


Introduction
In the US, sows are typically housed in stalls during gestation. The favourability of stalls includes that individual housing provides a safe environment for both the sows and employees, protecting sows from injury and physical aggression from other sows, and enabling each female to receive an individual feed ration without competition (Salak-Johnson Reference Salak-Johnson2017). Additionally, stall housing is economically efficient, allowing more sows to be housed in fewer square feet of space, resulting in improved economies of scale compared to group-housing facilities (Buhr Reference Buhr2010).
However, stall housing has been criticised (Ryan et al. Reference Ryan, Fraser and Weary2015) for limiting sows’ movement and normal behaviours, thereby reducing sow welfare in comparison to group-housed females. The ability of the animal to perform normal behaviours that they are motivated to perform is important and will impact their affective state, and definitions of welfare now consider this (Englund & Cronin Reference Englund and Cronin2023). The ability of an animal to perform behaviours and have choice in their environment is linked to a higher welfare status in many species (Englund & Cronin Reference Englund and Cronin2023). A sow’s choice regarding where to walk or lie, or whether to interact with other pigs (Sus scrofa) is restricted in stall housing, but group housing provides broader options. Pork consumers have demanded higher welfare standards for gestating sows and see group housing as promoting improved sow welfare, resulting in an international trend for banning gestation stalls (European Commission 2008; Canadian Pork Council and National Farm Animal Care Council 2014; Schulz & Tonso Reference Schulz and Tonsor2015). This has also been seen in the US, most recently with the passing of Proposition 12 in California, which requires 2.23 m2 of floor space per sow in gestation (Proposition 12 – Prevention of Cruelty to Farm Animals Act 2018). However, the research conclusions are conflicting as to whether sows housed in stalls experience poorer welfare than sows in group housing, as measures of physical health and affective state must both be considered. Measures of reproductive performance such as farrowing rate and litter size have been reported to be both negatively and positively influenced or not impacted by stall housing, as reviewed by Koketsu and Iida (Reference Koketsu and Iida2017). Stereotypical behaviours are considered indicators of chronically poor welfare because they develop as symptoms of frustration and boredom and impoverished housing conditions, and they have been observed with greater frequency in stall-housed sows (Chapinal et al. Reference Chapinal, Ruiz De La Torre, Cerisuelo, Gasa, Baucells, Coma, Vidal and Manteca2010). However, other measures of sow welfare like frequency and severity of lameness have been shown to be worse in group-housed sows (Koketsu & Iida Reference Koketsu and Iida2017). Biological metrics such as cortisol in blood and saliva have been used to assess stress related to housing systems (Barnett et al. Reference Barnett, Winfield, Cronin, Hemsworth and Dewar1985; Anil et al. Reference Anil, Anil, Deen and Baidoo2006), however, blood and salivary cortisol are measures of acute stress, as cortisol is secreted as a response of the hypothalamic-pituitary-adrenal axis to a stressor, following the secretion of adrenocorticotropic hormone (Heimbürge et al. Reference Heimbürge, Kanitz, Tuchscherer and Otten2020b). Cortisol is secreted into the bloodstream and incorporated into saliva within minutes after a stressor, such as restraint or handling during a blood draw (Bozovic et al. Reference Bozovic, Racic and Ivkovic2013); blood cortisol has been observed in swine to return to pre-stressor levels within an hour of an acute stress event (Guzik et al. Reference Guzik, Matthews, Kerr, Bidner and Southern2006). Cortisol in blood and saliva is therefore impractical as a measure of chronic stress, and measures that reflect long-term cortisol secretion may be more accurate in characterising chronic stress and as an indicator of sow welfare. Recently, there has been increasing interest in cortisol deposited in hair as a measure of long-term stress; with one theory being that cortisol circulating in the bloodstream is incorporated into the hair slowly as it grows capturing the longer-term activity of the HPA axis (Heimbürge et al. Reference Heimbürge, Kanitz, Tuchscherer and Otten2020b). Studies in multiple species have examined hair cortisol concentration (HCC) as an indicator of chronic activation of the hypothalamic-pituitary-adrenal axis and have observed elevated HCC in hypercortisolaemic subjects (Russell et al. Reference Russell, Koren, Rieder and Van Uum2012), infants on ventilators compared to healthy infants (Yamada et al. Reference Yamada, Stevens, De Silva, Gibbins, Beyene, Taddio, Newman and Koren2007), and animals such as dairy cattle (Bos taurus; González-de-la-Vara et al. Reference del González-de-la-Vara, Valdez, Lemus-Ramirez, Vázquez-Chagoyán, Villa-Godoy and Romano2011), goats (Capra hircus; Endo et al. Reference Endo, Yamane, Rahayu and Tanaka2018), and lynx (Lynx canadensis; Terwissen et al. Reference Terwissen, Mastromonaco and Murray2013) undergoing simulated stress conditions via injection of exogenous adrenocorticotropic hormone. Elevated hair cortisol has also been observed in swine experiencing chronic stress conditions (Prims et al. Reference Prims, Vanden Hole, Van Cruchten, Van Ginneken, Van Ostade and Casteleyn2019; Morgan et al. Reference Morgan, Meyer, Novak, Younis, Ahmad and Raz2021). Cortisol in hair may be used to identify chronic stress and hence provide information on sow welfare by acting as a record of elevated cortisol over a prolonged period.
In previous research, hair cortisol concentration in group-housed sows was not influenced by parity, but litter size and HCC were positively correlated (Roelofs et al. Reference Roelofs, Godding, De Haan, Van Der Staay and Nordquist2019); it has also been noted that HCC fluctuates during the reproductive cycle of sows housed both individually and in groups (Bacci et al. Reference Bacci, Nannoni, Govoni, Scorrano, Zannoni, Forni, Martelli and Sardi2014). However, little research to date has directly compared HCC of gestating sows kept in individual housing versus those housed in groups. The purpose of this study was to determine the HCC of sows in stall- and group-gestation housing systems within the same facility and to compare whether they differed. Results are to contribute knowledge on HCC as a metric of chronic stress influenced by housing system during gestation and to assess the influence of housing system on sow welfare.
Materials and methods
Ethical approval
The experimental protocols used in this study were approved by The South Dakota State University Institutional Animal Care and Use Committee (IACUC #17-072A and 18-064A).
Study animals and experimental design
The study was conducted in the sow barn at the South Dakota State University Swine Education and Research Facility from November 2018 until June 2019. A total of 123 multiparous and primiparous females (PIC 1050) across four breeding groups and housed in gestation stalls (two breeding groups) or group pens (two breeding groups) were used in this study. The sow herd was managed in a batch farrowing system where 20–30 sows per breeding group were bred, farrowed, and/or weaned every four weeks. The east half of the gestation facility contained 76 individual gestation stalls and the west half three group pens. The gestation facility was artificially lit between 0600 and 1400h and mechanically ventilated, and there was no wall separating the group pens from the stalls. The group pens (8.53 × 8.20 m; length × width) were slatted concrete, with a solid concrete portion along the west wall (8.20 × 3.18 m; length × width). The solid portion of each group pen had two partitions, dividing the solid portion into three resting areas (3.18 × 2.73 m; length × width). There were two water nipples along the south side of each pen, and a Mannabek electronic sow feeder (PigTek, Milford, IN, USA) on the east side of each pen. The stall-housed sows were housed in fully slatted gestation stalls (1.98 × 0.61 m; length × width) from breeding until day 111 (± 1.4) of gestation. Each stall had an individual feeder and a water nipple. Rations were delivered once per day and water was accessible ad libitum. The first stall-housed breeding group was bred in November 2018, and the second in December 2018. The first pen-housed breeding group was bred in February 2019, and the second in March 2019. The group-housed sows were divided among three group pens approximately 48 h after breeding. The group housing was managed as a dynamic system with females being added at approximate 28-day intervals depending upon housing needs, illustrated in Figure 1. Over the course of this work, gilts and small parity 1 sows from both pen-housed breeding groups were housed in one group pen (e.g. when gilts and small parity 1 sows from the first group reached approximately day 28 of gestation, the second breeding group of gilts and small parity 1 sows was added to the same pen). Multiparous sows were housed in the other two group pens. Each group pen housed up to 25 sows, providing at least 2.79 m2 per female, and as noted above, over the course of this work the group pens were dynamic due to the introduction or removal of non-experimental herd sows as outlined in Figure 1. During the gilt development period, all gilts were housed in groups of eight females per pen until undergoing acclimation to stalls four weeks prior to first breeding. All multiparous females were housed in stalls at least five days prior to breeding. Sows were moved from their assigned housing system to farrowing crates at day 111 (± 1.1) of gestation.
Entry and exit of breeding groups through the pen housing system in gestation.
1 Non-experimental breed groups entered pen after pregnancy check at approximately day 28 of gestation; 2 Experimental breed groups entered pen at day 2 after breeding.

All females were included in one of two nutrition trials being conducted during this experiment. All stall-housed sows were in a trial examining dietary antioxidants fed in gestation and lactation (Hernandez et al. Reference Hernandez, Dunn, Wenninghoff, Hesse and Levesque2021). There were four treatments, receiving a standard gestation diet plus a carrier top dress including either peppermint oil, whole yeast cell, gamma-tocopherol, or nothing (control). All group-housed sows were in a trial examining varying dietary lysine:energy ratios in gestation (Bruhn Reference Bruhn2020). There were three treatments: constant lysine:energy ratio throughout gestation; higher lysine:energy ratio from day 90–110 of gestation (phase-feed 1) or lower lysine:energy ratio from day 2–89; and higher:energy ratio from day 90–110 of gestation (phase-feed 2). In both nutrition trials, gestation diets were formulated to meet or exceed gestation nutrient requirement estimates according to National Research Council (2012). Daily feed allotment was managed similarly across both housing systems (i.e. daily feed allotment adjusted at 30-day intervals throughout gestation to maintain body condition score of 3); thus, differences in diets between the trials were not expected to impact sow response to housing. Further details regarding the nutritional studies can be found in Hernandez et al. (Reference Hernandez, Dunn, Wenninghoff, Hesse and Levesque2021) and Bruhn (Reference Bruhn2020).
Hair sample collection
If not already housed in a stall, animals were moved one-by-one to an individual stall to facilitate collection of hair; animals were replaced into their original group pen immediately following hair collection, which was completed within 5 min for each animal. An electric trimmer (Remington HC5855B, Middleton, USA) was fitted with a flexible plastic shield to better facilitate the direct movement of trimmed hair into the paper envelope, and hair was shaved as close to the skin as possible. The trimmer was wiped following each individual procedure to remove any hair fragments. Approximately 400 cm2 on the right hip was shaved at 6.0 (± 3.8) days post-breeding and hair was discarded. Hair regrowth samples (150 mg) from the hip at day 37, day 74, and day 111 of gestation were collected. All samples were collected and stored in individual paper envelopes and kept in a dark drawer in the laboratory at 20oC until analysis (not more than seven months).
Laboratory analysis
Samples were processed in accordance with a modified version of the MacBeth et al. (Reference Macbeth, Cattet, Stenhouse, Gibeau and Janz2010) protocol. Samples were examined, hairs with disturbed cuticles or broken hairs were discarded, split ends were trimmed off and the remaining intact portion of the hair was retained, debris removed, and samples were weighed. Samples ≤ 50 mg were placed into 2-mL cryogenic vials (Corning #430659, NY, USA) and > 50 mg were placed into 5-mL disposable glass culture tubes (Fisher Scientific #14-961-26, MA, USA). The samples were washed three times in methanol (40 μL mg–1 sample), blotted dry between rinses, then left to dry overnight. Samples were ground into a fine powder with a Retsch MM310 mixer mill (Retsch, Haan, Germany) at 30 Hz; 25 mg of powdered hair were weighed into 0.6-mL tubes. The analyte was extracted using 0.5 mL methanol per 25 mg of powdered hair, and the samples incubated for 16–24 h on a rotator. Tubes were centrifuged at 20°C for 15 min at 2,150 × g, and the supernatant collected and transferred to a 12-mm glass culture tube. To ensure all extracted steroids were recovered, the powdered hair sample was rinsed twice by adding 0.5 mL of fresh methanol, then gently vortexing (40 s), centrifuging, and pooling supernatants. The pooled supernatant was dried at 38°C under a gentle stream of nitrogen gas and reconstituted with 200 μL of extraction buffer from the cortisol assay kit, and samples were frozen at –80°C until analysis with enzyme immunoassay (EA65, Oxford Biomedical, Rochester Hills, MI, USA). The optical density value was read at 450 nm after 30 min on a SpectraMax 190 absorbance plate reader (Molecular Devices, Sunnyvale, CA, USA). A standard curve of optical density value versus cortisol concentration was generated, and hair cortisol concentration was then determined according to the standard curve and expressed in pg cortisol mg–1 powdered hair. According to the manufacturer, cross-reactivity of the antibody used for the cortisol kit is as follows: cortisol (100.00%), prednisolone (66.9%), 11-deoxycortisol (58.1%), cortisone (15.9%), prednisone (13.7%), 17-hydroxyprogesterone (5.4%), dexamethasone (4.6%), oestriol 4.5%, oestrone (4.1%), d-aldosterone (3.6%), progesterone (3.5%) 6-β-hydroxycortisol (3.4%), trans dehydroandrosterone (1.9%), testosterone (1.7%), corticosterone (1.4%), and pregnenolone (1.3%).
Sample selection for analysis
All available stall- (n = 64) and group-housed sows (n = 59) were shaved and samples collected 6.0 (± 3.8) days post-breeding. At day 111, 50 stall- and 39 group-housed sows remained, hair was shaved, and samples collected. Overall, 14 stall- and 20 group-housed females were removed from the trial due to failure to become or remain pregnant or removed from group pen due to injury or illness, or insufficient hair growth. During the laboratory selection and analysis, a further 16 samples from stall-housed sows and seven from group-housed females were removed due to inadequate sample size or poor sample quality (badly damaged or soiled). In total, samples from 34 stall-housed sows and 32 group-housed sows were analysed for HCC and included in the statistical analysis. Hair regrowth samples from the hip at day 37, day 74, and day 111 of gestation were collected with only samples from days 37 and 111 analysed for cortisol concentration. These time-points were selected because day 37 captured the period when sows were introduced to groups, and day 111 captured the final stage of gestation. Unless otherwise noted, results are expressed as an average of day 37 and day 111 cortisol concentrations. Samples from day 74 were not analysed in this experiment due to budget constraints. The final analysis contains the following number of samples from stall-housed sows from each dietary treatment: peppermint oil (n = 1); whole yeast cell (n = 11); gamma-tocopherol (n = 9); and control (n = 13). The final analysis contains the following number of samples from group-housed sows from each treatment: control (n = 12); phase-feed 1 (n = 10); and phase-feed 2 (n = 10).
Statistical analysis
The UNIVARIATE procedure of SAS (Version 9.4, SAS Institute Inc, Cary, NC, USA) was used to review distribution of the data, test for normality, and identify outliers. Data were analysed using the MIXED procedure of SAS with ‘sow’ as the experimental unit and ‘HCC’ as the dependent variable. Sows were assigned to one of three parity classes, 0–1 (gilts and sows having completed one successful parity) (n = 23), 2–3 (n = 17), and ≥ 4 (n = 26) and to one of three litter size classes based on total born, ≤ 14 (n = 22), 15–17 (n = 20), and ≥ 18 (n = 24). There were potential confounding effects of dietary treatment, breed group, parity class, and litter size; therefore, the main effect of each was tested. Main effect of dietary treatment and breed group were analysed for each housing system separately. Where the effect of a confounding variable was significant, the interaction with housing system was tested. Non-significant interactions were removed from the model. The final model for housing system and day of gestation included the main effects of housing system, day of gestation, and their interactions were tested with parity class as a random effect. ‘Pen’ was not included in the model either as confounding or random variable because of the difference in absolute number of pens between housing system (i.e. each sow equalled a ‘pen’ for stall-housed females) and because pen was synonymous with group as outlined in Figure 1. Data are presented as least significant means and standard error. Tukey’s adjusted means test was used to detect differences where statistical significance and tendencies were set at P ≤ 0.05 and P < 0.10, respectively, for all statistical tests.
Results
Dietary treatment did not influence HCC in that the HCC of control-fed females did not differ from the HCC of experimentally fed females in either the stall-housed or group-housed females (P > 0.1). Summary statistics of the sow groups used in this study are shown in Table 1.
Summary statistics regarding the specific groups used in this study into hair cortisol in gestating sows.

SD: Standard deviation.
There was no difference in HCC between breed groups in stall-housed females (66.5 pg mg–1 for the first breed group and 46.4 pg mg–1 for the second breed group; SEM = 13.3) and a tendency for group-housed females in the second breed group to have higher HCC (18.4 pg mg–1 for the first breed group and 23.7 pg mg–1 for the second breed group; SEM = 2.9; P = 0.0752).
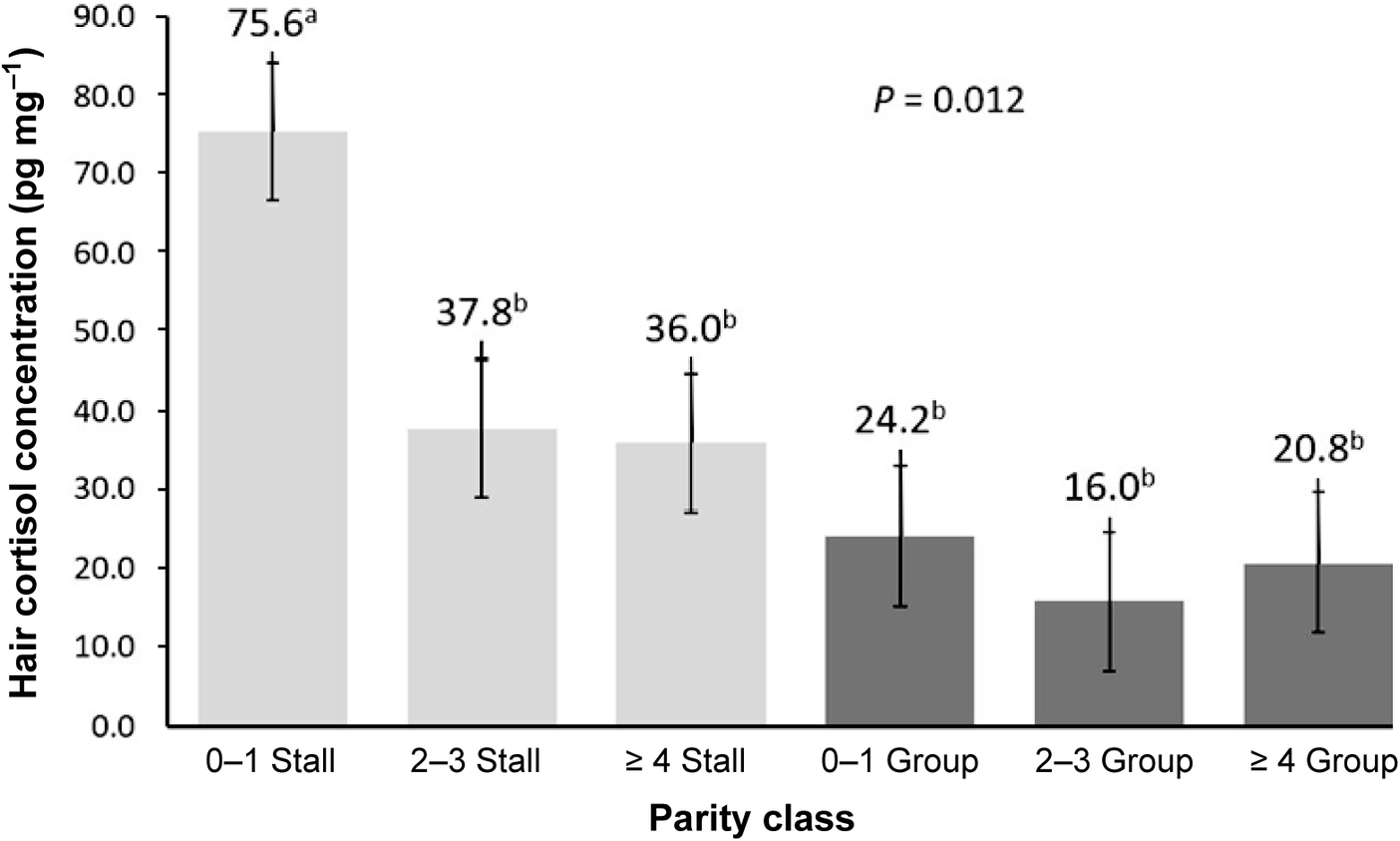
There was a significant interaction between housing system and parity class on HCC; parity 0–1 females housed in stalls had higher HCC than older sows housed in stalls (parity ≥ 2) and there was no difference among parity class for females housed in group pens (Figure 2).
Interaction between parity class1 and housing system (stall- and group-housed females) as regards hair cortisol concentration2,3. 1Parity 0–1 refers to gilts and sows that have successfully completed one parity, 2–3 refers to sows successfully completing 2 or 3 parities, and ≥ 4 refers to sows successfully completing 4 or more parities. 2 Bars with different superscript a, b, c denote significant differences where P ≤ 0.05. 3 0–1 Stall (n = 11), 2–3 Stall (n = 10), ≥ 4 Stall (n = 13); 0–1 Group (n = 12), 2–3 Group (n = 7), ≥ 4 Group (n = 13). Light grey represents stall-housed females and dark grey represents group-housed females.

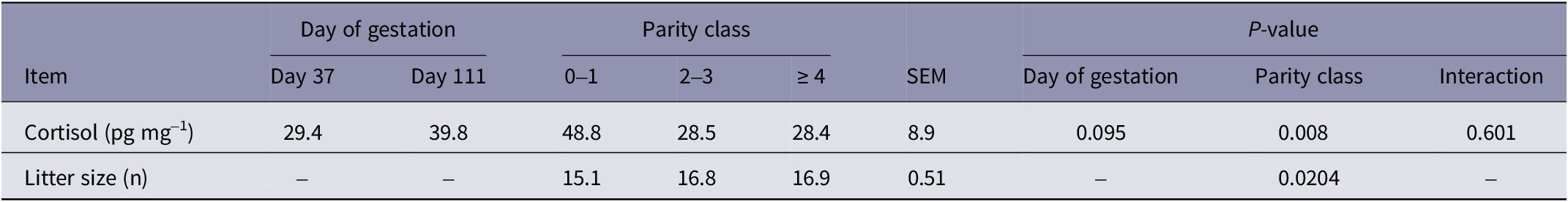
There was a tendency for day of gestation to influence HCC, with an increase in HCC at day 111 of gestation (Table 2). The HCC of stall-housed females increased 39% from day 37 to day 111 of gestation (41.3 pg mg–1 to 57.5 pg mg–1; SEM = 8.8) and 26% in group-housed sows (17.5 pg mg–1 to 22.1 pg mg–1; SEM = 9.0), but there was no interaction between day of gestation and housing system.
Hair cortisol concentration in gestating sows by day of gestation 1 and parity class 2; and average litter size 3 by parity class

1 Hair cortisol concentration was measured in hair growth between day 0 (breeding) and day 37 of gestation, and growth between day 74 and day 111 of gestation, representing the first and last third of gestation, respectively
2 Parity 0–1 refers to gilts and sows that have successfully completed one parity, 2–3 refers to sows successfully completing 2 or 3 parities, and ≥ 4 refers to sows successfully completing 4 or more parities
3 Litter size refers to total piglets born per litter
SEM: Standard error of the mean.
Litter size was affected by parity class, where parity ≥ 4 females had more total piglets born than parity 0–1, and parity 2–3 did not differ significantly from parity ≥ 4 (Table 2). However, litter size did not affect overall HCC (P = 0.212).
Discussion
Elevated HCC has been observed in animals undergoing simulated chronic stress conditions via injection of exogenous adrenocorticotropic hormone (González-de-la-Vara et al. 2011; Terwissen et al. Reference Terwissen, Mastromonaco and Murray2013; Endo et al. Reference Endo, Yamane, Rahayu and Tanaka2018). Further, elevated HCC has been detected in multiple species (humans [Iob & Steptoe Reference Iob and Steptoe2019], non-human primates [Davenport et al. Reference Davenport, Lutz, Tiefenbacher, Novak and Meyer2008; Fairbanks et al. Reference Fairbanks, Jorgensen, Bailey, Breidenthal, Grzywa and Laudenslager2011; Dettmer et al. Reference Dettmer, Novak, Suomi and Meyer2012; Carlitz et al. Reference Carlitz, Kirschbaum, Stalder and Van Schaik2014], wolves [Canis lupus; Bryan et al. Reference Bryan, Smits, Koren, Paquet, Wynne-Edwards and Musiani2015], grizzly bears [Ursus arctos horribilis; Bryan et al. Reference Bryan, Darimont, Paquet, Wynne-Edwards and Smits2013], sheep [Ovis aries; Nejad et al. Reference Nejad, Lohakare, Son, Kwon, West and Sung2014], and pigs [Prims et al. Reference Prims, Vanden Hole, Van Cruchten, Van Ginneken, Van Ostade and Casteleyn2019; Heimbürge et al. Reference Heimbürge, Kanitz, Tuchscherer and Otten2020b; Morgan et al. Reference Morgan, Meyer, Novak, Younis, Ahmad and Raz2021]) experiencing chronic stress conditions, either induced experimentally or naturally occurring. This suggests that in the present study, the parity 0–1 stall-housed females may have experienced greater chronic stress than all other group-housed and stall-housed females, suggesting stall housing reduced welfare of gilts and parity 1 sows, whereas group-housing did not negatively affect the welfare of pregnant females regardless of parity. However, because older sows in stall housing had indistinguishable HCC compared to older group-housed sows, acclimation to stall housing may occur over the course of multiple parities. Stall-housed females are observed as exhibiting more frequent stress behaviours, such as sham chewing and oronasofacial stereotypies, compared to females in group pens (Chapinal et al. Reference Chapinal, Ruiz De La Torre, Cerisuelo, Gasa, Baucells, Coma, Vidal and Manteca2010). Sows in group pens also can experience stress from exposure to fighting for social hierarchy, aggression, and injury from other sows (Salak-Johnson Reference Salak-Johnson2017). Other research has shown that frequent, repeated mixing of unfamiliar piglets over the course of three weeks does increase HCC (Prims et al. Reference Prims, Vanden Hole, Van Cruchten, Van Ginneken, Van Ostade and Casteleyn2019). In this trial, behavioural measures and injury scores were not recorded, so it is not possible to evaluate the acute stress experienced by the group-housed sows during mixing. Group-housed females were kept in dynamic groups, and it is probable therefore they experienced acute stress during mixing and for assembling order on the electronic sow feeder. Nevertheless, this period of acute stress at mixing was not sufficiently stressful or prolonged to raise the HCC of group-housed females above the HCC of their stall-housed counterparts. The HCC of group-housed sows was lower than stall-housed females younger than parity 2 despite the presumption acute stress at mixing in group-housing systems increases overall sow stress. This suggests that the stress of confinement parity 0–1 stall-housed females experienced over the course of gestation was greater and persistent and led to chronically elevated cortisol. Adaptation, defined as lower prolonged stress, to stall housing may not occur until the third or fourth pregnancy.
The lack of difference in HCC between parity classes in group housing demonstrates that parity 0–1 females in group housing do not experience significantly higher stress than their older and more experienced counterparts. The experience of being bred for the first time and carrying a litter does not appear to cause significantly greater stress to a naïve female compared to multiparous females when housed in group pens. This result is consistent with Roelofs et al. (Reference Roelofs, Godding, De Haan, Van Der Staay and Nordquist2019) who, similarly, did not detect a difference in HCC between multiparous and primiparous females in group housing.
It is generally recommended to acclimate gilts to stall housing for two to three weeks prior to boar exposure (Kraeling & Webel Reference Kraeling and Webel2015), as pigs are known to be social animals (Barnett et al. Reference Barnett, Winfield, Cronin, Hemsworth and Dewar1985) and individual housing can induce stress, which can disrupt oestrus, prolong the oestradiol peak, and decrease early embryo survival (Roongsitthichai et al. Reference Roongsitthichai, Suwimonteerabutr, Kaeoket, Koonjaenak and Tummaruk2011). Gilts in this study were acclimated to stall housing four weeks prior to first breeding. Despite this acclimation period, the results of this study indicate that gilts and parity 1 sows housed in stalls throughout gestation experienced higher stress than group-housed gilts and parity 1 sows and therefore were not adequately acclimated to stall confinement. This may be because of the restricted ability to choose whether to interact with other pigs and where to lie or walk: this freedom of choice is associated with improved welfare status (Englund & Cronin Reference Englund and Cronin2023).
An increase in HCC with progressing pregnancy is expected, as both fetal and maternal cortisol rise as parturition approaches. Fetal cortisol increases as gestation progresses (Silver & Fowden Reference Silver and Fowden1989) and in many mammals plays a role in inducing parturition (Decaluwe et al. Reference Decaluwe, Janssens, Declerck, De Kruif and Maes2012; Wood Reference Wood2013). Cortisol is critical for late fetal development, assisting in lung and kidney maturation (Challis et al. Reference Challis, Matthews, Gibb and Lye2000), skeletal growth in sheep (Fowden et al. Reference Fowden, Apatu and Silver1995), gastrointestinal development, and the transition from trans-placental glucose supply to liver glycogen and gluconeogenesis (Fowden et al. Reference Fowden, Apatu and Silver1995; Fishman et al. Reference Fishman, Vortman, Shanas and Koren2018). Maternal cortisol also contributes to fetal growth by promoting maternal insulin resistance (Herrera Reference Herrera2000), which reduces the female’s ability to utilise glucose from the bloodstream, thereby increasing glucose availability for the litter. As glucose is the primary nutrient crossing the placenta, this promotes fetal growth in late gestation (Herrera Reference Herrera2000). The rise in cortisol in late gestation in stall-housed females may also be related to the increase in discomfort due to progressing pregnancy and space restriction as reported by Anil et al. (Reference Anil, Anil, Deen and Baidoo2006); however, there are no data to our knowledge similarly associating late gestation HCC increase to discomfort in group-housed females.
There may be consequences regarding chronically heightened cortisol in all stages of gestation for litter productivity and piglet welfare. Elevated maternal glucocorticoids during gestation have been linked with lower offspring birth weights and weaning weights (Kranendonk et al. Reference Kranendonk, Hopster, Fillerup, Ekkel, Mulder, Wiegant and Taverne2006), and lower lactation and nursery performance (Jarvis et al. Reference Jarvis, Moinard, Robson, Baxter, Ormandy, Douglas, Seckl, Russell and Lawrence2006; Otten et al. Reference Otten, Kanitz and Tuchscherer2015). Piglets from sows experiencing elevated glucocorticoids during gestation may be more likely to have poorer welfare, demonstrating more anxiety-related behaviour or aggression in novel situations and higher pain score during tail docking (Otten et al. Reference Otten, Kanitz and Tuchscherer2015). In addition, gilts raised from mothers who had experienced stress during gestation appeared more restless and uncomfortable in the periparturient period (Jarvis et al. Reference Jarvis, Moinard, Robson, Baxter, Ormandy, Douglas, Seckl, Russell and Lawrence2006; Rutherford et al. Reference Rutherford, Piastowska-Ciesielska, Donald, Robson, Ison, Jarvis, Brunton, Russell and Lawrence2014), and in one trial tended to be more aggressive towards piglets (Jarvis et al. Reference Jarvis, Moinard, Robson, Baxter, Ormandy, Douglas, Seckl, Russell and Lawrence2006); both frequent posture changes and sow aggression can result in newborn piglets being injured or killed, contributing to pre-weaning mortality. Therefore, chronic stress in sows can detrimentally affect the productivity and welfare of their offspring. Our results suggest that parity 0–1 sows housed in stalls are more likely to experience chronic stress than group-housed females and may therefore produce less economically productive offspring with lower welfare status. In this trial, piglet birth weights and mortality were not evaluated, so there is a need for future research to consider the productivity and welfare of piglets born from stall- and group-housed sows.
Study limitations
Research into sow housing can be challenging due to the statistical limitations of comparing individual sows (such as those in stalls) to groups of sows. The choice was made in this research to use individual ‘sow’ as experimental unit rather than ‘pen’: with respect to the primary variable of interest (hair cortisol), the individual animal is the most appropriate experimental unit. The ‘stress’ associated with a given group-housing pen is a dynamic interaction between individual animal characteristics (i.e. age, size, dominance) (Li et al. Reference Li, Wang and Johnston2012; Wang & Li Reference Wang and Li2016) and cannot be suitably represented with an average value for all females within a pen. There is a precedent for this choice of statistical analysis in sow housing research, when using ‘pen’ as experimental unit is impractical (Chapinal et al. Reference Chapinal, Ruiz De La Torre, Cerisuelo, Gasa, Baucells, Coma, Vidal and Manteca2010; Bacci et al. Reference Bacci, Nannoni, Govoni, Scorrano, Zannoni, Forni, Martelli and Sardi2014; Zhou et al. Reference Zhou, Sun, Wang, Zhou, Lu, Marchant-Forde and Zhao2014; Min et al. Reference Min, Choi, Kim, Kim, Jeong, Kim, Song and Jung2020; Liu et al. Reference Liu, Song, Yan, Zhang, Wang, Zhao, Gao, Hou, Shi, Li and Wang2021; Tokareva et al. Reference Tokareva, Brown, MacPhee, Janz and Seddon2022).
In this study, the floor space allowance and group size in the group pens is not analogous to what is commonly seen in modern commercial sow facilities. Proposition 12 – Prevention of Cruelty to Farm Animals Act (2018) requires 2.23 m2 per sow, the EU requires 2.25 m2, and Canada requires at least 1.8 m2; and group sizes can be upwards of 50 sows per pen in group housing systems currently in use in the US (Li et al. Reference Li, Cui, Yang, Johnston and Baidoo2018). In this study, floor space per sow was, on average, 2.79 m2 and group pens held up to 25 sows. Therefore, the apparent lower stress of parity 0–1 sows in group housing compared to their stall-housed counterparts may, here, be related to large space allowance. Further investigation into the influence of stocking density and large group sizes on HCC in group-housed gestating sows is needed.
Despite differences in diets between the group- and stall-housed females, the lack of diet effect indicates gestation diet had limited impact on HCC. Further, total macro- and micro-nutrient supply for all females met or exceeded National Research Council (2012) requirement estimates and daily diet allocation was the same between all groups (i.e. allocated to maintain body condition score of 3) which means nutrient supply for fetal growth and metabolism of the pregnant females was similar across both groups.
In this study, possible seasonal influence on HCC was confounded with group. Due to farm management and research project requirements, stall-housed sows had to be bred in late fall and the group-housed sows bred in late winter; thus, having both housing types in each group was not possible. However, when the two stall-housed breed groups were compared to each other there was no difference in HCC; when group-housed breed groups were compared, there was a tendency for the second group to have a higher HCC (P = 0.0752). For the stall-housed sows, the average outside temperature ranged from 4–16°C, and the ambient barn temperature was maintained in the sows’ thermoneutral zone. During the last part of gestation for the group-housed females (May and June) the average outside temperature ranged from 22–27°C, so it is likely that the ambient barn temperature was at times above the sows’ thermoneutral zone. Previous research is inconclusive regarding the influence of season on HCC in swine. Heimbürge et al. (Reference Heimbürge, Kanitz, Tuchscherer and Otten2020a) failed to detect a difference in HCC between samples collected in July versus January, and Wiechers et al. (Reference Wiechers, Brunner, Herbrandt, Kemper and Fels2021) also failed to associate HCC with barn temperature. Bacci et al. (Reference Bacci, Nannoni, Govoni, Scorrano, Zannoni, Forni, Martelli and Sardi2014) found that HCC of sows was lower during summer months and higher barn temperatures. However, Escribano et al. (Reference Escribano, Contreras-Jodar, López-Arjona, Cerón, Fàbrega, Aymerich and Dalmau2023) observed elevated cortisone in the hair of pigs under heat stress conditions, and López-Arjona et al. (Reference López-Arjona, Tecles, Mateo, Contreras-Aguilar, Martínez-Miró, Cerón and Martínez-Subiela2020) found higher cortisone:cortisol ratios in the hair of gestating sows during summer months. In this trial, the group-housed sows in the second breed group may have experienced periods of heat stress, elevating cortisone and increasing HCC due to cross-reactivity in the assay. If hair grown in the summer has higher HCC, we expect in this trial that the group-housed females would have had higher HCC; however, that was not the case. There was little difference between HCC of stall-housed and group-housed females. Further, HCC was greatest in stall-housed gilts collected during cooler months, thus it is unlikely seasonality influenced HCC in this trial. In future research, consideration for and investigation into the possibility of seasonal effect on HCC is recommended, along with making use of highly precise analytical techniques, such as liquid chromatography mass spectometry, to distinguish and quantify cortisol and cortisone where heat stress could be a factor.
Animal welfare implications
Housing parity 0–1 sows in stalls during gestation promoted more severe and/or prolonged stress than the stress experienced by their group-housed counterparts. These results add to the body of data that demonstrate that group-housing gestating sows has the potential to improve their welfare by providing greater freedom of movement, freedom of choice, and opportunities for social interaction.
Conclusion
This study demonstrated that parity 0–1 females housed in stalls during gestation have higher HCC compared to group-housed sows, suggesting greater chronic stress. Further, stress associated with group sow housing may be more characteristic of acute stress around mixing rather than chronic. Finally, litter size did not influence HCC, and the impact of greater HCC on postnatal litter performance is unknown and should be explored in further work.
Acknowledgements
Financial support for this project was provided by USDA National Institute of Food and Agriculture (Hatch Project: SD00H666-18). We thank the University of Saskatchewan Western College of Veterinary Medicine for the training and resources involved in the laboratory processing of hair samples.
Competing interests
None.



 Open access
Open access