


Introduction
Monocotylidae Taschenberg, 1879 is a monogenean family that infects chondrichthyans (sharks, rays, and chimaeras). The family has been object of taxonomic interest, since many species within the family are thought to remain undescribed (Chisholm et al. Reference Chisholm, Wheeler and Beverley-Burton1995). At present, the Monocotylidae comprises eight subfamilies including the Calicotylinae Monticelli, 1903. Species of Calicotylinae can be distinguished by a variety of morphological features, such as the shape of the hamuli and male copulatory organ, the direction of the vaginae, the position of vaginal pores, the arrangement of the vitellarium and intestinal caeca, and the presence or absence of eye-spots, among others (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997). Morphological differentiation between individual species is often based on minor differences and, in certain cases, this has caused disagreements about the validity of certain taxa (Chisholm, Whittington et al. Reference Chisholm, Whittington, Morgan and Adlard2001). In addition, the historically generalized assumption that monogeneans are highly host-specific may have contributed to the misidentification of monocotylids, including species of Calicotyle Diesing, 1850 (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997; Chisholm and Whittington Reference Chisholm and Whittington1996; Whittington et al. Reference Whittington, Cribb, Hamwood and Halliday2000).
Calicotylinae contains two genera: Dictyocotyle Nybelin, 1941, a monotypic genus for D. coeliaca Nybelin, 1941, and Calicotyle Diesing, 1850 (see Chisholm, Morgan et al. Reference Chisholm, Morgan, Adlard and Whittington2001), which at present includes 21 recognized taxa (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997; WoRMS 2025 and references therein). Unlike most monogeneans, which are ectoparasites, species of Calicotyle typically infect the spiral valve, cloaca, rectum, rectal gland, or oviducts of their hosts. Species of this genus are distinguished from other monocotylids by the morphological features of the haptor, having one central and seven peripheral loculi and lacking haptoral accessory structures other than two hamuli and several hooklets (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997).
Only recently, an unidentified species of Calicotyle has been reported for the first time from deepwater catsharks (Pentanchidae) (Higueruelo et al. Reference Higueruelo, Schaeffner, Soler-Membrives and Dallarés2026), a group that remains generally poorly known despite being the most species-rich of all shark families. As fisheries and research expand to deeper waters, new species and new information regarding their biology are expected to be revealed (Ebert et al. Reference Ebert, Dando and Fowler2021). Studies on parasites of deepwater catsharks from the northern Atlantic are particularly scarce, and a predominant portion of their parasite diversity (including endoparasitic helminths) is neglected.
Among deepwater catsharks, the white ghost catshark, Apristurus aphyodes Nakaya & Stehmann, 1998, the Iceland catshark, Apristurus laurussonii (Saemundsson 1922), and the mouse catshark, Galeus murinus (Collett 1904) are commonly detected in Icelandic waters (NE Atlantic) (Jakobsdóttir et al. Reference Jakobsdóttir, Hjörleifsson, Pétursson, Björnsson, Sólmundsson, Kristinsson and Bogason2023). These three species are small-bodied (less than 80 cm) bottom dwellers distributed in the northern Atlantic Ocean (with A. aphyodes and G. murinus having a more restricted NE Atlantic distribution) in depths of 380 to 2,060 m (Ebert et al. Reference Ebert, Dando and Fowler2021). They are currently listed as ‘Least Concern’ in the International Union for the Conservation of Nature’s (IUCN) Red List of Threatened Species, with a stable population trend (Finucci and Armstrong Reference Finucci and Armstrong2024; Kulka et al. Reference Kulka, Cotton, Anderson, Crysler, Herman and Dulvy2020; Rigby et al. Reference Rigby, Iglésias and Rohner2024). Their parasite fauna was essentially unknown until a recent comprehensive parasitological study identified up to 15 different parasite taxa infecting these three pentanchids from Icelandic waters (Higueruelo et al., Reference Higueruelo, Schaeffner, Soler-Membrives and Dallarés2026). The mentioned study highlighted the potential parasite biodiversity and host-parasite associations still to be revealed in deep water marine habitats and included an undescribed species of Calicotyle (Higueruelo et al. Reference Higueruelo, Schaeffner, Soler-Membrives and Dallarés2026).
The present study provides the morphological description of this new species of Calicotyle, recovered from Apristurus aphyodes, Apristurus laurussonii, and Galeus murinus off Iceland. In addition, a phylogenetic framework is provided and patterns related to morphology and host-specificity of the genus are discussed.
Materials and methods
Collection of specimens
Twenty specimens of A. aphyodes, 20 A. laurussonii, and 32 G. murinus were obtained between 2022 and 2024 during the Icelandic Autumn Groundfish Survey performed annually by the Marine and Freshwater Research Institute of Iceland (MFRI). Specimens were obtained by bottom trawling using the research vessels Árni Friðriksson and Breki, at depths ranging from 700 to 1,300 m in Southwestern Icelandic waters in the North Atlantic Ocean (Fig. 1).
Map of the study area. Red dots indicate the sampling stations where the pentanchid sharks examined for the presence of Calicotyle carmenae n. sp. were collected.

A sub-sample of spiral valves of the three species was removed onboard and stored either in 95% ethanol or subjected to boiling saline and later transferred to 4% buffered formalin for molecular and morphological identification purposes, respectively. The remaining individuals were frozen at –20°C until further parasitological examinations. In the laboratory, both frozen shark individuals and already fixed spiral valves were examined for the presence of parasites and specimens of Calicotyle were stored in 70% ethanol. Individuals were collected from the spiral valve and were normally found attached to the mucosa of the rectal region. Prevalence and intensity data are provided only for the type-host (sensu Bush et al. Reference Bush, Lafferty, Lotz and Shostak1997), for which the largest data set was available.
Morphological study
For morphological observations, monogeneans were stained either in haematoxylin or iron acetocarmine, dehydrated in a graded series of ethanol, cleared in clove oil, and permanently mounted on microscope slides in Canada balsam. The haptor, which covered part of the body in ventral view, was detached from the body of few specimens prior to staining in order to reveal the extension of internal structures. For proper observations of haptoral features, both hamuli from three specimens recovered from A. aphyodes were detached and mounted in Canada balsam. Drawings were made with the aid of a drawing tube attached to an Olympus BH light microscope, and measurements obtained with a stage micrometer. All measurements are reported in micrometres (μm) as the range followed, in parentheses, by the mean ± standard deviation and the number of measurements taken (n). The description provides exclusively measurements from specimens found in spiral valves of A. aphyodes which have been exposed to hot, almost boiling, saline prior to fixation in formalin.
Three additional specimens were examined using scanning electron microscopy (SEM). Specimens were dehydrated in a graded ethanol series, placed in hexamethyldisilazane (HMDS), and allowed to air dry. After dehydration, samples were coated with an alloy of gold (80%) and palladium (20%) at 5 nm thickness using an Emitech K550X sputter coater and subsequently examined with a SEM Zeiss Merlin high-resolution scanning electron microscope (Carl Zeiss Microscopy GmbH, Jena, Germany).
The terminology for morphological characteristics follows Chisholm et al. (Reference Chisholm, Hansknecht, Whittington and Overstreet1997). Type and voucher material have been deposited in the Naturhistorisches Museum (Vienna, Austria) (NM), the Natural History Museum (London, United Kingdom) (NHM), and in the helminthological collection of the Zoology unit of the Universitat Autònoma de Barcelona (Barcelona, Spain) (UAB).
Molecular and phylogenetic analyses
A specimen from each host was used for genomic DNA extraction. DNA extraction was performed using a DNA QIA Cube HT (QIAgen) or with a QIAgen DNA extraction kit (QIAgen) following manufacturer’s instructions. Nuclear large subunit ribosomal DNA (28S rDNA) was amplified through polymerase chain reaction (PCR) as described in Constenla et al. (Reference Constenla, Padrós and Palenzuela2014) adjusted for Taqman Expression mastermix. Amplification was performed using one of two primer sets. In some reactions, primers C1 (5′-ACCCGCTGAATTTAAGCAT-3′) and D2 (5′-TGGTCCGTGTTTCAAGAC-3′) were used, together with the internal primers Rob1 (5′-GTC CAA TAG CAA ACA AGT CCC G-3′) and Rob2 (5′-CAC GYA CTR TTT ACT CTC-3′) (Chisholm, Morgan et al. Reference Chisholm, Morgan, Adlard and Whittington2001; Hassouna et al. Reference Hassouna, Mithot and Bachellerie1984). In other reactions, amplification was carried out using the primers LSU5 (5′-TAGGTCGACCCGCTGAAYTTAAGCA-3′) and 1500R (5′-GCTATCCTGAGGGAAACTTCG-3′) (Littlewood et al. Reference Littlewood, Curini-Galletti and Herniou2000; Olson et al. Reference Olson, Cribb, Tkach, Bray and Littlewood2003). Obtained PCR products were analysed by capillary electrophoresis using a High-Resolution DNA kit in a QIAxcel Advanced Instrument and viewed in the QIAxcel ScreenGel (QIAgen) or analysed on RedGel-stained 1% TAE agarose gels. Sequencing of PCR products was performed by Genewiz-Azenta (Leipzig, Germany) or Macrogen Inc. (Seoul, South Korea) using either the Sanger method or capillary electrophoresis, respectively. Obtained sequences were assembled either with the software Sequencher 5.4.6 from Gene Codes Corporation or with BioEdit 7.7.1 (Hall Reference Hall1999).
For phylogenetic analyses, the obtained sequences were aligned with the Muscle algorithm as implemented in MEGA v11 together with available sequences of Calicotyle and Dictyocotyle in GenBank that exceeded 900 base pairs (bp). Empruthotrema orashken Irigoitia, Braicovich, Rossin & Timi, Reference Irigoitia, Braicovich, Rossin, Canel, Levy, Farber and Timi2019 (Monocotylidae) (GenBank accession number MN190269) was used as an outgroup. The obtained alignment was trimmed to the shortest sequence length.
Maximum likelihood (ML) and Bayesian inference (BI) algorithms were used for phylogenetic tree reconstruction. Prior to analyses the best-fit model of nucleotide substitution and model parameters (i.e. nst, rates) were determined with jModelTest v2.1.4 (Darriba et al. Reference Darriba, Taboada, Doallo and Posada2012) using the Akaike Information Criterion (AIC) and the Bayesian Information Criterion (BIC), respectively. The best-fitting model selected for the ML and BI algorithms was GTR + G (nst = 6, rates = gamma, ngammacat = 4). Maximum likelihood analyses were performed in PhyML 3.0 (Guindon et al. Reference Guindon, Dufayard, Lefort, Anisimova, Hordijk and Gascuel2010) with a non-parametric bootstrap of 100 replicates. Bayesian inference analysis was carried out with MrBayes 3.2.7 (Ronquist et al. Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhn, Larget, l, Suchard and Huelsenbeck2012) through the CIPRES Science Gateway. In this case, log likelihoods were estimated over 10,000,000 generations using Markov chain Monte Carlo searches on two simultaneous runs of four chains, sampling trees every 1,000 generations. The first 25% of the sampled trees were discarded as ‘burn-in’, and a consensus topology and nodal support estimated were calculated as posterior probability values (Huelsenbeck and Ronquist Reference Huelsenbeck and Ronquist2001) from the remaining trees. Phylograms were edited with FigTree v.1.4.4 and Adobe Illustrator CS6. Pairwise genetic distance matrices (‘uncorrected p-distance’ and ‘number of differences’ models) were calculated in MEGA v11.
Results
Phylogenetic analyses
The newly-obtained 28S rDNA sequences were 1,018 bp long for a specimen of Calicotyle ex A. aphyodes, and 1,167 bp long for specimens of Calicotyle ex A. laurussonii and G. murinus. The three sequences have been deposited in the GenBank database under accession numbers PX282454, PX282455, and PX282456, respectively. The resulting alignment was 1,016 bp in length. Newly generated sequences were identical and, when compared to sequences from those of congeneric species, showed the lowest genetic divergence (8.2%, 74 bp) with Calicotyle japonica Kitamura, Ogawa, Shimizu, Kurashima, Mano, Taniuchi & Hirose, Reference Kitamura, Ogawa, Shimizu, Kurashima, Mano, Taniuchi and Hirose2010 infecting the shortspine spurdog, Squalus mitsukurii Jordan & Snyder 1903 (Squalidae) and the greatest divergence (11.1%, 99 bp) with Calicotyle kroyeri Diesing, 1850 infecting the cuckoo skate, Leucoraja naevus (Müller and Henle 1841) (Rajidae) (Table 1).
Nucleotide genetic divergence for 28S rDNA among species of Calicotyle and Dictyocotyle included in the molecular phylogenetic analyses, based on an alignment of 1,016 bp. Values below the diagonal are expressed as a percentage (p-distance ×100) while values above the diagonal represent number of bp differences. Accession numbers and references to the original studies from which the sequences were obtained are also provided

The newly generated sequences formed a strongly supported clade with those of other members of the genus infecting selachians and holocephalans (Fig. 2), namely Calicotyle affinis Scott, 1911, C. japonica, C. palombi Euzet & Williams 1960, and C. stossichi Braun, 1899, from which they differed by 8.2–9.1% (74–85 bp). This group of Calicotyle spp. from selachians and holocephalans was sister to a clade including species of Calicotyle infecting batoids, from which the newly generated sequences differed by 9.5–11.9% (84–99 bp). Collectively, sequences of Calicotyle infecting sharks and chimaeras differed by 9.5–12.6% (84–113 bp) from batoid-infecting congeners. Among the species of Calicotyle infecting batoids, there appear to be two clades with very strong support from both BI and ML algorithms (Fig. 2). One group infects batoids of the family Urolophidae whereas the other infects members of the family Rajidae. Sequences of Dictyocotyle form a separate clade differing from the Calicotyle sequences by 9.9–12.4% (88–111 bp) (Table 1).
Maximum likelihood (ML) and Bayesian inference (BI) consensus phylogenetic tree constructed using new partial 28S ribosomal DNA sequences of Calicotyle carmenae n. sp. and retrieved sequences from GenBank of the subfamily Calicotylinae. Outgroup: Empruthotrema orashken. Nodal support values for BI and ML analyses are indicated as BI/ML. The scale bar indicates the expected number of substitutions per site. Host families parasitised are displayed as black figures.

Description
Family Monocotylidae Taschenberg, 1879
Subfamily Calicotylinae Monticelli, 1903
Genus Calicotyle Diesing, 1850
Calicotyle carmenae n. sp. (Figs. 3–5)
Type-host: White ghost catshark, Apristurus aphyodes Nakaya & Stehmann 1998 (Carcharhiniformes: Pentanchidae).
Other hosts: Iceland catshark, Apristurus laurussonii (Saemundsson 1922); mouse catshark Galeus murinus (Collett 1904) (Carcharhiniformes: Pentanchidae).
Type-locality: Southwestern Icelandic waters (North Atlantic Ocean) at the edge of the Icelandic continental shelf, between the Reykjanes Ridge and the Denmark Strait (63°4′48″N, 26°31′48″W–65°15′36″N, 28°9′36″W), at depths between 700 and 1,300 m (Fig. 1).
Site of infection: Rectum, spiral valve.
Prevalence and intensity of infection: Prevalence 30.4%, mean intensity 3.6 in A. aphyodes.
Type-material: The holotype and paratypes, including an hologenophore, are deposited in NHM (accession numbers NHMUK2026.3.26.2 and NHMUK2026.3.26.3). Additional paratypes are deposited in NM (accession number NHMW-ZOO-EV-M5899) and in the helminthological collection of the Zoology Unit of UAB (accession numbers M9-M11).
Representative DNA sequences: Partial sequences of the nuclear large subunit ribosomal RNA gene (28S rDNA), 1,018–1,168 bp in length, deposited in GenBank under accession numbers PX282454, PX282455, and PX282456.
Conservation assessment: Least concern (conservation assessment methodology for animal parasites (CAMAP) criterion 6).
Etymology: This species is dedicated to Carmen Fernández, mother to the first author (AH).
ZooBank registration: urn:lsid:zoobank.org:act:2424544F-CB6C-481A-B6EA-41FC9E4646AD
[Description based on specimens recovered from the type-host, A. aphyodes: 12 stained and mounted adult specimens (including the holotype and one hologenophore), three specimens examined with SEM, and dissected hamuli of three additional specimens. Metrical data of specimens recovered from other hosts, namely A. laurussonii and G. murinus, are provided as supplemental material in Table S1.]
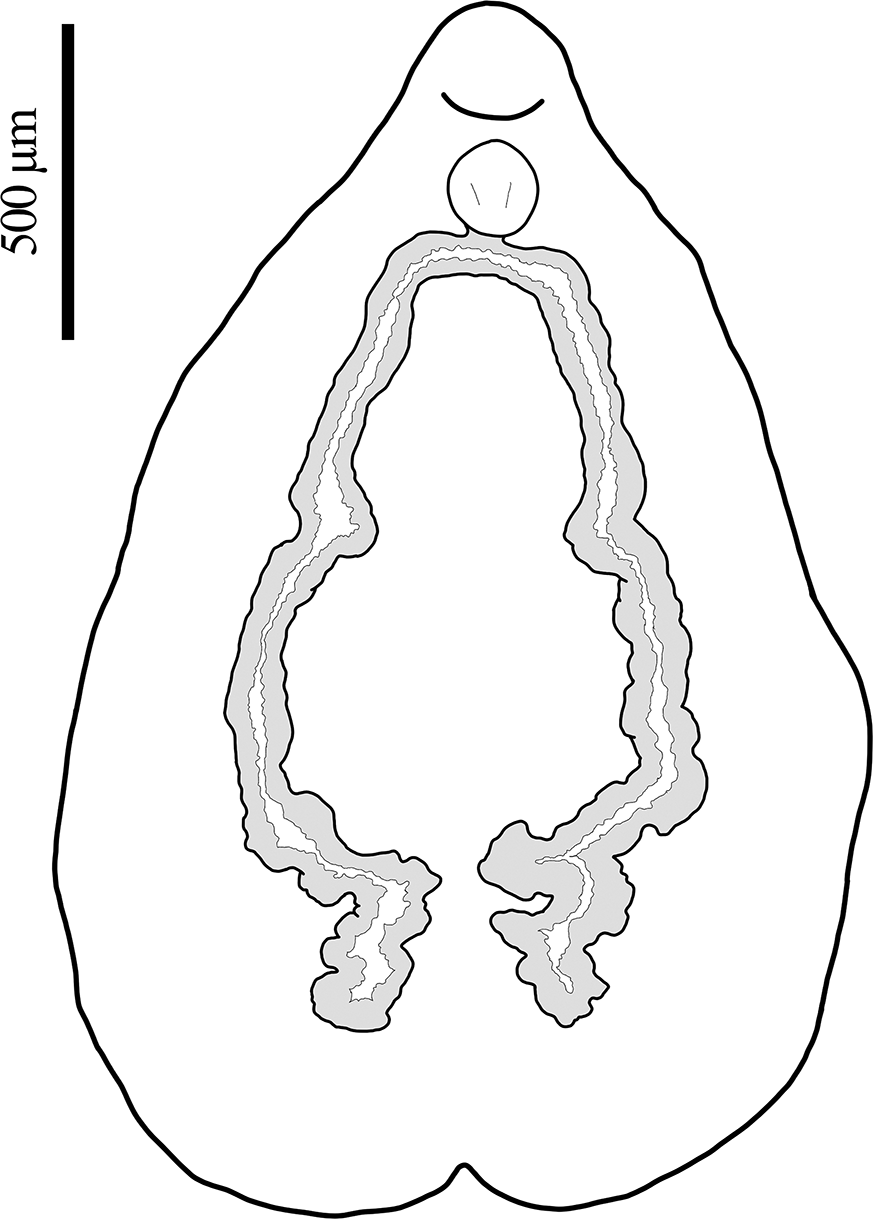
Body including haptor 2,561–3,537 (3,146 ± 431, n = 4) in length, dorsoventrally flattened; body proper pyriform, 1,902–3,049 (2,498 ± 492, n = 5) in length, greatest width at level of median region of testicular mass 1,902–2,244 (2,024 ± 190, n = 3), narrows significantly at level of rounded cephalic region (Figs. 3A; 5A, B). Haptor subcircular, bilaterally symmetrical, shallowly seven-septa lobed, 1,366–2,012 (1,555 ± 242, n = 6) in diameter, connected to main body by rather short peduncle; central loculus slightly immersed; peripheral loculi seven, approximately equal, further immersed, each delimited by muscular septum (Fig. 5D). Rim of haptor muscular, folds inwards with an undulation, becomes wider at level of radial septum (Fig. 3A). Hamuli in single pair, 211–227 (219 ± 6, n = 6) long, 59–72 (67 ± 6, n = 5) in maximum width, directed towards centre of haptor, provided with guard and relatively short blade (Fig. 3C), each at either side of posterior median loculus; tip of hamuli expands over marginal rim (Fig. 5E). Marginal hooklets 10–13 (11 ± 1, n = 5) long, located inside incurved margin of muscular rim of haptor, scattered throughout, apparently arranged symmetrically (Fig. 3D). Ratio of haptor length to body length 0.43–0.53 (0.48 ± 0.004, n = 4). Buccal cavity ventral, subterminal, not surrounded by sucker, encircled by several radial muscle fibres, surrounded by numerous small papillae (Fig. 5H). Eye-spots absent. Pharynx bulbous, 148–249 (184 ± 27, n = 10) in diameter; oesophagus indistinct. Oesophageal glands in two groups at each side, located between inferior part of pharynx and beginning of vitellaria, with ducts leading towards posterior centre of pharynx (Fig. 3F). Intestinal caeca two in number, without diverticula, bifurcate from pharynx and extend posteriorly along internal margin of vitellarium, with slight medial turn at level of Mehlis’ gland and second greater turn posterior to testicular mass, end blindly in several lobes at medial level (Fig. 4).
Line drawings of Calicotyle carmenae n. sp. from Apristurus aphyodes off Iceland. A, holotype, entire view, hooklets not in scale for the sake of visibility; B, male terminal genitalia; C, hamulus, lateral view; D, hooklet, lateral view; E, tetrahedral egg, mostly collapsed; F, anterior body region displaying terminal genitalia, detailed ventral view.

Line drawing of Calicotyle carmenae n. sp. from Apristurus aphyodes off Iceland with the haptor removed and illustrating the path and morphology of intestinal caeca.

Scanning electron micrographs (SEM) of Calicotyle carmenae n. sp. from Apristurus aphyodes off Iceland. A, whole specimen, ventral view; B, anterior region of body, ventral view; C, central region of body, ventral view, with male copulatory organ (MCO) everting from genital and vaginal pores (arrows); D, haptor; E, tip of haptor hamuli, close view; F, vaginal pore, close view; G, tip of MCO, close view; H, papillae from edge of buccal cavity, close view.

Testicular mass follicular, with 61–72 (65 ± 6, n = 3) testes; testes 57–133 (101 ± 19, n = 27) in diameter, occupy intercaecal space from level of ovary to region anterior to folding of intestinal caeca, distributed in two continuously juxtaposed (partially interspersed) rows. Vas deferens emerges from anterosinistral portion of testicular mass, runs approximately parallel to left caecum with some bends, thickens posterior to intestinal bifurcation to form the seminal vesicle. Seminal vesicle curved and elongate, maximum width 41–73 (52 ± 11, n = 10), turns posteriorly, narrows before connecting to proximal portion of the ejaculatory bulb. Prostatic gland cells weakly stained, larger than Mehlis’ gland cells, observed between vitellaria at level of ejaculatory bulb; prostatic gland ducts lead towards proximal ejaculatory bulb; ejaculatory bulb 61–80 (71 ± 6, n = 10) long by 76–95 (89 ± 6, n = 10) wide, with two spherical internal chambers; distal ejaculatory bulb ventrally elongated, connects to funnel-shaped base of male copulatory organ (MCO). MCO sclerotized, 573–783 (678 ± 63, n = 10) long, emerges posteriorly for short distance, then recurves, passes distally to ejaculatory bulb before turning and extending posteriorly towards proximal ejaculatory bulb, terminates at level of ootype, where it bends in different forms, displaying variability (Figs. 3B; 5G). Ejaculatory duct not observed. Common genital pore opens ventrally, just anterior to ootype, forms external bulbous structure when MCO is evaginated (Fig. 5C).
Ovary elongated, 40–119 (67 ± 17, n = 30) in width, lobular, extends from medial body region and dorsal to testicular mass, turns ventrally over right caecum and extends again towards medial region dorsal to fertilisation chamber; posterior region of ovary enlarged, 67–171 (108 ± 28, n = 10) in width, contains small immature oocytes; anterior ovarian region containing larger, mature oocytes. Vitellarium dendritic occupies space between body margins and caeca, in two lateral fields, from level of intestinal bifurcation to posterior end of body, not confluent posteriorly. Vitelline ducts dense, arise from inner medial margins of vitellarium, form transverse vitelline ducts at upper half of body, anterior to ovary. Transverse vitelline ducts 171–405 (288 ± 51, n = 20) long, broader in medial region, 52–138 (94 ± 21, n = 20), before narrowing considerably at conjunction at level of seminal receptacle. Fertilisation chamber oval, 57–133 (98 ± 25, n = 10) in diameter, dorsal to transverse vitelline ducts, connected to base of ootype through short duct. Seminal receptacle 62–140 (98 ± 23, n = 10) long by 86–181 (114 ± 29, n = 10) wide, with short marginal striations. Transverse vitelline ducts obscure junction of vaginae and seminal receptacle. Vaginae two, each 219–419 (317 ± 63, n = 14) long, U-shaped, uniform along their length, arise from seminal receptacle, located ventrally to conjunction of transversal vitelline duct, run approximately parallel to transverse vitelline ducts, turn anteriorly prior to reaching caeca, and slightly twist laterally before reaching vaginal pores. Small glands distributed along length of vaginae. Vaginal pores intercaecal (or slightly overlapping caeca), open ventrally at level of common genital pore (Fig. 5C, F). Mehlis’ gland posterior to transverse vitelline ducts, overlap ovary ventrally at right side, with two groups of collecting ducts that connect to base of ootype from both sides. Ootype muscular with thick walls, 110–171 (133 ± 21, n = 10) long by 112–150 (128 ± 13, n = 10) wide, lumen triangular in ventral view. Eggs tetrahedral, with short appendage, approximately 114 in diameter (n = 1) (Fig. 3E), present in 36% of specimens.
Remarks
Calicotyle carmenae has been described from the rectum of pentanchid catsharks from Icelandic waters. This constitutes the first record of a Calicotyle species in a pentanchid host, as well as the northernmost occurrence of a species of this genus reported to date. Present specimens are assigned to Calicotyle based on the following morphological characteristics: a haptor with one central and seven peripheral loculi, two hamuli and several hooklets, two vaginae, an ejaculatory bulb with two spherical internal chambers, and follicular testes (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997).
Calicotyle carmenae is distinguished from all other congeners by a unique combination of morphological characters. It possesses an MCO that loops once, a feature shared only with Calicotyle asterii (Szidat, 1970) Suriano, 1977, C. australis Johnston, 1934, C. mitsukurii Goto, 1894, and C. similis (Szidat, 1972) Timofeeva, 1985 (see Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997). However, C. carmenae has U-shaped vaginae with intercaecal vaginal pores slightly overlapping the intestinal caeca, in contrast to V-shaped vaginae and vaginal pores lateral to the caeca (i.e. extracaecal) of C. australis and C. mitsukurii, among other differences. It also differs from C. similis in the position of the vaginal pores, as well as in the absence of eye-spots and a vitellarium that is not confluent posteriorly. The absence of eye spots and shape of the ovary further distinguish C. carmanae from C. asterii. In general body appearance, C. carmenae closely resembles Calicotyle urobati Bullard & Overstreet Reference Bullard and Overstreet2000 and C. urolophi Chisholm, Beverley-Burton & Last 1991 by having a pyriform body with a large haptor (Bullard and Overstreet Reference Bullard and Overstreet2000; Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997). All three species are the only ones with a haptor length to total body length ratio larger than 0.4; however, they differ in internal structures such as the shape of the MCO, the position of vaginal pores, or the presence of anterolateral glands or eye-spots.
Unfortunately, all eggs were found mostly collapsed, though their tetrahedral shape, consistent with other congeneric species, could still be distinguished. A distinctive characteristic feature of C. carmenae is the subcircular, shallowly seven-septa lobed appearance of the haptor, in contrast to the predominantly circular haptors described in other species of Calicotyle (e.g. Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997; Neifar et al. Reference Neifar, Euzet and Ben Hassine2001; Ñacari et al. Reference Ñacari, Sepulveda, Droguet, Escribano and Oliva2020).
Papillae are documented for the first time in species of Calicotyle, distributed around the buccal cavity. Although they might occur in other members of the Calicotylinae, papillae have previously been reported only in Dictyocotyle coeliaca, where they occur not only around the buccal cavity but also around the genital and vaginal pores (Poddubnaya et al. Reference Poddubnaya, Hemmingsen and Gibson2015).
In taxonomic descriptions of species of Calicotyle, prostatic gland cells are rarely mentioned (e.g. Bullard and Overstreet Reference Bullard and Overstreet2000; Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997), and some authors have specifically noted the absence of ‘male accessory glands’ in C. australis (Glennon et al. Reference Glennon, Chisholm and Whittington2005; Whittington et al. Reference Whittington, Barton and Lester1989). It was not until transmission electron microscopy (TEM) studies of C. affinis that prostatic gland cells with ducts associated with the ejaculatory bulb were observed and described (Poddubnaya et al. Reference Poddubnaya, Hemmingsen, Bruñanská and Gibson2018a). In the present study, these cells were observed in whole mounts and are illustrated here for the first time for a species of Calicotyle.
This study also presents an SEM-based observation of an evaginated MCO. In this condition, the genital pore formed an external protruding bulbous structure, a condition which was not observed in individuals with non-evaginated MCO’s. Given that this marks the first record describing this condition, it remains unclear whether this is specific to the species examined or if it may also occur in other monocotylids.
Measurements in the description are provided only for individuals parasiting A. aphyodes, as it was the only host from which a sufficient number of specimens could be recovered from non-frozen material. Nonetheless, it is worth noting that measurements from other host species reported in Table S1 suggest potential intraspecific size variation linked to host size: smaller individuals were recorded in G. murinus, whereas larger specimens occurred in A. laurussonii, consistent with host size differences. However, the limited sample size and the fact that specimens from both A. laurussonii and G. murinus were not exposed to hot saline prior to fixation in formalin prevent definitive conclusions.
Interestingly, during the collection of specimens, some individuals were found firmly attached to the rectal wall, leaving clearly visible marks from the haptor fixation upon removal (Fig. 6).
Haptoral marks (arrows) of Calicotyle carmenae n. sp. on the rectal mucosa of Apristurus aphyodes. In the lower-left, detail of haptoral marks. Scale bar = 5 mm.

Several parasite species have been assessed for their conservation status (Ingelbrecht et al. Reference Ingelbrecht, Morgan, Lear, Fazeldean, Lymbery, Norman and Martin2022; Morgan et al. Reference Morgan, Tang and Peverell2010; Van der Spuy et al. Reference Van Der Spuy, Smit and Schaeffner2022). Kwak et al. (Reference Kwak, Heath and Cardoso2020) developed a conservation assessment methodology for animal parasites (CAMAP), which has since been applied by some authors, for example, to Carrassoniella sinuosiceps (Williams 1959) from the bluntnose sixgill shark, Hexanchus griseus (Bonnaterre 1788) (see Dallarés et al. Reference Dallarés, Barría and Schaeffner2025). The assignment of C. carmenae to the ‘Least concern’ category results from the application of criterion 6 of CAMAP, since required ecological and historical data (CAMAP criteria 1–5) have currently not been available for the new species. Calicotyle carmenae infects at least three pentanchids, all of which are suspected to have stable population trends and are currently assessed as ‘Least concern’ in the IUCN Red List (Finucci and Armstrong Reference Finucci and Armstrong2024; Kulka et al. Reference Kulka, Cotton, Anderson, Crysler, Herman and Dulvy2020; Rigby et al. Reference Rigby, Iglésias and Rohner2024). However, parasites, and monogeneans in particular, may be even more vulnerable than their hosts to anthropogenic disturbances (Gilbert and Avenant-Oldewage Reference Gilbert and Avenant-Oldewage2021; Wood et al. Reference Wood, Lafferty and Micheli2010), and therefore might face an increased risk of co-extinction (Koh et al. Reference Koh, Dunn, Sodhi, Colwell, Proctor and Smith2004), underscoring the importance of maintaining stable host populations to support parasite conservation.
Discussion
The present study integrates morphological observations with molecular techniques, supporting the recognition of C. carmenae as a distinct species. Ribosomal DNA sequences have been used as a taxonomic tool since the beginning of molecular phylogenetic studies and are considered to be markers for species differentiation (Aiken et al. Reference Aiken, Bott, Mladineo, Montero, Nowak and Hayward2007). The large subunit ribosomal DNA (28S rDNA) has been widely applied in numerous studies of monocotylid monogeneans, where it provides adequate resolution for species discrimination (Bullard et al. Reference Bullard, Warren and Dutton2021; Chisholm Whittington et al. Reference Chisholm, Whittington, Morgan and Adlard2001, Chisholm, Morgan et al. Reference Chisholm, Morgan, Adlard and Whittington2001; Fehlauer-Ale et al. Reference Fehlauer-Ale, Timothy, Littlewood, Fehlauer-Ale and Littlewood2011).
Although the genus Calicotyle underwent a comprehensive revision by Chisholm et al. (Reference Chisholm, Hansknecht, Whittington and Overstreet1997), the absence of well-preserved specimens for certain taxa (e.g. Calicotyle inermis Woolcock, 1936, C. mitsukurii, and C. ramsayi Robinson, 1961) and several gaps of knowledge in relation to biological traits (e.g. the development of individuals from juvenile to adult) (see Ñacari et al. Reference Ñacari, Sepulveda, Droguet, Escribano and Oliva2020), combined with the subsequent description of new species incorporating additional morphological characters, has resulted in inconsistencies and a lack of standardisation in the characterization of certain diagnostic features. These include the haptoral accessory structures, consisting of a pair of hamuli (absent only in C. japonica and C. inermis) and hooklets, which have been (if observed) reported in varying numbers. The hooklets are very small and difficult to observe without damaging the haptor, particularly when their position within the muscular rim is perpendicular, in which case only the tip is visible. Given that haptoral accessory structures are generally a conserved feature among monocotylid genera (Chisholm et al. Reference Chisholm, Wheeler and Beverley-Burton1995), it is likely that species of Calicotyle possess a fixed number of hooklets, most likely 14, arranged symmetrically, as observed in C. kroyeri (Chisholm Reference Chisholm, Hansknecht, Whittington and Overstreet1997). Similarly, other morphological features of the haptor are often insufficiently described and characterized. The shape of the haptor is typically described as ‘circular’, rarely addressing or illustrating the morphology of the haptor muscular rim. Further research is needed to clarify whether two haptoral morphologies (i.e. circular and subcircular seven-lobed, as observed in C. carmenae) occur within the genus and whether this represents a diagnostic trait among species.
In the review of Chisholm et al. (Reference Chisholm, Hansknecht, Whittington and Overstreet1997), the female reproductive system has not been addressed in detail beyond the V- or U-shaped vaginae. Details on the presence or absence of vaginal glands have not been discussed. These glands were observed in the present study and for additional species (e.g. C. californiensis, C. urobati) (see Bullard and Overstreet Reference Bullard and Overstreet2000). Therefore, it remains speculative if these glands represent a unique synapomorphic feature within the Calicotylinae. In some species descriptions, such as the one of C. carmenae, two distinct chambers associated with the female reproductive system were reported: the first is situated at the point where the vaginae converge, and the second, positioned posteriorly, connects the first chamber with the ootype and the ovary. Where only a single chamber was described in previous species records, it has generally been referred to as the seminal receptacle. However, in cases where two chambers have been described, the terminology becomes inconsistent. The first chamber is termed the ‘proximal region of the vaginae’ in C. californiensis and C. confusa Kanaeva, Vodiasova, Ha, Uppe & Dmitrieva Reference Kanaeva, Vodiasova, Ha, Uppe and Dmitrieva2025 (Bullard and Overstreet Reference Bullard and Overstreet2000; Kanaeva et al. Reference Kanaeva, Vodiasova, Ha, Uppe and Dmitrieva2025), ‘vaginal chamber’ in C. cutmorei Kritsky & Chisholm Reference Kritsky and Chisholm2020 (Kritskyand Chisholm Reference Kritsky and Chisholm2020), and ‘seminal receptacle’ in C. affinis (see Poddubnaya et al. Reference Poddubnaya, Hemmingsen and Gibson2018b). In contrast, the second chamber is referred to as the ‘seminal receptacle’ in C. cutmorei and C. californiensis (Bullard and Overstreet Reference Bullard and Overstreet2000; Kritsky and Chisholm Reference Kritsky and Chisholm2020), ‘ootype’ in C. confusa (Kanaeva et al. Reference Kanaeva, Vodiasova, Ha, Uppe and Dmitrieva2025), and ‘fertilisation chamber’ in C. affinis (Poddubnaya et al. Reference Poddubnaya, Hemmingsen and Gibson2018b). Based on the anatomical arrangement of these chambers, the first chamber at the convergence of the vaginae may primarily function as a seminal receptacle, storing spermatozoa, as demonstrated by TEM observations in C. affinis (see Poddubnaya et al. Reference Poddubnaya, Hemmingsen, Bruñanská and Gibson2018a). The second chamber which connects the distal extremity of the ovary with the ootype and receives the ducts emanating from the seminal receptacle and the vitelline reservoir, likely serves as the site where oocyte fertilisation occurs. Therefore, as implemented in the present description, it might be more appropriate to follow the terminology proposed by Poddubnaya et al. (Reference Poddubnaya, Hemmingsen and Gibson2018b), namely, seminal receptacle and fertilisation chamber.
In line with these observations, in some species (e.g. C. australis, C. confusa, C. kroyeri, C. urobati, C. urolophi, C. vicina Neifar, Euzet & Ben Hassine, Reference Neifar, Euzet and Ben Hassine2001), the vaginae appear to comprise two differentiated regions, with the proximal portion being thicker. These regions are usually referred to as the proximal and distal vaginae, or as the muscular and seminal portions of the vaginae (Bullard and Overstreet Reference Bullard and Overstreet2000; Glennon et al. Reference Glennon, Chisholm and Whittington2005; Kanaeva et al. Reference Kanaeva, Vodiasova, Ha, Uppe and Dmitrieva2025; Neifar et al. Reference Neifar, Euzet and Ben Hassine2001). It is likely that this enlarged section corresponds to a species-specific modification of the seminal vesicle rather than a part of the vaginae itself. However, this assumption requires further verification at the functional level. Detailed examination of the female reproductive system is important, as the features described above may represent diagnostic characters to distinguish closely related species.
It has been suggested that in Calicotyle spp., the absence of a marginal membrane and the presence of a thick haptoral rim have reduced the haptor’s ability to maintain an effective suction seal, leading these species to rely more heavily on their hamuli for attachment (Chisholm et al. Reference Chisholm, Wheeler and Beverley-Burton1995). Chisholm and Whittington (Reference Chisholm and Whittington1998) further argued that suction-based attachment may be less important for internal monocotylids, which are not exposed to strong water currents, compared with species inhabiting the nasal cavities or external surfaces of their hosts. Nonetheless, in the present study, several specimens were observed firmly attached to the host mucosa, leaving clear haptor attachment marks. Some species possess highly reduced hamuli, and the hooklets, located inside the marginal rim, are likely vestigial. Together, these observations suggest that species of Calicotyle may rely on the haptor for fixation to a greater extent than previously thought.
Monogeneans have historically been considered to exhibit high levels of host specificity (Whittington et al. Reference Whittington, Cribb, Hamwood and Halliday2000). However, C. carmenae was recorded from three host species, belonging to two genera of the same family and occurring within a relatively restricted geographical range. Our results are consistent with previous phylogenetic analyses, which suggest that species of Calicotyle tend to parasitize members of the same family, with hosts distributed within relatively small geographical areas (Chisholm, Whittington et al. Reference Chisholm, Whittington, Morgan and Adlard2001). Although C. kroyeri has been reported from 20 host species representing four families (i.e. Rajidae de Blainville 1816; Anacanthobatidae von Bonde & Swart 1923; Narcinidae Gill 1862; and Chimaeridae Rafinesque 1815), across geographically distant areas (Chisholm, Whittington et al. Reference Chisholm, Whittington, Morgan and Adlard2001; Kanaeva et al. Reference Kanaeva, Vodiasova, Ha, Uppe and Dmitrieva2025), phylogenetic analyses by Chisholm, Whittington et al. (Reference Chisholm, Whittington, Morgan and Adlard2001) indicate that many of these records may not refer to the same species. Instead, C. kroyeri appears to have served as a ‘dumping ground’ for individuals found in the cloaca of various species of rays and chimeras. Conversely, some species are presumed to be synonymous, subject to the examination of additional material. Calicotyle affinis is considered potentially synonymous with C. australiensis Rohde, Heap, Hayward & Graham 1992, and C. australis may be synonymous with C. mitsukurii and C. vicina (see Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997; Glennon et al. Reference Glennon, Chisholm and Whittington2005). Furthermore, following an earlier study (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997) and until additional material can be examined, C. sjegi Kusnetzova, 1970 is regarded as a species inquirenda, and C. inermis is considered a species incerta sedis (Chisholm et al. Reference Chisholm, Hansknecht, Whittington and Overstreet1997).
The validity and resurrection of Paracalicotyle for specimens collected from triakid and carcharhinid sharks (i.e. C. stossichi, C. palombi, C. asterii, C. californiensis, C. cutmorei) have been proposed by some authors based on a more elongate body appearance and smaller haptor size of specimens (Bullard and Overstreet Reference Bullard and Overstreet2000; Bullard et al. Reference Bullard, Warren and Dutton2021). However, C. carmenae, which belongs to the same clade, has a pyriform body shape and possesses a large haptor more comparable to the ones of C. urobati and C. urolophi (both from rajid hosts). Although the molecular divergence between the two principal Calicotyle clades (9.5–12.6%) is similar to that observed between Calicotyle and Dictyocotyle (9.9–12.4%), there are no consistent morphological synapomorphies to justify erecting a separate genus based solely on host type (i.e., parasites infecting batoids versus those infecting selachians and holocephalans). Consequently, we concur with the position of Chisholm et al. (Reference Chisholm, Hansknecht, Whittington and Overstreet1997) in retaining the current generic classification, unless subsequent multidisciplinary revisions demonstrate robust morphological or molecular justification for a generic separation.
Overall, the inconsistencies and knowledge gaps identified in this study highlight the need for a comprehensive revision of the subfamily, involving re-examination of deposited material and the acquisition of new specimens and molecular data.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0022149X26101527.
Acknowledgements
We are thankful to two anonymous reviewers for their suggestions and corrections, which have increased the quality of this research article. We are deeply grateful to the Marine and Freshwater Research Institute of Iceland (MFRI) for enabling B.C.S. and A.H. to participate in the Autumn Groundfish Surveys aboard the research vessel, with special appreciation to the survey leads Jón Solmundsson and Klara Björg Jakobsdóttir. We are grateful to Dr. Haseeb Randhawa (University of Iceland) for providing additional samples, and to Þórunn Sóley Björnsdóttir, Samuel Casas Casal, and Heida Sigurðardóttir for their valuable assistance with molecular analyses.
Financial support
This work was funded by a PhD student grant from FI SDUR (AGAUR 2021) awarded to A.H., supported by the Secretariat of Universities and Research of the Generalitat de Catalunya and the European Social Fund, and in parts by the University of Iceland Research Fund (award no. 15539) to B.C.S.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.









 Open access
Open access