


Introduction
Hearing problems (HP), including hearing loss, auditory processing deficits, and other related impairments, are among the most common chronic health conditions across lifespan (Alanazi, Reference Alanazi2023; Volter et al., Reference Volter, Gotze, Bruene-Cohrs, Dazert and Thomas2020). Recent studies have highlighted an increased prevalence of acquired HP among individuals affected by neurodevelopmental disorders, such as autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), and Tourette syndrome (TS) (Gunturu et al., Reference Gunturu, Saeidi, Alzein, Jafari, Salehi and Jaka2025; Lowick & Mbatha, Reference Lowick and Mbatha2025; McKenna et al., Reference McKenna, Prasad, Cooper, King, Shahzeidi, Mittal and Eshraghi2024). This epidemiological evidence supports possible shared pathogenesis linking neurodevelopmental disorders to altered auditory function later in life. In this context, the loss of SCN2A gene in oligodendrocytes has been associated with the disruption of myelin plasticity and neural circuit integrity as well as subsequent alterations in axonal and synaptic function, resulting in auditory processing abnormalities among individuals with ASD (Bae et al., Reference Bae, Wu, Nip, Gould and Kim2025). Additionally, other studies reported that sensory hypersensitivity, including auditory sensitivity, is significantly associated with autistic traits, with attention switching and communication difficulties serving as key predictors (Keshavarz & Esmaeilpour, Reference Keshavarz and Esmaeilpour2025).
To date, most studies regarding the comorbidity between neurodevelopmental disorders and acquired HP primarily focused on specific biological pathways and molecular targets (McKenna et al., Reference McKenna, Prasad, Cooper, King, Shahzeidi, Mittal and Eshraghi2024; Omidvar, Jeddi, Doosti, & Hashemi, Reference Omidvar, Jeddi, Doosti and Hashemi2020; Worley, Matson, & Kozlowski, Reference Worley, Matson and Kozlowski2011). Large-scale genome-wide association studies (GWAS) permitted investigators to generate novel insights into the pathogenesis of human traits and diseases (Abdellaoui, Yengo, Verweij, & Visscher, Reference Abdellaoui, Yengo, Verweij and Visscher2023), including neurodevelopmental disorders (Demontis et al., Reference Demontis, Walters, Athanasiadis, Walters, Therrien, Nielsen and Borglum2023; Grove et al., Reference Grove, Ripke, Als, Mattheisen, Walters, Won and Borglum2019; Yu et al., Reference Yu, Sul, Tsetsos, Nawaz, Huang and Zelaya2019) and acquired HP (De Angelis et al., Reference De Angelis, Zeleznik, Wendt, Pathak, Tylee, De Lillo and Polimanti2023). Integrating genome-wide information with multi-modal magnetic resonance imaging data, a brain-wide investigation demonstrated widespread pleiotropy between structural and functional changes in the nervous system and acquired HP in adults (He et al., Reference He, Cabrera-Mendoza, De Angelis, Pathak, Koller, Curhan and Polimanti2024). In this context, a systematic investigation of the shared genetic mechanisms linking neurodevelopmental disorders with hearing problems can provide deeper insight into the pathogenesis of these conditions and may reveal opportunities for therapeutic interventions that could improve the quality of life of the affected individuals. To our knowledge, although genome-wide data are available for these disorders (De Angelis et al., Reference De Angelis, Zeleznik, Wendt, Pathak, Tylee, De Lillo and Polimanti2023; Demontis et al., Reference Demontis, Walters, Athanasiadis, Walters, Therrien, Nielsen and Borglum2023; Grove et al., Reference Grove, Ripke, Als, Mattheisen, Walters, Won and Borglum2019; Yu et al., Reference Yu, Sul, Tsetsos, Nawaz, Huang and Zelaya2019), no study has fully leveraged them to dissect their genetic overlap. To fill this gap, we applied a multi-layered analytic framework to disentangle HP pleiotropy with ASD, ADHD, and TS. Our analyses investigated both global and local pleiotropy using complementary approaches. Then, we translated our genetic findings into their potential functional implications through gene ontology (GO) and drug-repurposing analyses. Overall, our study demonstrated that the comorbidity between HPs and neurodevelopmental disorders is at least partially due to widespread pleiotropy between these conditions. These findings could be translated into instruments for the early identification of individuals at high comorbidity risk and targets to develop tailored therapeutic strategies.
Materials and methods
We conducted a genome-wide investigation to assess global and local pleiotropy between HP and multiple neurodevelopmental disorders (ASD, ADHD, and TS). Owing to the use of previously collected, deidentified, and aggregated data, this study did not require institutional review board approval. Ethical approval and written informed consent had been obtained in all original cohorts that enrolled the participants investigated (De Angelis et al., Reference De Angelis, Zeleznik, Wendt, Pathak, Tylee, De Lillo and Polimanti2023; Demontis et al., Reference Demontis, Walters, Athanasiadis, Walters, Therrien, Nielsen and Borglum2023; Grove et al., Reference Grove, Ripke, Als, Mattheisen, Walters, Won and Borglum2019; Yu et al., Reference Yu, Sul, Tsetsos, Nawaz, Huang and Zelaya2019).
Genome-wide datasets
We leveraged genome-wide association statistics generated from large-scale studies (Supplemental Table 1). Specifically, GWAS of neurodevelopmental disorders were obtained from the Psychiatric Genomics Consortium (available at https://www.med.unc.edu/pgc/download-results/). ASD genome-wide information was derived from a meta-analysis including 18,381 ASD cases and 27,969 controls from clinical cohorts and population-based samples (Grove et al., Reference Grove, Ripke, Als, Mattheisen, Walters, Won and Borglum2019). ADHD GWAS meta-analysis included 38,691 cases and 186,843 controls from cohorts enrolling participants (Demontis et al., Reference Demontis, Walters, Athanasiadis, Walters, Therrien, Nielsen and Borglum2023). TS GWAS was based on 4,819 cases and 9,488 controls (Yu et al., Reference Yu, Sul, Tsetsos, Nawaz, Huang and Zelaya2019). PGC GWAS of these psychiatric disorders included individuals with different ages at diagnosis. With respect to HP, we leveraged genome-wide association statistics derived from 501,825 adult participants (29% cases) (De Angelis et al., Reference De Angelis, Zeleznik, Wendt, Pathak, Tylee, De Lillo and Polimanti2023) (available at https://zenodo.org/records/7897038). To focus only on acquired HPs, this study excluded cases of congenital HP forms. Due to the limited availability of cohorts representing global populations, the datasets investigated included only individuals of European descent (EUR).
Global genetic correlation and polygenic overlap
We applied linkage disequilibrium score regression (LDSC, version 1.0.1; available at https://github.com/bulik/ldsc) to estimate SNP-based heritability and global genetic correlations among HP, ASD, ADHD, and TS. LDSC analysis was conducted using HapMap variants and pre-computed LD scores from 1000 Genomes Project EUR populations (1000 Genomes Project Consortium et al., Reference Auton, Brooks, Durbin, Garrison, Kang and Abecasis2015). To further investigate HP polygenic overlap with neurodevelopmental disorders, we used the Mixture of eXperts Regression (MiXeR, version 1.2.0) (Frei et al., Reference Frei, Holland, Smeland, Shadrin, Fan, Maeland and Dale2019). All analyses were performed using the MiXeR Python wrapper (available at https://github.com/precimed/mixer), with parameter optimization performed via grid search with log-likelihood maximization. Similar to LDSC analysis, this analysis was also based on HapMap variants and LD structure of 1000 Genomes Project EUR populations (1000 Genomes Project Consortium et al., Reference Auton, Brooks, Durbin, Garrison, Kang and Abecasis2015). The bivariate MiXeR analysis was performed to estimate the number of shared causal SNPs and the degree of genetic overlap between traits, beyond what is captured by genetic correlation. Specifically, we estimated the proportion of shared polygenic signal and the concordance ratios indicating the proportion of shared SNPs with concordant effect directions. To define statistically meaningful bivariate MixeR models, we considered only those with Akaike information criterion (AIC) for the best model versus the model with minimal polygenic overlap greater than zero.
Local genetic correlation
We applied Local Analysis of Variant Association (LAVA) approach (version 1.8.0) (Werme, van der Sluis, Posthuma, & de Leeuw, Reference Werme, van der Sluis, Posthuma and de Leeuw2022) to investigate locus-specific pleiotropy between HP and neurodevelopmental disorders. Following LAVA documentation (available at https://github.com/josefin-werme/LAVA), we used 2,495 pre-defined, approximately independent LD blocks from 1000 Genomes Project EUR reference populations (1000 Genomes Project Consortium et al., Reference Auton, Brooks, Durbin, Garrison, Kang and Abecasis2015). For each trait, we estimate the locus-specific SNP-based heritability. Then, we performed LAVA bivariate analysis to estimate locus-specific genetic correlation between HP and neurodevelopmental disorders. False discovery rate (FDR q < 0.05) was applied to define statistically significant local genetic correlations after accounting for the number of loci tested.
Pleiotropic loci
To identify individual loci with pleiotropic effects on both HP and neurodevelopmental disorders, we used PolarMorphism approach (available at https://github.com/UMCUGenetics/PolarMorphism) (von Berg, Ten Dam, van der Laan, & de Ridder, Reference von Berg, Ten Dam, van der Laan and de Ridder2022). This method analyzes genome-wide data to identify genetic variants associated with multiple traits due to horizontal pleiotropy, attenuating the effect of vertical pleiotropy observed in the genetic correlation via a decorrelating transform (von Berg et al., Reference von Berg, Ten Dam, van der Laan and de Ridder2022). For each SNP, PolarMorphism computes two parameters: the overall strength of association across traits (r) and the relative contribution of the effect across the traits (theta). Theta q value < 0.05 was used to define significant pleiotropic loci after FDR multiple testing correction. PLINK 1.9 (available at https://www.cog-genomics.org/plink/1.9/) was used to identify LD-independent pleiotropic loci, considering a 250-kb window and an LD r 2 < 0.1 based on 1000 Genomes Project Phase 3 EUR reference populations (1000 Genomes Project Consortium et al., Reference Auton, Brooks, Durbin, Garrison, Kang and Abecasis2015). Functional annotation was performed using Open Targets Platform (Buniello et al., Reference Buniello, Suveges, Cruz-Castillo, Llinares, Cornu, Lopez and Ochoa2025) (available at https://platform.opentargets.org/) and GTEx v10 (GTEx Consortium, 2020) (available at https://www.gtexportal.org/home/).
Gene ontology and drug repurposing analyses
To explore the biological processes, molecular functions, and cellular components shared between HP and neurodevelopmental disorders, we applied GSA-MiXeR (available at https://github.com/precimed/gsa-mixer) to estimate gene-set heritability enrichments across these traits (Frei et al., Reference Frei, Hindley, Shadrin, van der Meer, Akdeniz, Hagen and Dale2024). Specifically, disorder-specific genome-wide association statistics were used as input for the GSA-MiXeR analysis. Then, to define GO terms shared between HP and neurodevelopmental disorders, we applied a Bonferroni multiple testing correction accounting for the number of GOs tested (p < 6.31 × 10−7), considering both pair-wise combinations (GO terms that were Bonferroni significant with respect to both HP and one of the neurodevelopmental disorders tested) and a four-way model (GO terms that were Bonferroni significant with respect to HP and all three neurodevelopmental disorders tested). To facilitate their interpretation, we pruned Bonferroni-significant GO terms similarly to GSA-MiXer original study (Frei et al., Reference Frei, Hindley, Shadrin, van der Meer, Akdeniz, Hagen and Dale2024). Specifically, we applied REVIGO (Supek, Bosnjak, Skunca, & Smuc, Reference Supek, Bosnjak, Skunca and Smuc2011), considering a similarity threshold = 0.7 and UniProt as the reference database. Non-redundant Bonferroni filtered GO terms were then used as input to perform a drug-repurposing analysis. Specifically, we used the Gene2drug approach (Napolitano et al., Reference Napolitano, Carrella, Mandriani, Pisonero-Vaquero, Sirci, Medina and di Bernardo2018) to identify molecular compounds associated with transcriptomic profiles overlapping with those of the shared GOs between HP and neurodevelopmental disorders. Transcriptomic reference data were derived from the Connectivity Map drug database (1.5 M gene expression profiles from ~5,000 small-molecule compounds, and ~ 3,000 genetic reagents) (Lamb et al., Reference Lamb, Crawford, Peck, Modell, Blat, Wrobel and Golub2006).
Results
Genetic correlation and polygenic overlap
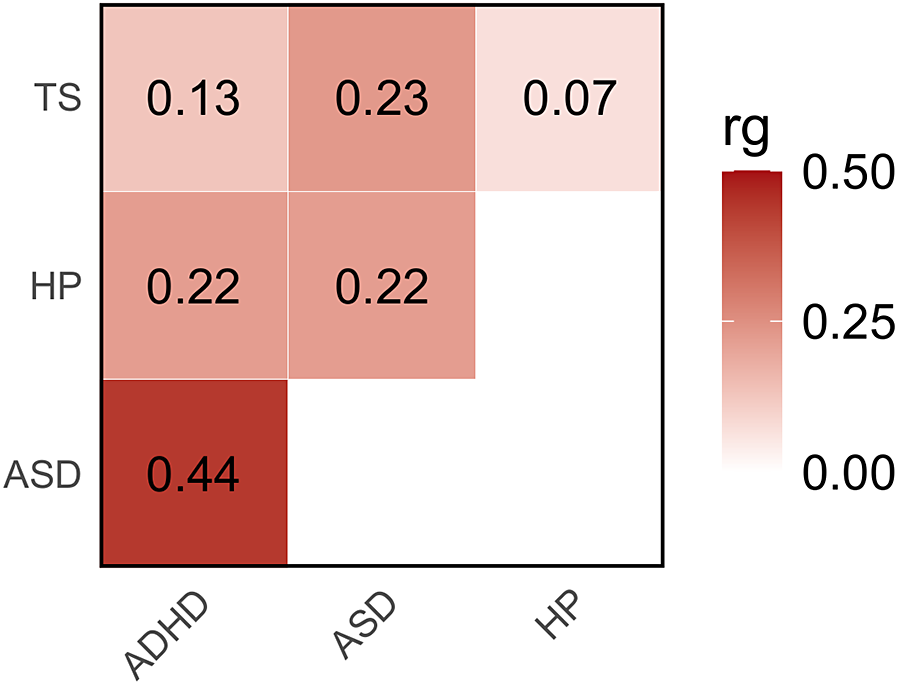
The observed-scale SNP-based heritability (SNP-h2) across the traits investigated ranged from 42% in ASD to 5% in ADHD (Supplemental Table 1). No evidence of inflation due to confounders was observed in the GWAS investigated (LDSC intercept < 1.03). Considering SNP-based heritability z-scores as a proxy of the statistical power of the GWAS investigated, we observed comparable statistical power for HP and ADHD (SNP-h2 z = 18.1 and 21.6, respectively), while reduced statistical power was present in TS and ASD GWAS (SNP-h2 z = 8.6 and 8.4, respectively). These differences were mostly explained by the effective sample size of the GWAS investigated (see section Genome-wide Datasets in METHODS). Nevertheless, HP genetic correlations were statistically significant for all neurodevelopmental disorders, although they differed in magnitude (Figure 1, Supplemental Table 2). Specifically, HP showed genetic correlations with both ASD (rg = 0.22 ± 0.04) and ADHD (rg = 0.22 ± 0.03), while the HP-TS genetic correlation was not statistically significant (p>0.05). The extent of HP genetic correlations reflected the pleiotropy among the neurodevelopment disorders investigated (Figure 1, Supplemental Table 2).
Genetic correlations (rg) among hearing problems (HP), autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), and Tourette syndrome (TS). Color intensity reflects rg magnitude (Supplemental Table 2).

We observed good fitting statistics only for the HP-ADHD bivariate MiXeR model (AICbestVSmininal = 6.13; Figure 2, Supplemental Table 3), with 34 ± 6% of the causal variants related to these traits were shared. Within these pleiotropic loci, only 74 ± 5% showed concordant effect directions. HP bivariate MiXeR models with ASD and TS were not statistically meaningful (AICbestVSmininal < 0) because the GWAS of these neurodevelopmental disorders had limited statistical power due to their smaller effective sample size.
Polygenic overlap between hearing problems (HP) and attention-deficit/hyperactivity disorder (ADHD). (a) Venn diagrams of unique and shared polygenic components at the causal level, showing polygenic overlap (gray) between HP (blue) and ADHD (orange). The numbers indicate the estimated quantity of causal variants (in thousands) per component, accounting for 90% of SNP heritability in each phenotype, along with the standard error between parentheses. The size of the circles reflects the degree of polygenicity. (b, c) Conditional Q–Q plots of observed versus expected −log10 p-values in the primary trait as a function of significance of association with a secondary trait at the level of p ≤ 0.1 (orange lines), p ≤ 0.01 (green lines), p ≤ 0.001 (red lines). Blue line indicates all SNPs. Dotted lines in blue, orange, green, and red indicate model predictions for each stratum. The black dotted line represents the expected Q–Q plot under the null hypothesis. (d) Log-likelihood profile for the estimated number of shared causal variants (k). The solid line represents the smoothed likelihood curve, while dotted lines indicate chromosome-wise estimates from the MiXeR fit. The minimum denotes the maximum likelihood estimate of k, supporting the presence of a finite set of shared causal variants.

Local genetic correlation
We identified 10 FDR-significant locus-specific genetic correlations, with nine of them related to HP-ADHD and one to HP-ASD (Table 1). There was no overlap between the FDR-significant local genetic correlations related to HP-ADHD and HP-ASD. Only the HP-ADHD local genetic correlation on chromosome 12 (hg19 position: 116,197,676–117,091,843; ρ = 0.76, p = 1.16 × 10−5, FDR q = 0.008) showed a nominal HP-ASD replication (p = 0.017). Within this chromosomal region, a total of 127 genome-wide associations were reported in the GWAS Catalog (Sollis et al., Reference Sollis, Mosaku, Abid, Buniello, Cerezo, Gil and Harris2023) (Supplemental Table 4), including variants associated with skeletal phenotypes (e.g. bone mineral density), immune-related traits (e.g. acute myeloid leukemia and monocyte count), and cardiovascular outcomes (e.g. blood pressure and atrial fibrillation). With respect to HP, only genome-wide significant gene–gene interaction was reported (rs4301303-rs10444487 p = 7 × 10−9). While no association was reported with respect to ADHD, ASD, or other psychiatric disorders, there was evidence related to behavioral traits (e.g. educational attainment, rs76130193 p = 8 × 10−18; smoking initiation, rs71469748 p = 4 × 10−11) and brain-imaging phenotypes (e.g. vertex-wise sulcal depth rs74550975 p = 9 × 10−10; cortical surface area rs74550975 p = 4 × 10−11). Interactive effects between smoking status and diastolic pressure were also identified within this chromosomal region (rs11067762 p = 5 × 10−18).
Local genetic correlations between HP and neurodevelopmental disorders (autism spectrum disorder, ASD; attention-deficit hyperactivity disorder) surviving false discovery rate multiple testing correction (FDR q < 0.05)

Abbreviations: BP, base pairs; NS, not significant.
Among the other loci specific to HP-ADHD pleiotropy, the most significant result was on chromosome 6 (hg19 position: 149,278,799–150,635,315; ρ = 1, p = 1.15 × 10−6, FDR q = 0.002). More than 240 genome-wide significant associations were reported within this chromosomal region in the GWAS catalog (Supplemental Table 5). The most significant included retinal thickness (rs57215159 p = 5 × 10−72), atrial fibrillation (rs117984853 p = 4 × 10−55), and several brain-imaging phenotypes (e.g. brain morphology rs7752089 p = 2 × 10−31; subcortical volume rs12523793 p = 2 × 10−24; vertex-wise sulcal depth rs14314 p = 7 × 10−17). Some HP-ADHD pleiotropic regions (i.e. hg19 chr2:9,960,671–10,611,654; chr4:79,880,102–81,206,182; chr9:37,687,113–38,822,977; chr17:27,344,402–29,783,141; chr20:58,780,895–59,560,457) showed genome-wide significant associations that converged on phenotypic traits, such as bone mineral density, blood pressure, atrial fibrillation, hematologic biomarkers, smoking initiation, and educational attainment (Supplemental Tables 6–10).
HP-ADHD pleiotropic regions showed different association patterns. For example, within hg19 chr2:234,115,093–234,945,577 region (Supplemental Table 11), we identified the locus most strongly associated with bilirubin levels (e.g. rs6704644 p = 1 × 10−1043). Within hg19 chr4:102,544,804–104,384,534 region, multiple genome-wide significant associations were reported for brain iron levels (e.g. pallidum iron levels rs13107325 p = 2 × 10−231; substantia nigra iron levels rs13107325 p = 2 × 10−197; caudate iron levels rs13107325 p = 2 × 10−167) along with several brain-imaging phenotypes (Supplemental Table 12). The only HP-ASD pleiotropic locus identified (hg19 chr19:51,259,179–51,903,804) showed an inverse local genetic correlation (ρ = −0.71, p = 9.81 × 10−5, FDR q = 0.028) and included many genome-wide significant associations with immune-related phenotypes (Supplemental Table 13), with those related to CD33 levels among the strongest ones (e.g. myeloid cell surface antigen CD33 levels rs33978622 p = 1 × 10−223).
Pleiotropy analysis
Our genome-wide pleiotropy analysis using PolarMorphism identified specific variants showing horizontal pleiotropy (shared pathogenic effects) between HP and neurodevelopmental disorders. Because PolarMorphism relies on the single-variant discovery power of the GWAS input GWAS (von Berg et al., Reference von Berg, Ten Dam, van der Laan and de Ridder2022), TS was excluded from this analysis due to the fact that its GWAS identified only one genome-wide significant locus (Yu et al., Reference Yu, Sul, Tsetsos, Nawaz, Huang and Zelaya2019). Modeling pleiotropic effects among HP, ASD, and ADHD, we identified two LD-independent variants with FDR-significant r and θ statistics, which correspond to the overall strength of association across traits and the relative contribution of the effect across the traits, respectively (Supplemental Table 14). On chromosome 5, rs325485*A allele showed genome-wide significant association with both HP (β = 0.026, p = 2.57 × 10−8) and ADHD (β = 0.067, p = 2.74 × 10−12) and consistent effect direction on ASD (β = 0.081, p = 0.005). According to Open Target Platform (Buniello et al., Reference Buniello, Suveges, Cruz-Castillo, Llinares, Cornu, Lopez and Ochoa2025), this variant is included within 57 95%-credible sets related to a range of complex traits (Supplemental Table 15). The top associations included hand grip strength, ASD, depression medications, risk-taking tendency, and asthma-depression comorbidity. Additionally, rs325485 is both an expression quantitative trait locus (QTL) and a splicing QTL for ENSG00000251574 in testis (Supplemental Table 16). The other pleiotropic locus was rs2207286 on chromosome 1, which showed concordant effects across HP (β = 0.019, p = 1.7 × 10−4), ASD (β = 0.096, p = 0.001), and ADHD (β = 0.036, p = 2.86 × 10−4). This variant is included within 11 95%-credible sets available from the Open Targets Platform (Buniello et al., Reference Buniello, Suveges, Cruz-Castillo, Llinares, Cornu, Lopez and Ochoa2025), with the most significant ones related to insomnia, middle ear disease, acquired hearing loss, general risk tolerance, and non-suppurative otitis media (Supplemental Table 17). No additional loci were identified when testing ADHD-HP and ASD-HP pairwise pleiotropy.
Gene enrichment and drug-repurposing analyses
Through GSA-MiXeR approach (Frei et al., Reference Frei, Hindley, Shadrin, van der Meer, Akdeniz, Hagen and Dale2024) and subsequent REVIGO pruning (Supek et al., Reference Supek, Bosnjak, Skunca and Smuc2011), we identified 56 non-redundant GO terms that reached Bonferroni significance with respect to both HP and all neurodevelopmental disorders investigated (p < 6.31 × 10−7; Figure 3, Supplemental Table 18). For instance, ‘plasma membrane region’ (GO:0098590) was the most significant GO term for both ASD (p = 9.87 × 10−52) and TS (p = 3.94 × 10−47), but it was also significant for ADHD (p = 1.47 × 10−24) and HP (p = 1.23 × 10−7). Similarly, ‘supramolecular complex’ (GO: GO:0099080) was the most significant for both HP (p = 1.51 × 10−64) and ADHD (p = 3.25 × 10−37), and it was also significant for ASD (p = 7.25 × 10−26) and TS (p = 1.48 × 10−27). For the subsequent drug-repurposing analysis, we considered non-redundant Bonferroni-significant GO terms shared across all four traits investigated and those shared between HP and each neurodevelopmental disorder tested. With respect to GO terms shared between HP and ASD, we identified a significant enrichment for the transcriptomic profile related to anisomycin (ES = −0.54, p = 1.93 × 10−5). Conversely, HP-ADHD shared GO terms were related to five molecular compounds: perphenazine (ES = 0.69, p = 3.11 × 10−5), bufexamac (ES = 0.74, p = 5.41 × 10−5), S-propranolol (ES = 0.67, p = 6.87 × 10−5), trichostatin A (ES = 0.73, p = 6.98 × 10−5), and propylthiouracil (ES = 0.73, p = 8.24 × 10−5).
Heatmap of GO Terms reaching Bonferroni significance (p < 6.31 × 10−7) shared among hearing problems (HP), autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), and Tourette syndrome (TS). The color scale represents the −log10 (p-value) of each GO term, with deeper red indicating stronger statistical significance.

Discussion
While extensive evidence exists regarding the comorbidity and the underlying pathogenic processes linking neurodevelopmental disorders and congenital hearing impairments (Akrich et al., Reference Akrich, Parlato de Oliveira, Favrot-Meunier, Rebichon, de Clerck, Ettori and Loundon2023), little is known regarding the dynamics contributing to the relationship observed between neurodevelopmental disorders and acquired HP (Danyluk & Jacob, Reference Danyluk and Jacob2023). Leveraging large-scale genome-wide datasets, the present study uncovered global and local pleiotropy between three neurodevelopmental disorders (ASD, ADHD, and TS) and HP, generating new insights into the underlying pathogenetic processes and potential molecular targets involved in this comorbidity. Our global genetic correlation analysis highlighted different degrees of pleiotropy between HP and the neurodevelopmental disorders investigated. Specifically, ASD and ADHD showed much larger genetic correlations with HP (rg > 0.20) than TS (rg < 0.1). HP genetic correlation with ASD and ADHD suggests their pleiotropy may be related to a shared neurodevelopmental factor identified previously (Grotzinger et al., Reference Grotzinger, Werme, Peyrot, Frei, de Leeuw, Bicks and Smoller2025). Clinical and epidemiological studies have reported that individuals affected by ASD are more likely to be affected by hearing impairment over their lifetime than the general population and age-matched population groups (Demopoulos & Lewine, Reference Demopoulos and Lewine2016; Nian et al., Reference Nian, Cheng, Cheng, Lin, Wang, Chien and Lin2024). Conversely, no information is available regarding the relationship between TS and HP.
With respect to HP-ADHD genetic correlation, a nationwide, population-based, cross-sectional study of adolescents reported a strong association between hearing impairment and severe ADHD with consistent relationships across hearing impairment type (sensorineural versus conductive) and severity (Tsur et al., Reference Tsur, Zloof, Rittblat, Reuven, Simchoni, Derazne and Beer2024). These previous findings are in line with HP-ADHD polygenic overlap identified by our MiXeR analysis. Indeed, 34% of causal variants were shared between these conditions, with >70% of them showing concordant effect directions. Interestingly, our analysis identified 10 chromosomal regions with evidence of local genetic correlation that was consistent with HP-ADHD global genetic correlation (i.e. increased ADHD genetic risk was correlated with increased HP genetic risk). This suggest that >70% of shared causal variants with concordant HP-ADHD effects have larger effect sizes than those with discordant effects. Several chromosomal regions identified have been previously associated with traits that may play a role in HP-ADHD co-occurrence. For instance, some of the loci identified were linked to brain structural changes, which may connect previous findings on neuroimaging differences in ADHD cases (Lohani & Rana, Reference Lohani and Rana2023; Pereira-Sanchez & Castellanos, Reference Pereira-Sanchez and Castellanos2021) and recent findings connecting central nervous system alterations to HP (He et al., Reference He, Cabrera-Mendoza, De Angelis, Pathak, Koller, Curhan and Polimanti2024). Additionally, one of the HP-ADHD loci was previously associated with iron levels in multiple brain regions. A systematic review of neuroimaging studies reported significantly reduced brain iron content in medication-naïve children with ADHD (Morandini, Watson, Barbaro, & Rao, Reference Morandini, Watson, Barbaro and Rao2024). Similarly, iron deficiency anemia was associated with both sensorineural and combined hearing loss in adults (Schieffer et al., Reference Schieffer, Chuang, Connor, Pawelczyk and Sekhar2017). These findings suggest that altered iron regulation may play a role in ADHD-HP shared pathogenesis.
Two other HP-ADHD loci supported additional pathogenic mechanisms. On chr 2 (hg19 position: 234,115,093–234,945,577), HP-ADHD local genetic correlation overlapped with the most significant locus associated with bilirubin levels (Sinnott-Armstrong et al., Reference Sinnott-Armstrong, Tanigawa, Amar, Mars, Benner, Aguirre and Rivas2021). Neonatal jaundice (i.e. elevated blood level of bilirubin in newborns) has been associated with ADHD and other neurodevelopmental disorders, although the pathogenic mechanisms remain unclear (Wei et al., Reference Wei, Chang, Lin, Chang, Li and Kao2015). In the context of HP, bilirubin-induced neurologic damage has been linked to auditory neuropathy spectrum disorder (Olds & Oghalai, Reference Olds and Oghalai2015). On chromosome 6 (hg19 position: 149,278,799–150,635,315), HP-ADHD local genetic correlation overlapped with a genome-wide significant association with retinal thickness. A recent study reported that ADHD patients have a significant reduction in total and outer retinal thickness across several macular sectors in both eyes, highlighting retinal thickness as a potential biomarker for this condition (Miquel-Lopez et al., Reference Miquel-Lopez, Garcia-Medina, Lopez-Hernandez, Garcia-Ayuso, De-Paco-Matallana, Hernandez-Olivares and Del-Rio-Vellosillo2025). With respect to HP, retinal thickness and related measurements have been investigated as predictors of aging-related functional decline (Lee et al., Reference Lee, Kim, Lee, Park and Bae2025).
Multiple ADHD-HP loci were related to smoking initiation. Extensive literature has reported both earlier smoking initiation among ADHD cases (van Amsterdam, van der Velde, Schulte, & van den Brink, Reference van Amsterdam, van der Velde, Schulte and van den Brink2018) and the impact of tobacco smoking on HP later in life (Garcia Morales et al., Reference Garcia Morales, Ting, Gross, Betz, Jiang, Du and Deal2022). This could point out smoking initiation as a moderator linking ADHD to HP. Similarly, other phenotypes associated across HP-ADHD loci (i.e. educational attainment, cardiovascular conditions, and immune alterations) that could connect ADHD to HP (Baiduc et al., Reference Baiduc, Sun, Berry, Anderson and Vance2023; Stonkute & Vierboom, Reference Stonkute and Vierboom2025; Tsur et al., Reference Tsur, Zloof, Rittblat, Reuven, Simchoni, Derazne and Beer2024; Zhang et al., Reference Zhang, Li, Andell, Garcia-Argibay, Quinn, D’Onofrio and Chang2024).
Beyond HP-ADHD pleiotropic loci, we also identified a genomic region on chromosome 19 (hg19: 51,259,179–51,903,804). Interestingly, the local genetic correlation in this chromosome region was inverse (inverse, increased ASD genetic risk was correlated with reduced HP genetic risk), while HP-ASD global genetic correlation was positive (increased ASD genetic risk was correlated with increased HP genetic risk). Within the genomic region on chromosome 19, there are several variants associated with immune-related traits, mainly related to CD33 gene (Supplemental Table 13), which encodes a transmembrane protein receptor expressed on myeloid cells. Genetically regulated CD33 levels in myeloid cells have been associated with increased odds of ADHD (Jue et al., Reference Jue, Dan-Fei, Fang-Fang, Ke-Pin, Jia-Ye, Hui-Ting and Jian2024). No information is currently available regarding whether CD33 may be implicated in HP pathogenesis. However, there is an extensive literature regarding CD33 role in Alzheimer’s disease and the different impact of its isoforms on neurodegeneration (Eskandari-Sedighi, Jung, & Macauley, Reference Eskandari-Sedighi, Jung and Macauley2023). We hypothesize that the inverse local genetic correlation between HP and ASD observed on chromosome 19 may be related to regulatory mechanisms that differentially influence these conditions. Supporting this, many CD33 expression QTLs have been identified by GTEx v10 (GTEx Consortium, 2020) (available at https://www.gtexportal.org/home/). While most of them showed cross-tissue effects, there were also CD33 expression QTLs in only a single tissue (Supplemental Table 19). Caudate nucleus showed the highest number of tissue-specific CD33 expression QTLs (Supplemental Table 19). Interestingly, this brain region has been linked to both ASD and HP, with increased caudate volume being associated with restricted and repetitive behaviors in ASD cases (Qiu et al., Reference Qiu, Chang, Li, Qian, Xiao, Xiao and Ke2016) and with sensory gating function of caudate nucleus linked to altered hearing perception (Wang et al., Reference Wang, Song, Liu, Du, Xiong, Fan and Ma2021).
While our local genetic correlation analysis did not identify genomic regions linking HP to both ADHD and ASD, we identified two single variants with shared genetic effects between HP and these neurodevelopmental disorders. According to Open Target Platform (Buniello et al., Reference Buniello, Suveges, Cruz-Castillo, Llinares, Cornu, Lopez and Ochoa2025), these loci are within 95% credible sets for multiple complex traits. With respect to rs325485 on chromosome 5, top associations were related to several mental health phenotypes, including ASD, depression medications, and risk-taking tendency, which support pleiotropy with neurodevelopmental disorders. This variant was also related to asthma-depression comorbidity (Supplemental Table 15), highlighting its potential implication in comorbidities between mental and physical health. In particular, its relationship with asthma may have a direct effect on HP risk, as asthma is associated with an increased risk of sensorineural hearing loss (Choi, Min, Lee, & Kim, Reference Choi, Min, Lee and Kim2022). With respect to rs2207286 on chromosome 1, the top 95% credible sets were related to several HL-related phenotypes in addition to general risk tolerance (a trait well-known to be associated with ADHD (Pollak, Shoham, Scheres, & Dekkers, Reference Pollak, Shoham, Scheres, Dekkers and Matson2023) and insomnia. The latter is commonly observed in neurodevelopmental disorders (Shelton & Malow, Reference Shelton and Malow2021) and HP (Clarke, Hoare, & Killan, Reference Clarke, Hoare and Killan2019), suggesting its potential role in contributing to the pleiotropy observed.
Through the GO analysis, we found a significant enrichment for multiple biological processes, molecular functions, and cellular components in the genetic heritability shared across HP and the three neurodevelopmental disorders investigated. These were mostly related to gene-sets related to broad domains, suggesting that the pathogenesis shared between these conditions converges on mechanisms acting across different biological systems. Nevertheless, when considering gene-sets shared between HP and each of the neurodevelopmental disorders tested, we identified specific molecular compounds showing transcriptomic signatures overlapping with the GOs identified. In some cases, it appears that there is evidence supporting the therapeutic potential for the molecular compounds identified in the context of the comorbidity between conditions investigated. For instance, ADHD-HP drug-repurposing analysis identified perphenazine, which is a first-generation antipsychotic medication that acts by blocking dopamine receptors in the brain (DrugBank Accession Number DB00850). This drug is also indicated for the treatment of Meniere’s disease, an inner ear disorder characterized by symptoms, such as vertigo, tinnitus, and hearing loss (Gupta & Jamwal, Reference Gupta and Jamwal2022). Trichostatin – also identified by the ADHD-HP drug-repurposing analysis – is an inhibitor of histone deacetylase (HDAC) enzymes (DrugBank Accession Number DB04297), which has been reported to reduce hearing loss (Nam et al., Reference Nam, Gi, Ko, Lee and Cho2025) and maladaptation-induced emotional abnormality in animal models (Kimijima et al., Reference Kimijima, Miyagawa, Kurokawa, Mochida-Saito, Takahashi, Takeda and Tsuji2022). Again with respect to ADHD-HP pleiotropy, we identified propranolol, which is a β-blocker medication used to treat cardiovascular and anxiety-related conditions (DrugBank Accession Number: DB00571). There was some earlier evidence regarding the possible benefits of propanol to treat ADHD symptoms (Mattes, Reference Mattes1986).
Some of the molecular compounds identified appears to be related to the disorders investigated because of their potential side effects. For instance, HP-ASD drug-repurposing analysis pointed to anisomycin, an antibiotic that inhibits bacterial protein and DNA synthesis (DrugBank Accession Number: DB07374). This appears to converge with the identification of HP-ASD local genetic correlation observed in CD33 locus, which plays an important role in innate immunity (Malpass, Reference Malpass2013). With respect to HP and ASD, anisomycin has been associated with impaired social recognition when administered in certain brain regions (Pena, Pereira-Caixeta, Moraes, & Pereira, Reference Pena, Pereira-Caixeta, Moraes and Pereira2014), and its chronic exposure also appears to induce hair cell death (Yuan, Qin, Wang, & Fan, Reference Yuan, Qin, Wang and Fan2021). Similarly, propylthiouracil – identified by the ADHD-HP drug-repurposing analysis – is an antithyroid medication (DrugBank Accession Number: DB00550) that has been associated with rare cases of sudden hearing loss (Tanabe et al., Reference Tanabe, Nishimura, Sugahara, Yamashita and Tanizawa2021) and is being used to induce hyperactive behaviors related to perinatal hypothyroidism in mice (Umezu, Kita, & Morita, Reference Umezu, Kita and Morita2019). Our drug-repurposing analysis also identified molecular compounds with unclear relationships with the disorders investigated. For example, the transcriptomic profile of HDAC inhibitors has been linked to the comorbidity among psychiatric disorders, spine degenerative disease, and chronic pain (Qiu et al., Reference Qiu, Friligkou, He, Cabrera-Mendoza, Aslan, Gupta and Polimanti2025). Similarly, bufexamac is a non-steroidal anti-inflammatory drug used to treat inflammatory skin conditions (DrugBank Accession Number: DB13346), but there is no evidence linking this drug to HP or ADHD. Overall, these molecular compounds highlight the presence of druggable pathways shared between HP and neurodevelopmental disorders, supporting the potential of identifying therapeutic interventions to reduce their comorbidity. Importantly, because of the pathogenesis of neurodevelopmental disorders originates in prenatal stages, some of the compounds identified in our studies may be useful to reduce the risk of developing age-related hearing problems in individuals affected by neurodevelopmental disorders.
While we believe our results open new directions in neuropsychiatric research, we also acknowledge three important limitations. First, our analysis was limited to EUR GWAS due to the limited availability of large-scale datasets for other population groups. Because of population differences in both neurodevelopmental disorders and HP (Gallin et al., Reference Gallin, Kolevzon, Reichenberg, Hankerson and Kolevzon2025; He, Curhan, Curhan, & Polimanti, Reference He, Curhan, Curhan and Polimanti2025; Shalaby, Sengupta, & Williams, Reference Shalaby, Sengupta and Williams2024), it will be important to assess the generalizability of our findings across human groups when novel datasets are available. Second, because of different GWAS statistical power, we could not investigate the three neurodevelopmental disorders with the same analytic depth, which led to an imbalance regarding the characterization of HP pleiotropy across conditions tested. Finally, longitudinal studies and causal inference analyses will be needed to explore possible mechanisms linking neurodevelopmental disorders to acquired HP later in life.
In conclusion, our study demonstrated that the pleiotropy between HP and neurodevelopmental disorders at least partially contributes to the comorbidity observed between these conditions. In particular, our findings highlighted that the relationship between neurodevelopmental disorders and altered hearing function can be due to pleiotropic mechanisms acting through both intrinsic and extrinsic pathogenic processes. Building on the evidence provided by the present investigation regarding the distinct pleiotropic mechanisms beyond global genetic correlation linking neurodevelopmental disorders to acquired HP, further studies will be needed to conduct more in-depth investigations regarding the underlying molecular mechanisms that could be responsible for the comorbidity between neurodevelopmental symptoms and altered hearing function later in life.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0033291726103201.
Data availability statement
The data generated by the present study are included in the manuscript and its supplemental material. Genome-wide association statistics from the Psychiatric Genomics Consortium can be accessed at https://pgc.unc.edu/for-researchers/download-results/. HP GWAS data can be accessed at https://zenodo.org/records/7897038.
Code availability
The analyses presented in the current study were conducted using publicly available code. LDSC, https://github.com/bulik/ldsc; MiXeR, https://github.com/precimed/mixer; LAVA, https://github.com/josefin-werme/LAVA; PolarMorphism, https://github.com/UMCUGenetics/PolarMorphism; PLINK, https://www.cog-genomics.org/plink/1.9/; GSA-MiXeR, https://github.com/precimed/gsa-mixer.
Acknowledgments
The authors thank the Psychiatric Genomics Consortium for making their data available.
Author contribution
Conceptualization: QZ, RP; Methodology: QZ, BCM, QC, JH, RP; Investigation: QZ, BCM, QC, DD, DQ, JH, RP; Visualization: QZ; Funding acquisition: QZ, RP; Supervision: RP; Writing – original draft: QZ, RP; Writing – review & editing: QZ, BCM, QC, DD, DQ, JH, RP.
Funding statement
The authors acknowledge support from the National Institutes of Health (Grant No. RF1MH132337) and the Jinan Municipal Health Commission (Grant No. 2023-BD-2-05).
Competing interests
RP received an honorarium from Karger Publishers for his work on Complex Psychiatry. The other authors declare no conflict of interest.






 Open access
Open access