


Management Implications
Lonicera maackii (Amur honeysuckle) is an aggressive and pervasive invasive plant that forms dense stands in a variety of environments, but especially in the forest understory, which significantly alters native plant community structure, ecosystem function, and forest succession. As such, persistent management is necessary to transpose a L. maackii–dominated forest understory back to native plant communities. Our findings corroborate other studies showing that cut stump glyphosate treatments effectively control L. maackii over the short term. However, our results do not demonstrate that the restoration plantings used in this study were a beneficial restoration tool to mitigate L. maackii reinvasion as hypothesized. Mature L. maackii stands produce copious amounts of seed, and overseeding with Elymus spp. (wildrye grasses) after L. maackii removal in effect promotes soil conservation and may provide competition and suppression of L. maackii reinvasion from seed rain or soil seedbank in response to increased light availability after L. maackii removal. Lindera benzoin (northern spicebush) is a common native shrub in deciduous forests of the eastern United States and occupies same niche as L. maackii (i.e., understory shrub layer); we hypothesized it might effectively compete with and suppress future L. maackii reinvasion. However, results of this study did not support that hypothesis, as the L. benzoin plantings experienced high mortality and were largely unsuccessful due to combination of low planting density, deer browsing, and late summer droughts. Nonetheless, we learned some practical management lessons regarding L. benzoin restoration plantings and recommend the following: (1) do not underestimate the impact of deer browsing—use tree protectors around newly transplanted shrubs or tree seedlings; (2) plant at high density (e.g., 1 shrub every 1 to 2 m2), as some mortality is likely; and (3) transplants may take several years to reach reproductive maturity depending on the restoration site, as we observed our first L. benzoin fruiting 5 to 6 yr after planting, in summer 2024 and 2025. Additional studies are needed to better determine whether Elymus spp. and L. benzoin (or other native understory shrub) plantings are a beneficial restoration tool to mitigate L. maackii reinvasion and promote forest understory native plant community recovery. Furthermore, more studies are needed to evaluate the efficacy of different herbaceous woodland native plant seed mixes and additional native shrubs in restoration plantings, especially local versus nonlocal ecotypes, to suppress L. maackii reinvasion and facilitate the recovery and succession of forest native plant communities.
Introduction
Aside from habitat loss, invasive species are one of the leading causes of ecological change and biodiversity loss (Didham et al. Reference Didham, Tylianakis, Hutchison, Ewers and Gemmell2005; Ehrenfeld Reference Ehrenfeld2003; Elton Reference Elton1958; Vitousek et al. Reference Vitousek, D’Antonio, Loope and Westbrooks1996). The discipline of invasion ecology has grown exponentially over the last three decades as a result of the profound and widespread impacts, both ecological and economic, that invasive species impart to their new environments (Seebens et al. Reference Seebens, Blackburn, Dyer, Genovesi, Hulme, Jeschke, Pagad, Pyšek, Winter, Arianoutsou, Bacher, Blasius, Brundu, Capinha and Laura Celesti-Grapow2017). In particular, non-native, invasive plants (hereafter invasive plants) have received a considerable amount of attention (Giora et al. Reference Giora, Hulme, Richardson and Pyšek2023; Richardson et al. Reference Richardson, Pyšek, Rejmánek, Barbour, Panetta and West2000), as many dramatically alter community structure and function of natural communities and ecosystems they invade (Ehrenfeld Reference Ehrenfeld2003; Giora et al. Reference Giora, Hulme, Richardson and Pyšek2023; McNeish and McEwan Reference McNeish and McEwan2016). Sources from which invasive plants establish in new environments vary (i.e., intentional vs. unintentional), but studies suggest the horticulture industry may be a main driver of some plant invasions, particularly amid current climate change conditions (Bell et al. Reference Bell, Wilen and Stanton2003; Niinemets and Peñuelas Reference Niinemets and Peñuelas2007; Reichard and White Reference Reichard and White2001; van Kleunen et al. Reference van Kleunen, Bossdorf and Dawson2018). It also has been well established that invasive plants often possess traits that afford them a “competitive edge” (rapid growth and reproductive rates, environmental plasticity, loss of natural enemies, novel weapons, allelopathy, etc.) relative to native plants (Keane and Crawley Reference Keane and Crawley2002; van Kleunen et al. Reference van Kleunen, Dawson, Schlaepfer, Jeschke and Fischer2010a, Reference van Kleunen, Weber and Fischer2010b, Reference van Kleunen, Dawson and Maurel2015, Reference van Kleunen, Bossdorf and Dawson2018; Lieurance and Cipollini Reference Lieurance and Cipollini2013; Liu and Stiling Liu and Stiling Reference Liu and Stiling2006). In addition, non-native, woody plant invasions in forest systems are especially problematic, as they impact multiple strata of the forest (Webster et al. Reference Webster, Jenkins and Jose2006).
Lonicera maackii (Rupr.) Herder (Caprifoliaceae), commonly known as Amur or bush honeysuckle, has a reputation as one of the most pervasive invasive plants in the eastern half of the United States. It is a perennial, deciduous shrub of east Asia origin introduced to the United States in the late 19th century for horticultural/ornamental and wildlife resource purposes (Luken and Thieret Reference Luken and Thieret1996). Since then, L. maackii has established populations in 33 states within the United States (see Boyce Reference Boyce2023). The invasive ecology of L. maackii has received considerable research attention in the last several decades because of its adverse ecological impacts (Henkin et al. Reference Henkin, Medley, Abbitt and Patton2013; McNeish and McEwan Reference McNeish and McEwan2016; Sena et al. Reference Sena, Gauger, Johnson, Shirkey, Caldbeck, Hammock, Kim, Mazza, Pethtel and Leuenberger2021, Reference Sena, Hackworth and Maugans2025). As such, L. maackii is a significant concern in many areas of Kentucky and the surrounding region where it has become the dominant forest understory species in some areas (Castellano and Boyce Reference Castellano and Boyce2007; Hartman and McCarthy Reference Hartman and McCarthy2008; Henkin et al. Reference Henkin, Medley, Abbitt and Patton2013; Sena et al. Reference Sena, Gauger, Johnson, Shirkey, Caldbeck, Hammock, Kim, Mazza, Pethtel and Leuenberger2021, Reference Sena, Hackworth and Maugans2025) and therefore greatly threatens natural succession, biodiversity, ecosystem processes, and overall conservation of natural areas (Crocker et al. Reference Crocker, Muller, Stringer, Cox and Thomas2023; McNeish and McEwan Reference McNeish and McEwan2016; Miller and Gorchov Reference Miller and Gorchov2004).
Lonicera maackii is very abundant across the landscape in central Kentucky, and it will inevitably remain a persistent feature of the landscape; however, it can be extirpated on a small spatial scale with persistent invasive species management efforts (Crocker et al. Reference Crocker, Muller, Stringer, Cox and Thomas2023; Frank et al. Reference Frank, Saunders and Jenkins2018; Hopfensperger et al. Reference Hopfensperger, Boyce and Schenk2017; Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010; Luken and Mattimiro Reference Luken and Mattimiro1991; Sena 2021, 2025). Nonetheless, L. maackii will undoubtedly continue to spread across the landscape and continue to impart adverse ecological impacts if left unmanaged in natural areas and areas of conservation priority. Furthermore, it will likely persist in the United States indefinitely as a result, in part, of isolated suburban and urban populations, which are persistent sources of propagule pressure dispersal to natural areas (Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010). The extended leaf phenology whereby L. maackii leaf emergence is much earlier than that of most native plants is especially problematic (McEwan et al. Reference McEwan, Birchfield, Schoergendorfer and Arthur2009; Smith Reference Smith2013). Further, it is well-documented that L. maackii adversely impacts forest structure, including species richness and diversity, tree regeneration (i.e., secondary succession), and forest ecosystem function (Arthur et al. Reference Arthur, Bray, Kuchle and McEwan2012; Boyce Reference Boyce2015; Collier et al. Reference Collier, Vankat and Hughes2002; Hartman and McCarthy Reference Hartman and McCarthy2008; Hopfensperger et al. Reference Hopfensperger, Boyce and Schenk2017; McNeish and McEwan Reference McNeish and McEwan2016; Meiners Reference Meiners2007; Miller and Gorchov Reference Miller and Gorchov2004; Sena et al. Reference Sena, Gauger, Johnson, Shirkey, Caldbeck, Hammock, Kim, Mazza, Pethtel and Leuenberger2021, Reference Sena, Hackworth and Maugans2025). The only way to reverse this trend and promote natural succession and biodiversity conservation is proactive and persistent L. maackii management in the conservation area of interest, whether it is private or public land (Dolan and Brown Reference Dolan and Brown2019; Dolan Reference Dolan2015 ; Emry et al. Reference Emry, Mercader, Bergeron, Eilert and Riddle2024).
Removal and subsequent management of L. maackii reinvasion are essential to begin revitalizing any forest or forest understory back to native plant communities (Boyce Reference Boyce2015; Emry et al. Reference Emry, Mercader, Bergeron, Eilert and Riddle2024; Fotis et al. Reference Fotis, Flower, Atkins, Pinchot, Rodewald and Matthews2022; Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010; McDonnell et al. Reference McDonnell, Mounteer, Owen and Todd2005; Sena et al. Reference Sena, Hackworth and Maugans2025). From anecdotal reports to experimental peer-reviewed research publications, L. maackii control methods have been relatively well established and successful, at least in the short term (Crocker et al. Reference Crocker, Muller, Stringer, Cox and Thomas2023; Emry et al. Reference Emry, Mercader, Bergeron, Eilert and Riddle2024 ; McDonnell et al. Reference McDonnell, Mounteer, Owen and Todd2005) yet controlling invasive woody shrubs can be challenging relative to other invasive plants (Luken and Mattimiro Reference Luken and Mattimiro1991). Lonicera maackii control methods may range from high-disturbance impacts (e.g., using an excavator such as a Bobcat® or tractor) to low-disturbance impact on forest understory and non-target species (e.g., basal bark and cut stump herbicide treatments). However, to our knowledge there is no “silver bullet” to effectively control and manage L. maackii, and management objectives may vary depending on the specific area. For instance, the most cost- and time-effective control method for a forest understory with a significant L. maackii canopy cover (i.e., 50% or higher) and depauperate in understory species richness and diversity may be to mechanically remove all L. maackii present (above- and belowground plant structures) with an excavator with no need for herbicide application. As such, the trade-off for excessive disturbance to forest understory is complete removal of established L. maackii. In contrast, if the objective is minimal forest understory and soil disturbance and the conservation of forest understory species (both herbaceous and woody), an effective L. maackii control require would require manual removal of individual shrubs by various options including: (1) manual removal; (2) mulching head treatment (Frank et al. Reference Frank, Saunders and Jenkins2018); (3) cutting with loppers or chainsaw followed by cut stump herbicide application; and/or (4) basal bark herbicide treatment (no cutting or removal of established L. maackii) (Hogan et al. Reference Hogan, Baker, Bach and Barber2024). Nonetheless, research suggests that control in any given area is most successful when L. maackii is managed on a persistent basis (e.g., annually) and not as a onetime event (Hopfensperger et al. Reference Hopfensperger, Boyce and Schenk2019; Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010).
Here we report the findings from a small-scale, permanent-plot L. maackii restoration study established in March 2019. The question we aimed to address is the following: Can native species restoration plantings better suppress L. maackii reinvasion over time after L. maackii removal compared with no restoration plantings? A recent study by LaPierre et al. (Reference LaPierre, Andreu, Adams, Miller, Hedman and Sharma2025) determined that certain ruderal native grass seed mixes provided better suppression of non-native species than other seed mixes for pine savanna restoration. For the restoration plantings, wildrye grasses (Elymus spp.) and northern spicebush [Lindera benzoin (L.) Blume] were selected, as they are common perennial native species found in understories of deciduous forests of the eastern United States. Furthermore, anecdotal experience from overseeding Elymus spp. indicates they have a high germination rate and establish a dense herbaceous layer that may suppress L. maackii reinvasion (MER, personal observation), while L. benzoin occupies similar forest understory niche as L. maackii, and we hypothesize it may provide competitive suppression of L. maackii in the long term. Accordingly, we hypothesize areas with L. maackii removal plus restoration plantings will experience reduced reinvasion (i.e., lower canopy cover) over time compared with areas with removal and no restoration plantings. Finally, we predict areas with L. maackii removal will also contain higher species richness compared with areas without removal, which would corroborate findings from other studies (Boyce Reference Boyce2015; Collier et al. Reference Collier, Vankat and Hughes2002; Meiners Reference Meiners2007; Sena et al. Reference Sena, Gauger, Johnson, Shirkey, Caldbeck, Hammock, Kim, Mazza, Pethtel and Leuenberger2021, Reference Sena, Hackworth and Maugans2025).
Materials and Methods
Study Site
Our study was conducted in a young, secondary growth forest on Asbury University (AU) property located in Jessamine County, KY (37.856846°N, 84.684301°W) in the Inner Bluegrass physiographic region of Kentucky. AU’s property extends along an area known as the Kentucky River Palisades region; the palisades are a rock outcrop derived from Ordovician limestone that towers above and along portions of the Kentucky River. Soils in the forested study area are relatively shallow (∼43 cm [∼17 in.]), with underlying Ordovician limestone bedrock. Soils are classified as Fairmount–Rock Complex series (clayey, mixed, active, mesic Lithic Hapludolls) with flaggy silty clay (0 to 17.8 cm; 0 to 7 in.), flaggy clay (17.8 to 43.2 cm; 7 to 17 in.), and unweathered Ordovician limestone below on 12% to 30% slopes (USDA-NRCS 2025). Mean annual precipitation and mean annual high and low temperatures from 1991 to 2020 in central Kentucky were 126.6 cm (49.84 in.), 19.1 C (66.3 F), and 7.9 C (46.3 F), respectively (NWS 2024).
The Kentucky River Palisades region is a conservation area of interest, because it supports a high level of biodiversity and is home to a number of unique, rare, and/or threatened species (Campbell and Meijer Reference Campbell and Meijer1989). Several nature preserves have been established to preserve the unique rolling hills topography, geological features, and biodiversity of the Kentucky River Palisades (TNC 2025). The forest canopy at the study location is co-dominated by common hackberry (Celtis occidentalis L.), black walnut (Juglans nigra L.), elm species (Ulmus spp.), chinquapin oak (Quercus muehlenbergii Engelm.), and Shumard’s oak (Quercus shumardii Buckley) (USDA-NRCS 2016). In some areas of the forested property where L. maackii is less abundant or not established, the forest herbaceous layer is rich and diverse in native spring ephemeral wildflowers, ferns, and other native plants. In contrast, much of the forest understory on the upper slopes and ridgetops has been invaded by L. maackii and is highly depauperate in native plant species richness and diversity, including spring ephemerals, shrubs, and other herbaceous plants (Sena et al. Reference Sena, Gauger, Johnson, Shirkey, Caldbeck, Hammock, Kim, Mazza, Pethtel and Leuenberger2021, Reference Sena, Hackworth and Maugans2025).
Experimental Design
In early March 2019, a randomized block design with three replications was established in a secondary growth forest containing a mature and dense understory population of L. maackii. Some shrubs ranged from 6- to 7.6-m tall and 10- to 25-yr old (from observed cut stump annual rings). Three different treatments were randomly assigned within each of three 18.3 by 15.2-m (60 by 50 ft) treatment blocks (i.e., replication units). Treatment plots (n = 3) within each block (n = 3) measured 6.1 by 15.2-m(20 by 50 ft), and treatments were as follows: (1) untreated control; (2) L. maackii removal + cut stump glyphosate (20% v/v; or 0.023 kg ae L−1; Rivera et al. Reference Rivera, Meilan, Scharf, Karve and Jenkins2022) application (hereafter CH); and (3) L. maackii removal + cut stump glyphosate application (0.023 kg L−1) + restoration plantings (hereafter CHP). Lonicera maackii shrubs were cut near the soil surface using a Stihl MS170 chainsaw (Stihl Incorporated, Virginia Beach, VA, USA), but younger shrubs with basal stems less than 3 to 4 cm were cut near the soil surface using handheld loppers, while the small, immature L. maackii shrubs (i.e., <1-cm diameter) were manually removed from the soil (i.e., both shoot and root system). All aboveground slash (i.e., cut stems) was removed from all removal plots. Immediately after cutting, a 20% (v/v) glyphosate solution with blue dye additive (Mark-It Blue®, Monterey Lawn and Garden Products, Fresno, CA, USA) was applied to the entire cut stump surface of each L. maackii shrub using a modified handheld herbicide sprayer with sponge applicator attached to the spray nozzle. The blue dye additive visually insured each L. maackii cut stump received the herbicide application and none were overlooked and left untreated.
From late March through early April 2019, 16 L. benzoin bare-root seedlings (0.5 to 0.75 m in height) purchased from the Missouri Department of Conservation (MDC; https://mdc.mo.gov/trees-plants/tree-seedlings) were transplanted into each of the three CHP treatment plots (total L. benzoin n = 48). Lindera benzoin saplings were planted approximately 1 m from treatment plot edge and 1.5 m apart along two rows. On December 2, 2019, each of the three CHP plots was over-seeded (by hand-broadcast seeding) at 13.4 kg pure live seed (PLS) acre−1 (33.5 kg PLS ha−1) with a forest native cool-season grass mixture of three native wildrye species: eastern bottlebrush grass (Elymus hystrix L.), Canada wildrye (Elymus canadensis L.), and Virginia wildrye (Elymus virginicus L.), purchased from Roundstone Native Seed (www.roundstoneseed.com).
Whitetail deer (Odocoileus virginianus) browsing and uprooting led to an approximate 90% L. benzoin mortality (Cipollini et al. Reference Cipollini, Ames and Cipollini2009; Donoso et al. Reference Donoso, Leonard and Gorchov2024), which was observed within a couple of weeks after L. benzoin plantings. To rectify the loss of experimental L. benzoin plantings, six additional L. benzoin seedlings (also purchased from MDC) were transplanted into each CHP plot (total n = 18) in mid-March 2020 (only six additional seedlings were transplanted due to limited availability). The L. benzoin density after replanting was six to eight shrubs per CHP plot, as several of the original 16 shrubs planted per CHP plot had survived deer browsing. To discourage and prevent further deer browsing and uprooting of newly transplanted L. benzoin seedlings, we enclosed each sapling within a “tree protector” constructed from galvanized metal hardware cloth (1.3 cm (½-in.) grid; approximately 15.2 cm (6-in.) diameter by 45.7 cm (18-in.) height).
Evaluating Treatment Efficacy: Vegetation Sampling
To evaluate and monitor the efficacy of the management treatments over time, plant communities were sampled annually in late May or early June 2021 to 2025. Five 1-m2 quadrat subsamples were randomly placed within each of three replicate treatment plots (total n = 15 quadrat subsamples per treatment). The percent cover of L. maackii, herbaceous plants (Elymus spp. canopy cover was included), and bare ground was visually estimated (0 to 100 m−2) (Daubenmire Reference Daubenmire1959). In addition, a direct count of species richness per square meter was also conducted during each sampling period. Moreover, we did not quantify L. benzoin canopy cover, as it was seldom captured in quadrat subsamples in the CHP treatment plots.
Data Analyses
All data were analyzed using SPSS v. 28 software (IBM Corporation, Armonk, NY, USA). Percentage data, L. maackii cover, herbaceous cover, and bare ground were arcsine square-root transformed to meet assumptions of homogeneity and homoscedasticity (Zar Reference Zar2010). A repeated-measure ANOVA was used to test for effects of time, treatment, and their interaction (time*treatment) on L. maackii cover, herbaceous cover, bare ground, and species richness per square meter at the α = 0.05 level (Zar Reference Zar2010). If an effect was significant (P < 0.05), a Tukey’s honestly significant different (HSD) post hoc test was performed to determine which plant community variable means differed by time, treatment, or time*treatment (Zar Reference Zar2010). Additionally, a Spearman rank-correlation test was conducted on raw data to investigate the relationship between percent L. maackii cover and herbaceous cover, bare ground, and species richness per square meter (Zar Reference Zar2010).
Results and Discussion
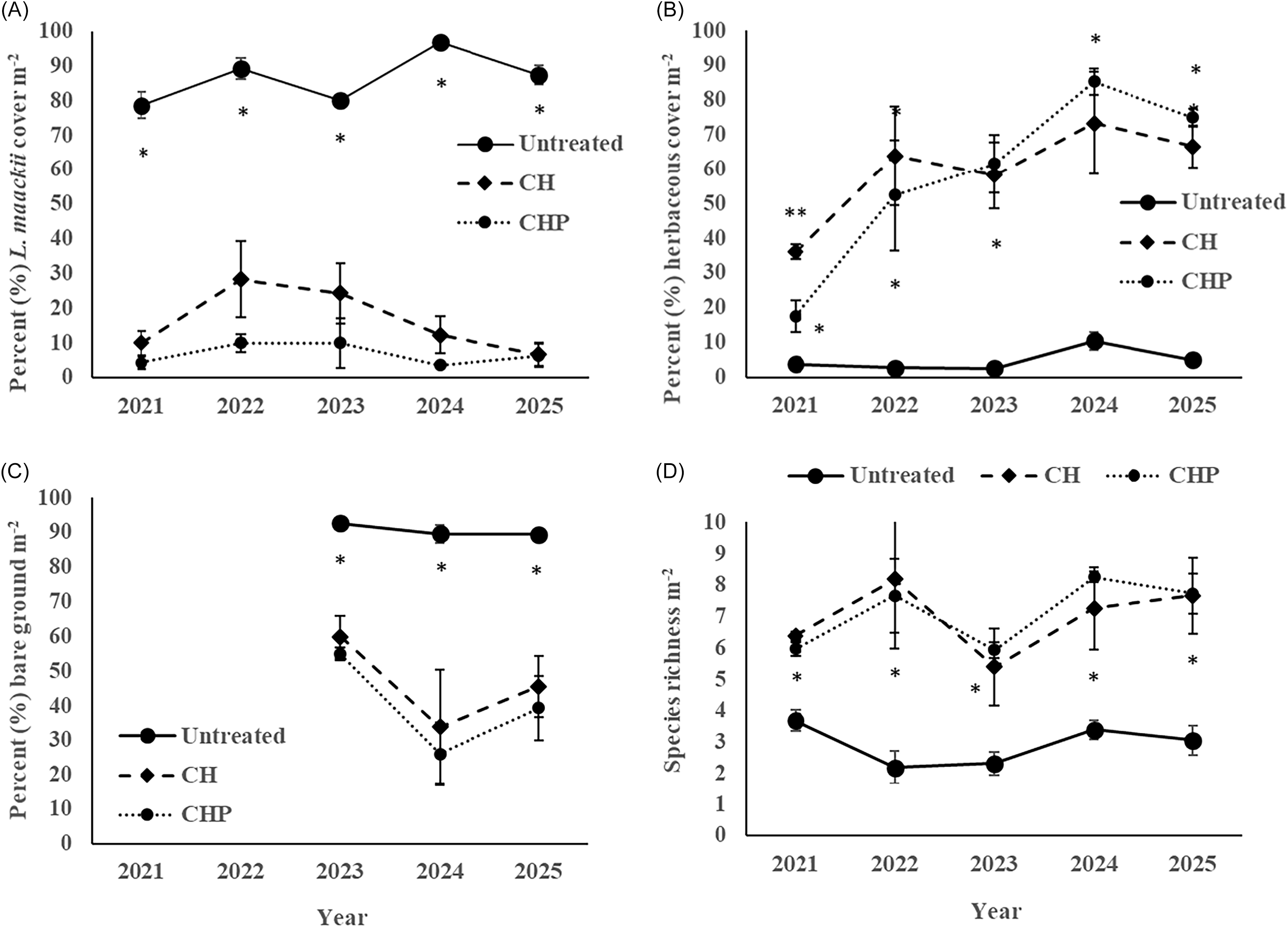
The cut stump glyphosate treatments applied at 0.023 kg ae L−1 in both the CH and CHP treatment plots provided effective L. maackii control over time, wherein removal plots (CH and CHP) averaged 30% or less L. maackii canopy cover m-2 from 2021 to 2025 (Figure 1A). This result corroborates with recent findings of Rivera et al. (Reference Rivera, Meilan, Scharf, Karve and Jenkins2022) using same concentration of glyphosate for L. maackii cut stump applications. However, L. maackii canopy cover in the CH and CHP plots did not differ (P > 0.05; Figures 1A and 2E) throughout 2021 to 2025, indicating the restoration plantings alone did not suppress L. maackii invasion relative to removal alone. As predicted, the untreated plots maintained significantly higher L. maackii canopy cover compared with both the CH and CHP removal treatments since 2021 (Figures 1A and 2). For instance, average L. maackii cover over time (2021 to 2025) was 5.4 and 12.3 times higher over 5 consecutive years (2021 to 2025) in the untreated plots compared with CH and CHP treatments, respectively (P ≤ 0.003) (Figure 1A). Lonicera maackii is pervasive invasive plant with high propagule pressure, and to effectively control it, herbicide treatment on established shrubs is necessary (foliar or cut stump), otherwise it will likely vigorously resprout especially from non–herbicide treated cut stumps (Frank et al. Reference Frank, Saunders and Jenkins2018; Hartman and McCarthy Reference Hartman and McCarthy2004; Hogan et al. Reference Hogan, Baker, Bach and Barber2024; McNeish and McEwan Reference McNeish and McEwan2016). In addition, we observed substantial L. maackii seedlings in removal plots (MER, personal observation), which corroborates with Hopfensperger et al. (Reference Hopfensperger, Boyce and Schenk2019), suggesting it will likely reinvade an area over time without further management.
(A) Percent Lonicera maackii canopy cover, (B) percent herbaceous cover, (C) percent bare ground, and (D) species richness per square meter by treatment over time from 2021 to 2025. A single asterisk (*) indicates a significant difference (P < 0.05) between L. maackii removal (CH and CHP) and untreated treatment plots based on Tukey’s honestly significant different (HSD) post hoc test. A double asterisk (**) in (B) indicates significantly higher herbaceous plant cover (P < 0.05) in the CH plots compared with the CHP and untreated plots based on Tukey’s HSD post hoc test and untreated plots. CH, cut + herbicide treatment; CHP, cut + herbicide + restoration plantings.

Photographs of representative experimental, long-term research plots in May 2021 in comparison to July 2025: (A) Cut + herbicide + restoration planting (CHP) plot in May 2021, (B) cut + herbicide (CH) plot in May 2021, (C) CHP plot with untreated plot in background for comparison in May 2021, (D) untreated plot in July 2025, (E) CHP plot in July 2025, and (F) fruiting Lindera benzoin shrub in CHP plot July 2025. a CH = cut + herbicide; CHP = cut + herbicide + restoration planting. CH plots in July 2025 have similar herbaceous plant cover and species composition as shown in E.

While we found no significant difference in L. maackii cover between CH and CHP plots over time (P > 0.05), it was interesting how it varied in the CH plots over time compared with the CHP plots—it was two times or more higher in CH than in CHP plots in 2022 and 2023 (Figure 1A). This variability likely reflects variation in random quadrat subsampling and is not an effect of restoration plantings suppressing L. maackii reinvasion. However, we observed a substantial amount of chlorotic and necrotic leaves on young shoots of reinvading L. maackii shrubs, which may reflect residual glyphosate effects, but this is unlikely, as these observations were 2 or more years after glyphosate application. A more plausible explanation is leaf mortality from honeysuckle leaf blight (Insolibasidium deformans) infection (Boyce Reference Boyce2018, Reference Boyce2023; Boyce et al., Reference Boyce, Brossart, Bryant, Fehrenbach, Hetzer, Holt, Parr, Poynter, Schumacher, Stonebraker, Thatcher and Vater2014; Boyce et al., Reference Boyce, Castellano, Marroquin, Uwolloh, Farrar and Wolfe2020), although we did not confirm presence of I. deformans leaf infection. Recent research suggests small/young L. maackii shrubs are susceptible to I. deformans infection-related mortality (Helton and Boyce Reference Helton and Boyce2022). Thus, I. deformans infection and subsequent mortality in young reinvading L. maackii shrubs may account for lower average L. maackii canopy cover in CH plots in 2024 and 2025 (Figure 1A). Nonetheless, like many invasive species, L. maackii is an aggressive invader, and regular L. maackii management is necessary and recommended to prevent it from reinvading any given restoration area to sustain native plant community recovery (Hopfensperger et al. Reference Hopfensperger, Boyce and Schenk2019; Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010). Under the assumption we removed and herbicide treated all L. maackii cut stumps in the CH and CHP treatment plots, it is possible some of the L. maackii cover reflects reinvasion from a combination of newly established shrubs and resprouting of some L. maackii cut stumps, based on posttreatment observations during vegetation sampling (MER, personal observation). Lonicera maackii reinvasion is highly probable following removal, and post-removal management efforts, in our opinion, should be routinely performed at a minimum of every 3 to 5 yr to sustain recovery of native plant communities and mitigate L. maackii reinvasion (Hopfensperger et al. Reference Hopfensperger, Boyce and Schenk2017, Reference Hopfensperger, Boyce and Schenk2019; Loeb et al. Reference Loeb, Germeraad, Treece, Wakefield and Ward2010).
Plant Community Response after Lonicera maackii Removal
As predicted, the forest understory plant communities in the CH and CHP plots changed dramatically after L. maackii removal (Figures 1A–D and 2A–C and E). Percent herbaceous plant cover and bare ground were significantly higher and lower, respectively, in both L. maackii removal plots (CH and CHP) compared with untreated plots (P ≤ 0.002), yet they did not differ between CH and CHP treatment plots (P > 0.05) (Figures 1B and C and 2A–C and E). Additionally, species richness was significantly higher over time (P ≤ 0.05) from 2021 to 2025 in CH and CHP plots compared with untreated plots (Figure 1D). The higher herbaceous plant cover and species richness were positive results, yet not surprising, and corroborate many other L. maackii management studies (Boyce Reference Boyce2015; Conover and Sisson Reference Conover and Sisson2016; McDonnell et al. Reference McDonnell, Mounteer, Owen and Todd2005; Sena et al. Reference Sena, Hackworth and Maugans2025)
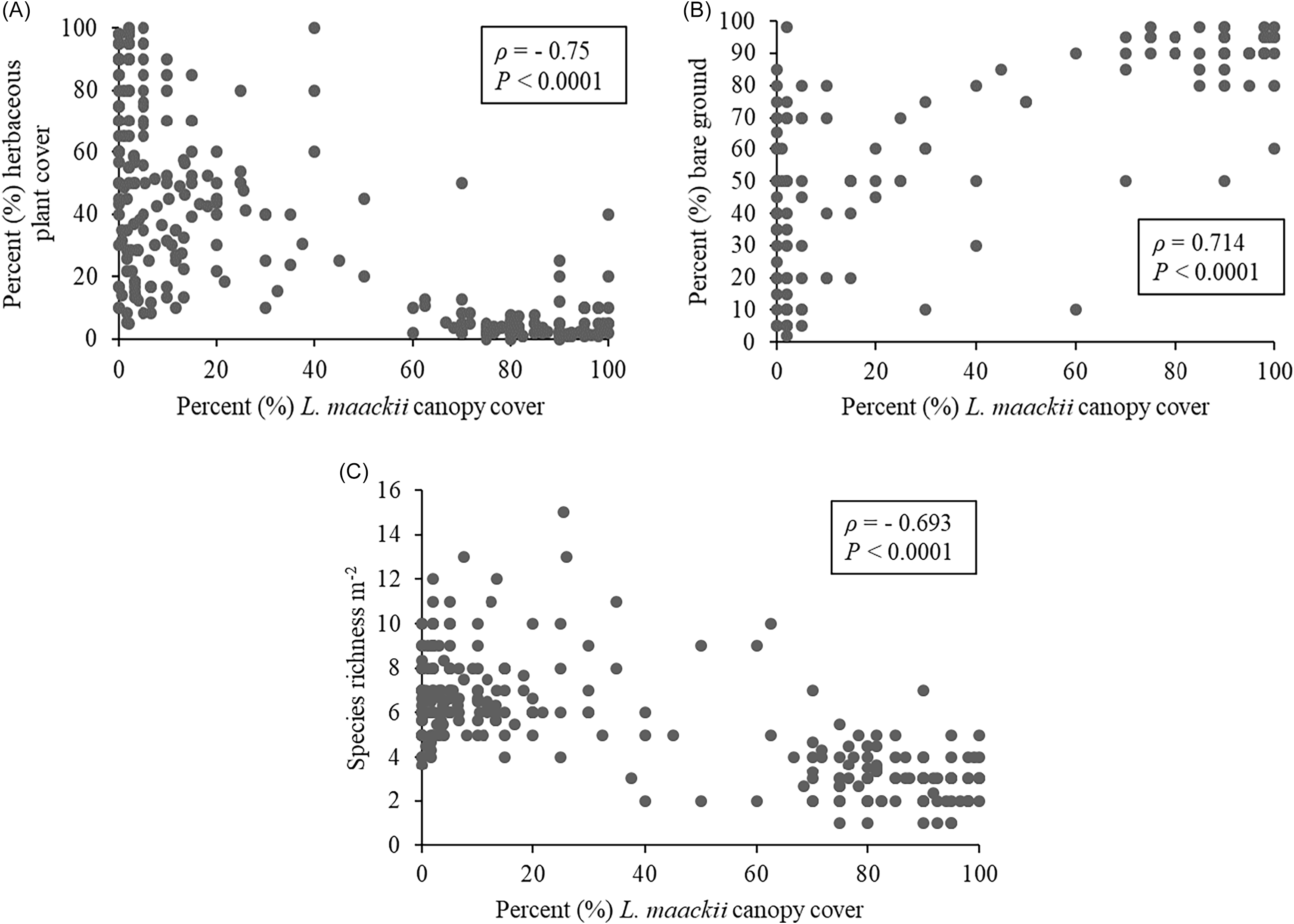
Lonicera maackii cover in untreated plots negatively correlated to low percent herbaceous plant cover (Spearman rank correlation, ρ = −0.75, P < 0.0001), low species richness per square meter (Spearman rank correlation, ρ = −0.69, P < 0.0001), and positively correlated to high percent bare ground (Spearman rank correlation, ρ = 0.714, P < 0.0001) (Figures 1–2). Meanwhile, both the CH and CHP treatment plots, which have experienced low L. maackii canopy cover for the last 6 yr, averaged significantly higher percent herbaceous cover, lower percent bare ground, and higher species richness per square meter compared with untreated plots (Figures 1 and 3).
Scatter plots depicting relationship between percent (%) Lonicera maackii canopy cover, and (A) percent (%) herbaceous plant cover, (B) percent (%) bare ground, and (C) species richness per square meter. Spearman rank correlation (ρ) test results are shown in text boxes.

During the first several growing seasons after L. maackii removal (i.e., 2019 to 2022), we observed a significant release of herbaceous plants from the soil seedbank and/or seed rain in the forest understory in both the CH and CHP plots. In the CHP plots, Elymus spp. established well in spring 2020 following seeding in December 2019 and dominated the forest understory herbaceous plant community through 2022 (Figure 2A and 2C). From 2023 to 2025, most of the established Elymus spp. decreased in abundance, as the plant communities have undergone succession and are now dominated by woodnettle [Laportea canadensis (L.) Weddell] with other broadleaved plants, Elymus spp., and sedges (Carex spp.) being present. Currently, the understory plant communities in the CH and CHP plots are similar in herbaceous plant cover, species composition, and richness 6-yr posttreatment (Figures 1B–D and 2E).
Many L. benzoin seedlings replanted in spring 2020 in the CHP plots have survived, and several shrubs are estimated to be 0.5 to 1.5 m in height (Figure 2F). Nonetheless, there also has been significant L. benzoin mortality (≥50%) in the CHP plots, likely attributable to summer droughts (2022 and 2024) and deer browsing. Excitingly, however, we observed two female L. benzoin shrubs that produced fruit in 2025 for the first time since planting (Figure 2F). In addition, we observed many eastern red cedar (Juniperus virginiana L.) saplings established in both the CH and CHP plots but never under L. maackii shrubs, demonstrating L. maackii removal promotes understory plant community succession and increases woody plant species richness. We also have observed other non-native, invasive species present in all treatment plots, most notably wintercreeper [Euonymus fortunei (Turcz.) Hand.-Maz], garlic mustard [Alliaria petiolata (M. Bieb.) Cavara & Grande], and Chinese privet (Ligustrum sinense Lour.), but there has not been a substantial observed increase in their abundance in the CH or CHP plots since L. maackii removal.
From the results of this study, L. maackii removal alone (i.e., without restoration plantings) facilitated native plant community recovery as described by other studies and equally well as removal plus restoration plantings. Thus, the benefits of restoration plantings as a management tool to suppress L. maackii reinvasion after removal needs further testing. Nevertheless, restoration plantings are beneficial in terms of reintroducing native species formerly displaced or unable to establish amid the competitiveness of L. maackii and its dense canopy cover and alleopathic effects (McNeish and McEwan Reference McNeish and McEwan2016), as well as potentially providing biotic resistance and competition to suppress L. maackii reinvasion. In addition, biotic resistance to invasive species establishment and success is afforded with higher species richness in many cases (Levine et al. Reference Levine, Adler and Yelenik2004; Maron and Vila Reference Maron and Vila2001; Beaury et al. Reference Beaury, Finn, Corbin, Barr and Bradley2020). The successful establishment of L. benzoin restoration plantings could have been greatly improved in this study by not underestimating the slow growth of L. benzoin and impacts of whitetail deer browsing; enclosing each L. benzoin seedling with “tree protectors” after initial planting to discourage deer browsing (Peebles - Spencer et al. Reference Peebles-Spencer, Gorchov and Crist2017); and planting L. benzoin seedlings at a higher density, as some mortality is inevitable, especially during the first year after planting. At the time of our study, we purchased L. benzoin seedlings from the MDC due to no availability of central Kentucky ecotype L. benzoin seedlings. Use of local ecotype sources in restoration plantings is often advocated by restoration ecologists, and planting of nonlocal L. benzoin ecotypes in a “new” environment may have, in part, affected their overall fitness in this study (Hufford and Mazer Reference Hufford and Mazer2003). Consequently, future studies should test for fitness differences between local and nonlocal L. benzoin or other native shrub ecotypes in L. maackii removal and restoration studies.
Future studies should also test the efficacy of different native plant herbaceous seed mixtures and/or different species and restoration planting densities of forest understory native shrubs to suppress L. maackii reinvasion over the long term, as Hopfensperger and colleagues (2019) recently found substantial seedling reinvasion, despite a low seedbank, 5 to 9 yr after removal and restoration efforts. Moreover, L. maackii seedlings are known to readily establish across a variety of soil types, temperatures, and light conditions (Hidayati et al. Reference Hidayati, Baskin and Baskin2000; Luken and Goessling Reference Luken and Goessling1995). Of note, Florey and Clay (Reference Flory and Clay2006, Reference Flory and Clay2009) found that L. maackii seedling germination success and density increased in early to mid-successional forests, and the present study was conducted in an early secondary successional forest that is likely to experience a persistent influx of L. maackii seed (from neighboring populations and bird dispersal), with subsequent seedling establishment and reinvasion likely to be a constant threat in the forest understory. Therefore, more studies are needed to evaluate various restoration planting strategies (herbaceous and native species seeding and planting densities) to suppress L. maackii seedling establishment and mitigate future reinvasion threats.
Acknowledgments
This study and article represent the efforts of three undergraduate students—MC, EW, AG—as part of their undergraduate senior research project. We are grateful for the statistical expertise, advice, and assistance provided by P Stephens, Ph.D (Asbury University) and the helpful feedback and suggestions from anonymous reviewers that greatly helped improve this article.
Competing interests
No competing interests have been declared.
Funding
Financial support for this research was graciously provided by the Walt and Rowena Shaw School of Science, Asbury University.


 Open access
Open access