


1. Introduction
Trace fossils are evidence of animal–substrate interaction, and contrary to body fossils that can be transported, reworked and redeposited, they represent dominantly in situ fossils with great paleoecological and paleoenvironmental significance (Ekdale and Mason, Reference Ekdale and Mason1988; MacNaughton, Reference MacNaughton and Miller2007; Buatois and Mángano, Reference Buatois and Mángano2011, Reference Buatois and Mángano2018). However, ichnofossils can exhibit long stratigraphic ranges, with only a few examples of applications as biostratigraphic tools (e.g., Seilacher, Reference Seilacher, Crimes and Harper1970; Shahkarami et al. Reference Shahkarami, Mángano and Buatois2017; Verde et al. Reference Verde, Netto, Azurica, Lavina and Pasquo2022; Memória et al. Reference Memória, Netto and Sedorko2023).
Despite limitations, multi-proxy investigations using trace fossils has great potential for assessing the age of sedimentary successions. Indeed, recent investigations by Verde et al. (Reference Verde, Netto, Azurica, Lavina and Pasquo2022) applying ichnofossils, microfossil and geochronological data have reassessed the oldest evidence of trace fossils from Uruguay (Pecoits et al. Reference Pecoits, Aubet, Heaman, Philippot, Rosière, Veroslavsky and Konhauser2016). These authors convincingly demonstrated that trace fossils previously recording Ediacaran age were repositioned into the Late Paleozoic Ice Age (LPIA) (Frakes and Crowell, Reference Frakes and Crowell1969; Deynoux et al. Reference Deynoux, Miller, Domack, Eyles, Fairchild and Young1994; Verde et al. Reference Verde, Netto, Azurica, Lavina and Pasquo2022). This similarity between shallow-marine Ediacaran ichnofaunas and late Paleozoic freshwater ichnofaunas has been explained as resulting from comparable strategies during the initial exploitation of underutilized or vacant ecosystems, a phenomenon known as the ‘déjà vu effect’ (Buatois and Mángano, Reference Buatois and Mángano2011, Reference Buatois, Mángano, Mángano and Buatois2016).
During geological mapping of the Sanfranciscana Basin in the Ibiracatu (Minas Gerais) region, the Brazilian Geological Survey recognized a succession containing trace fossils of bilaterian organisms in Ediacaran–Cambrian rocks from the Bambuí Group, a unit associated with Cloudina fossils (Warren et al. Reference Warren, Quaglio, Riccomini, Simões, Poiré, Strikis, Anelli and Strikis2014) and treptichnids (Sanchez et al. Reference Sanchez, Uhlein and Fairchild2021) (Figure 1). However, the absence of detailed sedimentological, stratigraphic, paleontological and geochronological data raises doubts about the age and depositional environment of the trace fossil-bearing succession, consisting potentially of a déjà vu effect.
Location and geological representation of the Sanfranciscana Basin in the northern portion of the state of Minas Gerais. (A) São Francisco Craton geological map (modified from Alkmim and Martins-Neto, Reference Alkmim and Martins-Neto2012; Caetano-Filho et al. Reference Caetano-Filho, Sansjofre, Ader, Paula-Santos, Guacaneme, Babinski, Bedoya-Rueda, Kuchenbecker, Reis and Trindade2021). (B) Geological map of the Sanfranciscana Basin in the study area. (C) Stratigraphic column of outcrops of the Bambuí Group in the study area. Legend: SFC (São Francisco Craton) Based on Romano et al. (Reference Romano, Knauer, da Costa, Joncew and Vasconcelos2015) and Uhlein et al. (Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019). C- Clay; St- Silt; S- Sand; G/P- Gravel/Pebble; M- Mudstone; W/P- Wackestone/Packstone; fG- fine Grainstone; mG- medium Grainstone. S1 to S9 refers to the columnar section raised in each corresponding outcrop. S2 is the outcrop with ichnofossils and is not represented in the stratigraphic column, as it will be discussed throughout this article.

Figure 1. Long description
The image presents a detailed geological map of the Sanfranciscana Basin located in the northern part of the state of Minas Gerais, Brazil. The map includes three main sections: (A) a geological map of the São Francisco Craton, (B) a geological map of the Sanfranciscana Basin in the study area, and (C) a stratigraphic column of outcrops of the Bambuí Group in the study area. The map highlights various geological formations, including the Serra da Saudade Formation, Lagoa do Jacaré Formation, and Serra de Santa Helena Formation. Key locations such as Ibiracatu, Varzelândia, Bonança, and São João da Ponte are marked with stars and squares. The stratigraphic column details different sedimentary layers, with labels indicating the presence of clay, silt, sand, gravel, pebble, mudstone, wackestone, packstone, and various grain sizes. The sections S1 to S9 correspond to different outcrops, with S2 being the outcrop containing ichnofossils, which will be discussed further in the article.
In order to verify the age and environmental significance of this trace fossil-bearing site and its relationship with the Bambuí Group, this study aims to (i) define the stratigraphic framework and depositional environments involved in the sedimentation of the sedimentary succession that outcrops in the Ibiracatu region; (ii) determine the maximum age of deposition, possible sources of sediments, as well as the paleoenvironmental conditions during deposition; and (iii) describe the preserved ichnodiversity and their paleoenvironmental implications.
2. Geological context
The Sanfranciscana Basin (Figure 1A,B) is an intracratonic basin developed over the São Francisco Craton and bordered by Brasiliano orogenic belts, including the Araçuaí Belt to the east, the Brasília Belt to the west and the Rio Preto Belt to the north, as well as Archean–Paleoproterozoic basement to the south and the Paramirim Aulacogen to the northeast (Alkmim and Martins-Neto Reference Alkmim, Martins-Neto, Pinto and Martins-Neto2001, Reference Alkmim and Martins-Neto2012; Reis et al. Reference Reis, Alkmim, Fonseca, Nascimento, Suss, Prevatti and UG2017). Its sedimentary fill records multiple tectono-sedimentary events and comprises three major Precambrian first-order sequences: Espinhaço (Paleo–Mesoproterozoic), Macaúbas (Neoproterozoic) and Bambuí (Ediacaran–Cambrian), overlain by Phanerozoic units, including the Permian–Carboniferous Santa Fé Group (Figure 2) (Campos and Dardenne, Reference Campos and Dardenne1997a; Sgarbi, Reference Sgarbi2000; Alkmim and Martins-Neto, Reference Alkmim and Martins-Neto2012; Reis and Suss, Reference Reis and Suss2016; Uhlein et al. Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019).
Simplified chronostratigraphic chart of the studied succession of São Francisco Basin. Modified from Campos and Dardenne (Reference Campos and Dardenne1997a) and Caetano-Filho et al. (Reference Caetano-Filho, Sansjofre, Ader, Paula-Santos, Guacaneme, Babinski, Bedoya-Rueda, Kuchenbecker, Reis and Trindade2021). Ages: ∼593 Ma (Paula-Santos et al. Reference Paula-Santos, Babinski, Kuchenbecker, Caetano-Filho, Trindade and Pedrosa-Soares2015), ∼520 Ma (Moreira et al. Reference Moreira, Uhlein, Dussin, Uhlein and Misuzaki2020), and ∼527 Ma (Tavares et al. Reference Tavares, Souza Martins, Alkmim and Lana2020).

Figure 2. Long description
The diagram illustrates the chronostratigraphic chart of the São Francisco Basin, divided into geochronology, chronostratigraphy, and lithostratigraphy sections. The geochronology section on the left lists periods from Ediacaran to Permian. The chronostratigraphy section in the middle shows layers of limestone, siltstone, and sandstone with specific ages marked as approximately 593 million years, 520 million years, and 527 million years. The lithostratigraphy section on the right categorizes these layers into groups and formations, such as Bambuí and Santa Fé, with specific formations like Três Marias, Serra da Saúde, and Tabuleiro.
The Bambuí Group represents a large foreland basin related to the late stages of the Brasiliano orogeny, forming a wedge-shaped succession extending across central Brazil (Uhlein et al. Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019). It consists of up to ∼3,000 m of mixed siliciclastic–carbonate deposits, organized (from base to top) into the Carrancas/Jequitaí, Sete Lagoas, Serra de Santa Helena, Lagoa do Jacaré, Serra da Saudade and Três Marias formations (Dardenne, Reference Dardenne1978; Martins-Neto and Alkmim, Reference Martins-Neto, Alkmim, Pinto and Martins-Neto2001; Uhlein et al. Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019). The basal Carrancas Formation records pre-Marinoan glacial deposition (Uhlein et al. Reference Uhlein, Uhlein, Halverson, Stevensson, Caxito, Cox and Carvalho2016), overlain by the Sete Lagoas Formation, which includes Marinoan cap carbonates and terminal Ediacaran fossil assemblages (Warren et al. Reference Warren, Quaglio, Riccomini, Simões, Poiré, Strikis, Anelli and Strikis2014, Reference Warren, Becker-Kerber, Inglez, Varejão, Morais, Simões, Freitas, Arrouy, Gómez-Peral, Poiré, Okubo, Caxito, Uhlein, Antunes, Lehn, Romero and Fairchild2024; Paula Santos et al. Reference Paula-Santos, Babinski, Kuchenbecker, Caetano-Filho, Trindade and Pedrosa-Soares2015; Caxito et al. Reference Caxito, Lana, Frei, Uhlein, Sial, Dantas, Pinto, Campos, Galvão, Warren, Okubo and Ganade2021). Overlying units record a major transgressive–regressive cycle, with fine-grained siliciclastics and shallow-marine carbonates (Uhlein et al. Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019; Moreira et al. Reference Moreira, Uhlein, Dussin, Uhlein and Misuzaki2020), followed by the Três Marias Formation, composed of fluvial to shallow-marine siliciclastic deposits of probable Cambrian age (Uhlein et al. Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019; Tavares et al. Reference Tavares, Souza Martins, Alkmim and Lana2020).
The Santa Fé Group preserves an important record of the LPIA in central Brazil. It unconformably overlies the Bambuí Group and is preserved within paleodepressions interpreted as glacial valleys incised into the Três Marias Formation (Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a). The group comprises the Floresta Formation, subdivided into the Brocotó, Brejo do Arroz and Lavado members and the overlying Tabuleiro Formation (Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a). These units record proglacial, glacial-lacustrine, fluvio-glacial and periglacial aeolian environments, with abundant glaciogenic features and trace fossils (Cruziana diplopoda, Cruziana problematica, Diplichnites gouldi, Diplopodichnus biformis, Helminthoidichnites tenuis and Rusophycus carbonarius) that combined with paleomagnetic data support a Pennsylvanian–Cisuralian age (Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a; Brandt and Ernesto, Reference Brandt and Ernesto2006; Brandt et al. Reference Brandt, Ernesto, Rocha-Campos and Santos2009; de Barros et al. Reference de Barros, Carmo, Andrade, Assine and Sedorko2025).
3. Materials and methods
Sedimentological and stratigraphic data were obtained from nine columnar sections (1:100) in successions corresponding to the Serra de Santa Helena, Lagoa do Jacaré and Serra da Saudade formations (Bambuí Group) outcropping in the region of Ibiracatu and nearby municipalities in the northern state of Minas Gerais (Figure 1; Table 1). Several trace fossils were observed throughout Section 2, representing the interval of interest in this study. The description of the sedimentary facies was determined using the methodology of Miall (Reference Miall1996), with details of the mineralogical constituents, textural aspects, syn- and post-depositional sedimentary structures and trace fossil content.
Columnar sections measured in the north of the state of Minas Gerais

Table 1. Long description
The table presents data from nine columnar sections measured in the north of the state of Minas Gerais. It includes details such as the section number, location, geographic coordinates (latitude and longitude in UTM format), thickness in meters, and the corresponding geological unit. The locations range from between Januária and Bonança to BR-135, km 228, with varying thicknesses and units such as Serra de St. Helena Fm., Floresta Fm., St. Fé Group, and others. The table provides a comprehensive overview of the sedimentological and stratigraphic data collected from these sections.
Forty-five samples with trace fossils were collected in the field, organized, catalogued and stored in the Sedimentology Laboratory (DEGEO/UFOP) under the numbers LS-261 to LS-305. The material was cleaned and photographed using a Nikon D90 camera with an 18–55 mm lens. The images were processed using software to highlight trace fossils for identification and classification. The specimens were classified at the ichnogenus level and, when possible, ichnospecies. The samples with trace fossil content were also classified according to the form of preservation of the trace fossils and bioturbation index, in addition to the establishment of the trace plot and ichnodiversity. The classification in relation to the form of preservation in the layer was based on the work of Seilacher (Reference Seilacher1953, Reference Seilacher, Imbrie and Newell1964); the degree of bioturbation was based on the scale proposed by Reineck (Reference Reineck1963). In addition, reference works (Buatois and Mángano, Reference Buatois and Mángano2013; Buatois et al. Reference Buatois, Wisshak, Wilson and Mángano2017, Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025) were used to define the architectural designs of the studied ichnofauna.
At the outcrop of the Ibiracatu trace fossil unit, approximately 2 kg of material was collected from a sandstone bed at the top of the succession for U-Pb geochronological analyses of detrital zircons, with the aim of defining the maximum depositional age for the stratigraphic interval related to the trace fossils. The zircon grains were extracted from the sample using traditional heavy mineral concentration techniques, such as crushing, sieving, heavy mineral concentration in a pan and magnetic separation with the Frantz equipment. The entire sample processing stage was performed at the Geochronology Sample Preparation Laboratory (LOPAG) of the Geology Department of the Federal University of Ouro Preto (DEGEO/UFOP). After separation, the heavy mineral concentrate was taken for manual collection of zircon grains using a binocular magnifying glass and, subsequently, organized in an epoxy resin tablet (mount) containing approximately 220 zircon grains. The wafer was polished for cathodoluminescence (CL) imaging of the grains using a JEOL 6510 Scanning Electron Microscope (SEM) at the Microanalysis Laboratory of DEGEO/UFOP, in order to characterize the internal structures of the individual zircon grains. The U-Pb analysis was performed at the Isotopic Geochemistry Laboratory of the Federal University of Ouro Preto, using a Thermo-Finnigan Element II (sector field – SF) ICPMS instrument coupled to a CETAC Nd:YAG 213 nm ultraviolet laser with a Helix abrasion cell (LA-SFICP-MS). Data were acquired in automatic peak jumping mode with 20 µm diameter holes and with approximately 40 s of analysis, with 20 s of background acquisition followed by 20 s of data acquisition. The zircon standard GJ1, 602 ± 1 Ma (Jackson et al. Reference Jackson, Pearson, Griffin and Belousova2004) was used as the primary reference material for this study, and the BB, 560 ± 1 Ma (Santos et al. Reference Santos, Lana, Scholz, Buick, Kamo, Gerdes, Condon, Corfu, Tohver, Storey, Basei, Krambrock and Fantini2017; Lana et al. Reference Lana, Farina, Gerdes, Alkmim, Gonçalves and Jardim2017) and Plešovice, 337 ± 0.37 (Sláma et al. Reference Sláma, Košler, Condon, Crowley, Gerdes, Hanchar, Horstwood, Morris, Lutz, Norberg, Schaltegger, Schoene, Tubrett and Whitehouse2008) zircon standards as secondary and tertiary reference material, respectively. Data reduction was performed using the SATURN® software (Silva et al. Reference Silva, Lana, Mazoz, Buick and Scholz2022). Zircon grains that presented dislocations >5%, high common Pb content and U/Th ratio < 0.1, which may indicate metamorphic ages (Hartmann and Santos, Reference Hartmann and Santos2004), were discarded and are not presented in the supplementary tables. The probability density plot of U-Pb zircon ages and the concordia diagram were obtained using Isoplot/Ex software (Ludwig Reference Ludwig2003, Reference Ludwig2008). For ages below 1400 Ma, 206Pb/238U ages were used, and for ages above 1400 Ma, 207Pb/206Pb ages were used (Gehrels, Reference Gehrels2014; Luo et al. Reference Luo, Qi and Xia2023). Final ages are quoted with a 95% confidence interval.
4. Results
4.a. Sedimentary facies and stacking patterns of the Bambuí Group
The Sete Lagoas Formation, which outcrops on the west side of the São Francisco River near the Januária municipality, and the Três Marias Formation that occur further to the northeast were not included here as no outcrops of these units were found in the studied region. The study succession is 245 m thick and comprises the Serra da Saudade, Lagoa do Jacaré and Serra da Saudade formations (Figure 1C).
4.a.1. Serra de Santa Helena Formation
The Serra de Santa Helena Formation is ca. 105 m thick (see Figure 1C; Supplementary Materials, Table S1), and probably thicker as its lower and upper boundaries with the Sete Lagoas and Lagoa do Jacaré formations, respectively, were not observed in the studied area. The lower part of the preserved succession (16 m thick) is composed of coarsening upward successions (Figure 3A) of massive to laminated siltstone (Fm and Fl), interbedded siltstone with lenticular bedding horizontally (Hl), horizontally laminated (Sl) and wave ripple cross-laminated sandstone (Sr), and hummocky cross-stratified sandstone (Sh) (Figure 3B-3C), as well as sandstone with wavy bedding presenting desiccation cracks and microbially induced sedimentary structures (MISS) (Figure 3D). Sandstone facies are locally associated with post-depositional structures, such as convolute lamination and fluid escape structures (see Figure 3C). The intermediate succession (81 m thick) comprises 2–5 m thick coarsening upward successions of massive to laminated siltstone (Fm and Fl), horizontally laminated and wave ripple cross-laminated sandstone (Sr) (Figure 3E). Siltstone intraclasts and local siltstone drapes are observed in association with sandstone facies. The upper succession (9 m thick) of the unit is marked by horizontally laminated sandstone with MISS presenting irregular micro-polygons similar to elephant skin type and laminated siltstone with desiccation cracks (Figure 3F).
Facies from the Serra de Santa Helena Formation. (A) Shoreface-offshore transition facies (Hl, Sl, Fm) and shoreface facies (Hw). (B) Succession of linsen bedding (Hl) with sandstone with wave ripple cross-lamination (Sr) and hummocky cross-stratification (Sh). (C) Sandstone with horizontal lamination (Sl), displaying fluid escape structure at base of the tabular set. (D) Lower part of a sandstone bed over siltstone presenting desiccation crack and MISS. (E) Coarsening upward successions (Fl to Sl and Sr), thinning towards the top of the image. (F) MISS presenting irregular micro-polygons similar to elephant skin type.

Figure 3. Long description
The image displays a detailed geological representation of the Sanfranciscana Basin in the northern portion of the state of Minas Gerais, Brazil. It includes several panels showing different aspects of the geological formations. Panel A shows the So Francisco Craton geological map, highlighting various rock units and structures. Panel B presents a geological map of the Sanfranciscana Basin in the study area, detailing the distribution of different sedimentary facies. Panel C illustrates a stratigraphic column of outcrops of the Bambu Group, indicating the vertical succession of different sedimentary layers. The legend provides information on the composition of these layers, including clay, silt, sand, gravel, pebble, mudstone, wackestone, packstone, fine grainstone, and medium grainstone. Panels D, E, and F show specific sedimentary structures and facies, such as shoreface-offshore transition facies, linsen bedding with wave ripple cross-lamination, hummocky cross-stratification, sandstone with horizontal lamination, fluid escape structures, desiccation cracks, micro-polygons, and coarsening upward successions.
4.a.2. Lagoa do Jacaré Formation
The Lagoa do Jacaré Formation is ca. 54 m thick in the outcrop area, showing a transitional upper boundary with the Serra da Saudade Formation (see Figure 1C; Supplementary Materials, Table S2). The lower succession (34 m thick) is composed of marl, grainstone and boundstone organized in 1–4 m thick coarsening upward stacking patterns (Figure 4A). Grainstone have tabular to lenticular geometry and present horizontal lamination (Gl), wave ripple cross-lamination (Gr), and planar (Gt), swaley and hummocky cross-stratifications (Gh) (Figure 4B-C). Big-scale (10 cm high and 20 cm length) wave ripple cross-laminated grainstone (Gr) are common (Figure 4D). Boundstone is characterized by nodular stromatolites (Bn) as well as thrombolites (Bt) (Figure 4E). In the top of this succession, occur a ∼5 m thick succession of trough cross-stratified ooid grainstone (Gt), organized in wedge-shaped sets (Figure 4F).
Facies from the lower Lagoa do Jacaré Formation. (A) Coarsening and shallowing upward stacking patterns. (B) Detail of the transition from shoreface-offshore transition to shoreface facies. (C) Interbedded grainstone (Gl, Gr and Gh) and thin (mm-scale) marl facies. (D) Wave ripple cross-laminated grainstone (Gr) and thrombolites (Bt). (E) Nodular microbialites (Bn) and thrombolites (Bt). (F) Compound bar of ooid grainstone with trough cross-stratification (Gt).

Figure 4. Long description
The image displays a detailed cross-sectional view of various geological facies from the lower Lagoa do Jacaré Formation. In section A, there is a coarsening and shallowing upward stacking pattern. Section B shows the transition from shoreface-offshore transition to shoreface facies. Section C features interbedded grainstone types (Gl, Gr, and Gh) with thin marl facies. Section D highlights wave ripple cross-laminated grainstone (Gr) and thrombolites (Bt). Section E depicts nodular microbialites (Bn) and thrombolites (Bt). Section F illustrates a compound bar of ooid grainstone with trough cross-stratification (Gt).
The upper succession is a 19 m thick mixed carbonate-siliciclastic succession, which transitions upward to the Serra da Saudade Formation. The succession comprises laminated siltstone (Fl); sandstone with wave ripple cross-lamination (Sr) and wavy to flaser beddings (Hw and Hf); massive (Fm), horizontal (Fl) and wrinkle laminated marl; grainstone with horizontal lamination (Gl), wave ripple cross-lamination (Gr) and planar (Gt), swaley and hummocky cross-stratifications (Gh); and mudstone with wrinkle lamination (Figure 5A-D). In addition, post-depositional features such as fluid escape structures, tepees and evaporitic breccia (Re) are observed in grainstone (Figure 5A-B), while desiccation cracks are observed in the top of the succession in association with marl.
Facies from the upper Lagoa do Jacaré (A–D) and Serra da Saudade (E–I) formations. (A) Interbedded laminated marl (Fl(m)) and grainstone (Gr and Gl), with evaporitic breccia (Re) at top of the succession. (B) Detail of the evaporitic breccia. (C) Grainstone with horizontal lamination (Gl). (D) Upper view of a marl bed presenting desiccation cracks. (E) Interbedded laminated siltstone (Fl) and hummocky cross-stratified sandstone (Sh). (F) Detail of giant-scale (10 cm high and 30 cm length) wave ripple cross-laminated sandstone (Sr). (G) Detail of Swalley cross-stratified sandstone (Ss). (H) Load casts and pseudonodules in intensely deformed sandstone (Sm(d)). (I) Upper view of horizontally laminated sandstone bed showing gypsum pseudomorphs.

Figure 5. Long description
The image displays a cross-sectional view of geological formations from the upper Lagoa do Jacaré and Serra da Saudade formations. It includes interbedded laminated marl and grainstone with evaporitic breccia at the top. The detailed view of the evaporitic breccia is shown, along with grainstone exhibiting horizontal lamination. A marl bed with desiccation cracks is visible from an upper perspective. The section also features interbedded laminated siltstone and hummocky cross-stratified sandstone. A close-up of giant-scale wave ripple cross-laminated sandstone is included, as well as a detailed view of swaley cross-stratified sandstone. Additionally, there are load casts and pseudonodules in intensely deformed sandstone, and an upper view of a horizontally laminated sandstone bed showing gypsum pseudomorphs.
4.a.3. Serra da Saudade Formation
In the study area, the Serra da Saudade Formation is 84 m thick, comprising marl, siltstone and sandstone (see Figure 1C; Supplementary Materials, Table S3). The lower succession comprises up to 7 m thick stacking patterns characterized, from base to the top, by massive to laminated siltstone (Fm and Fl), sandstone with wave ripple cross-laminated (Sr) and hummocky cross-stratification (Sh) (Figure 5E), and up to 2 m thick sandstone with horizontal lamination (Sl) at top. The upper succession is organized in up to 4 m thick stacking patterns with interbedded massive siltstone (Fm) and sandstone with horizontal lamination (Fl), wave ripple cross-lamination (Sr) and hummocky/swaley cross-stratifications (Sh) (Figure 5F, G), with horizontally laminated sandstone (Sl) in the top. Locally, massive sandstone with pseudonodules (Sm(d)) (Figure 5H) also occurs. Sandstone bedding planes can present lenticular gypsum pseudomorphs at the upper part of the coarsening upward stacking patterns (Figure 5I).
4.b. Ichnofossil-bearing succession
A 33 m thick succession of interbedded sandstone and red siltstone and claystone, containing abundant trace fossils, are exclusively described in the vicinities of the Ibiracatu locality (Figure 6). This succession crops out laterally to the Lagoa do Jacaré and Serra da Saudade formations (Figure 7A, B). The boundary between the ichnofossil-bearing succession and the Serra da Saudade Formation is marked by a reverse fault, marking an inversed stratigraphy (Figure 7C, D). However, the ichnofossil-bearing succession presents normal faults near the boundary with the Serra da Saudade Formation (Figure 7E, F) with beds dipping (∼25º) to NE. Interestingly, the boundary between the Lagoa do Jacaré and Serra da Saudade is not transitional as regionally observed but made by a normal fault (Figure 7G, H). The letters A to F in Figure 6 refer to the sets of samples with trace fossil content collected in the field section. A) LS-261 to LS-266, B) LS-267 to LS-271, C) LS-272 to LS-275, D) LS-276 to LS-279, E) LS-280 to LS-283 and F) LS-284 to LS-288. It was not possible to define the stratigraphic level of the samples numbered LS-289 to LS-305.
Columnar section of the trace fossil-bearing succession, showing the distribution of the trace fossil occurrences. Letters A to F are references for position of chart diagrams displayed in Figure 12. T1-T4 refers to the intervals of the Bouma Sequence applied to the studied section. C – Clay; St – Silt; fS – fine Sand; mS – medium Sand.

Figure 6. Long description
The image presents a columnar section of a trace fossil-bearing succession, illustrating the distribution of trace fossil occurrences. The section is divided into intervals labeled T1 to T4, corresponding to the Bouma Sequence applied to the studied section. Different lithologies are represented by various colors: clay, siltstone, fine sandstone, and medium sandstone. The image includes labels A to F, which reference the positions of chart diagrams displayed in another figure. The lithologies are marked with different patterns indicating horizontal lamination, ripple cross-lamination, sole marks, and tool marks. Various ichnofossils such as Circulichnus, Cruziana, Diplichnites, Diplocraterion, Helminthoidichnites, Lockeia, Palaeophycus, Problematica, Rusophycus, and Treptichnus are also depicted.
Boundary of the ichnofossil-bearing succession and the Bambuí Group. (A, B) Panoramic view of the ichnofossil-bearing succession (in the background) outcropping laterally to the Serra da Saudade Formation. (C, D) Reverse fault between the Serra da Saudade Formation and ichnofossil-bearing succession. (E, F) Normal faults observed in laminated siltstone at base of the trace fossil succession. (G, H) Faulted contact between the Lagoa do Jacaré and Serra da Saudade formations.

Figure 7. Long description
The image presents a detailed cross-sectional view of geological formations. It highlights the boundary between the ichnofossil-bearing succession and the Bambuí Group. The ichnofossil-bearing succession is visible in the background, outcropping laterally to the Serra da Saudade Formation. The image also shows a reverse fault between the Serra da Saudade Formation and the ichnofossil-bearing succession. Additionally, normal faults are observed in laminated siltstone at the base of the trace fossil succession. The faulted contact between the Lagoa do Jacaré and Serra da Saudade formations is also depicted. The image includes annotations and labels to indicate these geological features.
The trace fossil-bearing succession comprises a thickening and coarsening upward stacking pattern with five microfacies (Figure 6; Table 2). The lower succession, deposited laterally and over the Bambuí Group in a fault boundary, is dominated by massive (Fm) and laminated (Fl) siltstone at base (Figure 8A). The succession above is marked by fining upward stacking patterns (0.15 to 1.1 m thick), characterized by different proportions of massive (Sm), horizontally laminated (Sl), current ripple cross-laminated (Sr) sandstone and laminated siltstone (Fl) beds (Figure 8B–E). Sandstone beds are 0.05–0.3 m thick and laminated siltstone beds are 0.2 to 1 mthick. Thickness and grain size of sandstone beds increase towards the top, which can present local occurrences of siltstone intraclasts. Sandstone is organized into lenticular to tabular beds with flat to erosive base and undulating top (Figure 8E), with high degree of cementation by calcium carbonate. Beds dominated by Sm facies often have an erosive base, while those with Sl and Sr show dominant flat bases. The base of sandstone beds commonly present groove marks (sensu Peakall et al. Reference Peakall, Best, Baas, Hodgson, Clare, Talling, Dorrell and Lee2020; Baas et al. Reference Baas, Tracey and Peakall2021) (Figure 8F) and sole marks (Figure 8G), the last presenting a length/width/depth ratio of a few centimeters and characterized as parabolic flutes (sensu Peakall et al. Reference Peakall, Best, Baas, Hodgson, Clare, Talling, Dorrell and Lee2020; Baas et al. Reference Baas, Tracey and Peakall2021). Cross-laminations and sole marks indicate N–W flow directions. Trace fossils are observed throughout this intermediate succession, occurring in the top of laminated siltstone (with negative to positive epirelief), sandstone with massive appearance and with asymmetric cross-lamination (with negative and positive hyporelief) (Figure 8H). The degree of bioturbation is low (0–1; Reineck, Reference Reineck1963).
Sedimentary facies identified in the ichnofossiliferous succession

Table 2. Long description
The table presents a comparison of sedimentary facies identified in an ichnofossiliferous succession. It includes four columns: Code, Facies, Description, and Interpretation. The table has five rows, each detailing different facies types such as massive claystone, laminated siltstone, massive sandstone, laminated sandstone, and asymmetric wavy cross-laminated sandstone. Each row provides specific descriptions and interpretations of the facies, including bed thickness, composition, and depositional environments. Notable trends include the decantation of fine suspended sediments in calm and oxygenated waters for siltstone facies, and the transport of sandy sediments by traction in flat bedforms for sandstone facies. The table also mentions the occurrence of ichnofossils and the colonization of organisms in various facies.
Facies of the ichnofossiliferous succession. (A) Interbedded laminated siltstone (Fl) and massive siltstone (Fm). (B) Thinning upward successions of sandstone and siltstone. (C) Amalgamation of four events, the first with sandstone with current-ripple cross-lamination (Sr), the second with massive sandstone (Sm), and the third and fourth with sandstone with horizontal lamination (Sl). (D) Flat base and undulated bed of current-ripple cross-laminated sandstone (Sr) interbedded in massive siltstone (Fm). (E) Ripple marks in the upper view of the bed. (F) Tool mark structures along with some trace fossils. (G) Sole mark structures observed in hand sample. (H) Trace fossils preserved as positive and negative hyporelief.

Figure 8. Long description
The image displays several types of sedimentary structures in a geological formation. Panel A shows interbedded laminated siltstone (Fl) and massive siltstone (Fm). Panel B illustrates thinning upward successions of sandstone and siltstone. Panel C depicts the amalgamation of four events, including sandstone with current-ripple cross-lamination (Sr), massive sandstone (Sm), and sandstone with horizontal lamination (Sl). Panel D features a flat base and undulated bed of current-ripple cross-laminated sandstone (Sr) interbedded in massive siltstone (Fm). Panel E highlights ripple marks in the upper view of the bed. Panel F shows tool mark structures along with some trace fossils. Panel G presents sole mark structures observed in a hand sample. Panel H displays trace fossils preserved as positive and negative hyporelief.
In the upper part of the studied section, the fining upward stacking patterns (0.4 to 1.5 m thick) are dominated by sandstone with massive aspect and laminated siltstone. Sandstone beds are 0.1 to 1.1 m thick, while the laminated siltstone beds are 0.1 to 1.5 m thick. Interestingly, trace fossils are rarely found in this part of the succession, and the rocks, particularly sandstone, are considerably friable.
4.b.1. Systematic ichnology
Ichnogenus Circulichnis Vialov (Reference Vialov1971)
Circulichnis ligusticus Uchman and Rattazzi (Reference Uchman and Rattazzi2019)
Material: Three samples preserving five specimens (LS-272, LS-278, LS-280; Figure 9A).
Ichnofossiliferous succession trace fossils. (A) Sandstone sample with Circulichnis ligusticus (Ci), Treptichnus isp. (Tr). (B) Sandstone sample with Cruziana diplopoda (Cd), Cruziana problematica (Cp) and Diplopodichnus biformis (Db). (C) Sandstone sample with Cruziana diplopoda (Cd), Cruziana problematica (Cp), Diplopodichnus biformis (Db). (D) Sandstone sample with Diplichnites gouldi (Dg). (E) Sandstone sample with Diplichnites gouldi (Dg), Diplopodichnus biformis (Db) and Palaeophycus tubularis (Pa).

Figure 9. Long description
The image displays five sandstone samples, each containing different types of trace fossils. Sample A features Circulichnis ligusticus and Treptichnus isp. Sample B contains Cruziana diplopoda, Cruziana problematica, and Diplopodichnus biformis. Sample C also shows Cruziana diplopoda, Cruziana problematica, and Diplopodichnus biformis. Sample D exhibits Diplichnites gouldi. Sample E includes Diplichnites gouldi, Diplopodichnus biformis, and Palaeophycus tubularis. Each sample is labeled with the corresponding scientific names of the trace fossils.
Description: Horizontal ring-shaped, subcylindrical, smooth and unlined trail. The course of the trace is sinuous or irregular and may present breaks along the way. The external width of the trail varies from 3.08 to 1.20 mm, varying in some specimens. Preserved in sandstone as positive hyporelief.
Remarks: Originally described by Vialov (Reference Vialov1971), Circulichnis is interpreted as a grazing trail (pascichnia) produced by annelids or other worm-like organisms (Pickerill and Keppie, Reference Pickerill and Keppie1981; Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a; Dernov, Reference Dernov2024). In a recent study, Uchman and Rattazzi (Reference Uchman and Rattazzi2019) distinguished a new ichnospecies (Circulichnis ligusticus) and proposed that the ichnogenus Circulichnis represents a feeding behaviour of exploration of different sediment layers in search of food. The burrow would be produced by polychaetes in marine sediments and oligochaetes in continental sediments (Uchman and Rattazzi, Reference Uchman and Rattazzi2019). Circulichnis (at the ichnogenus level) is reported in different depositional environmental since the Ediacaran (Uchman and Rattazzi, Reference Uchman and Rattazzi2019; Dernov, Reference Dernov2024; Gougeon et al. Reference Gougeon, Mángano, Buatois, Narbonne, Laing and Paz2025), such as lacustrine, fluvial, deltaic, shallow marine, deep marine facies and turbidite deposits (Pickerill and Keppie Reference Pickerill and Keppie1981; Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a; Jackson et al. Reference Jackson, Hasiotis and Flaig2016; Uchman and Rattazzi Reference Uchman and Rattazzi2019; Bouchemla et al. Reference Bouchemla, Bendella, Benyoucef, Vinn and Ferré2021; Dernov Reference Dernov2024; Gougeon et al. Reference Gougeon, Mángano, Buatois, Narbonne, Laing and Paz2025).
Ichnogenus Cruziana d’Orbigny (Reference D’Orbigny1842)
Cruziana diplopoda Schatz et al. (Reference Schatz, Mángano, Buatois and Limarino2011)
Material: Two samples preserving six specimens (LS-282, LS-301; Figure 9B, C).
Description: Small, straight and slightly curved bilobed trails. On the surface of the lobes, very fine and poorly preserved striations are observed perpendicular to the axis of the trail, with angles of 90–110º to the midline. The external width of the lobes varies from 1.5 to 2.8 mm. The width of the central groove varies considerably, being 1.2 mm when the lobes are parallel and short, reaching 3.3 mm when the lobes are further apart. The length of the track can reach 60 mm. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by d’Orbigny (Reference D’Orbigny1842), Cruziana is interpreted as a locomotion trail (repichnia) produced by arthropods, mainly trilobites, isopods, and myriapods (Keighley and Pickerill, Reference Keighley and Pickerill1996; Lima et al. Reference Lima, Minter and Netto2017). The specimens described resemble Cruziana diplopoda (Schatz et al. Reference Schatz, Mángano, Buatois and Limarino2011) in the pattern of morphological variation along the trail, changes in the distance between narrow and parallel lobes to separate lobes with an increase in the width of the central groove. In addition, some specimens also present less relief along the trail, indicating a change in burrowing behaviour (Schatz et al. Reference Schatz, Mángano, Buatois and Limarino2011). Cruziana diplopoda was first reported in glacially influenced diamictite deposits of the late Carboniferous in Argentina (Schatz et al. Reference Schatz, Mángano, Buatois and Limarino2011) and was recently described by de Barros et al. (Reference de Barros, Carmo, Andrade, Assine and Sedorko2025) from a turbiditic deposit formed under glacial influence within the Santa Fé Group in the municipality of Santa Fé de Minas, northwestern Minas Gerais State (MG).
Ichnogenus Cruziana d’Orbigny (Reference D’Orbigny1842)
Cruziana problematica Schindewolf (Reference Schindewolf1921)
Material: Nine samples preserving 20 specimens (LS-275, LS-276, LS-282, LS-285, LS-286, LS-290, LS-297, LS-299, LS-301; Figures 9B-C and 11A, D).
Description: Bilobate trail composed of two lateral lobes divided by a central groove. Very fine curved striae perpendicular to the axis of the trace are observed on the surface of the lobes. The external width of the lobes varies from 1.90 to 2.85 mm, and the width of the central groove varies from 1.45 to 2.20 mm. The angle of the striae is approximately 91º with the midline. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by Schindewolf (Reference Schindewolf1921), Cruziana problematica is interpreted as a locomotion trail (repichnia) produced by arthropods, mainly trilobites, isopods and myriapods (Keighley and Pickerill, Reference Keighley and Pickerill1996; Lima et al. Reference Lima, Minter and Netto2017). Cruziana problematica has been reported in marine and continental deposits since the Cambrian (Hammersburg et al. Reference Hammersburg, Hasiotis and Robison2018) with a tendency to be found in LPIA deposits (Keighley and Pickerill, Reference Keighley and Pickerill1997; Garvey and Hasiotis, Reference Garvey and Hasiotis2008; Schatz et al. Reference Schatz, Mángano, Buatois and Limarino2011; Balistieri et al. Reference Balistieri, Netto and Sedorko2021; Buatois et al. Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025), being commonly described in glacial deposits of the Itararé Group (Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021; Lima et al. Reference Lima, Minter and Netto2017; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021; Balistieri et al. Reference Balistieri, Netto and Sedorko2021) and from a turbiditic deposit formed under glacial influence within the Santa Fé Group (de Barros et al. Reference de Barros, Carmo, Andrade, Assine and Sedorko2025).
Ichnogenus Diplichnites Dawson (Reference Dawson1873)
Diplichnites gouldi (Gevers et al. Reference Gevers, Frakes, Edwards and Marzolf1971)
Material: Twelve samples preserving 21 specimens (LS-262, LS-263, LS-265, LS-268, LS-269, LS-276, LS-278, LS-279, LS-286, LS-288, LS-302, LS-304; Figures 9D–E and 10E).
Ichnofossiliferous succession trace fossils. (A) Siltstone sample with Diplopodichnus biformis (Db), Helminthoidichnites tenuis (Ht). (B) Sandstone sample with Hormosiroidea meandrica (Ho). (C) Sandstone sample with Diplopodichnus biformis (Db), Lockeia siliquaria (Lo). (D) Sandstone sample with Lockeia siliquaria (Lo). (E) Sandstone sample with Diplichnites gouldi (Dg), Diplopodichnus biformis (Db) and Palaeophycus tubularis (Pa).

Figure 10. Long description
The image displays a siltstone sample featuring trace fossils. The sample includes two distinct types of trace fossils: Diplopodichnus biformis, labeled as Db, and Helminthoidichnites tenuis, labeled as Ht. The trace fossils are visible as distinct patterns within the siltstone, with Db appearing as linear structures and Ht as branching or meandering lines. The sample is shown at a scale of 1 centimeter for reference.
Ichnofossiliferous succession trace fossils. (A) Sandstone sample with Cruziana problematica (Cp), Rusophycus isp. (Ri). (B) Sandstone sample with Treptichnus isp. (Tr). (C) Sandstone sample with ‘problematica’ structures (Pr). (D) Sandstone sample with Cruziana problematica (Cp), Diplopodichnus biformis (Db). (E) Sandstone sample with Diplopodichnus biformis (Db). (F) Sandstone sample with Diplopodichnus biformis (Db), Rusophycus carbonarius (Rc).

Figure 11. Long description
The image displays six sandstone samples, each containing different trace fossils. Sample A features Cruziana problematica and Rusophycus isp. Sample B showcases Treptichnus isp. Sample C contains problematic structures. Sample D includes Cruziana problematica and Diplopodichnus biformis. Sample E highlights Diplopodichnus biformis. Sample F presents Diplopodichnus biformis and Rusophycus carbonarius. Each sample is labeled with the respective scientific names of the trace fossils.
Description: Simple trackways composed of two parallel and continuous rows of regularly spaced tracks. The tracks are elongated to an ellipsoid and perpendicular to the trace axis. The external width ranges from 1.8 to 3.10 mm, and the internal width ranges from 3.15 to 6.10 mm. The ratio between external and internal width is ∼1:1 to ∼1:2, varying in some specimens. Preservation in sandstone as positive hyporelief, with occasional transitions to Diplopodichnus biformis.
Remarks: Originally described by Dawson (Reference Dawson1873), Diplichnites was interpreted as being a product of the action of large worms or crustaceans. Gevers et al. (Reference Gevers, Frakes, Edwards and Marzolf1971) interpreted the ichnospecies Diplichnites gouldi as being a product of the locomotion activity (repichnia) of arthropods, probably trilobites. Later, other authors suggested myriapods as producers of the trace (Bradshaw, Reference Bradshaw1981; Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a, Reference Buatois, Mángano, Maples and Lanier1998b; Netto et al. Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021). Diplichnites gouldi has been reported in different depositional settings, although it is presently interpreted as typical of shallow-water lakes subjected to subaerial exposure, commonly associated with glacial environments (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a, Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025; Lima et al. Reference Lima, Netto, Corrêa and Lavina2015; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025; Netto et al. Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021). The association between Diplopodichnus biformis and Diplichnites gouldi has been reported by several authors as being a product of the animal displacement in sporadically exposed rhythmic facies (Gandini et al. Reference Gandini, Netto and Souza2007; Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021; Lima et al. Reference Lima, Netto, Corrêa and Lavina2015, Reference Lima, Minter and Netto2017; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021). Diplichnites gouldi is described in successions from the Terraneuvian to the Carboniferous (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a; Buatois and Mángano, Reference Buatois and Mángano2018).
Ichnogenus Diplopodichnus Brady (Reference Brady1947)
Diplopodichnus biformis Brady (Reference Brady1947)
Material: Thirty nine samples preserving 179 specimens (LS-261, LS-262, LS-263, LS-264, LS-265, LS-266, LS-267, LS-268, LS-269, LS-271, LS-272, LS-273, LS-274, LS-275, LS-276, LS-277, LS-278, LS-279, LS-280, LS-281, LS-282, LS-283, LS-285, LS-286, LS-287, LS-288, LS-290, LS-291, LS-292, LS-293, LS-296, LS-298, LS-299, LS-300, LS-301, LS-302, LS-303, LS-304, LS-305; Figures 9B–C, E, 10A, C, E, 11D–F).
Description: Two rows of continuous, parallel, straight-to-sinuous bilobated trails, separated by a central furrow. External width varies from 2.10 to 6.25 mm and internal width varies from 2.15 to 3.8 mm. The ratio between external and internal width is ∼1:1 to ∼1:2, varying in some specimens. Preservation in sandstone as positive hyporelief, with occasional transitions to Diplichnites gouldi.
Remarks: Originally described by Brady (Reference Brady1947), Diplopodichnus biformis is a very common ichnospecies in the samples studied. It was interpreted as a product of the locomotion activity (repichnia) of arthropods by Buatois et al. (Reference Buatois, Mángano, Maples and Lanier1998b), being generated by the same tracemaker as Diplichnites. This bilobed trail was recorded in different depositional environments, such as freshwater deposits, lacustrine facies and aeolian deposits (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a). The occurrences of Diplopodichnus biformis are more related to the mode of preservation and the consistency of the substrate than to a particular behaviour (Keighley and Pickerill, Reference Keighley and Pickerill1996). Associations between Diplopodichnus biformis and Diplichnites gouldi were reported by Netto et al. (Reference Netto, Balistieri, Lavina and Silveira2009) in the Mafra Formation (Itararé Group, Paraná Basin) and by de Barros et al. (Reference de Barros, Carmo, Andrade, Assine and Sedorko2025) in the Floresta Formation (Santa Fé Group, Sanfranciscana Basin). Diplopodichnus biforms is described in successions from the Terraneuvian to the Permian (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a; Buatois and Mángano, Reference Buatois and Mángano2018).
Ichnogenus Helminthoidichnites Fitch (Reference Fitch1850)
Helminthoidichnites tenuis Fitch (Reference Fitch1850)
Material: Two samples preserving three specimens (LS-268, LS-292; Figure 10A).
Description: Simple horizontal, unbranched trail, with a straight to curved trajectory, but not sinuous. Overcrossing between individuals is common. The width varies from 0.45 to 1.50 mm, changing along with the trajectory of individual specimens. The maximum length is 76.08 mm. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by Fitch (Reference Fitch1850), Helminthoidichnites tenuis is interpreted as a grazing trace (pascichnia) probably produced by annelids, insect larvae or arthropods (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a, Reference Buatois, Mángano, Maples and Lanier1998b; Lima et al. Reference Lima, Netto, Corrêa and Lavina2015, Reference Lima, Minter and Netto2017). Helminthoidichnites tenuis is reported in several depositional environments, being common in freshwater environments, especially in Carboniferous successions (Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998a; Buatois and Mángano, Reference Buatois and Mángano2003; Netto et al. Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a; Lima et al. Reference Lima, Minter and Netto2017; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025).
Ichnogenus Hormosiroidea Schaffer (Reference Schaffer1928)
Hormosiroidea meandrica (Balistieri et al. Reference Balistieri, Netto and Lavina2002)
Material: One sample preserving one specimen (LS-298; Figure 10B).
Description: Horizontal burrows composed by consecutive nodes, without branches and with a sinuous course. Burrows are 1 to 1.5 mm wide and up to 15 mm in length. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by Balisteri et al. (Reference Balistieri, Netto and Lavina2002), Hormosiroidea meandrica is interpreted as a feeding trace (fodinichnia). Although no organism is proposed as a possible producer, considering the small size of the described specimens, worm-like organisms can be considered as probable producers of the trace (Gandini et al. Reference Gandini, Netto and Souza2007). This ichnotaxon has been reported in freshwater environments, especially in Permian–Carboniferous rocks of the Itararé Group (Balistieri et al. Reference Balistieri, Netto and Lavina2002, Reference Balistieri, Netto and Sedorko2021; Gandini et al. Reference Gandini, Netto and Souza2007; Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009; Buatois et al. Reference Buatois, Netto, Mángano, López-Gamund and Buatois2010). Uchman (Reference Uchman1999) and Gaillard and Olivero (Reference Gaillard and Olivero2009) proposed that Hormosiroidea should be treated as a junior synonym of Halimedides. However, Hormosiroidea is fundamentally characterized by subspherical chambers, whereas Halimedides exhibit dominantly heart-shaped chambers (Gaillard and Olivero, Reference Gaillard and Olivero2009). In addition, Buatois et al. (Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025) used the nomenclature ‘rosary-shaped trace fossils’ to refer to the ichnotaxon Hormosiroidea meandrica. Despite the Halimedides having priority over Hormosiroidea, their ichnotaxonomy is still a discussion, and many authors have chosen to retain the classification within Hormosiroidea (e.g., Balistieri et al. Reference Balistieri, Netto and Lavina2002, Reference Balistieri, Netto and Sedorko2021; Gandini et al. Reference Gandini, Netto and Souza2007; Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a; Velásquez et al. Reference Velásquez, Silva, Mottin, Alfaro and Vesely2024); and we decided to follow those authors.
Lockeia James, (Reference James1879)
Lockeia siliquaria James, (Reference James1879)
Material: 15 samples preserving 50 specimens (LS-262, LS-263, LS-264, LS-265, LS-267, LS-269, LS-278, LS-280, LS-283, LS-285, LS-288, LS-298, LS-300; Figure 10C-D).
Description: Simple, unbranched mounds, shallow in relation to the bedding plane. Two closely spaced circular mounds are visible in the upper view of the specimens. The diameter of the mounds varies from 1.50 to 2.15 mm, and the distance between them varies from 3.20 to 4.00 mm. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by James (Reference James1879), Lockeia is interpreted as resting galleries (cubichnia) and/or dwelling structures (domichnia) produced by bivalve molluscs (Osgood, Reference Osgood1970; Seilacher and Seilacher, Reference Seilacher and Seilacher1994). However, small crustaceans (e.g., ostracods) should also be considered potential producers of Lockeia (Pollard, Reference Pollard1981; Goldring et al. Reference Goldring, Pollard and Radley2005). Lockeia has been reported in marine and freshwater continental environments since the Cambrian (Melchor et al. Reference Melchor, Cardonatto and Visconti2012; Netto et al. Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a; Pandey et al. Reference Pandey, Uchman, Kumar and Shekhawat2014; Sedorko et al. Reference Sedorko, Netto, Savrda, Assine and Tognoli2017; Memória et al. Reference Memória, Netto, Andrade, Sedorko, Cisneros and Agostinho2021). In LPIA deposits, Lockeia siliquaria is present on estuarine channels and fjord valley deposits of the Carboniferous Itararé Group (Balistieri et al. Reference Balistieri, Netto and Lavina2002, Reference Balistieri, Netto and Sedorko2021; Gandini et al. Reference Gandini, Netto and Souza2007; Netto et al. Reference Netto, Tognoli, Gandini, Lima, Gibert, Netto, Carmona and FMW2012b).
Ichnogenus Palaeophycus Hall (Reference Hall1847)
Palaeophycus tubularis Hall (Reference Hall1847)
Material: Nineteen samples preserving 76 specimens (LS-262, LS-263, LS-265, LS-266, LS-268, LS-269, LS-271, LS-272, LS-273, LS-276, LS-277, LS-278, LS-279, LS-280, LS-282, LS-285, LS-286, LS-296, LS-299; Figures 9E and 10E).
Description: Cylindrical, horizontal, unbranched, rectilinear to moderately sinuous burrows, with smooth wall surface and fill similar to the host rock. The burrow diameters range from 1.00 to 1.55 mm and are up to 10.92 mm in length. Intersections between burrows are occasional. Preservation in positive and negative hyporelief.
Remarks: Originally described by Hall (Reference Hall1847), Palaeophycus tubularis is interpreted as a dwelling structure (domichnia) produced by a predaceous or suspension-feeding organism (Pemberton and Frey, Reference Pemberton and Frey1982; Buatois and Mángano, Reference Buatois and Mángano1993; Sedorko et al. Reference Sedorko, Netto, Savrda, Assine and Tognoli2017). The polychaete Glicera was suggested by Pemberteon and Frey (Reference Pemberton and Frey1982) as a possible tracemaker. Palaeophycus is common in non-marine ichnocoenoses, probably being produced by larval stages of insects and other arthropods (Buatois and Mángano, Reference Buatois and Mángano1993). Palaeophycus is described in rocks from the Ediacaran to the Holocene, being reported in shallow to deep marine (Khalo and Pandey, Reference Khalo and Pandey2018) and continental environments, such as lacustrine (Scott et al. Reference Scott, Buatois, Mángano, Knaust and Bromley2012), alluvial (Padia et al. Reference Padia, Desai, Chauhan, Prizomwala, Chauhan and Thakkar2022), and aeolian settings (Melchor et al. Reference Melchor, Perez and Umazano2024).
Ichnogenus Rusophycus Hall (Reference Hall1852)
Rusophycus carbonarius Dawson (Reference Dawson1864)
Material: Three samples preserving eight specimens (LS-282, LS-297, LS-301; Figure 11A, F).
Description: Bilobate resting burrow with a median groove separating the lobes. The surface of the lobes is wrinkled with some specimens showing ornamentation. The diameter of the lobes varies from 10.85 to 17 mm in length and 3.05 to 5 mm in width, with an internal groove of 0.95 to 2.60 mm (length/width ratio ranging from 3.4 to 3.55). The smaller specimens resemble the shape of a coffee bean. Some specimens are arranged in straight, parallel, V-shaped trails about 60 mm long, characterizing several bilobate resting burrows that repeat across the horizontal plane. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by Hall (Reference Hall1852), Rusophycus is interpreted as a resting burrow (cubichnia) of arthropods (Buatois and Mángano, Reference Buatois and Mángano2003; Seilacher, Reference Seilacher2007). Rusophycus carbonarius is commonly reported in Paleozoic (Buatois and Mángano, Reference Buatois and Mángano2003; Pollard et al. Reference Pollard, Selden and Watts2008; Sedorko et al. Reference Sedorko, Netto, Savrda, Assine and Tognoli2017; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025; Verde et al. Reference Verde, Netto, Azurica, Lavina and Pasquo2022; Buatois et al. Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025) and Mesozoic successions (Melchor et al. Reference Melchor, Cardonatto and Visconti2012; Hminna et al. Reference Hminna, Lagnaoui, Zouheir, Saber and Schneider2020; Zouheir et al. Reference Zouheir, Hminna, Saber, Klein, Lagnaoui, Voigt, Rmich, Schneider and Lucas2023), occurring in marine and continental environments.
Ichnogenus Treptichnus Miller, (Reference Miller1889)
Treptichnus isp.
Material: Five samples preserving seven specimens (LS-266, LS-271, LS-280, LS-298; Figures 9A and 11B).
Description: Straight to slightly curved, horizontal burrows, with zigzag pattern and small nodes at semi-regular spacing. Small branches with a globular texture are observed projecting from the ends of the burows. The distance between the nodes varies from 0.15 to 0.25 mm. The burrow width is up to 0.2 mm, and the length can reach 23 mm. Preservation in sandstone as positive hyporelief.
Remarks: Originally described by Miller (Reference Miller1889), Treptichnus is interpreted as a feeding structure (fodinichnia) produced by vermiform organisms. The specimens described in this study present a well-marked zigzag pattern and nodes along the trace. The analysed specimens have an irregular pattern, and branches projecting beyond the zigzag terminations (Miller, Reference Miller1889). These branching projections commonly have globular textures. Therefore, due to the overlap of characteristics, these specimens will be classified only at the ichnogenus level. Treptichnus is a trace fossil reported in marine and continental environments since the Cambrian (Shahkarami et al. Reference Shahkarami, Mángano and Buatois2017; Kaur et al. Reference Kaur, Singh, Bhargava, Mikuláš, Singla, Prasad and Stopden2021), especially in continental Carboniferous deposits (Buatois et al. Reference Buatois, Labandeira, Mángano, Cohen, Voigt, Mángano and Buatois2016; Marchetti et al. Reference Marchetti, Petti, Zoboli and Pillola2018) and in the glacial deposits of the Itararé Group, Treptichnus pollardi is commonly reported (Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009; Lima et al. Reference Lima, Netto, Corrêa and Lavina2015; Balistieri et al. Reference Balistieri, Netto and Sedorko2021).
Problematica
Material: Three samples preserving 21 specimens (LS-280, LS-286, LS-290; Figure 11C).
Description: Small mounds, subcircular to ovoid, slightly elongated, without ornamentation, with filling similar to that of the host rock. The mounds occur in groups of three or more, having a smooth surface. The diameter of the mounds varies from 0.42 to 0.72 mm in width and up to 1.28 mm in length. Preservation in positive hyporelief.
Remarks: The specimens in this study are similar to Lockeia and with a general organization of Sagitichnus. Furthermore, the morphology is also similar to ice-rafted debris (IRD), such as those presented in the Santa Fé Group by de Barros et al. (Reference de Barros, Carmo, Andrade, Assine and Sedorko2025). Consequently, samples were sectioned to analyse their internal structure. Upon cutting, no grains were present (which would evidence the presence of IRD) displaying a convex morphology filled with sandstone, nor was there any lining of the possible trace (which would be evidence of an open burrow later infilled). Additionally, the presence of an aggregated morphology does not align with the interpretation of IRD deposition, as these particles are not found in such clusters, but rather are dispersed (Eyles and Eyles, Reference Eyles, Eyles, Walker and NP1992, Reference Eyles, Eyles, James and RW2010; Miller, Reference Miller and Reading1996). At the same time, however, there is no evidence of the production of tubes or burrows, which would substantiate the biogenic origin of the structure (see Lewy, Reference Lewy2008; Rindsberg, Reference Rindsberg, Knaust and RG2012; Inglez et al. Reference Inglez, Warren, Okubo, Simões, Quaglio, Arrouy and Netto2019; Saldanha et al. Reference Saldanha, Martins, Osés and Callefo2025). The specimens could alternatively represent the vertical component of Treptichnus isp. on bedding planes, reflecting a preservational variant of this ichnotaxon. However, these traces are not continuous and consist of dispersed structures, precluding assignment to Treptichnus. Due to the millimeter-scale size of the structures and poor preservation hampering ornamentation preservation, this classification remains uncertain. Future work by the research group will seek to address the questions regarding the biogenicity of this structure. Therefore, we decided to name these structures as ‘problematica’.
4.b.2. Trace fossil architectural designs, ethology and diversity
Eight architectural designs are identified in the studied succession, demonstrating differences between the studied ichnofauna within the stratigraphic succession and indicating low ichnotaxonomic richness (ten ichnogenera and eleven ichnospecies) with moderate ichnodisparity (variety of architectural designs). Among these are recognized (see Figure 12): (i) simple horizontal trails (Circulichnis ligusticus, Helminthoidichnites tenuis); (ii) bilobate trails and paired grooves (Cruziana diplopoda, Cruziana problematica, Diplopodichnus biformis); (iii) trackways and scratch imprints (Diplichnites gouldi); (iv) bilaterally symmetrical short, scratched impressions and burrows (Rusophycus carbonarius); (v) passively filled horizontal burrows (Palaeophycus tubularis); (vi) horizontal burrows with horizontal to vertical branches (Treptichnus isp.); (vii) horizontal burrows with serial chambers (Hormosiroidea meandrica); and (viii) isolated and serial oval to almond-shaped burrows (Lockeia siliquaria) (Buatois and Mángano, Reference Buatois and Mángano2013; Buatois et al. Reference Buatois, Wisshak, Wilson and Mángano2017). Regarding the behavioural aspect of the producer, we identified five classes: domichnia (dwelling structure), repichnia (locomotion activity), cubichnia (resting burrows), pascichnia (grazing trace) and fodinichnia (feeding burrows). The ichnotaxa Hormosiroidea meandrica is not represented in Figure 12, because they were recovered from loose samples.
Pie charts showing the described ichnotaxa and the behavioural aspects of the producing specimen. A to F refers to the stratigraphic position of the assemblages in the Ibiracatu section (see Figure 6). Ci: Circulichnis ligusticus, Cd: Cruziana diplopoda, Cp: Cruziana problematica, Dg: Diplichnites gouldi, Db: Diplopodichnus biformis, Ht: Helminthoidichnites tenuis, Lo: Lockeia siliquaria, Pa: Palaeophycus tubularis, Rc: Rusophycus carbonarius, Tr: Treptichnus isp. and Pr: ‘Problematica’ structures. Pr was not included in the pie charts. Lo is interpreted as resting galleries (cubichnia) and/or dwelling structures (domichnia).

Figure 12. Long description
The image contains six sets of pie charts and corresponding diagrams labeled A to F, representing different stratigraphic positions. Each set includes three components: ichnotaxa distribution, behavioral aspects, and the distribution of ichnofauna. The ichnotaxa pie charts display the relative abundance of various ichnotaxa such as Ci, Cd, Cp, Dg, Db, Ht, Lo, Pa, Rc, and Tr. The behavioral pie charts categorize these ichnotaxa into different behavioral aspects like cubichnia, domichnia, repichnia, pascichnia, and fodichnia. The distribution diagrams illustrate the spatial arrangement of these ichnotaxa within the stratigraphic layers. Each diagram shows the top and base of the layers with labeled ichnotaxa. The diagrams also depict the interaction and distribution of these ichnotaxa within the sedimentary layers. The image provides a comprehensive view of the ichnotaxa and their behavioral aspects across different stratigraphic positions.
4.c. U-Pb detrital zircon geochronology
Geochronological analysis was conducted on detrital zircons from a massive sandstone sample collected at the top of the Ibiracatu succession (see Figure 6). We obtained a total of 189 reliable ages (Supplementary Materials, Table S4), acquired after reduction and treatment of the U-Pb isotopic data. The zircons have an anhedral to subhedral shape, are light brown to translucent, and vary in length between 150 μm and 350 μm (Figure 13A). Cathodoluminescence images show that the zircon grains have distinct internal structures, some with well-defined internal zoning, while others are not zoned (Figure 13A). Ages range from 3413 to 834 Ma, with 61% of the ages between 1806 and 2739 Ma (Figure 13B-D). Two zircons with ages of 318 and 314 Ma showed low concordance (see Supplementary Materials, Table S4) and were discarded from the U-Pb geochronological data on detrital zircons. However, because their ages refer to the Carboniferous, these two zircons will be discussed in Section 5.d.
U-Pb geochronological data on detrital zircons for the Ichnofossiliferous succession. (A) Cathodoluminescence images of some zircon grains with analysis points marked, showing the variety of ages, sizes and internal structures. (B) Concordia diagram showing the ages of the zircon populations. (C) Probability density diagram. (D) Pie charts showing the proportion of zircon ages according to the periods/eras in the Chronostratigraphic Table (Cohen et al. Reference Cohen, Finney, Gibbard and Fan2013). The acronyms refer to the possible sources of zircons. AB: Orthogneissic domains of the Archean basement (Machado et al. Reference Machado, Schrank, Abreu, Knauer and Abreu1989; Silva et al. 2002; Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016); LD: Lagoa Dourada Suite (Seixas et al. Reference Seixas, David and Stevenson2012; Teixeira et al. Reference Teixeira, Avila, Dussin, Corrêa-Neto, Bongiolo, Santos and Barbosa2015); RA: Riacian magmatic arcs (Novo et al. Reference Novo, Noce, Pedrosa-Soares and Batista2011; Silva et al. 2002; Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016); SP: Paciência Suite (Silva et al. Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016; Bersan et al. Reference Bersan, Danderfer, Abreu and Lana2018); TA: Taphrogenic and/or anorogenic events from the Statherian to the Stenian (Danderfer et al. Reference Danderfer, DeWaele, Pedreira and Nalini2009; Chemale-Júnior et al. Reference Chemale, Dussin, Alkmim, Martins, Queiroga, Armstrong and Santos2012; Fonte-Boa et al. Reference Fonte-Boa, Novo, Pedrosa-Soares and Dussin2017) and TE: mafic dikes and Tonian anorogenic granites (Machado et al. Reference Machado, Schrank, Abreu, Knauer and Abreu1989; Silva et al. Reference Silva, Pedrosa-Soares, Noce, Delgado, Armstrong and McNaugthon2008; Danderfer et al. Reference Danderfer, DeWaele, Pedreira and Nalini2009).

Figure 13. Long description
The image contains four panels labeled A, B, C, and D. Panel A shows cathodoluminescence images of zircon grains with analysis points marked, displaying a variety of ages, sizes, and internal structures. Panel B presents a Concordia diagram illustrating the ages of the zircon populations. Panel C features a probability density diagram. Panel D includes pie charts showing the proportion of zircon ages according to the periods and eras in the Chronostratigraphic Table. The acronyms refer to possible sources of zircons, including AB, LD, RA, SP, TA, and TE.
5. Discussion
5.a. The Bambuí Group in the Ibiracatu area
The coarsening upward stacking patterns of the Serra de Santa Helena Formation in the studied area are interpreted as a product of wave influence in a shallow waterbody. Wave action is inferred based on the association of siltstone and sandstone with wave ripple cross-lamination, horizontal lamination and local hummocky cross-stratification, which characterizes deposition between storm and fair-weather wave bases. Storm was apparently more frequent at the base of the succession, where fluid escape structures and lenses of hummocky cross-stratification are found (see Figure 3B-3C). The intermediate succession is dominated by cm- to dm-scale alternations between Fl, Sl and Sr (see Figure 3E), which is interpreted to be related to low variation of base level in gentle ramp settings, reflecting deposition in offshore to shoreface-offshore transition settings. Interestingly, desiccation cracks and MISS are verified in association with thicker sandstone beds at the lower and upper parts of the unit, which reflects shallower waters with frequent subaerial exposition, characteristic of upper shoreface areas. The resultant stacking patterns are characterized by progradation and transgressive pattern at the base, retrogradational and transgressive pattern at the middle part, and progradational and regressive pattern at the top.
The Lagoa do Jacaré Formation occurs above the Serra de Santa Helena Formation, although its lower boundary could not be verified in the field. This dominantly carbonate unit is characterized at base by stratified ooid grainstone organized in tabular beds, interpreted as shoreface, ooid bar deposits. The succession briefly deepens upwards (transgressive surface) as verified by the presence of thin marl beds interbedded with grainstone presenting wave ripple cross-lamination, horizontal lamination and hummocky cross-stratification, characterizing deposition in the shoreface-offshore transition with important storm influence in sedimentation (see Figure 4C). Cross-stratified grainstone beds are found above, and are interpreted as shoreface, ooid bar deposits, characterizing progradational stacking pattern. Nodular stromatolites and thrombolites occur above and are considered as the result of deposition in protected settings, probably associated to a lagoon blocked by the ooid bars. Wedge-shaped beds of ooid grainstone with trough cross-stratification occur in the top of the carbonate-dominated succession. Interestingly, evidence of restriction (evaporitic breccias and columnar stromatolites), and subaerial exposition (desiccation cracks) are verified alongside with mixed marl/sandstone/carbonate deposition towards the top of the succession, confirming a progradational and regressive pattern towards the top of the unit.
A transgressive surface is verified above exposition features from the Lagoa do Jacaré Formation. This stratigraphic surface is marked by the occurrence of thick marls and laminated siltstone beds, characteristic of offshore deposition at the lower part of the Serra da Saudade Formation. Thin sandstone beds with horizontal lamination and swaley/hummocky cross-stratification are also verified at the base of the unit, where gypsum pseudomorphs occur, indicating local variations in salinity. The succession shallows upwards, as verified by the dominance of shoreface-offshore transition facies at the intermediate part of the unit and of shoreface facies at the top. The influence of storms was high, with recurrent occurrences of hummocky and swaley cross-stratifications. The shoreface facies (thicker sandstone beds with massive, horizontal lamination and some trough cross-stratifications) were identified in the uppermost 10 m, and possibly are related to a transitional boundary with the Três Marias Formation.
As presented here, animal activity was not verified in the detailed survey conducted in the area. Overall, the described Bambuí succession was dominated by fine-grained terrigenous deposition below storm wave base. High variations in salinity were interpreted in localized portions of all units, particularly in the shallower parts of the systems, but also locally evident in the shoreface-offshore transition. Thus, it is possible that basin restriction impeded animal occupation and thriving, as previously indicated by Warren et al. (Reference Warren, Quaglio, Riccomini, Simões, Poiré, Strikis, Anelli and Strikis2014, Reference Warren, Becker-Kerber, Inglez, Varejão, Morais, Simões, Freitas, Arrouy, Gómez-Peral, Poiré, Okubo, Caxito, Uhlein, Antunes, Lehn, Romero and Fairchild2024), Uhlein et al. (Reference Uhlein, Uhlein, Pereira, Caxito, Okubo, Warren and Sial2019) and Caxito et al. (Reference Caxito, Lana, Frei, Uhlein, Sial, Dantas, Pinto, Campos, Galvão, Warren, Okubo and Ganade2021).
5.b. Ichnofossil-bearing thin-bedded turbidites from the Floresta Formation
The data presented in this work show that the ichnofossil-bearing succession was deposited in a different depositional context than the units of the Bambuí Group (Serra de Santa Helena, Lagoa do Jacaré, and Serra da Saudade formations). While the Bambuí Group units represent sedimentation in a restricted marine system with shoreface to offshore deposits, the succession recorded in Ibiracatu represents turbidite facies developed in a shallow lake system (Figure 14A). The succession is marked by at least four stacking patterns that represent variations in deposition along lobes and outer fringes of turbidites (Figure 14A): (i) Fm to Fl; (ii) Sm to Fl; (iii) Sr to Fl; and (iv) Sl to Fl. These stacking patterns represent characteristic Bouma (Reference Bouma1962) and Mutti (Reference Mutti1992) facies, i.e., Sm = Ta, Sl = Tb, Sr = Tc, Fm = Td, and Fl = Te. Thus, the general stacking pattern, characterized, from base to the top of the succession by (i) Td-Te, Tc-Te, Ta-Te; (ii) Tc-Te, Td-Te, Ta-Te; and (iii) Ta-Te, represents variations in the degree of amalgamation, proximality–distality ratios, and lateral variations of channel-lobe complex. This depositional interpretation conflicts with the ichnological interpretation, and it is discussed later in Section 5.c.
Trace-fossils in the shallow turbiditic lake. (A) Block diagram of the sedimentation model of turbidite facies developed in a shallow lacustrine system described for the study area. (B) Substrate colonization in shallow lacustrine turbidites. Ci: Circulichnis ligusticus, Cd: Cruziana diplopoda, Cp: Cruziana problematica, Dg: Diplichnites gouldi, Db: Diplopodichnus biformis, Ht: Helminthoidichnites tenuis, Ho: Hormosiroidea meandrica, Lo: Lockeia siliquaria, Pa: Palaeophycus tubularis, Rc: Rusophycus carbonarius, Ri: Rusophycus isp., Tr: Treptichnus isp. and Pr: ‘Problematica’ structures.

Figure 14. Long description
The block diagram illustrates the sedimentation model of turbidite facies in a shallow lacustrine system. It shows the distribution of different turbidite facies labeled T1 to T5, with T1 being the most proximal and T5 the most distal. The diagram includes a legend indicating lithostratigraphy and sedimentary facies, such as massive sandstone, laminated sandstone, ripple cross-laminated sand, laminated siltstone, and massive claystone/siltstone. The lower part of the image depicts substrate colonization in shallow lacustrine turbidites, showing various trace fossils like Circulichnis ligusticus, Cruziana diplopoda, Diplichnites gouldi, and others.
The lower Td-Te succession is characteristic of distal portions of the turbidite system, and the upward increase in lower to upper flow regime facies represents the progradation of the lobe system. However, it is important to note that gradual transitions between Bouma facies were not observed, indicating a rapid deposition without bottom current reworking by traction (Shanmugam, Reference Shanmugam2016, Reference Shanmugam2021), followed by slow settling of suspended sediments. The observed groove marks are formed by debris flows, in which flow velocities are slow (Peakall et al. Reference Peakall, Best, Baas, Hodgson, Clare, Talling, Dorrell and Lee2020). The intermediate portion is also characterized by progradation (Tc to Ta), although it represents smaller axial variations, with superposition of several individual turbidite events (Chen et al. Reference Chen, Xian, Li, Wang, Yang, Zhang, Tian, Wu, Liu, Xiang and Wang2025). The sole marks described are formed by turbidity currents, in which flow velocities are fast (Peakall et al. Reference Peakall, Best, Baas, Hodgson, Clare, Talling, Dorrell and Lee2020). The stacking pattern type of the upper succession, with thick Ta-Te pairs and absence of bioturbation and erosion at the base of the facies stackings, indicates rapid deposition in the proximal portions of the turbidity current in a lobe complex with sandy debris flow and bottom-current reworking (Shanmugam, Reference Shanmugam2016, Reference Shanmugam2021). The increase in Sm beds at the top of the unit and the coarsening of cycles towards the upper succession indicate successive turbidite events (Chen et al. Reference Chen, Xian, Li, Wang, Yang, Zhang, Tian, Wu, Liu, Xiang and Wang2025).
Based on the results presented here, it is possible to define that the Ibiracatu succession is representative of the Brejo do Arroz Member, Floresta Formation of the Permian–Carboniferous Santa Fé Group (Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a), part of the LPIA events in Gondwana. While previous works (Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a) describe that the rocks of the Santa Fé Group are preserved in glacial paleovalleys carved in the Bambuí Group, the preservation of this Permo-Carboniferous unit in the Ibiracatu region was conditioned by a system of faults generated at different times in the basin evolution, representing a structural window within an area where the Bambuí Group is mapped. No glacial evidence was found in the field, such as striated pavements, striated clasts and dropstones, among others (see Campos and Dardenne, Reference Campos and Dardenne1994, Reference Campos and Dardenne1997a), though not ruling out the glacial influence in the origin of turbidity currents. Additionally, in relation to the works by Campos and Dardenne (Reference Campos and Dardenne1994) and de Barros et al. (Reference de Barros, Carmo, Andrade, Assine and Sedorko2025), this study shows a greater ichnodiversity for the Floresta Formation (Figure 14B; see discussion below).
The depositional and post-tectonic evolution of the Floresta Formation in the Ibiracatu region is as follows: (i) Generation of U-shaped valleys carved into rocks of the Bambuí Group, formed by the passage of glaciers during the Permian–Carboniferous. The glacial valleys were filled by glaciers, which, after melting, formed glacial lakes. With the deglaciation in steep areas outside the lake, turbidity currents were created, transporting and depositing sediments at the bottom of the glacial lake (Figure 15A). Despite no soft-sediment deformation structures being formed in the area, the presence of glaciation is inferred by regional correlations. Thus, it is possible that the turbidites formed in distal areas; (ii) The deglaciation event probably caused an isostatic readjustment, in which elastic deformation probably occurred. This process is known as post-glacial rebound and can generate or reactivate pre-existing faults (Wood, Reference Wood, Gregersen and Basham1989; Wu et al. Reference Wu, Johnston and Lambeck1999; Stewart et al. Reference Stewart, Sauber and Rose2000; Steffen et al. Reference Steffen, Wu, Steffen and Eaton2014), which reactivated with reverse movement, raising the Ediacaran–Cambrian units over the Carboniferous sediments and locally inverting the stratigraphy (Figure 15B). In the interior portion of Alto de Januária, extensional structures predominate, represented by normal faults with an NNE direction (Piatti, Reference Piatti2021); (iii) In the Early Cretaceous, a crustal stretching related to the development of the South Atlantic caused mechanical subsidence in the Sanfranciscana Basin, generating normal faults and causing the collapse of blocks related to the deposition of the Areado Group (Campos and Dardenne, Reference Campos and Dardenne1997b; Fragoso et al. Reference Fragoso, Uhlein, Sanglard, Suckau, Guerzoni and Faria2011). In the study area, these extensional tectonics probably generated normal faults in the inverted stratigraphy (Figure 15C-D), which occur in a series of hemi-grabens of the Floresta Formation’s siltstone below siltstones and sandstones of the Serra da Saudade Formation.
The depositional and post-tectonic evolution of the Floresta Formation in the Ibiracatu region. (A) Turbidite depositional system of the Floresta Formation deposited in U-shaped valleys carved into rocks of the Bambuí Group. Note the presence of glaciers over the units and preexisting normal faults in the Bambuí Group formations. (B) Deglaciation caused postglacial rebound and reactivated the preexisting normal fault with reverse motion, locally inverting the stratigraphy. (C) Generation of normal faults in the inverted stratigraphy, caused by extensional tectonics in the Early Cretaceous, related to the development of the South Atlantic. (D) Deposit of the Floresta Formation observed today, occurring laterally to the Serra da Saudade Formation with stepped normal faults observed near the boundary with the Cambrian unit, generated in an inverted stratigraphy. To the south, a normal fault contact between the Serra da Saudade and Lagoa do Jacaré formations.

Figure 15. Long description
The diagram illustrates the depositional and post-tectonic evolution of the Floresta Formation in the Ibiracatu region. Panel A shows the turbidite depositional system of the Floresta Formation deposited in U-shaped valleys carved into rocks of the Bambuí Group, with glaciers over the units and preexisting normal faults. Panel B depicts deglaciation causing postglacial rebound and reactivating the preexisting normal fault with reverse motion, locally inverting the stratigraphy. Panel C illustrates the generation of normal faults in the inverted stratigraphy due to extensional tectonics in the Early Cretaceous, related to the development of the South Atlantic. Panel D shows the current deposit of the Floresta Formation, occurring laterally to the Serra da Saudade Formation with stepped normal faults near the boundary with the Cambrian unit, generated in an inverted stratigraphy. To the south, a normal fault contact between the Serra da Saudade and Lagoa do Jacaré formations.
5.c. Benthic colonization of thin bedded turbidites
Other basins that register the LPIA present comparable ichnofossil assemblages to those reported here, such as the Karoo (South Africa), Paganzo and San Rafael (Argentina), Paraná (Brazil), Sydney (Australia) and Tarija (Bolivia), characterized by the presence of arthropod trace fossils and shallow burrows made by worm-like organisms (e.g., Alonso-Muruaga et al. Reference Alonso-Muruaga, Buatois, Mángano and Limarino2012, Reference Alonso-Muruaga, Limarino, Buatois and Pirrone2020; Buatois et al. Reference Buatois, Netto, Mángano and Balistieri2006, Reference Buatois, Netto, Mángano, López-Gamund and Buatois2010, Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025; Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a, Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021; Lima et al. Reference Lima, Netto, Corrêa and Lavina2015, Reference Lima, Minter and Netto2017; Verde et al. Reference Verde, Netto, Azurica, Lavina and Pasquo2022; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025). Indeed, the paleoichnocoenosis present in the studied succession is characterized by the occurrence of trails, resting burrows and dwelling structures produced by arthropods (Cruziana, Diplichnites, Diplopodichnus, Lockeia, Rusophycus), and grazing/feeding traces, produced by vermiform organisms or insect larvae (Circulichnis, Helminthoidichnites, Hormosiroidea, Palaeophycus, Treptichnus). These occurrence aspects characterize the non-marine mixed Scoyenia-Mermia ichnofacies (Buatois and Mángano, Reference Buatois and Mángano1993, Buatois et al. Reference Buatois, Mángano, Maples and Lanier1998b, Reference Buatois, Netto, Mángano, López-Gamund and Buatois2010, Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025; Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025) (Figure 14B).
The Scoyenia–Mermia ichnofacies is commonly attributed to shallow lacustrine basins or high-meltwater influenced marine deposits, with short-scale subaerial exposure of the deposited surfaces (Balistieri, Reference Balistieri2003; Buatois and Mángano, Reference Buatois and Mángano1998, Reference Buatois and Mángano2009; Buatois et al. Reference Buatois, Netto, Mángano, López-Gamund and Buatois2010). Additionally, the presence of Diplichnites gouldi commonly indicates colonization by terrestrial tracemakers (see Netto et al. Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021). This ichnocoenosis may record ephemeral, extremely shallow-waters to damp conditions, permitting colonization of terrestrial organisms. However, the primary mode of deposition and sedimentary structures in the studied section, suggests deposition by turbidity currents in a lacustrine basin. And differently from the deposits of the Bambuí Group, they also lack sedimentological evidence of prolonged subaerial exposure, such as desiccation cracks or pedogenic features (James and Dalrymple, Reference James and Dalrymple2010). This discrepancy of subaerial vs. subaquatic evidence is common in many deposits of the LPIA (see Netto et al. Reference Netto, Balistieri, Lavina and Silveira2009, Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021; Buatois et al. Reference Buatois, Netto, Mángano, López-Gamund and Buatois2010). One hypothesis is a temporal offset between sediment accumulation and subsequent colonization; tracemakers inhabiting a substrate under environmental conditions that differ from those prevailing at the time of deposition (see Rosa et al. Reference Rosa, Isbell, Vesely, Sedorko, Garcia and McNall2024). Rapid drainage events could be one explanation for this incongruence between sedimentary facies and punctual occurrence of terrestrial ichnological signatures (e.g., Huss et al. Reference Huss, Bauder, Werder, Funk and Hock2007; Loso et al. Reference Loso, Anderson, Anderson and Reimer2007; Meghani and Anders, Reference Meghani and Anders2025); such short-lived exposure windows could be sufficient for arthropod colonization (Noll and Netto, Reference Noll and Netto2018; Netto et al. Reference Netto, Corrêa, Lima, Sedorko and Villegas-Martín2021). A model could explain the apparent environmental contradiction by invoking a temporal disconnect between deposition and colonization: following the initial emplacement of sediment by turbidity currents in a relatively deep lacustrine setting (e.g., within a channel-lobe complex), a rapid drop in water level briefly exposed these surfaces, creating a short-lived window of damp or shallow conditions that allowed terrestrial arthropods to colonize the substrate before subsequent rise in water level or a new depositional event buried and preserved the traces.
Glacial environments create stressful conditions for most biota, such as cyclical erosion and sedimentation, reduced salinity due to dilution by glacial meltwater, nutrient-poor environments and low temperatures, causing only adapted and opportunistic organisms to survive in these hostile scenarios (Netto et al. Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a). These environmental characteristics induce a biota with low ichnodisparity, low ichnodiversity and reduced size (Netto et al. Reference Netto, Benner, Buatois, Uchman, Mángano, Ridge, Kazakauskas, Gaigalas, Knaust and Bromley2012a; de Barros et al. Reference de Barros, Becker-Kerber, Sedorko, Lima and Pacheco2021, Reference de Barros, Carmo, Andrade, Assine and Sedorko2025), demonstrating a colonization of strategist organisms, adapted to the glacial environment. The ichnoassemblage from the Floresta Formation are characterized by low ichnodiversity (ten ichnogenera and eleven ichnospecies) with moderate ichnodisparity (eight architectural designs). These characteristics may indicate that substrate colonization was affected by different sedimentation rates originating from turbidity flows, accommodation change, and oxygenation levels.
Prolonged intervals with settling of suspended sediments must have occurred between turbidite events, favouring the colonization of organisms with different behaviours over time (see Figure 12). In this scenario, Diplopodichnus biformis and Diplichnites gouldi are epistratal structures recording different stages of locomotion and proximity of the body plan to the substrate (Keighley and Pickerill, Reference Keighley and Pickerill1996). Cruziana diplopoda and Cruziana problematica were probably produced by the same tracemaker as Diplopodichnus biformis and Diplichnites gouldi, with the difference that the producer penetrated deeper into the substrate (Keighley and Pickerill, Reference Keighley and Pickerill1996; Schatz et al. Reference Schatz, Mángano, Buatois and Limarino2011). The low occurrence of Rusophycus carbonarius in the studied ichnoassembly may indicate a high rate of locomotion in search of food and few rest breaks.
Turbidity currents are responsible for oxygenating the waters at the bottom of the basin and providing suspended particles, enabling the colonization of organisms and playing an important role in environmental dynamics (Buatois and Mángano, Reference Buatois and Mángano2013; Zheng and Cao, Reference Zheng and Cao2023). The presence of grazing traces (pascichnia) and feeding burrows (fodinichnia) indicates that freshly deposited turbidites provided nutrient-rich substrates that supported rapid and opportunistic colonization under oxygenated conditions. Opportunistic colonizers likely responded quickly to short-lived windows of ecological stability following turbidity events.
Therefore, substrate colonization was directly influenced by turbidity flows, as well-documented by studies of trace fossils in lacustrine to fjord turbidite successions of the LPIA in Argentina (Alonso-Muruaga et al. Reference Alonso-Muruaga, Buatois, Mángano and Limarino2012, Reference Alonso-Muruaga, Limarino, Buatois and Pirrone2020; Buatois and Mángano, Reference Buatois and Mángano1993, Reference Buatois and Mángano2003; Buatois and del Papa, Reference Buatois and del Papa2003). The reported association is indicative of different levels of mixing according to the position within the turbidite lobe (from T1 to T5; Figure 14A and 14B). The abundance of dominichia and cubichnia is associated with a greater occurrence of T1-T5 cycles, at the edges of the turbidity lobes, where the environment is calmer and has higher amounts of siltstone. Meanwhile, the dominance of repichnia is concentrated in the inner part of the lobes, associated with the greater presence of T1-T3 cycles, indicating shallower settings. This assemblage can be attributed to the Diplopodichnus association (Buatois et al. Reference Buatois, Alonso-Muruaga, Mángano, Luo and Shi2025).
5.d. The regional extent of the Carboniferous glaciation in Gondwana
This study presented the first U-Pb geochronological analysis of detrital zircon grains for the Santa Fé Group, providing data on the sedimentary provenance of the unit. The concordance, pie and probability density plots of U-Pb zircon ages (Figure 13B–D), show a polymodal pattern with different ages ranging from the Paleoarchean to the Tonian. Therefore, Precambrian ages comprise 100% of the entire set of dated detrital grains.
Paleoarchean zircon populations (17 grains) range from 3413 to 3203 Ma, corresponding to 9% of the entire set of dated grains. Mesoarchean grain populations (26 grains) range between 3196 and 2858 Ma, representing 14% of the entire set of dated grains. Neoarchean grains (39 grains) record ages between 2739 and 2514 Ma, representing 21% of all grains analysed.
Paleoproterozoic zircon populations (84 grains) account for 43% of the entire dated grain set, being Siderian (12 grains; 2491-2316 Ma; 6%), Rhyacian (27 grains; 2277-2050 Ma; 14%), Orosirian (37 grains; 2048-1806 Ma; 20%) and Statherian (8 grains; 1784-1738 Ma; 4%). Mesoproterozoic zircon populations (14 grains) account for 7% of the dated zircon set, being Calymmian (8 grains; 1599-1404 Ma; 4%), Ectasian (2 grains; 1283-1208 Ma; 1%) and Stenian (4 grains; 1178-1165 Ma; 2%). Neoproterozoic zircon populations (9 grains) range between 984 and 834 Ma (Tonian), representing 5% of the entire set of dated grains. Phanerozoic populations are rare among the dated detrital zircon grains, with ages restricted to the Carboniferous, including the Bashkirian (1 grain; 318 Ma) and Moscovian (1 grain; 314 Ma). However, these Carboniferous-aged grains presented high common Pb values and low concordance and were therefore not included in the analysis.
Therefore, Archean and Proterozoic populations correspond to 44% and 56%, respectively, of the entire set of dated and analysed detrital grains, with a main age peak at ∼2.1 Ga and a secondary one at ∼2.6 Ga, in addition to other minor peaks. The polymodal age distribution, containing larger proportions of older ages, reflects the history of the underlying basement, a characteristic feature of intracratonic basins (Cawood et al. Reference Cawood, Hawkesworth and Dhuime2012). The predominance of Precambrian ages suggests that most of the zircons are inherited from different orogenic events throughout the evolution of the São Francisco Craton.
It is important to highlight that recent reconstructions of the Neopaleozoic glaciation show ice flow directions with a preferential NW orientation in the LPIA basins of Gondwana, especially in the eastern portions of South America (Paraná and Chaco Paraná basins) and western Africa (Congo, Kalahari and Karoo basins) (Isbell et al. Reference Isbell, Henry, Gulbranson, Limarino, Fraiser, Koch, Ciccioli and Dineen2012) (Figure 16A). Furthermore, cross-laminations and sole marks in the Santa Fé Group outcrop in Ibiracatu studied in this work indicate NW flow directions. However, data on the ice flow directions of glaciers during the Permian–Carboniferous glaciation in the Sanfranciscana Basin are still scarce. Therefore, based on the flow directions of the Neopaleozoic glaciers during Gondwana, we infer that the Araçuaí Belt was the main source for the zircon population (Figure 16B). However, based on the observation of paleocurrents from striated pavements, the optical behaviour of detrital quartz, and the association of heavy minerals and dropstones in the glacial-lacustrine facies, Campos and Dardenne (Reference Campos and Dardenne1994) interpreted the directions of ice flow of glaciers from NE to SW. Therefore, it is possible that source areas that were further north contributed to the sediment supply during the deposition of the Santa Fé Group (Figure 16C). Further studies are needed to define the directions of ice flow of glaciers and the respective sediment sources.
Reconstruction of maximum glaciation during the Late Paleozoic Ice Age and possible ice flows directions to the Floresta Formation. (A) Reconstruction of maximum glaciation during the Late Paleozoic Ice Age based on recent ice flow direction data. Gondwana basins (C: Congo; CP: Chaco Paraná; Kl: Kalahari; Kr: Karoo and P: Paraná). AW: Araçuaí-West Congo orogen. Modified from Isbell et al. (Reference Isbell, Henry, Gulbranson, Limarino, Fraiser, Koch, Ciccioli and Dineen2012). (B) Simplified geological map highlighting the Archean-Orosirian basement units of the southern São Francisco craton (SFC) and Araçuaí orogen, and their location in Western Gondwana (A: Amazonian craton; AW: Araçuaí-West Congo orogen; C: Congo craton; KA: Kalahari craton; P-LA-RLP: Paranapanema, Luiz Alves and Rio de La Plata cratonic blocks; WA: West Africa craton). Modified from Silva et al. (Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016). (C) Geological map of the Sanfranciscana Basin in the northwestern portion of the State of Minas Gerais with possible ice flow directions.

Figure 16. Long description
The map illustrates the reconstruction of maximum glaciation during the Late Paleozoic Ice Age, highlighting possible ice flow directions. It includes three sub-maps: (A) a global view of Gondwana with labeled basins and ice flow directions, (B) a simplified geological map of the southern São Francisco craton and Araçuaí orogen, and (C) a detailed geological map of the Sanfranciscana Basin in the northwestern portion of the State of Minas Gerais, indicating possible ice flow directions.
The Araçuaí Belt is composed of Archean/Paleoproterozoic rocks, consisting of TTG gneisses and migmatites (tonalite-trondhjemite-granodiorite), granitic plutons and greenstone belt-type sequences (Noce et al. Reference Noce, Pedrosa-Soares, Silva, Armstrong and Piuzana2007; Silva et al. Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016). The oldest zircons, with ages between 3413 Ma and 2514 Ma, may be derived directly from the Archean basement, represented by the Porteirinha, Guanhães and Gouveia orthogneissic domains (Machado et al. Reference Machado, Schrank, Abreu, Knauer and Abreu1989; Silva et al. Reference Silva, Armstrong, Noce, Carneiro, Pimentel, Pedrosa-Soares, Leite, Vieira, Silva, Paes and Cardoso Filho2002, Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016).
Possible Paleo-Mesoproterozoic sources of zircons are the following: (i) recycled zircons from the Mineiro Belt, including Siderian orthogneisses, dated between 2450 and 2410 Ma (Barbosa et al. Reference Barbosa, Teixeira, Leal and Leal2013), and biotite metatrondhjemite, orthogneisses and hornblende metatonalite from the Lagoa Dourada Suite from 2356 to 2328 Ma (Seixas et al. Reference Seixas, David and Stevenson2012; Teixeira et al. Reference Teixeira, Avila, Dussin, Corrêa-Neto, Bongiolo, Santos and Barbosa2015); (ii) Rhyacian magmatic arcs of the Juíz de Fora and Mantiqueira complexes, dated at 2195 to 2107 Ma and 2137 to 2018 Ma, respectively. In addition to the Caparaó-Pocrane magmatic arcs, with crystallization ages around 2200 Ma (Novo et al. Reference Novo, Noce, Pedrosa-Soares and Batista2011; Silva et al. Reference Silva, Armstrong, Noce, Carneiro, Pimentel, Pedrosa-Soares, Leite, Vieira, Silva, Paes and Cardoso Filho2002, Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016); (iii) Orosirian alkaline intrusions, including syenites and plutonites of the Paciência Suite, dated between 2053 and 2039 Ma (Silva et al. Reference Silva, Pedrosa-Soares, Armstrong, Pinto, Reis Magalhães, Pinheiro and Santos2016; Bersan et al. Reference Bersan, Danderfer, Abreu and Lana2018); and (iv) Taphrogenic and/or anorogenic events that preceded the uplift of the Araçuaí-West Congo orogen, related to the development of the Espinhaço Supergroup and Chapada Diamantina Group, encompassing rocks from the Statherian to the Stenian, dated between 1750 and 1096 Ma (Brito-Neves et al. Reference Brito-Neves, Sá, Nilson and Botelho1995; Danderfer et al. Reference Danderfer, DeWaele, Pedreira and Nalini2009; Pedrosa-Soares and Alkmim, Reference Pedrosa-Soares and Alkmim2011; Chemale-Júnior et al. Reference Chemale, Dussin, Alkmim, Martins, Queiroga, Armstrong and Santos2012; Fonte-Boa et al. Reference Fonte-Boa, Novo, Pedrosa-Soares and Dussin2017).
Possible sources of Tonian zircons are associated with mafic dikes and anorogenic granites (Pedrosa-Soares and Alkmim, Reference Pedrosa-Soares and Alkmim2011), among them: (i) swarm of mafic dikes of the Pedro Lessa Suite, with ages of 906 Ma, related to the Southern Espinhaço Mountain Range (Machado et al. Reference Machado, Schrank, Abreu, Knauer and Abreu1989); (ii) basic dikes dated at 850 Ma, associated to the Northern Espinhaço Mountain Range (Danderfer et al. Reference Danderfer, DeWaele, Pedreira and Nalini2009); and (iii) anorogenic granites of the Salto da Divisa Suite, with ages around 880 Ma (Silva et al. Reference Silva, Pedrosa-Soares, Noce, Delgado, Armstrong and McNaugthon2008).
The Carboniferous zircons (318 and 314 Ma) are probably associated to post-glacial rebound processes. With deglaciation and isostatic readjustment, deformation and faulting occurred, facilitating the percolation of fluids through grain fractures and altering the magmatic zircons. Dias et al. (Reference Dias, Martins-Ferreira, Pereira, Sales and Chemale2024), in a geochronological study on the Phanerozoic evolution of the São Francisco Craton, found detrital zircons with ages ranging from 330 to 280 Ma and interpreted them as a consequence of the ice retreat and isostatic rebound interior of the Gondwana Paleocontinent. Additionally, fluid (meteoric, ice melt and metamorphic) percolation may enrich zircon grains in U, Th and Pb common (Breeding et al. Reference Breeding, Ague, Grove and Rupke2004). Thus, it is difficult to determine the possible sources of the Carboniferous-aged zircons dated in this work, since they probably come from the post-glacial rebound reworking of pre-existing zircons.
Carboniferous-age zircons were discarded from the U-Pb geochronological data on detrital zircons, and the maximum depositional age of the Santa Fé Group was calculated at 834 ± 12 Ma, referring to the youngest detrital zircon among the entire grain set, too old for unity. However, the data presented in this work show that the Santa Fé Group was deposited in a different depositional context than the units of the Bambuí Group in Ibiracatu. Furthermore, the ichnofossil assemblage is similar to other LPIA basins. Therefore, our data provides certainty that the ichnofossil succession in Ibiracatu is indeed the Floresta Formation, Santa Fé Group, as defined by Campos and Dardenne (Reference Campos and Dardenne1994).
6. Conclusion
The sedimentological, ichnological and geochronological data acquired and presented in this work provide new insights into the depositional evolution, benthic colonization, and age of the Floresta Formation, Santa Fé Group, in the Ibiracatu region of Minas Gerais. The Floresta Formation consists of turbidite facies developed in a shallow lacustrine system, presenting a different depositional context than the Bambuí Group units, which represent sedimentation in a restricted marine system. The depositional evolution and preservation of the Floresta Formation in the Ibiracatu region was marked by a system of faults generated at different times in the evolution of the Sanfranciscana Basin. The paleoichnocoenosis of the Floresta Formation is dominated by the activity of arthropods, annelids, worm-like organisms, and/or insect larvae. Eleven ichnotaxa (Circulichnis ligusticus, Cruziana diplopoda, Cruziana problematica, Diplichnites gouldi, Diplopodichnus biformis, Helminthoidichnites tenuis, Hormosiroidea meandrica, Lockeia siliquaria, Palaeophycus tubularis, Rusophycus carbonarius, and Treptichnus isp.) in addition to a problematic structure were identified. This paleoichnocoenosis is characteristic of the mixed Scoyenia-Mermia ichnofacies, commonly found in other basins that record the LPIA. The low ichnodiversity, moderate ichnodisparity, and different morphological plans indicate that substrate colonization was directly influenced by turbidity fluxes. The abundance of Dominichia and Cubichnia is concentrated at the edges of the turbidity lobes, associated to low sedimentation rates. Meanwhile, Repichnia dominates the inner part of the lobes, in the region where water-level fluctuations can induce a shallower setting. Regarding geochronology, the polymodal spectrum with a predominance of Precambrian ages indicates that a large portion of the zircons is inherited from different orogenic events throughout the evolution of the São Francisco Craton. Based on the flow directions of Neo-Paleozoic glaciers during Gondwana, the Araçuaí Belt is the main source of detrital zircons. The zircons dated from the Carboniferous are probably associated with post-glacial rebound processes, in which deglaciation and isostatic readjustment facilitated the percolation of fluids through fractures in the grains, altering the magmatic zircons.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0016756826100739
Acknowledgements
We would like to thank Editor Emese Bordy and the reviewers Luis A. Buatois, Mariano Verde and Renata G. Netto, as well as two anonymous reviewers, for their valuable comments, which significantly improved the manuscript. LMC thanks the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for a Master’s scholarship and the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) for a doctoral scholarship, as well as the PPGECRN (UFOP) for infrastructure and logistical support through the research funding projects FAPEMIG (APQ-02529-24) and FINEP (1340/24). FGV is a research fellow of CNPq (304881/2024-4). GEBdeB thanks the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the doctorate grant Finance Code 001 (88887.799772/2022-00) and Sandwich Doctorate Program (PDSE) grant (88881.980843/2024-01). The authors acknowledge the Brazilian Geological Survey for providing data and location of the studied material, and Gabriel Jubé Uhlein and Lucas Inglez dos Reis for their insightful comments on the article.
Author contributions
LMC: conceptualization (equal), data curation (lead), formal analysis (lead), investigation (equal), methodology (equal), writing – original draft (lead); FGV: conceptualization (equal), formal analysis (supporting), investigation (equal), methodology (equal), project administration (lead), writing – original draft (supporting); GEBdeB: formal analysis (supporting), writing – original draft (supporting); MGR: formal analysis (supporting), investigation (equal), writing – original draft (supporting); JBM: formal analysis (supporting), investigation (equal), writing – original draft (supporting); IDR: formal analysis (supporting), writing – original draft (supporting); CCL: formal analysis (supporting), writing – original draft (supporting); DCB: formal analysis (supporting), writing – original draft (supporting); LBG: formal analysis (supporting), writing – original draft (supporting).
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.


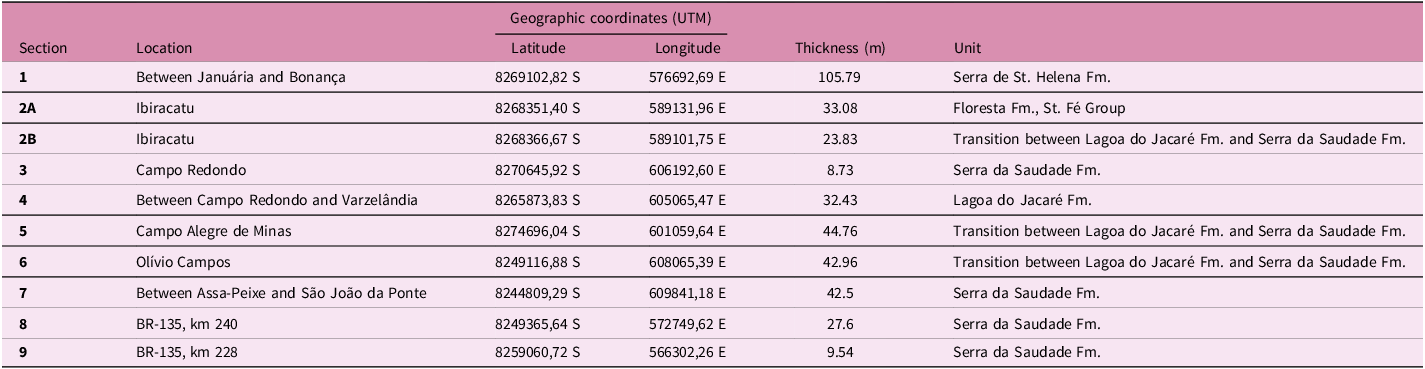





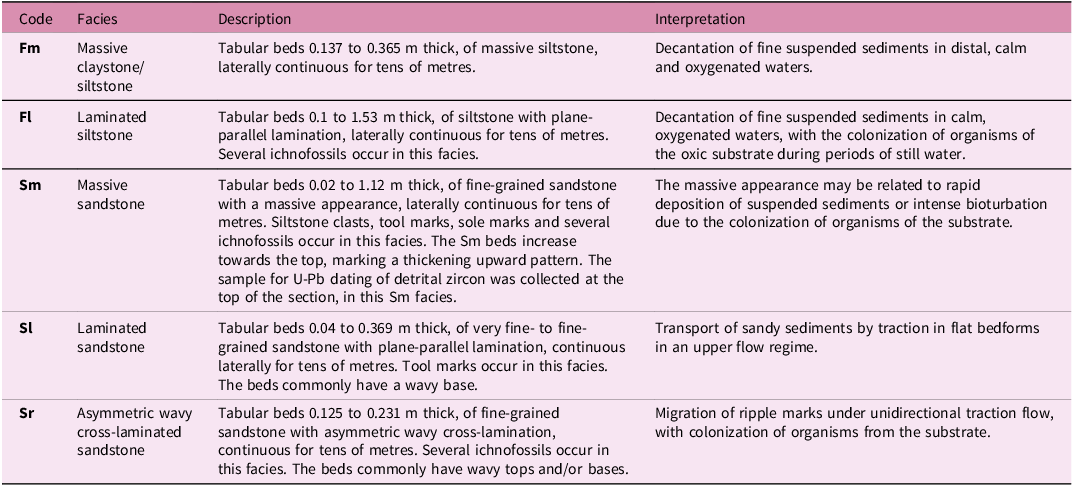












 Open access
Open access