


Impact statement
Interactions among long-term weather factors, vegetation characteristics and soils present numerous permutations for soil organic carbon (SOC) and nitrogen (N) dynamics in semiarid rangeland ecosystems. As a result, assessing livestock grazing impacts on SOC and N are difficult to categorize given this complexity and site specificity of conditions. Moreover, long-term studies in semiarid rangelands with consistent grazing treatments and repeated soil samplings are lacking to understand the influences of grazing with respect to environmental influences, such as directional trends in long-term weather factors. This longitudinal study capitalizes on a multi-decadal grazing experiment in the semiarid North American Great Plains, northern mixed-grass prairie, with grazing season (June-late September) grazing treatments of season-long light and heavy stocking rates, and exclosure (non-grazed) established in 1982 and soils sampled in 1993, 2003 and 2013. Increasing grazing intensity from non-grazed to heavy grazed, reduced annual aboveground standing crop biomass and shifted the plant community dominance from cool-season (C3) to warm-season (C4) perennial grasses but did not affect SOC and N in the top 30 cm of soils with the exception of soil N in the upper soil depth (0-5) cm higher with season-long light grazing. Soil dynamic findings demonstrate that concentrations of SOC and N decreased over 20 years (from 1993 to 2013) irrespective of grazing treatments (ranging from no grazing to light to heavy stocking) for the top 30 cm. Long-term weather factors, such as an observed decreasing trend in precipitation but no temperature trends, may impact SOC and N dynamics more than livestock grazing, suggesting that expectations of increased SOC and N with livestock grazing are not reasonable in semiarid rangeland ecosystems of the North American Great Plains with a long evolutionary grazing history.
Introduction
Livestock grazing is a primary land use in drylands (Wang et al., Reference Wang, Jiao, MacBean, Rulli, Manzoni, Vico and D’Odorico2022) with benefits to the provision of numerous ecosystem goods (e.g., forage and livestock production) and services (e.g., soil carbon storage, biodiversity) (Sanderson et al., Reference Sanderson, Beutler, Brown, Burke, Chapman, Conant, Derner, Easter, Fuhlendorf, Grissom, Herrick, Liptzin, Morgan, Murph, Pague, Rangwala, Ray, Rondeau, Schulz and Sullivan2020). Adapting grazing management to biotic attributes such as soils, plant communities and climatic variability, including extremes like deluges and drought, at local scales can result in potential benefits for ecosystem structure, function and resilience (Maestre et al., Reference Maestre, Eldridge, Soliveres, Kéfi, Delgado-Baquerizo, Bowker, García-Palacios, Gaitán, Gallardo, Lázaro and Berdugo2016; Copeland et al., Reference Copeland, Hoover, Augustine, Bates, Boyd, Davies, Derner, Duniway, Porensky and Vermeire2023; Willard et al., Reference Willard, Gennet, Anderson, Knapp, Lavallee, Ritten, Feldman, Hulvey, Nasto, Jacobs and Watts2025). Livestock grazing can contribute to increased carbon sequestration in drylands, but these impacts are context-dependent due to unique ecosystem responses and climate–soil–plant interactions (Hanan et al., Reference Hanan, Milne, Aynekulu, Yu and Anchang2021; Maestre et al., Reference Maestre, Le Bagousse-Pinguet, Delgado-Baquerizo, Eldridge, Saiz, Berdugo, Gozalo, Ochoa, Guirado, García-Gómez and Valencia2022).
Drylands play a critical role in the earth system due to their large land area (>50% of the terrestrial surface) and socioeconomic and ecological importance, including 30% of the global carbon in above- and belowground biomass and soils (Hanan et al., Reference Hanan, Milne, Aynekulu, Yu and Anchang2021). Half to two-thirds of the earth’s drylands is either already degraded or highly susceptible to degradation, primarily through aeolian and hydrologic erosion (Lal, Reference Lal2019). Proper stocking rate is a recommended practice of managing hot spots in degraded drylands (Sanderman et al., Reference Sanderman, Hengl and Fiske2017) to resequester lost soil carbon (C) (Lal, Reference Lal2004).
Measuring concentrations and changes in soil C and nitrogen (N) in drylands is complicated for numerous reasons, including 1) presence of fertile islands that present substantial small scale heterogeneity (Vinton and Burke, Reference Vinton and Burke1995; Derner et al., Reference Derner, Briske and Boutton1997; Ding and Eldridge, Reference Ding and Eldridge2021), 2) thousands of years of legacy grazing by native herbivores and more recently domesticated livestock have established an evolutionary history of grazing that can mask effects of grazing management treatments (Milchunas et al., Reference Milchunas, Sala and Lauenroth1988), 3) these water limited ecosystems with low plant productivity and young developing soils are substantially influenced by inter- and intra-annual variability in precipitation inputs and pulses of biogeochemical cycling (Klausmeier, Reference Klausmeier1999; Ahlström et al., Reference Ahlström, Raupach, Schurgers, Smith, Arneth, Jung, Reichstein, Canadell, Friedlingstein, Jain and Kato2015; Smith et al., Reference Smith, Cotrufo, Rumpel, Paustian, Kuikman, Elliott, McDowell, Griffiths, Asakawa, Bustamante and House2015; Augusto et al., Reference Augusto, Achat, Jonard, Vidal and Ringeval2017), 4) reported C sequestration rates in these systems have been small relative to the pool sizes (Derner and Schuman, Reference Derner and Schuman2007; McKenna et al., Reference McKenna, Grams, Barasha, Antoninka and Johnson2022) and 5) livestock grazing can induce changes in the plant community from one dominant plant functional group to another. For example, in northern mixed-grass prairie of the North American Great Plains, heavy grazing intensity during the growing season (early June through September) changes vegetation from C3 perennial grass-dominated to C4 perennial grass-dominated over decadal scales (Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016). Associated with this plant community shift, heavy grazing reduces aboveground productivity by about 20% (Derner and Hart, Reference Derner and Hart2007), thereby reducing inputs of C and N to soils. However, increases in belowground productivity due to resource allocation shifts in C4 perennial grasses (Frank et al., Reference Frank, Tanaka, Hofmann and Follett1995), could offset lowered aboveground inputs to result in an outcome of limited to no net changes in soil C and N.
Syntheses and meta-analyses of livestock grazing influences on soil C in drylands have demonstrated high variability regarding the effects of grazing (Milchunas and Lauenroth, Reference Milchunas and Lauenroth1993; Derner and Schuman, Reference Derner and Schuman2007; McSherry and Ritchie, Reference McSherry and Ritchie2013; Conant et al., Reference Conant, Cerri, Osborne and Paustian2017; Bai and Cotrufo, Reference Bai and Cotrufo2022). This variability can be attributed to limited treatment replications, context-specificity of the site and local-to-regional drivers of change (Jordon et al., Reference Jordon, Buffet, Dungait, Galdos, Garnett, Lee, Lynch, Röös, Searchinger, Smith and Godfray2024), and spatial variation in soil texture, precipitation, vegetation type, grazing intensity, study duration and sampling depth (McSherry and Ritchie, Reference McSherry and Ritchie2013). Furthermore, grazing-induced changes to plant communities and productivity, typically used to predict soil organic C (SOC) change over time, work poorly in dryland, semiarid rangeland ecosystems (Reinhart et al., Reference Reinhart, Rinella, Waterman, Worogo and Vermeire2024; but see Liang et al., Reference Liang, Smith, Chen, Wu, Guo, Gornish and Liang2021). In contrast, for mesic environments with prior management (e.g., tillage, overgrazing) that degraded SOC, establishment of perennial vegetation (e.g., planted pastures) and livestock grazing increases SOC (Conant and Paustian, Reference Conant and Paustian2002; Conant et al., Reference Conant, Cerri, Osborne and Paustian2017). Despite these complications, measuring soil C and N is critical to understanding management effects on soil C, as it comprises over 90% of the total ecosystem C in low productivity ecosystems like semiarid rangeland ecosystems (Derner et al., Reference Derner, Boutton and Briske2006).
Lacking in the literature are long-term grazing studies in drylands with longitudinal soil sampling events to help in assessing livestock grazing management effects over time. Here, we capitalize on one of the few multi-decadal grazing experiments in North America with long-term continuous season-long (June-late September) grazing treatments of light and heavy intensity, along with exclusion (no grazing) (Hart et al., Reference Hart, Samuel, Test and Smith1988), and controls for soil texture (loamy), precipitation (local), study duration (since 1982) and sampling depth (0–60 cm, increments of 0–5, 5–15, 15–30 and 30–60). A moderate grazing intensity treatment was included in the experimental design for vegetation and livestock responses (Hart et al., Reference Hart, Samuel, Test and Smith1988), but this grazing intensity was not soil sampled throughout the experiment. Prior soil sampling events addressed the first decade (1982–1992, Schuman et al., Reference Schuman, Reeder, Manley, Hart and Manley1999) and second decade (1993–2003, Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008) of grazing influences on SOC and soil N. Here, we assess the full study period (1982–2013) with comparison of grazing influence on SOC and N across time (from 1993 to 2013). We predicted that long-term grazing will result in lower SOC and N concentrations as grazing intensity increases from none to light to heavy due to decreasing aboveground C inputs.
Methods
Site description
This experiment was conducted in the North American northern mixed-grass prairie, a dryland rangeland ecosystem at the United States Department of Agriculture – Agricultural Research Service (USDA-ARS) High Plains Grasslands Research Station near Cheyenne, Wyoming, USA (41°11′N, 104°53′W). Long-term (>100 years) mean annual precipitation for this site is 381 mm, and most of the rainfall occurs in late spring/early summer. Dominant native vegetation is a mix of C3-C4 perennial grasses, including C4 blue grama (Bouteloua gracilis) and C3 graminoids western wheatgrass (Pascopyrum smithii), needleandthread (Hesperostipa comata) and needleleaf sedge (Carex duriscula) (Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016).
Experimental design
Fixed stocking rates, initiated in 1982 on land that experienced very light grazing by various forms of livestock and wildlife in the past (Hart et al., Reference Hart, Samuel, Test and Smith1988), included replicate pastures of gently rolling topography each of light (15.7 ± 2.8 animal unit days per hectare [AUD ha−1]; mean ± 1SD; or 15 yearlings per 82 ha) and heavy (43.4 ± 7.3 AUD ha−1 or 4 yearlings per 9 ha) grazing intensities. One AUD equals one 454 kg animal for 1 day. These stocking rates were established to be ~35% below and ~33% above stocking rates recommended by the USDA-Natural Resource Conservation Service (Hart et al., Reference Hart, Samuel, Test and Smith1988). Two 0.5 ha exclosures were fenced in 1982 to prevent livestock grazing, but not wildlife or small mammal access. Yearling cattle grazed the light and heavy stocking rate treatments from early June to late September, about 115 days, each year. Tillers of the dominant C3 perennial grass western wheatgrass and C4 grass blue grama were grazed more frequently with heavy grazing (Hart et al., Reference Hart, Clapp and Test1993). For additional details, see Hart et al. (Reference Hart, Samuel, Test and Smith1988) and Derner and Hart (Reference Derner and Hart2007).
Aboveground standing crop biomass
To quantify aboveground standing crop biomass, three 1.5 m2 wire cages to prevent livestock grazing for that growing season were randomly located on a permanently marked 50-m transect in a flat topographical position in each pasture. Soils at these topographic positions are well drained, coarse loams comprised mainly of Albinas, Ascalon and Altvan loams (mixed mesic Aridic Argiustolls) (Stevenson et al., Reference Stevenson, Baumgartner and Schuman1984). The wire cages were moved a random distance in a random cardinal direction each spring prior to initiation of grazing treatments but remained on the flat topographical positions. A 0.18 m2 quadrat was clipped to the ground level in the middle of each wire cage at peak standing crop, mid-July. Current-year biomass production was separated into plant functional groups: C3PG (cool-season perennial graminoids), C4PG (warm-season perennial grasses), C3AG (cool-season annual grasses), forbs, and sub-shrubs and dried at 60°C and weighed. We combined C3PG, C3AG, forbs and subshrubs into a C3 group and the C4PG represented the C4 group. For additional details, see Derner and Hart (Reference Derner and Hart2007).
Soil sampling and analyses
Soil samples were taken in early May 2013 from each permanently marked 50-m transect for the aboveground standing crop biomass in each replicate pasture and exclosures where prior sampling events occurred in 1993 (Schuman et al., Reference Schuman, Reeder, Manley, Hart and Manley1999) and 2003 (Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008). Soil moisture is adequate in May coinciding with the peak in precipitation to obtain representative soil samples. Dry soil conditions in mid-July preclude timing of soil sampling with the aboveground standing crop biomass harvests. As previously sampled, two soil cores of 4-cm diameter to 60 cm depth were taken at the 0, 10, 20, 30, and 40 m locations on each transect using a hydraulic soil sampler (Giddings Machine Company, Windsor, Colorado, USA). Each core was separated with a knife into 0–5, 5–15, 15–30 and 30–60 cm increments. Increments were composited across the cores in each transect to reduce variability. All soil samples were air-dried and 2-mm sieved to remove plant crowns, visible roots and root fragments as in the prior soil sampling events (Schuman et al., Reference Schuman, Reeder, Manley, Hart and Manley1999; Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008).
A 100-g subsample was taken from each replicated grazing treatment and depth increment for air elutriation to remove fine root material. These subsamples were finely ground, carbonates were removed by acid (HCl) fumigation prior to analysis (Stewart et al., Reference Stewart, Roosendaal, Sindelar, Pruessner, Jin and Schmer2019) and analyzed by combustion for SOC and total soil N on a Europa Scientific C/N analyzer (Europa Scientific Ltd., England). For the 1993 soil sampling, SOC was determined using the Walkley-Black dichromate procedure and soil N with a modified micro-Kjeldahl procedure (Schuman et al., Reference Schuman, Reeder, Manley, Hart and Manley1999). For the 2003 soil sampling, total soil C and N were determined using a Carlo-Erba C/N analyzer (Carlo-Erba Instruments, Italy) with inorganic C, determined using a modified pressure-calcimeter procedure (Sherrod et al., Reference Sherrod, Dunn, Peterson and Kolberg2002), subtracted from the total soil C values to obtain SOC (Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008). Concentration of C and N do not differ between dry combustion and dichromate oxidation or micro-Kjeldahl methods (Kerven et al., Reference Kerven, Menzies and Geyer2000; Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008). For all soil sampling events, SOC and N are expressed on an oven-dried basis (55°C).
Statistical analysis
Concentrations were used as the response metric rather than stocks of SOC and N as bulk density is differentially affected by grazing intensity (Van Haveren, Reference Van Haveren1983; Tate et al., Reference Tate, Dudley, McDougald and George2004) thereby providing a confounding factor when using stocks in analysis. For SOC and N concentrations, mixed linear models were fitted for each soil depth (averaged across the five location data points on each transect) using the lmer function from the lme4 package (version 3.1–152) (Bates et al., Reference Bates, Maechler, Bolker and Walker2015 in R version 4.2.2; R Core Team, 2022). Fixed effects included the interaction of year and grazing treatment, while pasture was set as a random effect. To meet assumptions of normality and homogeneity of variance, log transformations were used for SOC and N concentrations at every soil depth. Standard errors and estimated marginal means (Lenth, Reference Lenth2021) were derived from linear models. The function r.squaredGLMM in the ‘MuMIn’ package (version 1.43.17: R version 4.2.2) (Bartoń, Reference Bartoń2023) was used to calculate Pseudo R 2 for each model. Year and grazing treatment effects on SOC and N were tested using the anova function with a Kenward–Roger approximation using the ‘car’ package (Fox and Weisberg, Reference Fox and Weisberg2018).
Results
Aboveground standing crop biomass
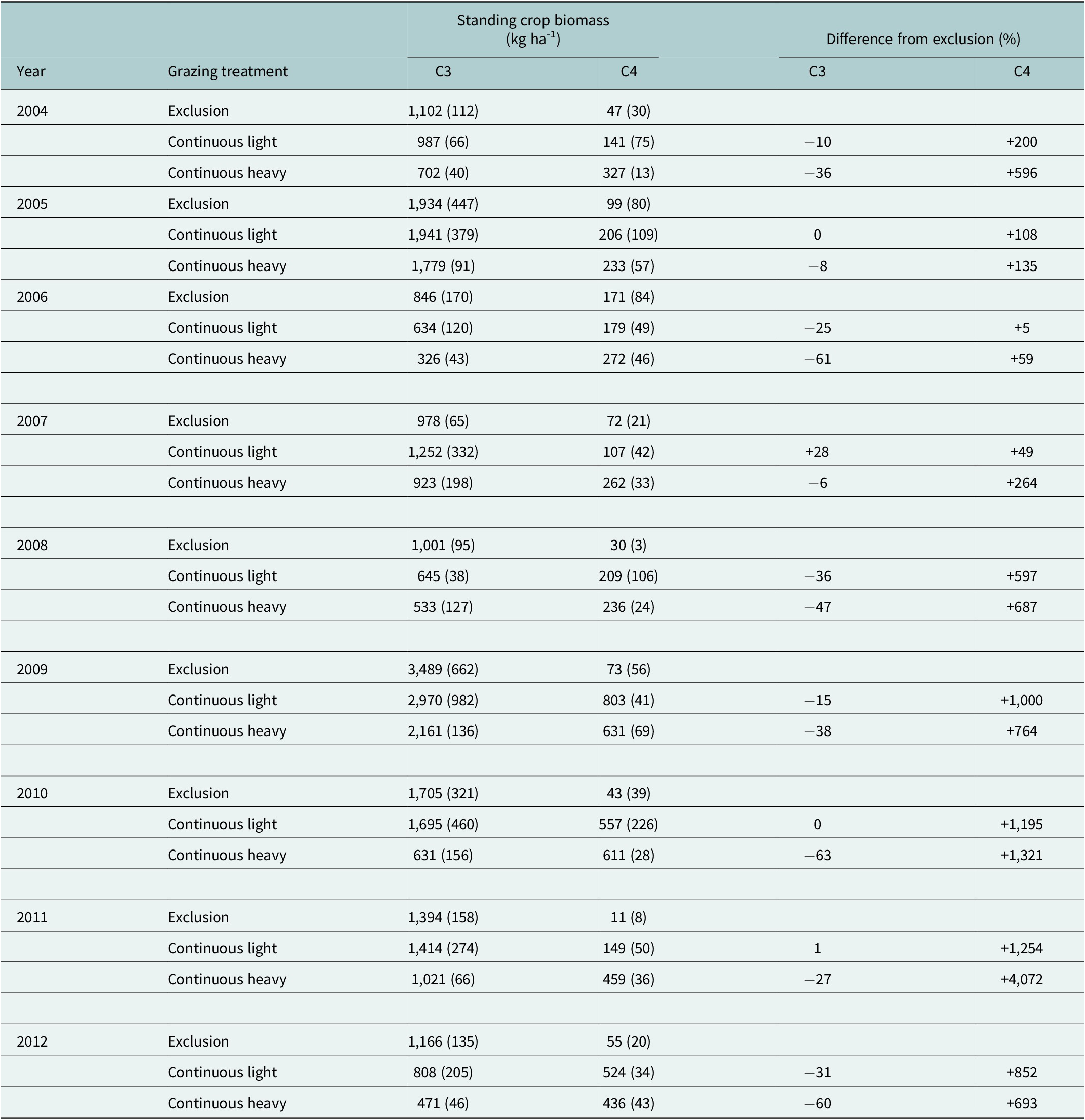
High years of aboveground standing crop biomass during the 2004–2012 period occurred in 2005 and 2009, with low years occurring in 2006, 2008, and the year preceding soil sampling 2012 (Table 1). Increasing grazing intensity from the exclusion (no grazing) to heavy treatment reduced mean annual aboveground standing crop biomass by 16% (1,335 vs. 1,580 kg ha−1) with a 37% (950 vs. 1,513 kg ha−1) reduction in the cool-season (C3) component but a 474% (385 vs. 67 kg ha−1) increase in the warm-season (C4) component. Aboveground standing crop biomass did not exhibit a directional change with time for any of the grazing treatments (P = 0.56, data not shown)
Mean (± one SE) cool-season (C3) and warm-season (C4) aboveground standing crop biomass in mid-July (kg ha-1) for three grazing treatments (exclusion, continuous light and continuous heavy) from 2004 to 2012 in northern mixed-grass prairie near Cheyenne, Wyoming, USA

Precipitation and air temperatures
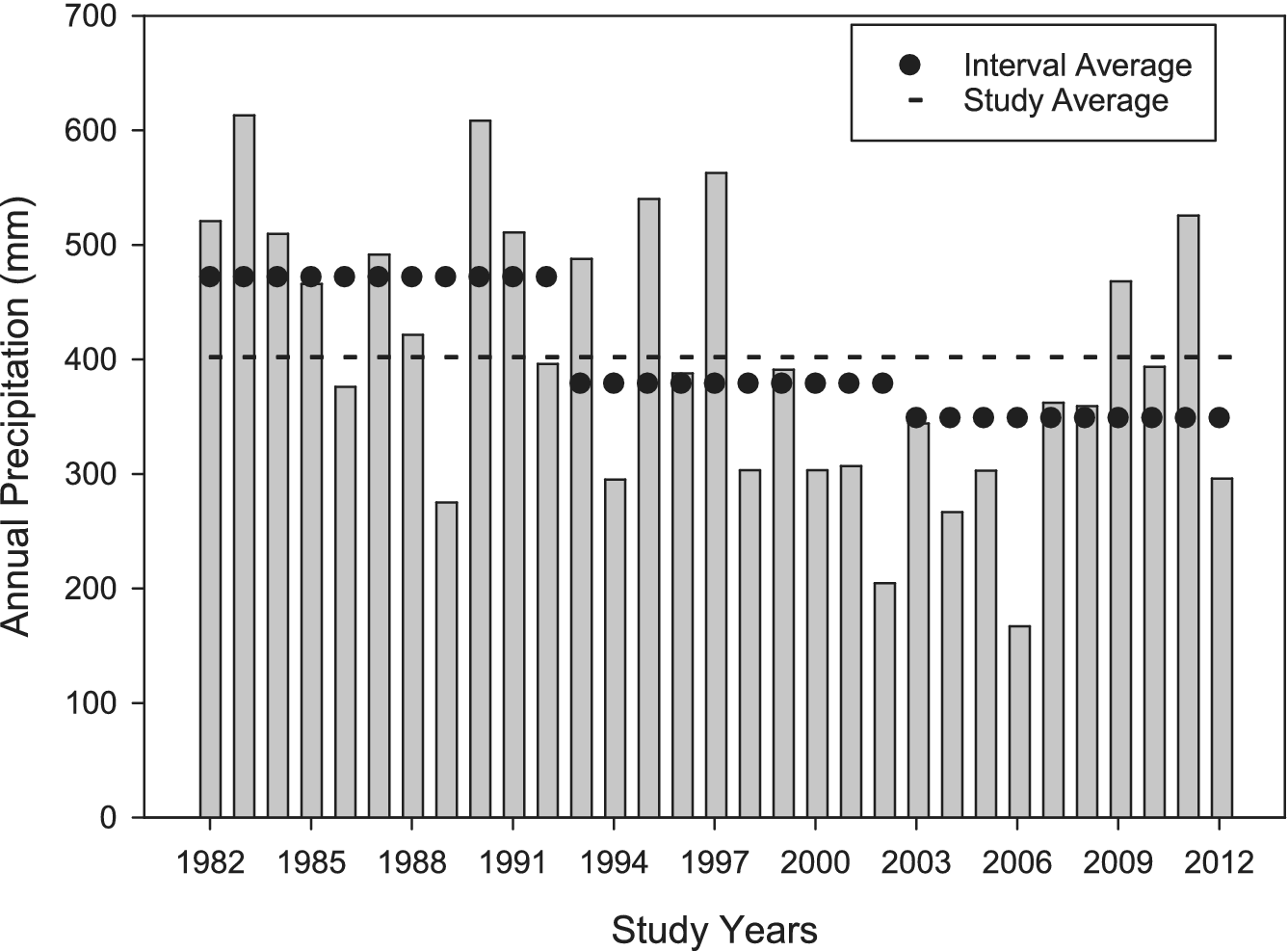
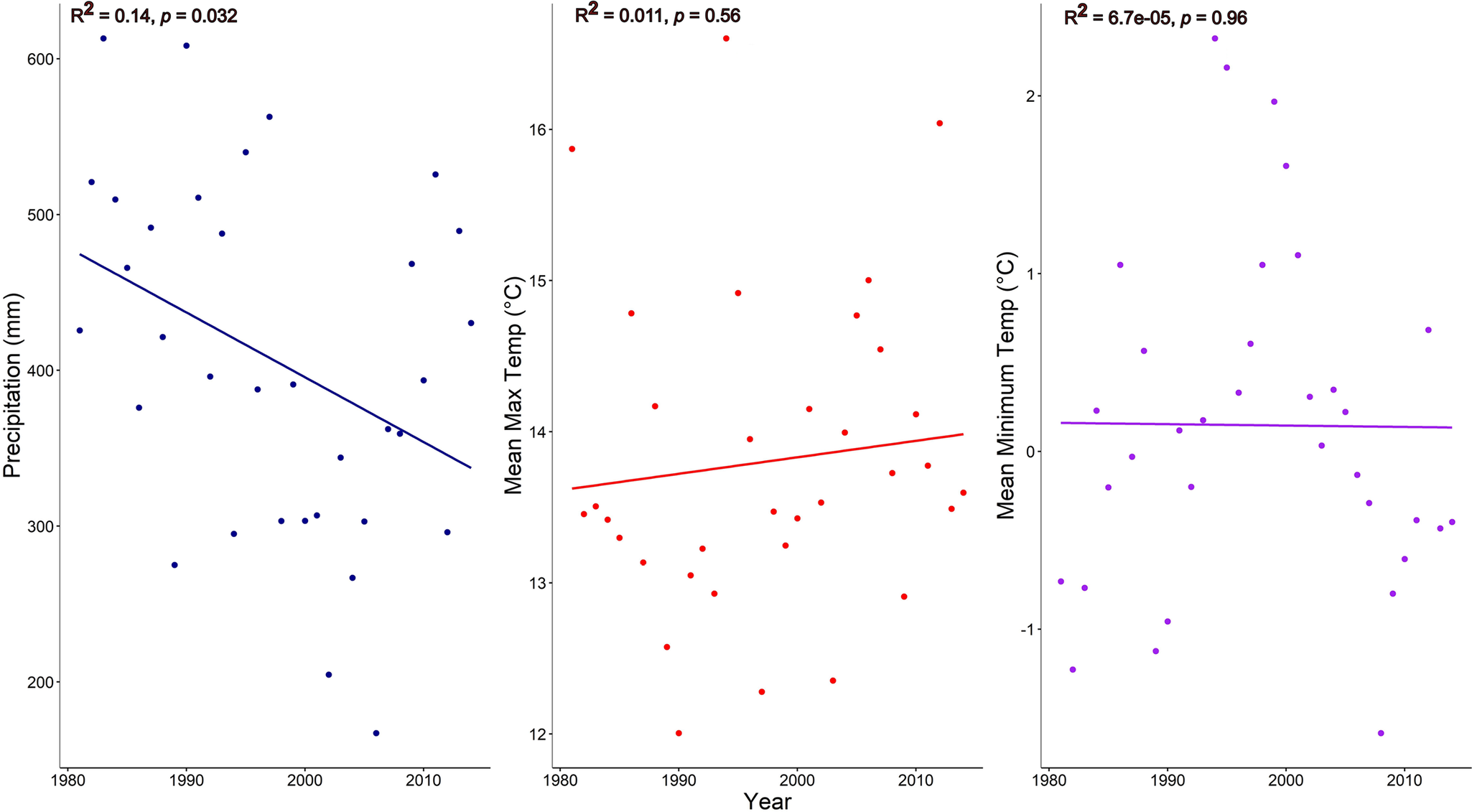
For the full grazing study period (1982–2012), mean annual precipitation was 400 mm (Figure 1), with the 10-year interval means between soils sampling events (May 1993, 2003 and 2013) decreasing with time. Linear regression analyses of annual precipitation revealed a decreasing linear trend of 4.2 mm yr−1 from 1982 (initiation of the grazing study) to the early 2010s (Figure 2, P = 0.032). Conversely, mean annual maximum (P = 0.56) and minimum (P = 0.96) temperatures did not exhibit a trend during this study period.
Annual precipitation (mm) at the USDA-ARS High Plains Grasslands Research Station near Cheyenne, Wyoming, USA, from 1982 (initial year of grazing treatments) to 2012, with three decadal increments (1982–1992, 1993–2002 and 2003–2012) showcased prior to the soil sampling events in May 1993, 2003, and 2013).

Directional trends in annual precipitation (mm), mean annual maximum temperature (°C) and mean annual minimum temperature (°C) at the USDA-ARS High Plains Grasslands Research Station near Cheyenne, Wyoming, USA, from 1981 (year prior to the initiation of grazing treatments) and 2013.

SOC and N concentrations
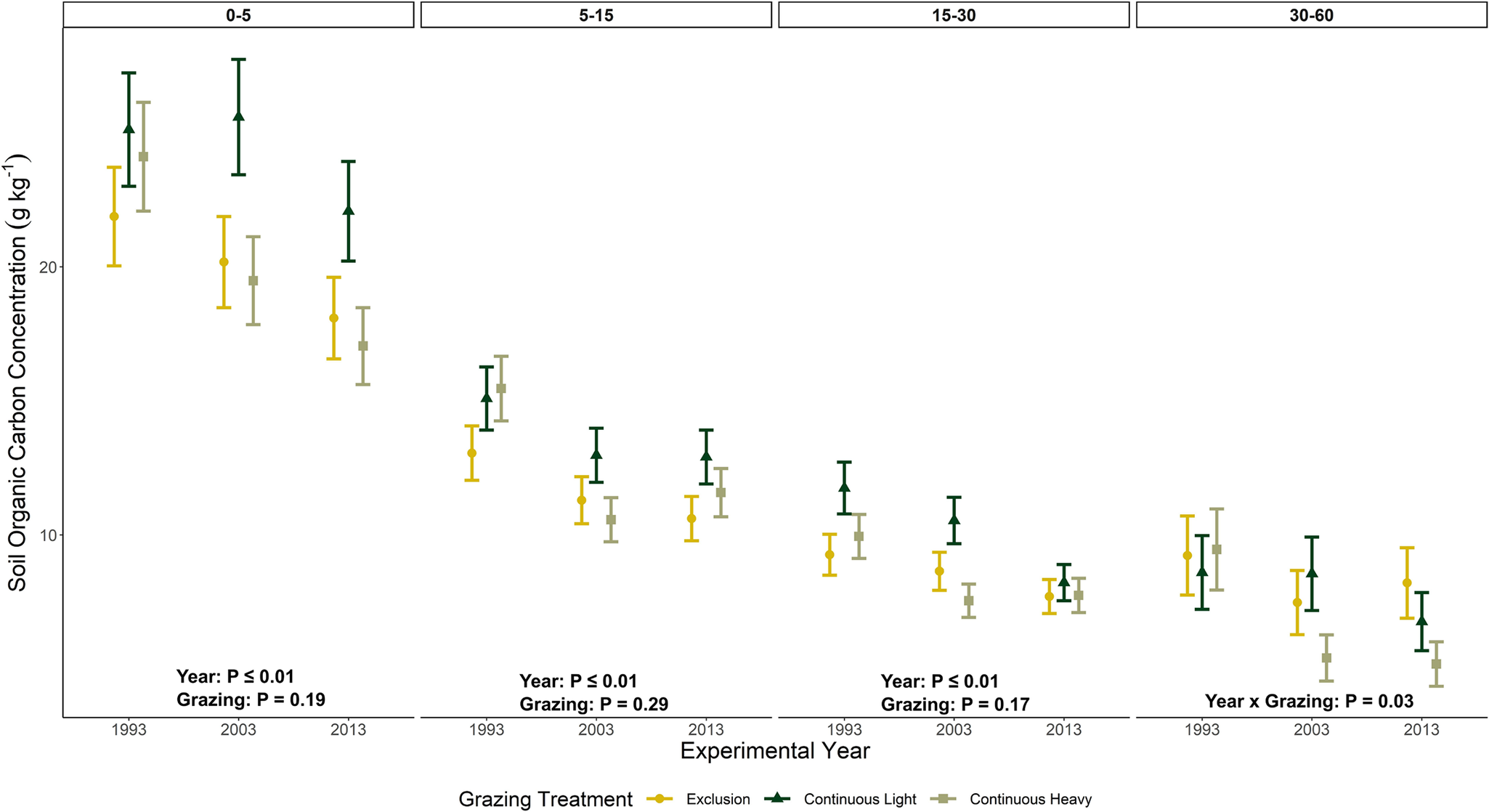
Concentrations of SOC were influenced (P < 0.01) by year with consistent decreasing trends observed from 1993 to 2013 for the top three depths (0–5, 5–15 and 15–30) cm for all grazing treatments (Figure 3, Supplementary Tables 1 and 2). Grazing did not affect SOC concentrations for these depths but did interact (P = 0.03) with year of sampling (1993, 2003 and 2013) at the 30–60 cm depth.
Soil organic carbon concentrations (mean ± one SE, g kg-1) as affected by grazing treatment (exclusion, continuous light, continuous heavy) at four soil depths (0–5, 5–15, 15–30 and 30–60 cm) for three experimental years (1993, 2003 and 2013) at the USDA-ARS High Plains Grasslands Research Station near Cheyenne, Wyoming, USA. No significant pairwise comparisons occurred between grazing treatments within a given year.

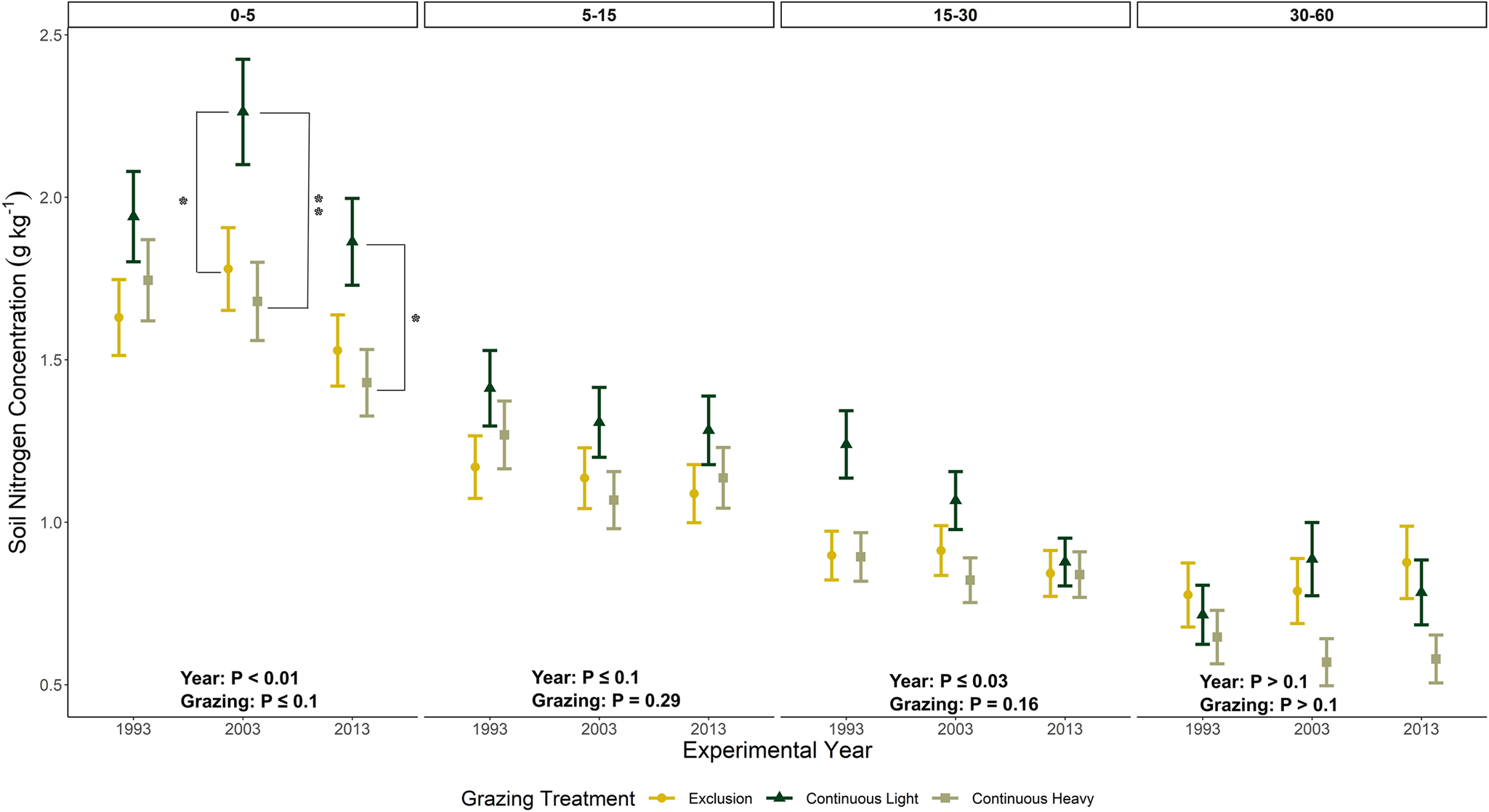
Concentrations of soil N exhibited similar patterns as SOC in general, with decreasing trends over the two decades between soil samplings for the upper three soil depths (Figure 4, Supplementary Tables 1 and 2). Grazing did affect (P ≤ 0.1) soil N in the upper soil depth (0–5) cm with higher concentrations observed with continuous light compared to exclusion and continuous heavy, which did not differ. At the deepest soil depth (30–60 cm) no differences with year or grazing treatment were observed.
Total soil nitrogen concentrations (mean ± one SE, g kg-1) as affected by grazing treatment (exclusion, continuous light, continuous heavy) at four soil depths (0-5, 5-15, 15-30 and 30-60 cm) for three experimental years (1993, 2003 and 2013) at the USDA-ARS High Plains Grasslands Research Station near Cheyenne, Wyoming, USA. Asterisks indicate significant pairwise comparisons between grazing treatments within a given year. * Significant at P ≤ 0.1, ** Significant at P ≤ 0.05.

Discussion
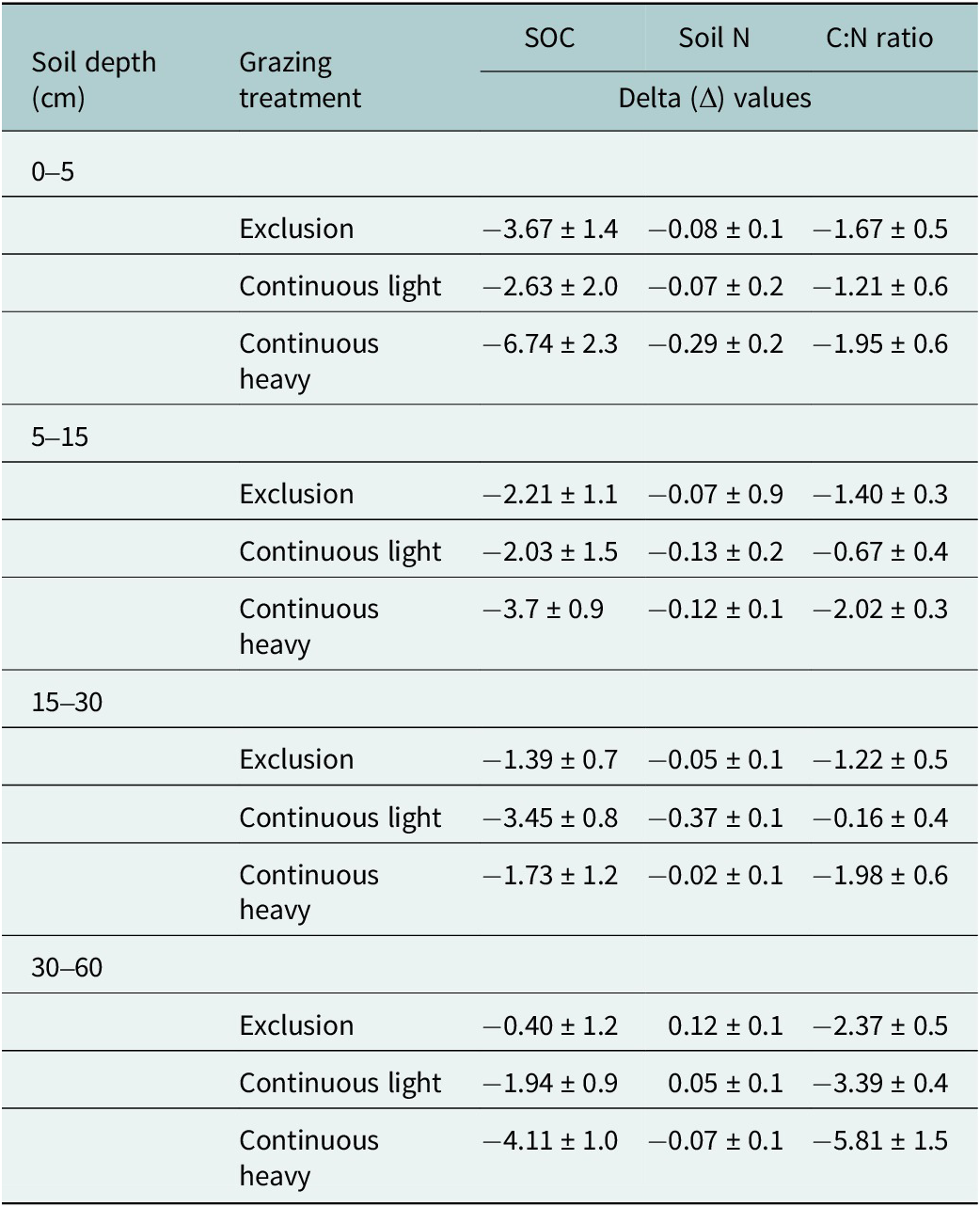
Understanding the temporal impacts of livestock grazing on SOC and N in dryland rangeland ecosystems is constrained by the lack of long-term grazing experiments with longitudinal soil samplings. Our findings from the three soil samplings (1993, 2003 and 2013) indicate soil SOC (Figure 3, Table 2) and soil N (Figure 4, Table 2) loss over the 30+ years of this experiment (1982–2013), irrespective of different grazing intensities from none (exclusion) to light to heavy during the grazing season. Grazing treatments, in general, did not affect SOC and N concentrations. Exceptions included grazing increasing soil N in the 0–5 cm depth with the light grazing treatment and grazing interacting with year at the 30–60 cm depth for SOC. Even with the increase in soil N at the uppermost soil depth with light grazing, improvements in soil health were not observed within this study site (Taylor et al., Reference Taylor, Derner, Liptzin, Porensky, Lavallee, Augustine and Hoover2025). In this dryland, plant communities differ with grazing intensity (Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016) and aboveground standing crop biomass decreases from exclosures to the heavy grazing intensity (Table 1, Derner and Hart, Reference Derner and Hart2007). We did observe a decreasing trend in mean annual precipitation for the three sampling intervals (Figure 1) resulting in a decline of 42 mm per decade (Figure 2) suggesting that other ecological processes may have a much larger influence than livestock grazing on SOC and N concentrations in dryland semiarid rangeland ecosystems (Wu et al., Reference Wu, Chen, Delgado-Baquerizo, Liu, Wang, Wu, Hu and Bai2022).
Mean (± one SE) raw delta values for soil organic carbon concentration (SOC; g kg−1), total soil nitrogen concentration (Soil N; g kg−1), and soil carbon-to-nitrogen ratios (C:N Ratio) for four soil depths (0–5, 5–15, 15–30 and 30–60 cm) and three grazing treatments (exclusion, continuous light and continuous heavy)

Note: Delta (Δ) values were calculated as the change in concentrations or ratio between experimental years 2013 (third decade) and 1993 (first decade). Negative values indicate a decrease in respective soil responses by 2013 relative to 1993.
Our findings of little to no change in SOC due to grazing with several longitudinal soil sampling events over the thirty years agree with single soil sampling events after decades of livestock grazing treatments in similar drylands. For example, seven decades of moderate or no grazing in a C4-dominated, dryland shortgrass steppe did not affect SOC (Derner et al., Reference Derner, Augustine and Frank2019). Fifty-eight years with no grazing, moderate, or heavy grazing did not change SOC in a C3-dominated dryland grassland (rough fescue, Festuca campestris) in Canada (Henderson et al., Reference Henderson, Ellert and Naeth2004). Differences between no grazing and heavy grazing for SOC after 78 years were not detected in a dryland northern mixed-grass prairie of North Dakota (Frank et al., Reference Frank, Tanaka, Hofmann and Follett1995). Furthermore, grazing management strategy (e.g., rotational, continuous or seasonal grazing) did not impact soil C across grazing studies in Australian drylands (McDonald et al., Reference McDonald, Badgery, Clarendon, Orgill, Sinclair, Meyer, Butchart, Eckard, Rowlings, Grace and Doran-Browne2023).
Heavy grazing intensity shifts the native plant community in this northern mixed grass prairie from a C3/C4 mix to one dominated by the low productivity C4 perennial grass blue grama (Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016). This grazing-induced change reduces aboveground productivity by approximately 20% (Derner and Hart, Reference Derner and Hart2007, Table 1) and decreases plant interspaces and litter cover as grazing intensity increases (Derner and Whitman, Reference Derner and Whitman2009). These aboveground production and vegetation changes associated with grazing treatments, however, were not consistent with observed effects of grazing on concentrations of SOC and soil N in this semiarid rangeland ecosystem. The lack of grazing treatment differences on SOC concentrations in the upper 30 cm of soil across this multidecadal study agrees with prior findings of no grazing effect from other multidecadal studies in the neighboring C4-dominated semiarid shortgrass steppe in Colorado, USA (Derner et al., Reference Derner, Augustine and Frank2019; Taylor et al., Reference Taylor, Derner, Liptzin, Porensky, Lavallee, Augustine and Hoover2025). These studies, in the dry portion of the North American Great Plains, contrast with general findings from global metanalyses where SOC decreases as grazing intensity increases (Aryal, Reference Aryal2022; Liu et al., Reference Liu, Liu, Yang, Li, Feng, Yu, Zhang and Dong2022; Deng et al., Reference Deng, Shangguan, Bell, Soromotin, Peng, An, Wu, Xu, Wang, Li and Tang2023; Xu et al., Reference Xu, You, Tan, Xu, Liu, Wang, Xu, Sardans and Peñuelas2023; Hao et al., Reference Hao, Yang, Dong, He and Zhang2024; Ren et al., Reference Ren, Terrer, Li, Cao, Yang and Liu2024); this suggests that factors including the evolutionary history of grazing in the studied ecosystem (Milchunas et al., Reference Milchunas, Sala and Lauenroth1988) should be considered regarding SOC responses of rangelands to grazing. For example, both the northern mixed-grass prairie (this study) and the shortgrass steppe (Derner et al., Reference Derner, Augustine and Frank2019; Taylor et al., Reference Taylor, Derner, Liptzin, Porensky, Lavallee, Augustine and Hoover2025) are in semiarid environments with long evolutionary histories of grazing where plant communities have a high grazing tolerance (Milchunas et al., Reference Milchunas, Sala and Lauenroth1988) and grazing-indued changes to plant communities occur are slow, occurring at decadal scales and reversible (Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016, Reference Porensky, Derner, Augustine and Milchunas2017). Thus, it is not surprising that our findings concur with Reinhart et al. (Reference Reinhart, Rinella, Waterman, Worogo and Vermeire2024) as grazing treatment induced changes to aboveground vegetation have limited predictive ability for SOC changes in semiarid rangelands of the North American Great Plains where ecosystem phase but not state changes occur with grazing (Bestelmeyer et al., Reference Bestelmeyer, Brown, Havstad, Alexander, Chavez and Herrick2003, Reference Bestelmeyer, Tugel, Peacock, Robinett, Shaver, Brown, Herrick, Sanchez and Havstad2009; Porensky et al., Reference Porensky, Mueller, Augustine and Derner2016, Reference Porensky, Derner, Augustine and Milchunas2017) as thousands of years of legacy grazing by native herbivores and more recently domesticated livestock have established an evolutionary history of grazing that can mask effects of recent grazing management treatments (Milchunas et al., 1998).
Edaphic factors, rather than management, are an overarching controlling factor for SOC and N in drylands (Stanley et al., Reference Stanley, Wilson, Patterson, Machmuller and Cotrufo2024). For example, the observed downward directional change in annual precipitation could have been a potential factor in the reduced concentrations of SOC and soil N during the duration of this multidecadal experiment. Similarly, SOC and N decrease with increasing duration of the study was a finding from the Xu et al. (Reference Xu, You, Tan, Xu, Liu, Wang, Xu, Sardans and Peñuelas2023) metanalysis of 95 livestock grazing studies, as well as for long-term (1947–2018) dryland cropping sites in the Great Plains (Liebig et al., Reference Liebig, Calderon, Clemensen, Durso, Duttenhefner, Eberly, Halvorson, Jin, Mankin, Margenot and Stewart2024). Our field observations of reduced SOC concentrations through time also concur with declines of SOC in this same northern mixed-grass prairie under experimental warming (Carrillo et al., Reference Carrillo, Dijkstra, LeCain, Blumenthal and Pendall2018). This indicates that environmental influences on plant–soil water impact soil microbial communities, function and structure may be important in driving soil nutrient changes.
Livestock grazing alters soil bacterial and fungal community structure in grasslands with meta-analysis results from 48 grazing experiments indicating greater impacts on the bacterial than fungal community (Jing et al., Reference Jing, Li, Mipam, Jiang, Liu, Hu and Tian2025). At this field site, grazing intensity influences microbial biomass, respiration and N-mineralization with rates decreasing as grazing intensity increases (Taylor et al., Reference Taylor, Derner, Liptzin, Porensky, Lavallee, Augustine and Hoover2025), and the microbial community structure differs between grazing treatments (Ingram et al., Reference Ingram, Stahl, Schuman, Buyer, Vance, Ganjegunte, Welker and Derner2008). The abandonment of grazing (i.e., exclosure, non-grazed treatment) has been demonstrated to enhance aboveground biomass in the short-term with soil C sequestration exhibiting a lagged response (Qu et al., Reference Qu, Deng, Shangguan, Sun, He, Wang, Zhou, Li and Peñuelas2024), and excluding grazing decouples soil microbial networks in Mediterranean grasslands (Serrano et al., Reference Serrano, Peco, Morillo and Ochoa-Hueso2024). Grazing exclusion in drylands can be a short-term effective management strategy for increasing SOC stocks when substantial prior degradation has occurred (Phukubye et al., Reference Phukubye, Mutema, Buthelezi, Muchaonyerwa, Cerri and Chaplot2022; Qu et al., Reference Qu, Deng, Shangguan, Sun, He, Wang, Zhou, Li and Peñuelas2024).
Losses of SOC and N in this semiarid rangeland are likely attributed to the combined influences of grazing-induced plant interspaces and litter cover (Derner and Whitman, Reference Derner and Whitman2009), which interact with pulses of precipitation in semiarid rangelands to affect the functioning of ecosystems (Schwinning et al., Reference Schwinning, Sala, Loik and Ehleringer2004; Schwinning and Sala, Reference Schwinning and Sala2004). As a result, the pulsiness of precipitation induces large short-term losses of soil N (Dijkstra et al., Reference Dijkstra, Augustine, Brewer and von Fischer2012) and soil C due to soil microbial and root respiration (Huxman et al., Reference Huxman, Snyder, Tissue, Leffler, Ogle, Pockman, Sandquist, Potts and Schwinning2004; Chen et al., Reference Chen, Lin, Huang and He2008; Munson et al., Reference Munson, Benton, Lauenroth and Burke2010; Jiang et al., Reference Jiang, Bian, Xu, Li and He2021). These short-term losses accumulate with time to result in larger magnitudes of SOC and N loss as experienced in this multidecadal field study.
Conclusions
Expectations of increased SOC and N with livestock grazing during the growing season are not reasonable in semiarid rangeland ecosystems of the North American Great Plains with a long evolutionary grazing history such as this northern mixed-grass prairie. Rather, environmental controls on soil microbially mediated processes likely exert substantial influence on SOC and N concentrations in these drylands, through respiration losses via soils and plants associated with pulses of precipitation. Observed downward directional trends in precipitation in this multi-decadal study may likely impact ecosystem functioning for soil SOC and N concentrations more than grazing-induced changes in plant communities and structure. As aridity increases in drylands globally (Berdugo et al., Reference Berdugo, Vidiella, Solé and Maestre2022), it is important to understand how long-term weather drivers and livestock management may influence biogeochemical cycles at local to global scales.
Open peer review
For open peer review materials, please visit http://doi.org/10.1017/dry.2026.10027.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/dry.2026.10027.
Data availability statement
Data is available at the USDA National Agricultural Library.
Acknowledgements
We gratefully acknowledge the establishment of the grazing study by Dr. Richard H. Hart in 1982, and the technical support persons throughout the study, including Pam Freeman and numerous summer students. We sincerely appreciate the scientific and soil sampling efforts of Dr. Gerald (Jerry) Schuman on this study. Liz Pruessner conducted the laboratory analyses. King Ranch provided the yearlings for grazing treatments. This research was a contribution from the Long-Term Agroecosystem Research (LTAR) network. LTAR is supported by the United States Department of Agriculture.
Author contribution
Conceptualization: J.D.D., K.M.T., C.E.S., D.L.H.; Data Collection: M.C.M.; Formal Analysis: K.M.T., M.C.M., C.E.S.; Funding Acquisition: J.D.D.; Investigation: J.D.D.; Methodology: J.D.D., K.M.T., M.C.M.; Project Administration: J.D.D.; Resources: J.D.D.; Supervision: J.D.D.; Validation: K.M.T.; Visualization: K.M.T., C.E.S., D.L.H.; Writing – Original Draft & Editing: J.D.D.; Writing – Review & Editing: K.M.T., M.C.M., C.E.S., D.L.H.
Financial support
This research was a contribution from the Long-Term Agroecosystem Research (LTAR) network. LTAR is supported by the US Department of Agriculture. The primary project number for this study was 3012-21500-001-000D. Research was funded by the USDA-Agricultural Research Service. USDA is an equal opportunity provider, employer and lender. Any use of trade, firm or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Competing interests
Co-author David L. Hoover is an editor for this journal.



 Open access
Open access
Comments
Thank you for the opportunity to submit this manuscript entitled “Multi-decadal grazing effects on soil organic carbon and nitrogen concentrations in a semiarid rangeland” as a Research Paper for consideration of publication in Drylands. Assessing livestock grazing impacts on soil organic carbon and nitrogen in rangelands are difficult to categorize given complexity of interactions among long-term weather factors, vegetation characteristics, and soils, as well as site-specificity of conditions. Moreover, long-term studies in semiarid rangelands with consistent grazing treatments and repeated soil samplings are lacking to understand the influences of grazing with respect to environmental influences such as directional trends in long-term weather factors.
This study capitalizes on a multi-decadal grazing experiment in the semiarid North American Great Plains, northern mixed-grass prairie, with grazing season (June-late September) grazing treatments of season-long light and heavy stocking rates, and exclosure (non-grazed) established in 1982 and soils sampled in 1993, 2003, and 2013.
This work has not been published before, is not being considered for publication elsewhere and has been read and approved by all authors. Thanks again for the opportunity to submit this manuscript. If I can be of any assistance or if questions arise, please contact me using the information provided below.
Sincerely,
Justin Derner
USDA-ARS Rangeland Resources and Systems Research Unit