visited Australia in 1986–1987 to study them. Walt was a guest of the CSIRO’s Rangelands Research Laboratory in Deniliquin, NSW. We studied termites in the dry mulga woodlands about 40 km northwest of Louth, NSW. We studied how, when mulga trees died, they often fell over, and soil accumulated around the fallen logs to form accretion mounds. Walt showed us how termites made coverings of soil over the surface of these mulga logs to feed on them, and how they also went inside the logs to feed. The termites lived below the surface and would build tunnels from nests deep in the soil to bring sand-sized soil particles up to build their coverings. We found that not only were the termites consuming the dead logs but their activity helped to create mounds of soil that were much richer in nutrients, such as nitrogen, than the soils away from the log mounds. Furthermore, because of the network of termite tunnels below the mounds, rainwater would soak into their soils much faster than away from the mounds. Thanks to Walt, these findings not only gave us a new perspective on the importance of termites in drylands but also this perspective led to our book on “Restoring Disturbed Landscapes,” written to help land owners around the world better manage their vast drylands.
Introduction
Social relationships are critically important in all disciplines, and no less in science, yet examples of these relationships are seldom reported in formal scientific literature. An example of these social relationships is the synergy that results from personal associations among scientists, often from different countries and disciplines and with different levels of experience. The combination of different perspectives often leads to significant innovations and to outcomes that are greater than would occur when scientists work alone (Leigh Reference Leigh2026). These relationships are also important for motivating early-career scientists (Yang Reference Yang2016) where mentors provide advice and support freely (Lee Reference Lee2002). An example of such a synergy in dryland ecology occurred when Professor Walter G. (Walt) Whitford visited Australia in 1986–1987 to review the Commonwealth Scientific and Industrial Research Organisation’s (CSIRO) Rangelands Research program. The partnerships resulting from this visit have endured and led to a better understanding of ecological processes and functions across the world’s drylands.
Walter (Walt) Whitford loved to study small fauna such as soil mites and termites, as well as large fauna, as seen in his passion for observing birds. In fact, Walt loved and had a remarkable understanding of all desert organisms and the ecosystems and landscapes they lived in. Walt was truly a rare general ecologist who integrated studies across scales (Eldridge and Kerley, Reference Eldridge and Kerleyin press), as Walt’s career advanced from a study of localized salamander populations to deserts. He fully embraced collaborative and interdisciplinary studies. Walt believed that the positive synergies emerging from working closely with others produced a greater understanding of dryland systems, far more than from working alone. Synergies from interdisciplinary research are also important in many other systems (Gupta et al., Reference Gupta, Mera, Seshanna, Suryanarayana, Sharma and Coral2025).
Rarely did Walt publish as a single author (e.g., Whitford, Reference Whitford2002). Walt planned, conducted and published interdisciplinary studies on dryland systems in collaboration with established researchers from many continents, including Asia, Australia, Europe, South Africa and South America. Walt enjoyed mentoring, and he fostered and published dryland studies with early career academics, postdoctoral fellows, graduate students and undergraduate students. Walt firmly believed that ideas should be shared and that knowledge is gained when freely given (Vangelisti and Perlman, Reference Vangelisti and Perlman2018).
In this article, we aim to provide a perspective on the synergies resulting from research that Walt did in Australia in 1986–1987 with us and other colleagues at the CSIRO Rangelands Research Laboratory in Deniliquin, New South Wales. As background, in the 1960s, a Rangeland Research Unit had been created in CSIRO in Canberra, which was led by Mr. Ray Perry, who promoted the need to study Australia’s vast and dry rangelands. In 1969, when Ted Moore was appointed Officer-in-Charge of the CSIRO Riverina Laboratory at Deniliquin, the research focus changed from precision irrigated agriculture to dryland-rangeland science.
To illustrate Walt’s collaboration with us and other colleagues in Australia, we summarize two case studies published with Walt on termites in the semi-arid woodlands. First, we examine how termites played a role in the functioning of finer-scale landscape features, such as in mulga (Acacia aneura) log mounds (Tongway et al., Reference Tongway, Ludwig and Whitford1989). Second, we will look at coarser scales – how termites contributed to the functioning of systems where mulga trees formed distinctive patches and banded patterns in landscapes (Tongway and Ludwig, Reference Tongway and Ludwig1990; Whitford et al., Reference Whitford, Ludwig and Noble1992). Finally, we provide a perspective on how Walt’s research in Australia, and elsewhere, has led to current theoretical and practical progress in ecology.
The Lake Mere study area
Our studies with Walt were conducted in the semiarid woodlands of southeastern Australia. These woodlands are described by Harrington et al. (Reference Harrington, Mills, Pressland, Hodgkinson, Harrington, Wilson and Young1984) as having a vegetation of small trees, mostly mulga (A. aneura), and native grasses such as woollybutt (Eragrostis eriopoda) and Bandicoot grass (Monachather paradoxa). These woodlands occur on massive red-earth soils of coarse-to-medium texture and neutral acidity (Mabbutt and Fanning, Reference Mabbutt and Fanning1987). They are strongly patterned from fine-scale clumps of grasses and soil mounds formed by fallen mulga logs to coarser-scale bands of grasses and trees (Tongway and Ludwig, Reference Tongway and Ludwig1990; Ludwig and Tongway, Reference Ludwig and Tongway1995).
Within these semiarid woodlands, we conducted specific studies at a CSIRO study site located 40 km northwest of Louth on a sheep station, Lake Mere. Louth is on the Darling River in NSW. This site was established in 1985 to explore differences in how kangaroos and sheep utilize available dryland forage (Wilson, Reference Wilson1991). We conducted baseline measurements on the Lake Mere site (Tongway and Ludwig, Reference Tongway and Ludwig1990) and found that the topography, vegetation and soils exhibit the patchy patterns typical of the semiarid woodlands (Figure 1). Although not shown in Figure 1 because of their small size, in the field, fine-scale patches of grass clumps and mulga log-mounds are readily observed on the more open, higher runoff areas, but they become obscured within denser bands of grasses and trees occurring in run-on zones such as drainage lines, which are ephemeral and nonincised.
A diagram of the 200 ha (1,250 m × 1,600 m) Lake Mere study site showing boundaries of drainage lines with dense mulga and smaller mulga groves with gray fill. Although not shown as dots, smaller mulga log mounds are dispersed throughout the site. Slightly elevated ridge crests are indicated by dashed lines, and the direction of overland water flows as arrows. Figure 1 is redrawn from a portion of Figure 1 in Tongway and Ludwig (Reference Tongway and Ludwig1990) using ChatGPT with specific instructions to “Within the irregular boundaries of the image, replace weak dashed fill with light grey fill” and to “On the image created slightly thicken the outer boundary line”.

Mulga log-mounds: Termites affect fine-scale patch processes
Initially, it was a surprise to us when Walt showed us how termites could play a notable role in the structuring and functioning of soil mounds formed by logs of fallen mulga (Acacia aneura) trees. We had observed that mulga log mounds were a prominent finer-scale feature in many landscapes in the semiarid woodlands of eastern Australia, such as on our CSIRO Lake Mere study site (Figure 1). Our CSIRO colleague Jim Noble estimated the density of mulga log mounds at 137/ha (Nobel et al., Reference Noble, Diggle and Whitford1989). Walt showed us how termites (e.g., Amitermies spp.) build galleries of soil over the surface of the logs as they feed on the decaying woody material beneath the galleries. It was hypothesized that the origin of these gallery materials was coarser-textured soils from deep below the surface. Further, Walt showed us that other termites (e.g., Drepanitermies pernige and Tumulitermes tumuli) build “pipes” of soil inside the logs as they feed, again using more coarser-textured soils from deep below the surface. With time, the soils in these galleries and pipes, enriched by termites, would be added to build the soil mound. We hypothesized that soils in mulga log mounds would have a sandier texture and greater fertility than the soils surrounding the mound.
To bring soil up to the surface from below, these termites formed a network of “tunnels” below the mulga log mound (Figure 2). We hypothesized that these termite tunnels form stable biotic macropores of 1–2 mm in diameter, which effectively increase the rate at which water would infiltrate into the soil during rainfall events.
A diagram of a typical mulga log mound cross-section showing orientation and location of termite surface feeding and subsurface storage galleries and tunnels. The size of termite tunnels, typically 1–2 mm in diameter, has been exaggerated for illustration. Figure 2 is redrawn from Figure 1 in Tongway et al. (Reference Tongway, Ludwig and Whitford1989) with final revisions using ChatGPT with specific instructions to “Redraw the image at 6 cm by 8 cm and at a resolution of 300 dpi”.

To investigate these hypotheses, Walt worked with us to randomly select five plots on mulga log mounds on the Lake Mere site, which were paired with five plots located 3 m upslope of the selected log mounds. Mulga log mounds are typically 3–5 m long and 1–1.5 m wide. On the 10 plots, we collected samples of termite gallery and pipe material and soil samples at 0–1, 1–3, 3–5 and 5–10 cm below the surface. Six weeks after a 35 mm rainfall event in mid-April 1987, we harvested and oven-dried at 60 °C and weighed in the lab all above-ground herbage by species in 1-m2 quadrats centered within the 10 plots; this provided us with comparative plant biomass and biodiversity data for on-and-off mulga log mounds. Termite gallery and pipe materials and soil samples were analyzed in the lab for texture, organic carbon, mineralizable and total nitrogen and available phosphorus (see Tongway et al., Reference Tongway, Ludwig and Whitford1989).
Findings confirmed our hypotheses – termites strongly affected soil texture, infiltration and fertility, hence herbage biomass and diversity, on mulga log mounds at our Lake Mere site. We found from soil texture analyses that soils constructed by termites into galleries and pipes and those soils at the surface of the log mound did not differ but had significantly (P < 0.05) higher proportions of sand, hence lower proportions of silt and clay, than in soils collected from 3-m upslope (76.9% vs. 61.7% sand at 0–1 cm; see Table 1 in Tongway et al., Reference Tongway, Ludwig and Whitford1989). This indicates that termites are selectively bringing larger sand-sized soil particles to the surface to construct their galleries and pipes, and with time, these particles are deposited at the surface to help form the elevated mulga log mound.
We found that upper soil layers (0–10 cm) in mulga log mounds compared to soils 3 m upslope had on average, based on paired t-tests at 3 degrees of freedom, significantly more mineralizable nitrogen (8.73 ppm vs. 4.80 ppm; p = 0.018), marginally more extractable phosphorus (13.2 ppm vs. 10.8 ppm; p = 0.058) but no significant difference in organic carbon (0.63% vs. 0.55%; p = 0.19). These data by soil depths are illustrated in Figure 3 in Tongway et al. (Reference Tongway, Ludwig and Whitford1989) and presented in Table 2.4 in Tongway and Ludwig (Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997).
The soils within mulga log mounds had consistently higher rates of water infiltration (1.0–3.8 mm/min) than soils 3 m upslope (0.25–0.48 mm/min), as illustrated in Figure 2 in Tongway et al. (Reference Tongway, Ludwig and Whitford1989). The greater capacity of log mound soils to infiltrate water was reflected in our above-ground herbage biomass data. On average, the five mulga log mounds had 10.7 g/m2 compared to only 0.34 g/m2 on the five comparable plots 3 m upslope of the mounds (for data by species, see Table 3 in Tongway et al. [Reference Tongway, Ludwig and Whitford1989]).
From these findings, we concluded that mulga log mounds form what might be considered “fertile patches” on our Lake Mere site in the semiarid woodlands (Tongway et al. Reference Tongway, Ludwig and Whitford1989). Their sandier soils and abundant macropores created by termites provided richer habitats for plants and animals, forming what are called “environmental resource patches” (Forman and Godron, Reference Forman and Godron1981). Mulga log mounds form resource-rich patches that persist for many years, although we do not know their exact ages. Walt noted that although termites build galleries to store organic materials within the log mounds, they did not appear to use the mounds as nest sites.
Mulga tree groves: Termites affect landscape-scale processes
As illustrated by Figure 1, the CSIRO Lake Mere study site was strongly patterned with dense bands of mulga trees occurring in weakly dendritic, non-incised drainage lines and with mulga trees also forming small groves on open runoff slopes (Tongway and Ludwig, Reference Tongway and Ludwig1990). Importantly, the mulga groves on these open slopes formed bands with intervening intergroves repeating downslope, and this pattern functioned to capture and retain water, soil sediments and litter during rainfall events (Ludwig and Tongway, Reference Ludwig and Tongway1995; Tongway and Ludwig, Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997). Further, we found these grove-intergrove banding patterns occurred on other sites in addition to Lake Mere; they are now known for many other areas across the drylands of Australia and in other countries with extensive drylands having a similar topography (Tongway et al., Reference Tongway, Valentine and Segheri2001).
During our studies at Lake Mere on the role of termites in the structure and functioning of fine-scale mulga log mounds, described above, Walt showed us examples of how termites were also quite abundant and likely to be very important in the functioning of the mulga grove-intergrove system. Walt showed us that termites would build soil-based foraging galleries over piles of litter, on dung, over dead grass tussocks and on dead wood. During Walt’s visit in 1986–1987, we only had time to complete the mulga log-mound study. But, when Walt came for a second visit in 1990 (with his son Brett Whitford), we all teamed up with CSIRO colleague Jim Noble to conduct a study on Lake Mere to quantify the abundance of different termite foraging galleries in mulga groves and intergroves. Later, after the Whitford returned to New Mexico, we conducted follow-up studies to test if sites with the greatest abundance of termite galleries also had greater soil fertility and higher infiltration rates.
To survey termite foraging gallery abundance in mulga groves and intergroves, 100-m lines were laid out at random within each of 12 paddocks established for a kangaroo-sheep forage utilization study (Wilson, Reference Wilson1991). Along each 100-m line, ten 1-m2 quadrats were examined at 10-m intervals. In the 120 quadrats, the percentage cover of termite galleries on different materials was recorded. The study was conducted 3 weeks after a 72 mm rainfall event.
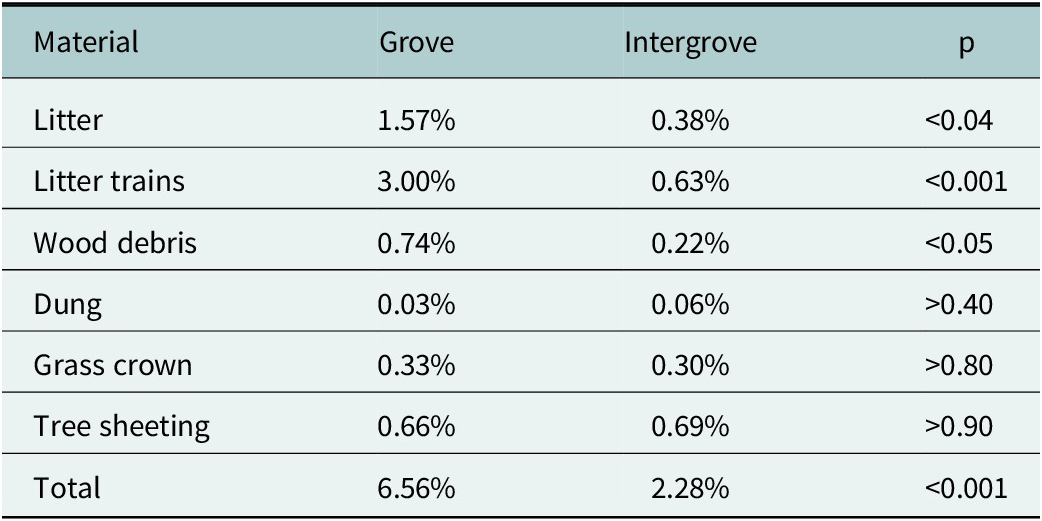
We found that termite foraging galleries significantly (p < 0.05) covered on average over 6.5% of the 1-m2 quadrats in mulga groves compared to only 2.3% for intergroves (Table 1). The greatest cover of foraging galleries was on litter materials, especially on litter trains, which form from overland flows during notable rainfall events, where litter materials, such as leaves and stems, are deposited in distinctive lines (trains) across gentle slopes in places where water flows are impeded by obstructions such as grass tussocks and rocks. Woody debris, such as sticks within quadrats, also had significantly more termite gallery covering in mulga groves than in intergroves. Termites would even construct sheetings of foraging galleries on the trunks of old, but living, mulga trees in both groves and intergroves. Other materials, dung and crowns of old grass clumps, had insignificant differences in gallery coverings.
Percentage cover of termite foraging galleries on six different materials in mulga groves compared to intergroves at the CSIRO Lake Mere study site

Note: Data after Whitford et al. (Reference Whitford, Ludwig and Noble1992). Statistical significance (p) based on paired t-tests.
The termites that build these above-ground foraging galleries are subterranean, primarily xylophagous, termites such as the Amitermes spp., which are found in the mulga log mounds, and harvester termites, such as Drepanotermes perniger, which build hydrophobic caps or soil pavements over their nests – at Lake Mere, there were an estimated 58 termite pavements per hectare (Noble et al., Reference Noble, Diggle and Whitford1989). These subterranean termites bring soil particles up through networks of tunnels to build their foraging galleries. As we found for mulga log mounds (Tongway et al., Reference Tongway, Ludwig and Whitford1989), we asked if termites would also enhance water infiltration and enrich fertility in mulga tree groves?
To answer this question, we conducted another study on Lake Mere where we established a line transect oriented down a gentle slope that intercepted a number of repeating mulga tree groves and intergroves. Soil samples were collected within and between these groves at depth intervals of 0–1, 1–3, 3–5 and 5–10 cm. Soil samples were taken to the Deniliquin CSIRO Lab for nutrient analyses.
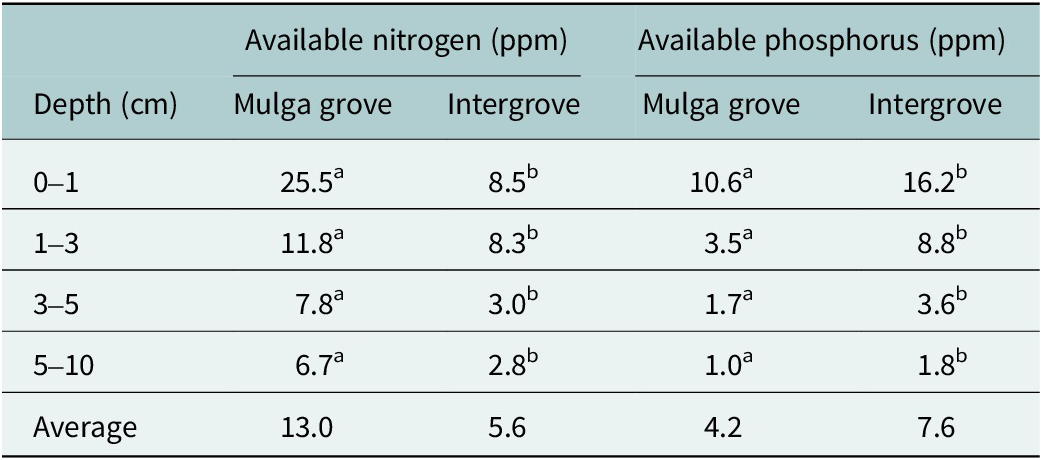
We found that soils down to 10 cm had significantly (p = 0.05) higher concentrations of potentially available nitrogen (AN) within mulga tree groves (average = 13.0 ppm) compared to 5.6 ppm in the intergroves (Table 2). Potentially available phosphorus (AP) was opposite to AN, where AP was significantly higher down to 10 cm in the intergroves (7.6 ppm) compared to within the mulga groves (4.2 ppm).
Available nitrogen and phosphorus in soils sampled to 10 cm depth within mulga tree groves and intergroves on the CSIRO Lake Mere site in the semiarid woodlands of eastern Australia
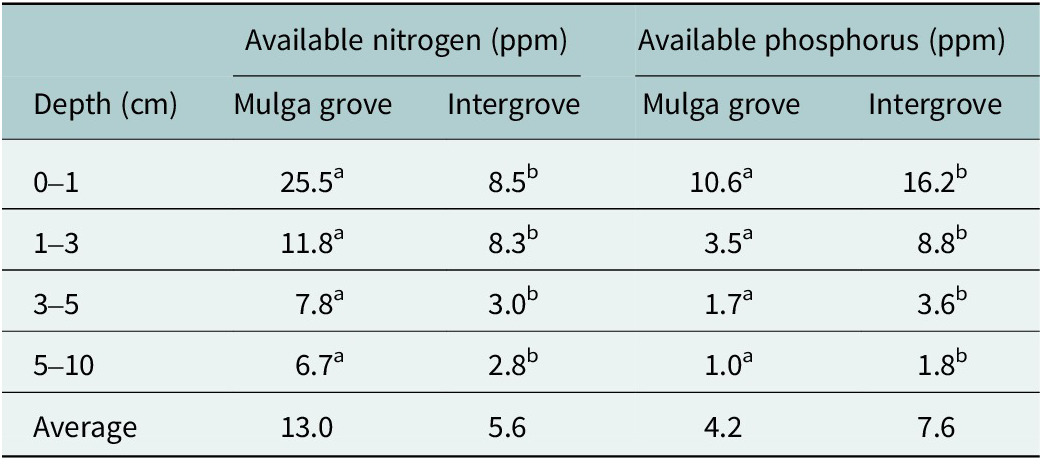
Note: Data from Table 3 in Tongway and Ludwig (Reference Tongway and Ludwig1990). For each nutrient and soil depth, soils with different superscript letters are significantly different (p = 0.05). Statistical significance (p) based on paired t-tests.
Do the subterranean tunnels created by termites also enhance soil infiltration rates in mulga tree groves compared to intergroves, as was the case for mulga log mounds? To answer this question, we looked at data collected by our CSIRO colleague Richard Greene after a single rainfall event of 37.5 mm at Lake Mere (Greene Reference Greene1992). Richard found that soils of intergroves only infiltrated 15.7 mm or 42% of the incident rain – thus, over half of the rainfall was runoff. Much of this runoff was captured as run-on by a mulga grove, which infiltrated 51.7 mm or 138% of the incident rainfall. Furthermore, Richard found that within a mulga tree grove, more than 90% of the rainwater infiltrated through the biopores created by termites and root channels.
We summarized and interpreted these findings within a theoretical framework for the conservation of water and nutrients with dryland landscapes (Tongway and Ludwig, Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997). As illustrated in Figure 3, intergrove zones between individual groves or bands of mulga trees have low soil infiltration rates, hence generate runoff during notable rainfall events, which in turn is largely captured as run-on by mulga groves, where the leading or upslope edge is often grassy with a moderate rate of water infiltration. The termites not only enhance infiltration by creating stable biopores, but they also actively help to decompose organic matter in litter and dung, for example, to enhance soil fertility, notably available nitrogen. We can conclude that mulga tree groves are another type of “fertile patch” in the semiarid landscapes of eastern Australia, and that mulga grove-intergrove systems enable greater control of excessive runoff and erosion at the local watershed scale.
A typical cross-section of the mulga grove-intergrove landscape found in the semiarid woodlands of eastern Australia. The steepness of the intergrove slope has been exaggerated (typically 1–2%). Figure 3 is redrawn from Figure 2.3 in Tongway and Ludwig (Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997) with final revisions using ChatGPT with specific instructions to “Redraw the image at 4 cm by 12 cm and at a resolution of 300 dpi”.

A landscape-level perspective
The visit by Walt to Australia in 1986–1987 was a revelation of how the biological processes of termites were strongly related to soil condition properties, including water infiltration, organic matter decomposition and nutrient availability in semiarid landscapes. From ongoing discussions with Walt, we concluded that four scales of observation/sampling were needed: (1) fine scale (mm to about half-a-meter), mostly biological features and processes, (2) medium scale (0.5–50 m), dominated by mostly physical processes such as runoff, sediment deposition and erosion, (3) macro-scale (50 m to more than 1 km), the size of the local catchment and (4) regional scale (much greater size, as in mapping land systems). Walt alerted us to the importance of the finest scale by demonstrating how above and below-ground dwelling termites have processes such as consumption, digestion and defecation of macro-organic matter that are critically important mechanisms for recycling recalcitrant organic matter (e.g., mulga logs and sheep feces). Termites also establish stable pores in the soil, which greatly affect the water infiltration rate and the transport of oxygen into lower soil layers by diffusion (Tongway et al., Reference Tongway, Ludwig and Whitford1989). In return, as colleagues, we perhaps expanded Walt’s view from how termites functioned at fine scales to a greater emphasis on their role in patterns and processes within broader landscapes – an interaction we know produced a combined great effect, a synergy.
While Walt’s time in Australia was limited to a study leave in 1986–1987, and a shorter return visit in 1990, these visits not only resulted in new publications with Walt – as illustrated above by the two case studies – but his mentoring encouraged us to expanded into whole new lines of theoretical and practical research, such as the use of soil surface indicators and landscape function analysis in landscape ecology (Tongway and Ludwig, Reference Tongway and Ludwig1994, Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997; Tongway and Hindley, Reference Tongway and Hindley2004) and in restoration ecology (Tongway and Ludwig, Reference Tongway and Ludwig2011). Walt also opened up whole chapters of research with other colleagues, such as “bioturbation studies” involving organisms smaller and larger than termites that “turn over” soil and facilitate seed germination (Noble et al., Reference Noble, Müller, Whitford and Pfitzner2009; Eldridge et al., Reference Eldridge, Koen, Killgore, Haung and Whitford2012). These studies demonstrated that termites serve a crucial role in rangeland functioning, an understanding that would not have occurred within the original “precision agriculture” methods of study, where the farmer takes responsibility for the physics, chemistry and biology of the soil.
Walt’s ability to recognize and identify fine-scale, biota-mediated processes and to integrate the role of these processes at the local watershed scale expanded our capacity to be more “complete” ecologists. Walt demonstrated that no matter how cryptic or notionally irrelevant termites might initially appear, there was always a functional, process-based explanation for their presence in the ecosystem. Walt was not satisfied with just lists of “those present” or “fancy that, what’s it doing here?.” Walt strived to demonstrate their role in biophysical processes, that is, not simply compile a list of what happens, but sequences of how it actually happens – a functional integration. Walt placed more importance on functional diversity than on simple biodiversity assessments, which are often incomplete or neglect cryptic organisms; this is where we need to be as scientists if we are to inform our emerging successors.
Walt had a strong mentoring or pedagogic streak, which enthused those new to ecology to rapidly uptake new information. Walt’s contributions to the importance of fine-scaled biological processes in ecosystems went well beyond those with us in Australia. To note a few of many, Walt promoted the importance of assessing soil condition in other rangelands, such as the drylands of the western United States (Whitford et al., Reference Whitford, De Soyza, Van Zee, Herrick and Havstad1998; DeSoyza et al., Reference DeSoyza, Whitford, Turner, Van Zee and Johnson2000). Walt encouraged colleagues to study the role of microbes, lichens and fungi in drylands, notably their role in the ecohydrology of dryland watersheds (Belnap et al., Reference Belnap, Welter, Grimm, Barger and Ludwig2005; Ludwig et al., Reference Ludwig, Wilcox, Breshears, Tongway and Imeson2005).
Before we close, we would be remiss if we did not say that besides Walt’s enduring collaborations with CSIRO colleagues, Walt also established long and productive collaborations with other Australian ecologists, such as David Eldridge (e.g., Eldridge and Whitford, Reference Eldridge and Whitford2014). We also want to emphasize Walt’s collaboration with our CSIRO colleague Jim Noble, which occurred during 1986–1987, and again in 1990. We quote from Walt’s memoir: “Another fun collaboration was with a rangeland ecologist, Jim Noble, who had a farm just north of Deniliquin. He invited me to do a field trip with him to his research sites in Western New South Wales and then to the Lake Mere field station where I worked with Ludwig and Tongway. Traveling with Jim was a great education because of his broad interests in the history of the country and his knowledge about indigenous peoples.” After 1990, Walt’s research on Australian termites continued in collaboration with Jim and others (e.g., Noble et al., Reference Noble, Whitford and Kaliszweski1996, Reference Noble, Müller, Whitford and Pfitzner2009).
A closing perspective
We close our perspective on Walt’s experiences in Australia with another quote from Walt’s memoir. Walt wrote: “Spending time in the field with David [Tongway] and John Ludwig was a great education because John knew Australian plant communities; David understood how the Australian arid landscapes worked; I added a perspective on animals, especially unseen termites that are important to how the arid landscapes function. We had very productive collaboration that resulted in several publications and I left Australia with a greater appreciation for linkages across landscapes and the complexity of arid ecosystems.” As true for many others, we had a very positive synergy with Walt.
Open peer review
For open peer review materials, please visit https://doi.org/10.1017/dry.2026.10034.
Data availability statement
As noted within the two case studies, additional data on the results presented are available on mulga log mounds in Tongway et al. (Reference Tongway, Ludwig and Whitford1989) and Tongway and Ludwig (Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997), and on mulga tree groves in Tongway and Ludwig (Reference Tongway and Ludwig1990, Reference Tongway, Ludwig, Ludwig, Tongway, Freudenberger, Noble and Hodgkinson1997).
Acknowledgments
The authors would like to thank CSIRO, the research organization that provided us with long and productive careers, with the opportunity to work with many colleagues and with guests such as Walt Whitford.




 Open access
Open access
Comments
Dear David and Graham,
Thank you for the invite to submit a manuscript for consideration for a special issue of Drylands, a tribute to Walt Whitford. We aimed to provide a personal perspective based on research we did with Walt in the semi-arid woodlands of southeastern Australia in 1986-87. As described in two previously published case studies with Walt, we illustrate how Walt demonstrated to us that termite activities enhanced nutrient and infiltration processes in fine-scale patches, and we demonstrated to Walt how these patch processes scaled up to local landscapes—we learned together—a synergy.
We closed with the perspective that Walt’s legacy of mentoring 40 years later not only contributed to our professional development but to that of many others around the world.
We affirm that we both contributed to the concept of this perspective manuscript, and to its drafting and revisions. We both approved the final version of the submitted manuscript.
We also affirm that we have no conflicts of interest with regards to this manuscript and that this research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Cheers,
John and David
PS: David is currently not shown as Author 2 in the submission processes because he doe not have an ORCID ID. I am not sure how to fix this because ORCID does not seem to like personal email addresses—it wants email addresses indicating an institutional affiliation. John