


Introduction
The Ross Sea region, commonly defined as the region between 160°W and 170°E and south of 60°S, encompasses the largest continental shelf and most productive region of the Southern Ocean (Smith et al. Reference Smith, Nelson, Di Tullio and Leventer1996, Pinkerton et al. Reference Pinkerton, Boyd, Deppeler, Hayward, Hofer and Moreau2021) and supports rich and complex ecosystems that have been the focus of ecological study for decades (Smith et al. Reference Smith, Ainley and Cattaneo-Vietti2007, Reference Smith, Ainley, Cattaneo-Vietti, Hofmann, Rogers, Johnston, Murphy and Clarke2012, Pinkerton et al. Reference Pinkerton, Bradford-Grieve and Hanchet2010). In 2008, the Ross Sea region was recognized as the ocean region likely to have been least affected by human activity in the world (Ainley Reference Ainley2007, Halpern et al. Reference Halpern, Walbridge, Selkoe, Kappel, Micheli and D’Agrosa2008) and one of the few continental shelf ecosystems with its full complement of piscine and air-breathing predators (e.g. Schipper et al. Reference Schipper, Chanson, Chiozza, Cox, Hoffmann and Kataryia2008, Ainley Reference Ainley2009). Since 1997, the Antarctic toothfish (Dissostichus mawsoni Norman) has supported an longline fishery in the Ross Sea region managed by the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR) with a mandate to protect dependent and associated species and ensure potential reversibility of the effects of fishing within 20–30 years (CCAMLR 1982). Although the Ross Sea toothfish fishery is certified to the Marine Stewardship Council’s (MSC) global standard for a well-managed and sustainable fishery (Marine Stewardship Council 2021), the potential for ecosystem impacts from the fishery remains controversial (Abrams Reference Abrams2014, Abrams et al. Reference Abrams, Ainley, Blight, Dayton, Eastman and Jacquet2016, Ainley et al. Reference Ainley, Crockett, Eastman, Fraser, Nur and O’Brien2017).
The Antarctic toothfish (hereafter ‘toothfish’, noting that its congener, the Patagonian toothfish Dissostichus eleginoides Smitt, is generally found north of the Antarctic Convergence (Hanchet et al. Reference Hanchet, Rickard, Fenaughty, Dunn and Williams2008) and is not known from the study area) is the largest teleost in Antarctic waters, growing up to 2.1 m in length, weighing up to 120 kg and living up to ~50 years (Gon & Heemstra Reference Gon and Heemstra1990, Hanchet et al. Reference Hanchet, Rickard, Fenaughty, Dunn and Williams2008, Reference Hanchet, Dunn, Parker, Horn, Stevens and Mormede2015b). It is both an important predator in the ecosystem (Pinkerton et al. Reference Pinkerton, Bradford-Grieve and Hanchet2010, Pinkerton & Bradford-Grieve Reference Pinkerton and Bradford-Grieve2014) and a prey species for Weddell seals Leptonychotes weddellii (Lesson), ecotype-C killer whales Orcinus orca (Linnaeus) and sperm whales Physeter macrocephalus Linnaeus (Yukhov Reference Yukhov1971, Pinkerton et al. Reference Pinkerton, Dunn and Hanchet2008, Ainley & Siniff Reference Ainley and Siniff2009, Torres et al. Reference Torres, Pinkerton, Pitman, Durban and Eisert2013, Ainley et al. Reference Ainley, Cziko, Nur, Rotella, Eastman and Larue2020). Large colossal squid Mesonychoteuthis hamiltoni Robson are also known to depredate longline-caught toothfish and possibly ‘weakened and dying individuals’ (Remelso et al. Reference Remelso, Yukhov, Bolstad and Laptikhovsky2019), while toothfish are known to consume portions of large colossal squid, probably either ‘torn from living animals or scavenged from the seafloor’ (Roberts et al. Reference Roberts, Xavier and Agnew2011).
Understanding the trophic connections of toothfish is needed to anticipate potential effects of fishing and climate change on this species (Pinkerton & Bradford-Grieve Reference Pinkerton and Bradford-Grieve2014, Abrams et al. Reference Abrams, Ainley, Blight, Dayton, Eastman and Jacquet2016, Caccavo et al. Reference Caccavo, Christiansen, Constable, Ghigliotti, Trebilco and Brooks2021, McCormack et al. Reference McCormack, Melbourne-Thomas, Trebilco, Griffith, Hill and Hoover2021). Climate change is affecting primary productivity in the Ross Sea region (Pinkerton et al. Reference Pinkerton, Boyd, Deppeler, Hayward, Hofer and Moreau2021), with potential effects through the food web that are potentially synergistic with fishing (Kirby et al. Reference Kirby, Beaugrand and Lindley2009).
The diet of toothfish on the continental slope and oceanic features of the Ross Sea region (where the majority of the toothfish biomass resides; Hanchet et al. Reference Hanchet, Dunn, Parker, Horn, Stevens and Mormede2015b, Grüss et al. Reference Grüss, Pinkerton, Mormede and Devine2024) is relatively well-studied (e.g. Fenaughty et al. Reference Fenaughty, Stevens and Hanchet2003, Kokorin Reference Kokorin2010, Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014). In these regions, toothfish are largely adaptive piscivores, with icefish (mainly Chionobathyscus dewitti Andriashev & Neyelov) and grenadiers (Macrourus caml McMillan, Iwamoto, Stewart & Smith, and M. whitsoni (Regan)) dominating the diet on the continental slope and Macrourus spp. and Antimora rostrata (Günther) dominating the diet on oceanic features (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014). Toothfish are likely to consume 64% of the annual production of medium-sized demersal fish (dominated by macrourids and icefish) and probably exert strong top-down control on piscine prey species (Pinkerton & Bradford-Grieve Reference Pinkerton and Bradford-Grieve2014). Squid are also important prey items for toothfish in the Ross Sea region, in particular Psychroteuthis glacialis Thiele on the continental slope and Moroteuthopsis (formerly Kondakovia) longimana (Filippova) on oceanic features (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014).
The diet of toothfish on the Ross Sea continental shelf is more poorly understood than in more heavily fished areas of the Ross Sea region and is expected to differ from that in other parts of the Ross Sea region for two reasons. First, there is pronounced geographical variation in the age and size of toothfish within the Ross Sea region: subadult individuals (aged up to ~10 years) dominate the continental shelf and larger/older individuals are located further north on the slope and oceanic features (Hanchet et al. Reference Hanchet, Rickard, Fenaughty, Dunn and Williams2008, Grüss et al. Reference Grüss, Pinkerton, Mormede and Devine2024). Second, the potential prey species present over the continental shelf also vary geographically.
Knowledge of the diet of toothfish over the Ross Sea continental shelf is important given the potential for fishing to lead to trophic cascades. Several studies (including Ainley et al. Reference Ainley, Nur, Eastman, Ballard, Parkinson, Evans and Devries2013, Reference Ainley, Crockett, Eastman, Fraser, Nur and O’Brien2017, Reference Ainley, Morandini, Salas, Nur, Rotella and Barton2024, Lyver et al. Reference Lyver, Barron, Barton, Ainley, Pollard and Gordon2014) have hypothesized that depletion of toothfish by fishing led to a predation release of Antarctic silverfish (Pleuragramma antarcticum Boulenger) over the continental shelf, and that this, in turn, led to an increase in Adélie penguin (Pygoscelis adeliae (Hombron & Jacquinot)) numbers. Antarctic silverfish (hereafter silverfish) are an important prey for several top predators (La Mesa et al. Reference La Mesa, Eastman and Vacchi2004), and previous studies of the diet of toothfish caught below sea ice in McMurdo Sound have pointed to a dominance of silverfish (Eastman Reference Eastman1985b, Parker et al. Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019). Theoretical and empirical studies show that the effects of fishing on the ecosystem as a whole are dependent on the food-web structure, especially the pattern of trophic connections in the system (Pace et al. Reference Pace, Cole, Carpenter and Kitchell1999, Brose et al. Reference Brose, Berlow and Martinez2005, Pascual & Dunne Reference Pascual and Dunne2006). Based on ecosystem modelling, Pinkerton et al. (Reference Pinkerton, Lyver, Stevens, Forman, Eisert and Mormede2016) suggested that the predation release of silverfish because of fishing on toothfish was not likely to be strong enough to explain observed changes in Adélie penguin populations, although this conclusion was disputed (Ainley et al. Reference Ainley, Crockett, Eastman, Fraser, Nur and O’Brien2017). The importance of silverfish in the diet of toothfish on the Ross Sea continental shelf is an important factor in this debate, but there have been no detailed studies on the diet of toothfish over the wider Ross Sea continental shelf published in the peer-reviewed literature.
Toothfish diet has been studied in McMurdo Sound, at the south-western corner of the Ross Sea continental shelf, based on the opportunistic examination of Weddell seal prey (Calhaem & Christoffel Reference Calhaem and Christoffel1969), a specific feeding study by Eastman (Reference Eastman, Siegfried, Condy and Laws1985a,Reference Eastmanb) and a comparison of sea ice-based and vessel-based diets in McMurdo Sound (Parker et al. Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019). These studies caught toothfish on or near the sea floor through holes cut in fast sea ice. Calhaem & Christoffel (Reference Calhaem and Christoffel1969) obtained several toothfish (from 6.8 to 29.5 kg in weight) that had been captured by a Weddell seal, and the toothfish stomachs contained numerous silverfish. Eastman (Reference Eastman, Siegfried, Condy and Laws1985a,Reference Eastmanb) captured 58 toothfish (~93–166 cm total length (TL), 10–60 kg) on setlines at 300–500 m depth, and the toothfish diet was predominantly silverfish (71% occurrence and 89% dry weight), with some mysids (Antarctomysis spp.), icefish and other notothens (Pagothenia borchgrevinki (Boulenger) and Trematomus loennbergii Regan). Parker et al. (Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019) captured 182 toothfish (93–166 cm TL) on setlines through ice in McMurdo Sound at 518–588 m depths and observed a similarly silverfish-dominated diet (99% of stomachs). Silverfish have thus been considered characteristic (even dominant) in toothfish diet over the Ross Sea continental shelf, whereas silverfish are absent from toothfish stomachs in fish captured on the slope and oceanic features to the north (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014).
Since January 2012, an annual summer longline survey has been conducted in the southern part of the Ross Sea continental shelf to monitor the abundance of sub-adult toothfish (< 100 cm TL) in order to understand and monitor variability in toothfish recruitment as an abundance index for the Ross Sea stock assessment (Fig. 1; Devine Reference Devine2022). The survey was designed to cover the areas of highest abundance of juvenile toothfish on the continental shelf, based on previous fishery data.
Locality map showing the Core (A–C) and experimental (D, E, I, K, N, O) stratum boundaries and the locations of non-empty stomach samples for the 2012–2015 sub-adult surveys. Inset shows the position of the survey area (black lines) in relation to the General protection Zone, Special Research Zone and Krill Research Zone (grey shading) within the Ross Sea region Marine Protected Area, which came into enforcement in 2017. Lines indicate the position of the 600, 1000 and 2000 m isobaths. Colours indicate the year of sampling.

Figure 1 Long description
The inset at the top right shows the Ross Sea region with the survey area outlined in black, overlaid on the General Protection Zone, Special Research Zone and Krill Research Zone (grey shading). The main map below details the survey area, divided into labeled strata: A, B, C (core), and D, E, I, K, N, O (experimental). Stratum boundaries are marked with black lines. Sample locations are shown as dots colored by year: black for 2012, magenta for 2013, brown for 2014, and blue for 2015. Strata A, B, D, E, I, K, N, and O contain varying densities of sample points, with the highest concentration in strata A, B, and E. Isobaths at 600, 1000, and 2000 meters are indicated by thin contour lines. The map is oriented with latitude and longitude gridlines, and the legend at the lower left explains the color coding for years.
In this study, we use toothfish stomachs collected from four consecutive years of this survey series (2012–2015) to describe in detail the diet of toothfish on the continental shelf of the southern Ross Sea, with a focus on three subregions.
Materials and methods
Survey area
The survey comprised a Core area on the south-west continental shelf to monitor the abundance of sub-adult (< 100 cm TL) toothfish (strata A–C, 500–900 m depths). The Core area was extended during the first two surveys to examine the extent of sub-adult toothfish outside the core strata, with stratum D (400–500 m depths) sampled in 2012 and 2013 and stratum E (300–400 m depths) sampled in 2012. There were also two additional survey areas: outer McMurdo Sound (strata I and N, sampled in 2014, 600–1000 m depths) and Terra Nova Bay (strata K and O, sampled in 2015, 600–1200 m depths); these were assessed in order to monitor the abundance and size distribution of toothfish in areas of importance to predators (Fig. 1). The survey area is within the Ross Sea region Marine Protected Area (MPA), which came into effect in December 2017 (Fig. 1). For this paper, ‘outer McMurdo Sound’ is defined as the area beyond the fast ice and is distinguished from ‘inner McMurdo Sound’, which is predominantly under fast ice.
Stomach samples
Toothfish stomach samples were obtained from four surveys conducted in January–February 2012 (Hanchet et al. Reference Hanchet, Mormede, Parker and Dunn2012), 2013 (Parker et al. Reference Parker, Hanchet, Mormede, Dunn and Sarralde2013), 2014 (Mormede et al. Reference Mormede, Parker, Hanchet, Dunn and Gregory2014) and 2015 (Hanchet et al. Reference Hanchet, Sharp, Mormede, Parker and Vacchi2015a). Toothfish were captured by the bottom longliner FV San Aotea II at depths from 328 to 1205 m using size 15/O EZ-Baiter hooks baited with Chilean squid (Dosidicus gigas (A. d’Orbigny)). Toothfish were sampled from each haul, measured, weighed, sexed and macroscopically staged, and their stomachs were excised, placed in labelled plastic bags and frozen at −20°C.
In the laboratory, each stomach was examined and graded on a qualitative scale according to its fullness as empty, trace, part full or full. Individual food items were identified to the lowest possible taxonomic level, counted and weighed. The state of prey digestion was recorded qualitatively as fresh, slightly digested, moderately digested or digested. Prey items were identified using reference material and identification guides: fish (Gon & Heemstra Reference Gon and Heemstra1990), otoliths (Williams & McEldowney Reference Williams and McEldowney1991, Reid Reference Reid1996), cephalopod beaks (Xavier & Cherel Reference Xavier and Cherel2021) and crustaceans (Xavier et al. Reference Xavier, Cherel, Boxshall, Brandt, Coffer and Forman2020).
Analysis of diet composition
The composition of the diet was described by the numerical importance (%N), frequency of occurrence (%F), weight (%W) and percentage index of relative importance (%IRI; Pinkas et al. Reference Pinkas, Oliphant and Iverson1971). Although there are known problems with %IRI and basic descriptors may be preferred (Ahlbeck et al. Reference Ahlbeck, Hansson and Hjerne2012, Baker et al. Reference Baker, Buckland and Sheaves2013), we included this metric to allow for comparison with Stevens et al. (Reference Stevens, Dunn, Pinkerton and Forman2014). In the analysis of diet variability, we excluded 1) prey classed as digested or unidentifiable, 2) scavenging amphipods and isopods (considered to be ingested with larger depredated fish they were feeding on or from toothfish that had died on the longline) and 3) parasites (including gnathiid praniza isopods, sphyrion copepods, anisakid nematodes and trematodes). Large amphipods and isopods were considered legitimate prey. Prey were then aggregated into taxonomic categories at the family level or higher.
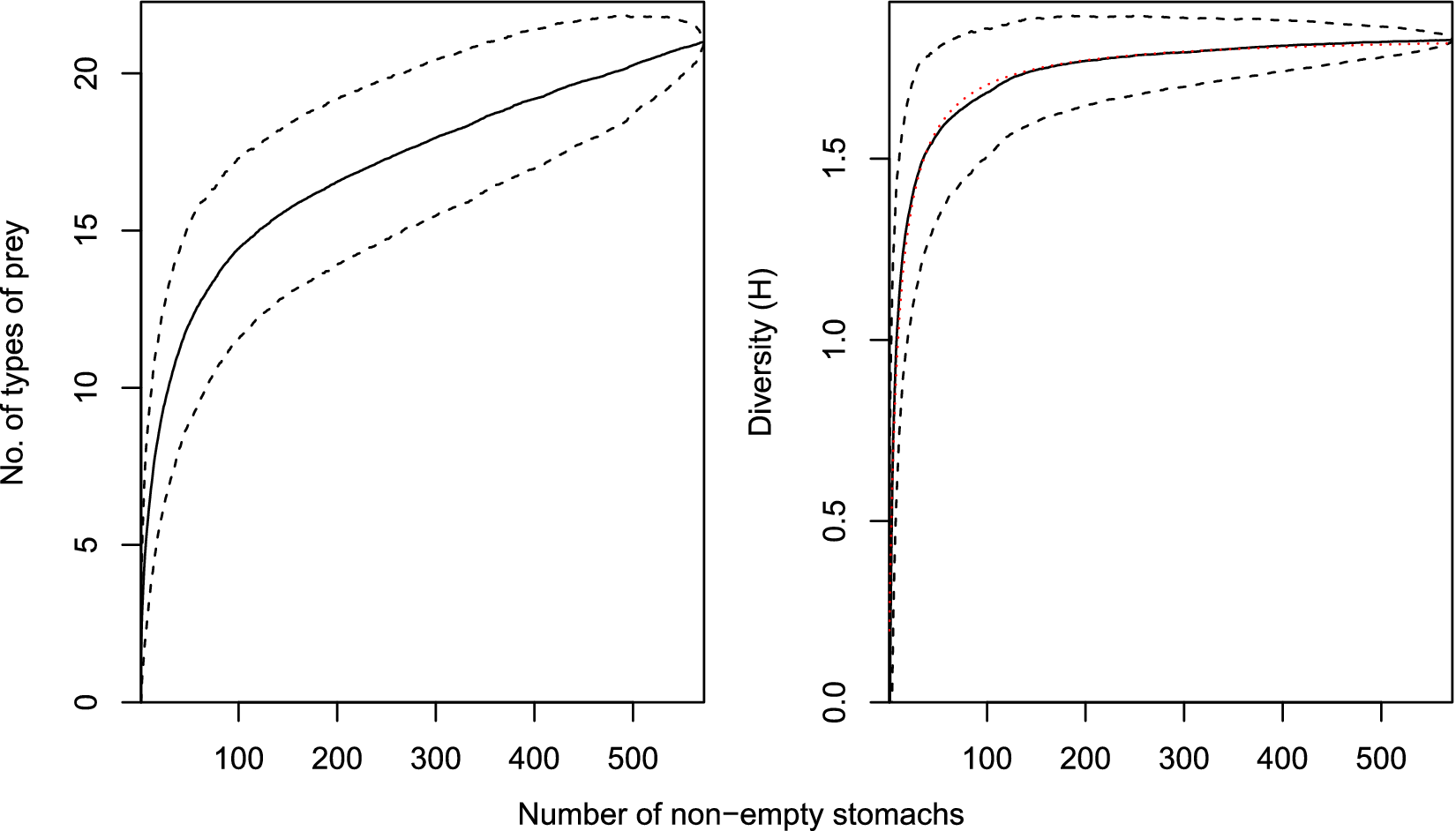
To assess the adequacy of the samples, the cumulative number and cumulative diversity (Brillouin index of diversity, H) of categorized stomach contents were plotted against the cumulative number of stomachs containing food. The mean cumulative curve and 95% credible intervals were estimated from 1000 curves based upon different random orders of the stomachs. The total sample size (N) was considered adequate if the mean sample diversity was at least 95% of the asymptotic diversity estimated from a fitted curve of the form H = aN(1 + bN)−1 (Dunn Reference Dunn2009).
Multivariate analyses using PRIMER v7 (Clarke & Gorley Reference Clarke and Gorley2015) were conducted to identify the differences between samples and factors affecting diet composition. The prey were first categorized, then prey weight was standardized and square-root transformed, and a dissimilarity matrix was calculated using Bray-Curtis distances. ANOSIM (analysis of similarities) was used to determine which samples or groups of stomachs were significantly different from each other, followed by SIMPER (similarity percentages) to identify the prey categories characteristic of each group. The mean of normalized prey weight for the prey categories identified by SIMPER was then calculated to show mean differences in diet composition between groups.
Distance-based linear model (DistLM) analysis in PRIMER v7 (Anderson et al. Reference Anderson, Gorley and Clarke2015) was used to identify which predictors explained a significant proportion of the variability in diet composition. The predictors investigated were fish length, weight, sex, macroscopic maturity stage, fishing depth, year, stratum (combination of year and region: Core 2012, Core 2013, Core 2014, McMurdo 2014, Core 2015, TerraNova 2015), latitude and longitude. We did not use environmental covariates because we wanted to focus on regional (spatial) variations in diet rather than year-to-year changes, which would probably dominate the environmental variability at the scale of the study. The results of the DistLM analysis were a marginal test, fitting each predictor individually, and a conditional test, fitting each predictor conditional on the predictor(s) already in the model. The most significant predictors in the conditional tests were selected using the ‘best’ selection method (Dunn Reference Dunn2009) and the Akaike information criterion (AIC), with a more complex model only being accepted if the additional predictor explained at least 0.5% of additional deviance. Continuous variables selected by DistLM were binned, with bin limits chosen so that the number of observations in each bin was approximately equal. This was considered objective given that there were no a priori known biologically meaningful boundaries for these predictors, and it prevented bins containing small, and so potentially biased, samples. The mean of normalized prey weight for the prey categories identified by SIMPER was then calculated to show mean differences in diet composition between binned groups.
Results
Sampling
Of 740 stomachs examined, 10.8% were empty, 20.7% contained a trace of prey, 45.2% were part full and 23.3% were full. Nine stomachs contained only parasites, scavenging crustaceans, bait or sand and rocks. Of the 651 stomachs containing prey, between 1 and 20 prey items were identified per stomach, most of the stomachs (80.0%) contained fewer than 5 prey items and 27.5% contained just a single item. There were 18 stomachs containing only well-digested prey items, 1 containing only unidentifiable cephalopod remains and 60 containing only unidentifiable fish, leaving 572 (77.3%) for detailed analyses of diet composition.
The diversity of prey categories by sample approached the asymptote in all samples (79.1–95.8% of the estimated asymptote), with outer McMurdo Sound in 2014 being the least completely sampled in terms of being the furthest from asymptotic prey diversity (Table I). Although new types of prey category were steadily encountered with increasing sample size, each new prey type was increasingly rare, and consequently the diversity of prey categories more rapidly reached an asymptote, being 75% of the estimated asymptote after 24 stomachs, 85% after 51 stomachs and 95% after 156 stomachs (Fig. 2).
Mean of standardized percentage prey weight within stomach samples of toothfish from the Ross Sea shelf. Prey types shown are those that contributed the most to 95% of the SIMPER (similarity percentages) within-sample similarity for one or more samples. Note that silverfish (Pleuragramma antarcticum) did not contribute to 95% of the SIMPER and so are not included separately in this table but are included in the ‘All Nototheniidae’ group.

Table I. Long description
Column headers list strata: Core (2012, 2013, 2014, 2015), McMurdo Sound (2014), Terra Nova Bay (2015). Rows list prey types: Crangonidae, Amphipoda, Megaleledonidae, Liparidae, Bathydraconidae, Artedidraconidae, Trematomus species, Channichthyidae, Scavenged, All Nototheniidae (including Trematomus species and Pleuragramma antarcticum). For Core, Trematomus species have the highest mean percentage prey weight, increasing from 32.2 in 2012 to 52.8 in 2015. Channichthyidae is next highest, ranging from 22.8 to 31.0 except 30.4 in 2012 and 26.4 in 2015. All Nototheniidae group rises from 62.6 in 2012 to 79.2 in 2015. McMurdo Sound 2014 shows Trematomus species at 42.8, Channichthyidae at 34.8, All Nototheniidae at 77.6. Terra Nova Bay 2015 shows Channichthyidae highest at 58.7, Trematomus species at 31.1, All Nototheniidae at 89.8. Other prey types have lower values, mostly below 10. Sample sizes (n) range from 43 to 150. Mean total weights (Wt) and interquartile ranges are given for each stratum-year, e.g., Core 2012 is 17.6 kilograms (7.0–27.0). Percentage prey diversity (%PD) ranges from 79.1 to 95.8 across strata and years.
%PD = sample total prey diversity as a percentage of the estimated asymptotic prey diversity; n = sample size; Wt = mean total toothfish weight in kilograms (interquartile range).
The mean cumulative number of prey categories and diversity of prey categories (measured using the Brillouin index of diversity, H) for all samples combined. Dashed lines indicate 95% credible intervals. Dotted line is the fitted diversity curve from which asymptotic diversity was estimated.
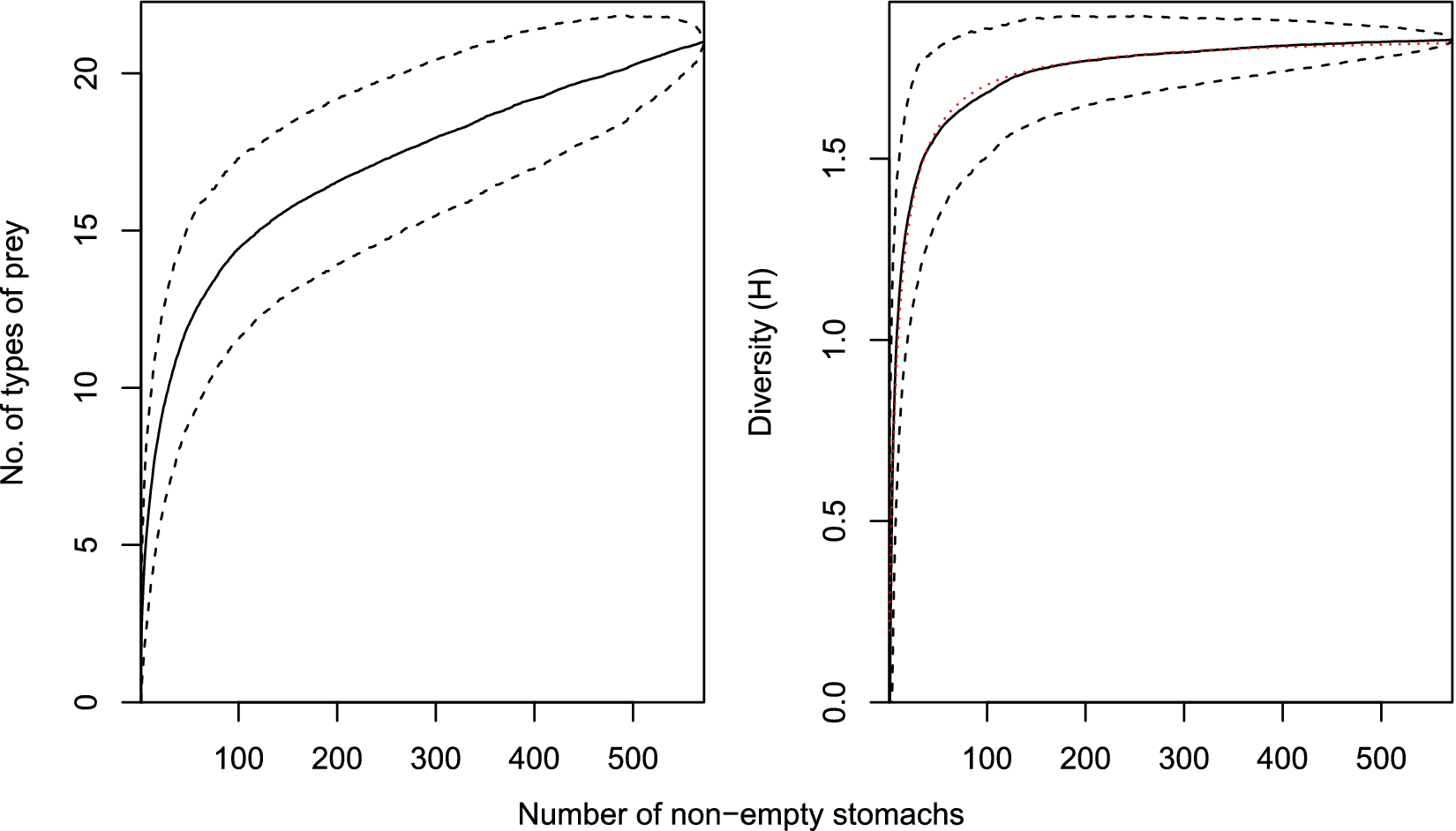
Figure 2 Long description
The left panel plots number of types of prey on the y-axis, ranging from 0 to 20, against number of non-empty stomachs on the x-axis, ranging from 0 to 500. A solid line shows a steep initial rise in prey types, then a gradual asymptotic increase. Dashed lines above and below the solid line indicate the 95 percent credible intervals, widening at lower sample sizes and narrowing as sample size increases. The right panel plots diversity H on the y-axis, ranging from 0 to 2, against number of non-empty stomachs on the x-axis, also from 0 to 500. The solid line shows a rapid initial increase in diversity, then levels off asymptotically. Dashed lines above and below the solid line represent the 95 percent credible intervals. A dotted line closely follows the solid line, representing the fitted diversity curve used to estimate asymptotic diversity. Both panels show that increases in sample size yield diminishing returns in new prey types and diversity, with curves flattening at higher sample sizes.
The mean weight of individual toothfish was significantly different between strata (ANOVA (analysis of variance) on log(weight), P = 0.005). Toothfish sampled in the Core area in 2015 were significantly smaller than those from the Core area in 2014 (t-test on log(weight), Benjamini-Hochberg corrected, P = 0.025), Terra Nova Bay in 2015 (P = 0.025) and outer McMurdo Sound in 2014 (P = 0.026).
Overall diet
In the core survey strata, the most commonly identified notothen prey species was T. loennbergii, which occurred in ~6–37% of stomachs (%F) and accounted for ~3–35% of total prey weight (%W) in 2012–2015 (Table II). The most important icefish prey were Chionodraco spp., of which Chionodraco hamatus (Lönnberg) was the most frequently identified, and Neopagetopsis ionah Nybelin. Silverfish and Trematomus scotti (Boulenger) occurred in ~2–10% and ~1–21% of stomachs, respectively, but they weighed little and so had little contribution by weight or %IRI. Two deep-water macrourid prey (Macrourus caml and Macrourus whitsoni) were identified from one toothfish stomach and may indicate movement of the toothfish into the survey area from deeper water. Dragonfish (Bathydraconidae) were rarely observed, except in 2012, when Gymnodraco acuticeps Boulenger occurred in ~6% of stomachs and accounted for ~11% of prey weight. The prawn Notocrangon antarcticus (Pfeffer) and amphipod Eusiris spp. were found in ~15–24% and ~3–6% of stomachs, respectively, but were small and contributed little to weight or %IRI. Most of the remaining prey were generally small, benthic species. Unidentified fish accounted occurred in ~36–55% of stomachs, but they were often partially or well-digested and so only accounted for ~4–7% of total prey weight.
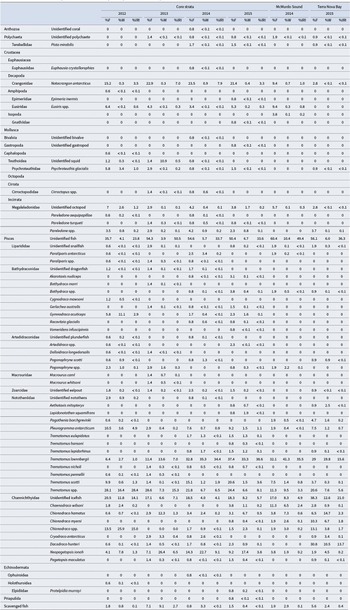
Comparison of the percentage occurrence (%F), percentage by weight (%W) and percentage by index of relative importance (%IRI) of prey items in the diet of Dissostichus mawsoni by sample location and year.
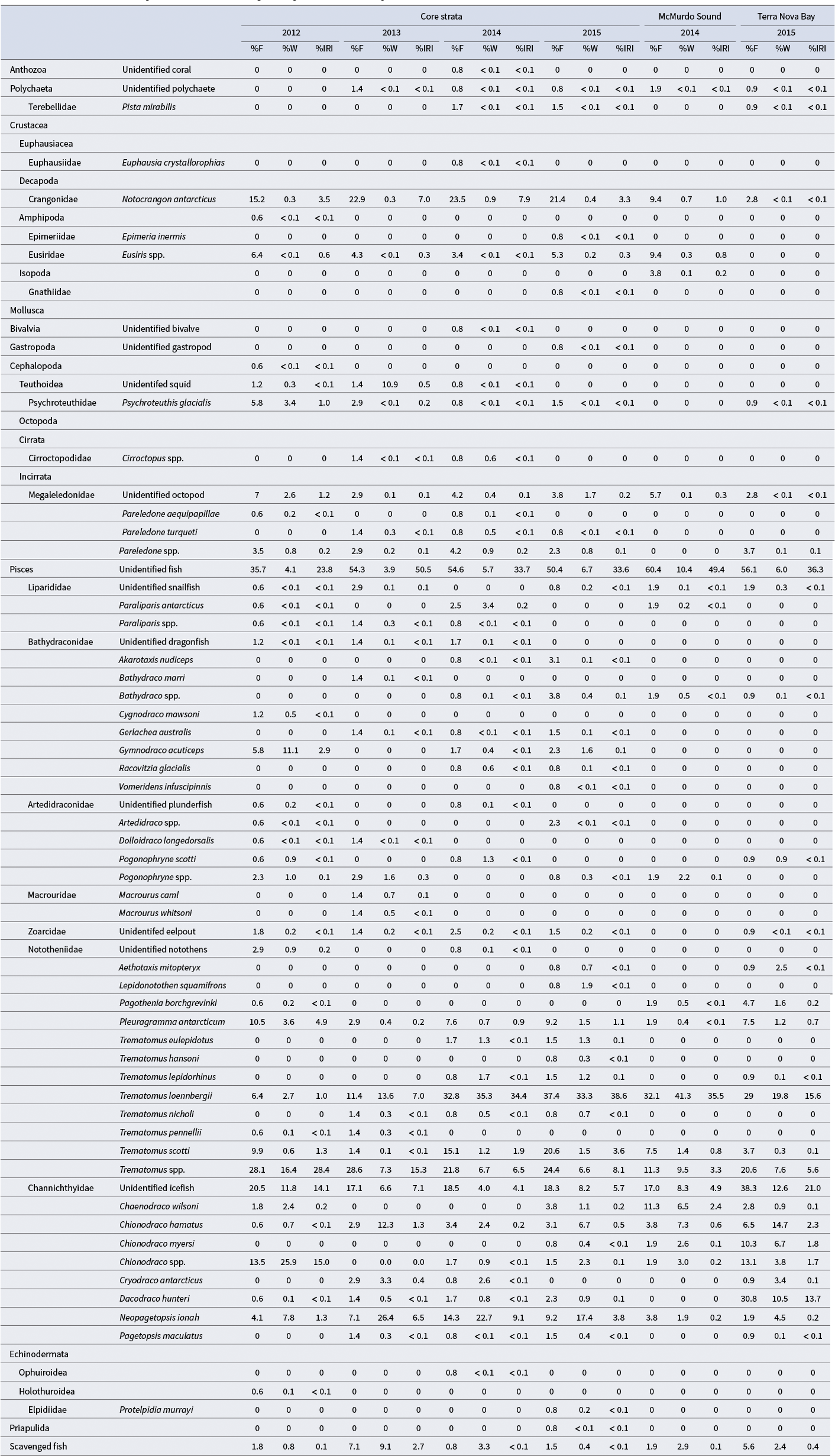
Table II. Long description
The table lists prey taxa in the diet of Dissostichus mawsoni, organized by taxonomic group and species in the first four columns. For each prey item, data are presented for Core strata (2012, 2013, 2014, 2015), McMurdo Sound (2014, 2015), and Terra Nova Bay (2014, 2015). Each location-year group has three columns: percent frequency of occurrence (%F), percent by weight (%W), and percent index of relative importance (%IRI). For example, unidentified fish in Core strata 2012 show 35.7 %F, 4.1 %W, 23.8 %IRI; in Core strata 2013, 54.3 %F, 3.9 %W, 50.5 %IRI; in Core strata 2014, 54.6 %F, 5.7 %W, 33.7 %IRI; and in Core strata 2015, 50.4 %F, 6.7 %W, 33.6 %IRI. Trematomus loennbergii in Core strata 2014 has 32.8 %F, 35.3 %W, 34.4 %IRI, and in Terra Nova Bay 2015, 29 %F, 19.8 %W, 15.6 %IRI. Chionodraco spp. in Core strata 2012 has 13.5 %F, 25.9 %W, 15.0 %IRI, and in Terra Nova Bay 2015, 13.1 %F, 3.8 %W, 1.7 %IRI. Channichthyidae (unidentified icefish) in Core strata 2012 has 20.5 %F, 11.8 %W, 14.1 %IRI, and in Terra Nova Bay 2015, 38.3 %F, 12.6 %W, 21.0 %IRI. Most invertebrate taxa have low or zero values across all locations and years. The table highlights that fish, especially Trematomus spp., Chionodraco spp., and Channichthyidae, are the dominant prey, with high values for all three metrics across multiple years and locations. Lower values or zeros are common for Anthozoa, Polychaeta, Mollusca, and Echinodermata.
The diet in outer McMurdo Sound was similar to the Core survey area, with T. loennbergii being the most important prey species, Chaenodraco wilsoni Regan and C. hamatus being the most commonly identified icefish prey species and silverfish being rarely identified (Table II). The cryopelagic notothen P. borchgrevinki was found in one stomach but was not identified in the Core survey area. The prawn N. antarcticus was less commonly observed than in the Core survey area, whereas the large amphipod Eusiris spp. was more commonly observed.
In Terra Nova Bay, icefish were more important than notothens in the diet (Table II). Seven icefish species were identified, and the otherwise rarely sampled Dacodraco hunteri Waite was the most common identified icefish species. T. loennbergii and Chionodraco spp. were also important prey, whereas silverfish was relatively rare. The ice-associated notothen P. borchgrevinki was found in five stomachs and the pelagic notothen Aethotaxis mitopteryx DeWitt was found in one stomach.
Variability in diet
ANOSIM showed that the diet of toothfish from Terra Nova Bay in 2015 was significantly different from diets in other strata/years (P = 0.001 for all pairwise comparisons) except the Core area in 2012 (P = 0.360). The diet from the Core area in 2013 was significantly different from those of the Core area in 2014 and 2015 (ANOSIM, P = 0.038 and P = 0.004, respectively). No other significant differences were detected. The main species contributing to the dissimilarity between samples are reported in Table I, with Terra Nova Bay in 2015 notably having a relatively high proportion of Channichthyidae. An increasing trend in the percentage of prey weight attributed to the two dominant fish prey categories (Channichthyidae and Trematomus spp.; Table I) was not likely to be attributable to a decrease in unidentified fish (i.e. to improvements in fish prey identification), as unidentified fish also increased (see Table II) in accordance with there being an overall increase in fish prey over time. Compared to the Core area 2014 and 2015 samples, the Core area 2013 sample had a relatively low proportion of Channichthyidae and Trematomus spp. and a high proportion of Crangonidae and scavenged prey (Table I).
The best model from the DistLM analysis attributed 3.3% of the variability in prey composition to latitude, followed by 1.3% to year and 1.1% to fish weight (total 5.7%), leaving most of the variability in diet unexplained. The latitude predictor captured the difference between the more northern Terra Nova Bay 2015 sample and the more southern Core area and outer McMurdo Sound samples. The year effect was also identified by ANOSIM (see above). After binning the toothfish weights, the only significant difference in diet between toothfish weight classes identified using ANOSIM was between the smallest (2.5–6.0 kg) and largest (> 24.5 kg) size classes (P = 0.001). The smaller fish notably contained greater proportions of Trematomus spp. and Crangonidae and lesser proportions of Channichthyidae than larger fish (Table III).
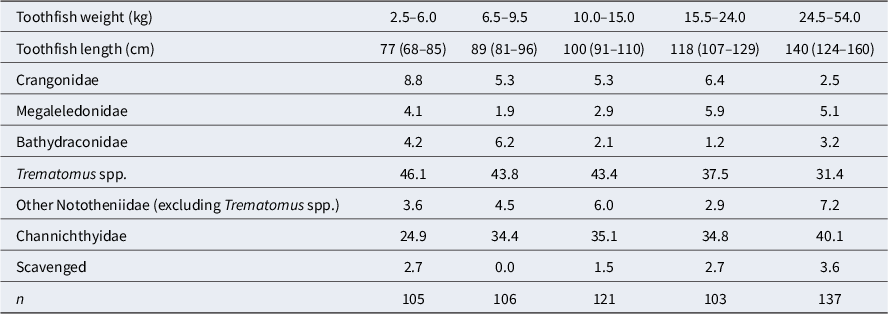
Mean of standardized percentage prey weight within stomach samples of toothfish from the Ross Sea shelf by toothfish weight bin (kg). Prey types shown are those that contributed the most to 95% of the SIMPER (similarity percentages) within-sample similarity for one or more samples. The length composition of the weight groups is shown (median and 95% interval). Nototheniidae excludes prey identified as Trematomus spp. Note that silverfish (Pleuragramma antarcticum) did not contribute to 95% of the SIMPER and so are not included separately in this table but are included in the ‘Other Nototheniidae’ group.
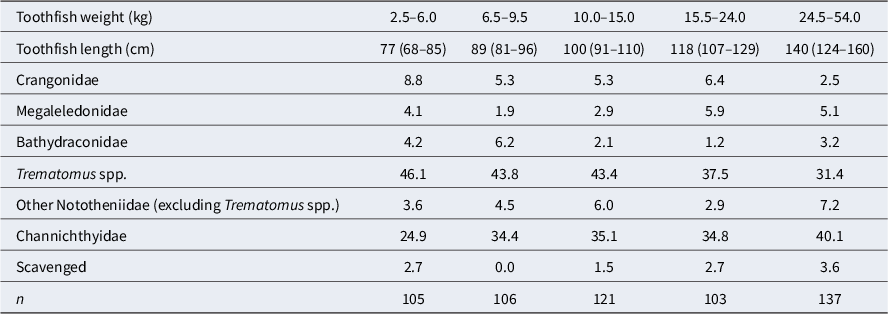
Table III. Long description
Starting from the leftmost column, toothfish weight bins are 2.5–6.0, 6.5–9.5, 10.0–15.0, 15.5–24.0, and 24.5–54.0 kilograms. The first row lists median toothfish lengths and 95 percent credible intervals: 77 (68–85), 89 (81–96), 100 (91–110), 118 (107–129), and 140 (124–160) centimeters. For Crangonidae, mean prey weights are 8.8, 5.3, 5.3, 6.4, and 2.5 percent. Megaleledonidae values are 4.1, 1.9, 2.9, 5.9, and 5.1 percent. Bathydraconidae values are 4.2, 6.2, 2.1, 1.2, and 3.2 percent. Trematomus species values are 46.1, 43.8, 43.4, 37.5, and 31.4 percent. Other Nototheniidae (excluding Trematomus) values are 3.6, 4.5, 6.0, 2.9, and 7.2 percent. Channichthyidae values are 24.9, 34.4, 35.1, 34.8, and 40.1 percent. Scavenged prey values are 2.7, 0.0, 1.5, 2.7, and 3.6 percent. Sample sizes (n) are 105, 106, 121, 103, and 137 for each bin respectively. Silverfish are included in the ‘Other Nototheniidae’ group.
n = sample size (number of toothfish stomachs).
Discussion
This study examined the diet of Antarctic toothfish on the Ross Sea continental shelf via an analysis of stomach contents. Although toothfish diet has been well-studied on the Ross Sea continental slope and on the features of the Pacific-Antarctic ridge to the north (Fenaughty et al. Reference Fenaughty, Stevens and Hanchet2003, Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014) and in the inner McMurdo Sound (Eastman Reference Eastman1985b, La Mesa et al. Reference La Mesa, Eastman and Vacchi2004, Parker et al. Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019), little has been previously published on the toothfish diet over the Ross Sea wider continental shelf. This is an important issue because of potential trophic cascades as the population size of toothfish is reduced by fishing (Ainley et al. Reference Ainley, Nur, Eastman, Ballard, Parkinson, Evans and Devries2013, Pinkerton et al. Reference Pinkerton, Lyver, Stevens, Forman, Eisert and Mormede2016). Sample sizes of non-empty toothfish stomachs were not large, but they were sufficient, as indicated by the diversity of prey approaching the asymptote diversity.
Toothfish diet over the Ross Sea continental shelf
This study found that the deep-water notothen T. loennbergii was the dominant toothfish prey in the Core survey area and outer McMurdo Sound and was second only to D. hunteri as a prey item in Terra Nova Bay. T. loennbergii is an epibenthic species that feeds predominantly on benthic prey (Eastman Reference Eastman, Siegfried, Condy and Laws1985a, La Mesa et al. Reference La Mesa, Vacchi, Castelli and Diviacco1997) and is therefore likely to be often associated with the seabed (La Mesa et al. Reference La Mesa, Eastman and Vacchi2004). The blackfin notothen T. scotti also feeds predominantly on benthic prey (Daniels Reference Daniels1981, Pakhomov Reference Pakhomov1997) and was a relatively frequent toothfish prey in this study.
Seven species of icefish were eaten by toothfish. Icefish were important prey and were likely to have been predated near or on the bottom. Based on buoyancy measurements (Eastman Reference Eastman2020), C. wilsoni, Chionodraco myersi DeWitt & Tyler (and possibly C. hamatus), Cryodraco antarcticus Dollo and Pagetopsis maculatus Barsukov & Permitin are thought to be demersal, while D. hunteri and N. ionah are thought to be semi-pelagic (occupying the water column and the substrate; Eastman Reference Eastman2020). Some species (e.g. C. hamatus, C. myersi and C. wilsoni) are thought to rest on the substrate when not feeding or while waiting for suitable prey, ascending periodically to feed near the bottom or in midwater (Kock & Jones Reference Kock and Jones2002, La Mesa et al. Reference La Mesa, Eastman and Vacchi2004). Nesting and parental guarding also occur in several icefish species, including those species frequently eaten by toothfish on the continental shelf (e.g. C. wilsoni, Kock et al. Reference Kock, Pshenichnov and Devries2006, Ziegler et al. Reference Ziegler, Smith, Edwards and Vernet2017; C. hamatus, Ferrando et al. Reference Ferrando, Castellano, Gallus, Ghigliotti, Masini, Pisano and Vacchi2014, Carlig et al. Reference Carlig, Di Blasi, Canese, Vacchi, Grant and Ghigliotti2024; N. ionah, Riginella et al. Reference Riginella, Pineda-Metz, Gerdes, Koschnick, Böhmer and Biebow2021, Purser et al. Reference Purser, Hehemann, Boehringer, Tippenhauer, Wege and Bornemann2022; Pagetopsis macropterus (Boulenger), La Mesa et al. Reference La Mesa, Piepenburg, Pineda-Metz, Riginella and Eastman2019). Resting on the substrate and nesting and parental guarding could make these icefish species particularly vulnerable to toothfish predation.
D. hunteri was an important prey species in Terra Nova Bay. It is not known to rest on the substrate, is thought to be a broadcast spawner and, based on a weakly ossified skeleton and buoyancy measurements, is regarded as semi-pelagic (Eastman Reference Eastman1999, 2020). D. hunteri larvae are one of the most abundant larval channichthyids (La Mesa et al. Reference La Mesa, Cali, Di Franco, Riginella, Donato and Russo2025); however, in contrast, adults are rarely captured, perhaps because they are often in the water column or in an area not sampled by research trawls. Both larval and adult D. hunteri appear to feed exclusively on silverfish (Eastman Reference Eastman1999, La Mesa et al. Reference La Mesa, Cali, Di Franco, Riginella, Donato and Russo2025). The importance of D. hunteri in the diet of toothfish in Terra Nova Bay (57 individuals, 30.8% frequency) may indicate some feeding by toothfish in the water column or close to the bottom, and that they may be aggregated in this region. However, they may be more abundant in deeper water (49 of 57 individuals were recovered from toothfish captured in 907–1205 m, x = 1065 m), a depth range not sampled in Core area strata (300–900 m). D. hunteri was not identified in the stomachs of toothfish captured in outer McMurdo Sound (600–1000 m).
Small stones were occasionally observed in stomachs, sometimes with appreciable amounts of fine silt or mud. The terebellid polychaete Pista mirabilis McIntosh was observed in five stomachs and a small ophiuroid in a further stomach. Substrate (stones, silt and mud) and benthic invertebrates may have been accidentally ingested while benthic feeding (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014) or, alternatively, the toothfish may have died on the longline and, with its mouth open, then been dragged along the seabed during retrieval, which forced benthic biota and substrate into the stomach. In some cases, these stomachs also contained numerous small scavenging amphipods, indicating the toothfish died prior to hauling the line.
Invertebrate prey were a smaller component of the toothfish diet, and their composition has been found to vary between the continental shelf, slope and oceanic features, probably reflecting latitudinal differences in habitat and potential prey (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014). On the continental shelf, the prawn N. antarcticus and large gammarid amphipods (primarily Eusiris spp.) were occasional prey. Mysids (Antarctomysis spp.) were a common prey in the diet of 58 toothfish captured through holes cut in fast sea ice in inner McMurdo Sound (Eastman Reference Eastman, Siegfried, Condy and Laws1985a,b), but they were not observed in toothfish stomachs in this study, although a small number of Antarctomysis mysids were recovered from toothfish stomachs on the 2018 Ross Sea shelf survey (J.S. Forman, unpublished data 2018). On the continental slope, Nematocarcinus prawns were occasional prey (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014). However, on a demersal trawl survey in the same area (Mawson and Iselin banks; O’Driscoll & Double Reference O’Driscoll and Double2015), Nematocarcinus lanceopes Spence Bate was commonly captured (915 individuals from 10 trawls at 1013–1990 m depths (Earth Sciences New Zealand, unpublished data 2015, New Zealand-Australia Antarctic Ecosystems Voyage 2015), despite using a rough bottom trawl probably not well-suited to catching benthopelagic prawns. This suggests that there may be some prey selection by toothfish. On oceanic features, king crabs (Paralomis spp.) were occasionally eaten (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014).
Not surprisingly, cephalopod prey also varied with locality, consistent with Stevens et al. (Reference Stevens, Dunn, Pinkerton and Forman2014). Octopods were occasional toothfish prey, but they have few diagnostic characters when partially digested; therefore, they were often not identified to species. However, small incirrate octopuses (probably mainly Pareledone species) were more common prey on the continental shelf, the large, robust cirrate Cirroctopus spp. was widespread and the delicate, gelatinous cirrate Stauroteuthis sp. seemed to be largely restricted to oceanic features. Squid were more common prey, and they have distinctive beaks, so they were often identified to species (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014). Glacial squid (P. glacialis) was the only squid species occasionally eaten on the continental shelf, but it was a relatively important prey on the continental slope, while the large onychoteuthid Moroteuthopsis longimana were relatively important prey on oceanic features. Other squid species were rare prey (Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014).
In general, the diet found here is consistent with predominantly demersal or benthic feeding of toothfish on the Ross Sea continental shelf. Benthic prey in the diet of toothfish on the shelf include notothens (T. loennbergii, T. scotti), probably icefish (C. wilsoni, C. myersi, C. antarcticus, P. maculatus), dragonfishes (Bathydraconidae), plunderfishes (Artedidraconidae), benthic prawns (N. antarcticus) and octopuses. However, we note the presence of cryopelagic prey (P. borchgrevinki) and potentially pelagic prey (P. antarcticum). That toothfish feed on a range of prey near the seabed is consistent with previous studies of this species on the Antarctic continental slope and oceanic features (Fenaughty et al. Reference Fenaughty, Stevens and Hanchet2003, Kokorin Reference Kokorin2010, Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014, Park et al. Reference Park, Yeon, Han, Lee, Hanchet and Baeck2015, Seong et al. Reference Seong, Choi, Chung, An, Kim and Baeck2021, Reference Seong, Chung, Kim, Kang, Jin and Baeck2023, Queirós et al. Reference Queirós, Stevens, Pinkerton, Rosa, Duarte and Baeta2022, Pérez-Pezoa et al. Reference Pérez-Pezoa, Cárdenas, González-Aravena, Gallardo, Rivero and Arriagada2023). These papers lead to a general picture of Antarctic toothfish as an opportunistic predator of predominantly benthic or demersal prey.
Silverfish as prey of toothfish
This study found that, in summer (January–February), silverfish was only a minor prey item (< 4% by weight) for toothfish in all three areas considered: the wider open continental shelf, outer McMurdo Sound and Terra Nova Bay. The low proportion of silverfish in the stomachs of toothfish is comparable with results obtained by Parker et al. (Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019) in the outer McMurdo Sound and in contrast with studies under shore fast ice in the inner McMurdo Sound (e.g. Eastman Reference Eastman1985b, Parker et al. Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019). In spring (October–November), the diet of toothfish caught under fast ice in the inner McMurdo Sound at 300–588 m depths was found to be dominated by silverfish (71% by occurrence, 89% by dry weight: Eastman Reference Eastman, Siegfried, Condy and Laws1985a,b; 99% occurrence: Parker et al. Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019). In the outer McMurdo Sound area in the present study, less than 2% of stomachs contained silverfish, and silverfish contributed less than 1% of total prey weight.
Silverfish are one of the only true pelagic fish of the Antarctic shelf, but they can occupy the whole water column (Gon & Heemstra Reference Gon and Heemstra1990, Vacchi et al. Reference Vacchi, La Mesa, Massimo and MacDonald2004, La Mesa & Eastman Reference La Mesa and Eastman2012). The depth where they occur varies with location, age/stage of individuals, season and possibly time of day, although the literature is conflicted on silverfish diel migration (Robison 2003, O’Driscoll et al. Reference O’Driscoll, Macaulay, Gauthier, Pinkerton and Hanchet2011, Reference O’Driscoll, Ladroit, Parker, Vacchi, Canese and Ghigliotti2018). In ice-free waters of the Ross Sea shelf, juvenile and small adult silverfish (< 10 cm) typically form diffuse layers shallower than 100 m (O’Driscoll et al. Reference O’Driscoll, Macaulay, Gauthier, Pinkerton and Hanchet2011), well above the seabed (~500–800 m). Eastman et al. (Reference Eastman2020) described silverfish to ‘hang in loose shoals in the water column’. However, larger adult silverfish (10–20 cm) are commonly found deeper than 200 m, sometimes close to the seabed (O’Driscoll et al. Reference O’Driscoll, Macaulay, Gauthier, Pinkerton and Hanchet2011), where they can form dense assemblages (O’Driscoll et al. Reference O’Driscoll, Ladroit, Parker, Vacchi, Canese and Ghigliotti2018, La Mesa et al. Reference La Mesa, Piepenburg, Pineda-Metz, Riginella and Eastman2019). The silverfish retrieved from toothfish stomachs in this study were mainly adults (150–223 mm standard length, n = 15) and may have been consumed near the seabed or in the water column.
Antarctic toothfish in the present study over the Ross Sea continental shelf weighing more than 12 kg (and so probably neutrally buoyant; Near et al. Reference Near, Russo, Jones and Devries2003, Eastman Reference Eastman2020) consumed more silverfish than did smaller toothfish, but the difference was modest, and silverfish remained a relatively minor prey item even for the largest toothfish in this study (24.5–54.0 kg, 125–160 cm, > 13 years old: 7.1% by weight silverfish). Overall, our study shows no evidence of significant changes in toothfish diet with individual size, and the diet information is consistent with predominant (but not exclusive) feeding near the seabed, as in previous toothfish feeding studies.
Furthermore, variations in catch rates of toothfish by depth provide no evidence that toothfish of any size forage in the water column, even in the inner McMurdo Sound below sea ice. The data used by Eastman’s (1985a,b) study were collected as described by Raymond (Reference Raymond1975): ‘initially the hooks were spaced at even intervals from the surface to the bottom which was at a depth of 500 m, but we soon learned that almost all the catches were made in the bottom 100 metres. Subsequently all the hooks were placed in this interval.’ Similarly, Parker et al. (Reference Parker, Mormede, Hanchet, Devries, Canese and Ghigliotti2019) deployed 28 hooks spanning the bottom 40 m of the water column, and interactions were higher within 5 m of the sea floor. Further work with baited vertical longlines deployed in the inner McMurdo Sound and Terra Nova Bay (Devine et al. Reference Devine, Parker and Prasad2021, Devine & Prasad Reference Devine and Prasad2022) also found that toothfish were not caught in the water column. Therefore, we conclude that differences in how toothfish were caught are not able to explain the differences in toothfish diet between this and previous studies.
The difference in silverfish in the diet of toothfish from ice-based samples in the inner McMurdo Sound in the spring months (November–December) compared with open water-based samples, including outer McMurdo Sound, in January–February is striking. One explanation is that silverfish mainly inhabit coastal fast-ice areas (i.e. inner McMurdo Sound or Terra Nova Bay; O’Driscoll et al. Reference O’Driscoll, Ladroit, Parker, Vacchi, Canese and Ghigliotti2018, La Mesa et al. Reference La Mesa, Piepenburg, Pineda-Metz, Riginella and Eastman2019), where they are available to toothfish. This may be an especially important prey item seasonally, as silverfish school to spawn in the platelet ice zone during early spring months (Vacchi et al. Reference Vacchi, La Mesa, Massimo and MacDonald2004, O’Driscoll et al. Reference O’Driscoll, Ladroit, Parker, Vacchi, Canese and Ghigliotti2018). Alternatively, the silverfish may be locally abundant under seasonal sea ice in the winter in outer McMurdo Sound but then leave the area, including the open water shelf areas, during summer months. Currently, no information on silverfish abundance is available from outer McMurdo Sound when it is ice free in the summer, and no information on silverfish abundance is available from under sea ice on the Ross Sea shelf area during early spring.
Scavenging by toothfish
The importance of scavenging is generally difficult to determine from stomach contents. On the Ross Sea continental slope and offshore oceanic features, scavenging could be an important component of toothfish diet, with bait, depredated fish bycatch, large squid remains and occasional fishing discards and seabird and seal remains reported (Fenaughty et al. Reference Fenaughty, Stevens and Hanchet2003, Kokorin Reference Kokorin2010, Stevens et al. Reference Stevens, Dunn, Pinkerton and Forman2014, Shepherd et al. Reference Shepherd, Miskelly, Cherel and Tennyson2021, Devine & Peron Reference Devine and Péron2023). On the continental shelf, bait (D. gigas) was found in 6.8–16.4% of stomachs over the four surveys, while depredated fish bycatch (identifiable by probable longline hook damage or partial consumption by scavenging amphipods - often present inside the prey) was observed in 0.8–7.1% of stomachs over the four surveys. Scavenged penguin remains have been recovered from toothfish stomachs on a few occasions in the Ross Sea (Fenaughty et al. Reference Fenaughty, Stevens and Hanchet2003, royal penguins Eudyptes schlegeli Finsch: Shepherd et al. Reference Shepherd, Miskelly, Cherel and Tennyson2021), including on the shelf surveys (Adélie penguins P. adeliae: Devine & Péron Reference Devine and Péron2023).
Variability in toothfish diet
This study identified some significant differences in diet between samples, but most of the variability in diet could not be explained by the predictors, and there was little evidence of systematic spatial variation in toothfish diet across the areas sampled on the Ross Sea shelf. Specifically, we found no statistical difference between the diets of toothfish from continental shelf areas covered by the survey Core area strata and outer McMurdo Sound, but the diet was different in Terra Nova Bay. In Terra Nova Bay, icefish were more important than notothens in toothfish diet, in particular D. hunteri, which was not observed in the diet of toothfish from outer McMurdo Sound and was a rare prey on the wider continental shelf (survey Core area).
Ontogenetic changes in diet are expected in fishes because of changes to habitat and gape-limited foraging (where feeding is restricted by mouth size). The smallest group of toothfish consumed relatively large proportions of Trematomus spp. and the benthic Crangonidae and relatively small proportions of Channichthyidae. Stevens et al. (Reference Stevens, Dunn, Pinkerton and Forman2014) found sub-adult toothfish (< 110 cm TL) on the continental slope consumed more of the relatively small Trematomus spp., dragonfish (mainly Bathydraco spp.) and Nematocarcinus sp. prawns than adults.
The proportion of fish prey, including the dominant notothen prey T. loennbergii, increased over the years in the survey Core strata. However, our ability to identify partially digested fish, including notothens, also improved over the surveys. Most unidentified Trematomus had black mouths, a character unique to T. loennbergii and Trematomus lepidorhinus (Pappenheim). However, most black-mouthed specimens were likely to be T. loennbergii, as this was the most common bycatch species on survey longlines and in survey pots (Devine & Péron Reference Devine and Péron2023, Devine et al. Reference Devine, Jones and Walker2024).
We note that our information on toothfish diet is restricted to only one time of year. Given the high seasonality in primary productivity and environmental conditions in the high-latitude Ross Sea shelf, seasonal variations in diet across the food web are expected. Biomarker (fatty acid and stable isotope) analyses can complement conventional stomach content dietary analysis, providing longer-timescale information on feeding relationships. However, the multiplicity of potential prey items coupled with their overlapping biomarker signatures over the Ross Sea shelf (Jo et al. Reference Jo, Lim, Hanchet, Lee and Kang2013) mean that these approaches do not, as yet, provide a viable alternative to stomach contents analysis for elucidating species-specific predator-prey connections. Seasonal variations in the importance of silverfish as a prey for toothfish would be especially interesting to study if a specific biotracer could be identified.
Final comments
This study provided new information on the diet of toothfish over the south-west Ross Sea continental shelf. In contrast to earlier studies under shore fast ice, silverfish were not important prey, and the diet mainly comprised small notothens (mainly T. loennbergii) and icefish. The feeding studies to date indicate that toothfish are primarily non-selective benthic feeders whose size and prey distributions change spatially. These changes should be incorporated into any study that examines the ecosystem effects of changes in the toothfish population.
Research associated with the Ross Sea region MPA aims to improve understanding of the structure and dynamics of the ecosystem, allowing us to anticipate and manage threats to the ecosystem, particularly those associated with the fishery for Antarctic toothfish (Dunn et al. Reference Dunn, Vacchi and Watters2017). The trophic connectivity between toothfish and silverfish is a key piece in the puzzle of how the dual stressors of climate change and fishing will affect the wider Ross Sea ecosystem. Our study advances this understanding and yet highlights the complexities of this trophic connectivity, especially variations in predator-prey relationships in space, season and potentially life-cycle stage of both species.
Acknowledgements
We thank Ben Sharp, Marino Vacchi (deceased, formerly ISMAR-CNR, Italy), David Bowden, Stuart Hanchet, Sophie Mormede, Susan Gregory (British Antarctic Survey, UK), Hyun-Su Jo (National Fisheries Research and Development Institute, Korea), Roberto Sarralde (Instituto Español de Oceanografía, Spain) and the New Zealand and international (CCAMLR) scientific observers Marli Dee, Geoff Dolan, John Schinkel (NZL), Jacques Combrinck, Peter Lafite, Keith Paterson and Schalk Visagie (CCAMLR) and Sanford Ltd science consultants Jack Fenaughty, Dave Bilton, and Monique Messina, who helped collect the stomachs used in this study. Thanks also to the crew of the San Aotea II for their excellent cooperation throughout the surveys. Peter McMillan (Museum of New Zealand Te Papa Tongarewa), Sadie Mills, Kareen Schnabel, Owen Anderson, Niki Davey (all Earth Sciences New Zealand) and Anne-Nina Lörz helped with the identification of toothfish prey items. The New Zealand Antarctic Fisheries Working Group provided useful comments on an earlier draft.
Author contributions
All authors were involved in the conception and design of this study. DWS and JSF examined the toothfish stomach contents. DWS, MRD and MHP drafted the manuscript. MRD and MHP performed the data analyses. JAD created Fig. 1. MP provided funding for the study. All authors edited, read and approved the final manuscript.
Financial support
The project was co-funded by: the New Zealand Ministry for Business, Innovation and Employment (MBIE) Endeavour Programme C01X1710 (Ross-RAMP); the New Zealand Antarctic Science Platform phase 1 (ANTA1801) and phase 2 (Tiaki Moana, ANTARCTICANZ2504); MBIE SSIF funding to Earth Sciences New Zealand (Healthy Ocean); and the New Zealand Ministry for Primary Industries project ANT201901C/RSSS.
Competing interests
The authors declare none.


 Open access
Open access