


Management Implications
The cumulative effects of altered hydrology and ever-increasing competitive pressure from invasive species have led to declines in forest health and cover in Upper Mississippi River (UMR) floodplain forests, and these trends are likely to continue into the future without active management. Tree plantings in the UMR are often met with limited success, and reforesting areas that have been taken over by invasive species such as Humulus japonicus (Japanese hop) and Phalaris arundinacea (reed canarygrass) poses even more difficulties. The results of this experimental tree planting indicate that large-diameter willow (Salix) cuttings can provide a viable alternative to traditional planting stock in efforts to reforest portions of the UMR that have experienced loss of forest cover coupled with colonization by invasive plant species. Salix is also a good fit for reforesting the lowest-lying areas that can still support tree cover given its high tolerance for flooded conditions.
The Salix cuttings planted at high densities for reforestation in this study had significantly better survival than containerized trees. Height growth was also impressive, with cuttings at the three southerly sites achieving mean annual height growth of about a meter per year, reaching mean cumulative heights of more than 5 m after just three to five growing seasons. Also noteworthy was that invasives cover was significantly lower in the Salix plantings than in the containerized plantings. Maintenance activities are recommended, as they had a significantly positive effect on survival. Sourcing and transporting cuttings of appropriate size may be an obstacle in some locations without viable Salix stock nearby. There are also labor costs involved, but the cost per planted tree is still likely much lower than the purchase price of containerized trees. Damage and loss of planting stock due to Castor canadensis (North American beaver) and other wildlife can also be significant, but using tree-planting guards and other protective measures can deter those actions to some degree.
Introduction
Invasive plant species are increasing in abundance and dominance in the Upper Mississippi River (UMR) and have the ability to dominate and maintain sites in an invasive disclimax community (Barrette et al. Reference Barrette, Bélanger, De Grandpré and Royo2017; Guyon and Cosgriff Reference Guyon and Cosgriff2022; Johnson et al. Reference Johnson, Amatangelo, Townsend and Waller2016; Price et al. Reference Price, Spyreas and Matthews2018; Sparks Reference Sparks2010). Specifically, invasive plant species colonize forest canopy gaps, suppress natural tree regeneration, and negatively impact native species (Guyon and Cosgriff Reference Guyon and Cosgriff2022; Weidlich et al. Reference Weidlich, Florido, Sorrini and Brancalion2020). Over time, this in turn can degrade the compositional and structural diversity of floodplain forests and complicate efforts to restore and sustainably manage these communities. This project evaluated the effectiveness of artificial reforestation methods to reduce the impact of invasive plant species by rapidly reestablishing trees in canopy openings and restoring UMR floodplain forest cover.
Forest communities of the UMR are highly productive, provide habitat for birds and wildlife, control erosion, and improve water quality (Guyon et al. Reference Guyon, Deutsch, Lundh and Urich2012, Reference Guyon, Sloan, Van Essen and Corcoran2016; Johnson and Hagerty Reference Johnson and Hagerty2008; Romano Reference Romano2010; Urich et al. Reference Urich, Swenson and Nelson2002; Yin et al. Reference Yin, Nelson and Lubinski1997). Four broad forest types are commonly recognized in UMR land-cover mapping efforts: Salix community (willow [Salix L.]), Populus community (cottonwood [Populus L.]), floodplain forest, and lowland forest (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019; Dieck et al. Reference Dieck, Ruhser, Hoy and Robinson2015). The Salix community is dominated by black willow (Salix nigra Marshall) and/or sandbar willow (Salix interior Rowlee). The Populus community is dominated by eastern cottonwood (Populus deltoides W. Bartram ex Marshall). The floodplain forest community is dominated by silver maple (Acer saccharinum L.) with common associates such as American elm (Ulmus americana L.), green ash (Fraxinus pennsylvanica Marshall), boxelder (Acer negundo L.), and American sycamore (Platanus occidentalis L.). The Salix and Populus communities are typically recognized as early successional, and eventually transition to the floodplain forest type depending on site elevation and edaphic conditions (Yin et al. Reference Yin, Nelson and Lubinski1997). For example, S. nigra is one of the most flood-tolerant trees in the UMR, and some Salix communities may persist as the dominant forest type in lower-lying, wetter areas (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019). The lowland forest type comprises mast-producing species such as oaks (Quercus), hickories (Carya), and associates such as common hackberry (Celtis occidentalis L.) (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019). The lowland forest type generally occurs on higher-elevation terraces and similar areas and is often considered a late successional community type in this system (Dieck et al. Reference Dieck, Ruhser, Hoy and Robinson2015; Hodges Reference Hodges1997).
However, these important communities have seen a significant decline in acreage, diversity, and health since pre-settlement times due to harvests, conversion to agriculture, and altered hydrology (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019; Theiling and Burant Reference Theiling and Burant2013; Yin Reference Yin1998; Yin et al. Reference Yin, Nelson and Lubinski1997). In addition, community composition has largely shifted to dominance by A. saccharinum, compounded by a lack of adequate natural regeneration in many areas (Guyon and Battaglia Reference Guyon and Battaglia2018; Johnson and Hagerty Reference Johnson and Hagerty2008; Urich et al. Reference Urich, Swenson and Nelson2002). Invasive species are a significant contributor to the ongoing decline in forest community health and diversity by impacting natural forest regeneration (De Jager et al. Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024; Guyon et al. Reference Guyon, Deutsch, Lundh and Urich2012; Urich et al. Reference Urich, Swenson and Nelson2002). Compounding this issue, emerald ash borer (Agrilus planipennis; EAB) is currently spreading throughout the UMR and is causing widespread mortality of F. pennsylvanica, creating numerous additional canopy openings (Burr and McCullough Reference Burr and McCullough2014; De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019; Herms and McCullough Reference Herms and McCullough2014; Siegert et al. Reference Siegert, McCullough, Liebhold and Telewski2014).
Reed canarygrass (Phalaris arundinacea L.) and Japanese hop (Humulus japonicus Siebold & Zucc.) are two of the most common invasive plant species that are causing significant natural regeneration issues in UMR floodplains (Adams et al. Reference Adams, Kauth and Sorenson2011; De Jager et al. Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024; Guyon and Cosgriff Reference Guyon and Cosgriff2022), and they are increasing in dominance and distribution. Phalaris arundinacea is a shade-intolerant perennial grass that establishes in dense, sod-forming monocultures, is widespread in the UMR, and negatively impacts native plant communities in wetland areas (Lavergne and Molofsky Reference Lavergne and Molofsky2004; Maurer and Zedler Reference Maurer and Zedler2002; Spyreas et al. Reference Spyreas, Wilm, Plocher, Ketzner, Matthews, Ellis and Heske2010). Mowing and herbicides are commonly used to control this species, but complete eradication rarely occurs, and sites can quickly be recolonized following treatments. Humulus japonicus is a more recent invasive annual or weakly perennial vine species that has been spreading throughout the UMR over the past 20 yr and is now commonly found in UMR floodplains from Minnesota to the Mississippi River and Ohio River confluence at Cairo, IL (Guyon and Cosgriff Reference Guyon and Cosgriff2022). It readily establishes in forest canopy openings in riverine and wetland habitats, forms dense patches that inhibit native species, and is particularly effective in suppressing tree and shrub regeneration (Pannill and Cook Reference Pannill and Cook2008; Pannill et al. Reference Pannill, Cook, Hairston-Strang and Swearingen2009). Preliminary analyses of auxiliary U.S. Army Corps of Engineers (USACE) forest resource inventory data from Mississippi River Pools 24, 25, and 26 suggest that 15% to 20% of the floodplain forest has H. japonicus existing in the understory, mostly in small canopy gaps (USACE, unpublished data). Humulus japonicus has a growth pattern typical of trailing vine species; it can generally climb to a height of about 5 m in a single growing season and dies back to the ground every winter. When it forms dense stands, it can quickly overtop native vegetation. It has a high light requirement and can rapidly colonize edge areas and canopy gaps in forested floodplain settings, where it can exploit full available sunlight. As a highly shade-intolerant species, it does not survive beneath a closed forest canopy (Guyon and Cosgriff Reference Guyon and Cosgriff2022).
Canopy gaps are a natural occurrence in upland and floodplain forests and are created as trees succumb to disturbance events (e.g., flooding and wind) or senescence (Guyon et al. Reference Guyon, Strassman, Oines, Meier, Thomsen, Sattler, De Jager, Hoy, Vandermyde and Cosgriff2020; Kern et al. Reference Kern, Burton, Raymond, D’Amato, Keeton, Royo, Walters, Webster and Willis2017; King and Antrobus Reference King and Antrobus2001; Poznanovic et al. Reference Poznanovic, Poznanovic, Webster and Bump2014; Runkle Reference Runkle1982). For example, forests of the UMR are dominated by species such as A. saccharinum and P. deltoides that are reaching the end of their life cycles in many areas and can create large canopy gaps when they die. Record flooding in 1993 and 2019 also created significant canopy gaps where between 30% and 70% mortality occurred in mature trees in lower UMR pools in 1993 and upper pools in 2019 (Cosgriff et al. Reference Cosgriff, Nelson and Yin2007; Yin et al. Reference Yin, Wu, Bartell and Cosgriff2009; SA Weiss, L Guyon, N De Jager, R Cosgriff, M Van Appledorn, unpublished data). Additional tree mortality has occurred due to other flooding events and increased windthrow from weakened canopy structure. Another disturbance that is impacting UMR forests is EAB, which causes near 100% mortality in all species of ash (Fraxinus L.) trees (Herms and McCullough Reference Herms and McCullough2014). Fraxinus pennsylvanica is a common tree species within UMR floodplain forests, and as it succumbs to EAB, additional canopy gaps will be created, providing further opportunity for invasive species such as P. arundinacea and H. japonicus to become even more widespread (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019).
Under natural forest successional dynamics, these disturbance events create canopy gaps of varying size and distribution that are then colonized by natural tree regeneration that eventually recruits into the next cohort of canopy trees (Frelich and Graumlich Reference Frelich and Graumlich1994; Hodges Reference Hodges1997; Lorimer Reference Lorimer1977; Oliver and Larson Reference Oliver and Larson1996; Runkle Reference Runkle1982). There has been a significant amount of work related to bottomland forest regeneration and restoration utilizing natural and artificial reforestation techniques along the Mississippi River, although mostly in the Lower Mississippi Alluvial Valley (Allen et al. 2004; King and Keeland Reference King and Keeland1999; Stanturf et al. Reference Stanturf, Gardiner, Shepard, Schweitzer, Portwood and Dorris2009). Species recruitment into canopy gaps in Mississippi River floodplains is tied to inundation regimes, elevation, gap size, soil texture, proximity to parent trees, herbivory, and seedling competition for light and nutrients (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019; Stanturf et al. Reference Stanturf, Conner, Gardiner, Schweitzer and Ezell2004). Initially, native light-seeded, early successional species such as P. deltoides, S. nigra, and P. occidentalis colonize large, open gaps with abundant sunshine. The life history of these species includes producing numerous small seeds that are readily distributed by wind and water. These species are also typically fast growing and may begin to recruit into the midstory and ultimately the canopy layer in a relatively compressed time frame, reducing competition from shade-intolerant ground-layer species in the process. There is potential to use native, fast-growing tree species such as these to quickly close existing forest gaps and reduce the impact of invasive species (Adams et al. Reference Adams, Kauth and Sorenson2011; Kim et al. Reference Kim, Ewing and Giblin2006). For example, Kim et al. (Reference Kim, Ewing and Giblin2006) was able to reduce P. arundinacea by 56% to 68% the second year after planting willow (Salix L.) cuttings in Washington wetlands. Planted cuttings in that study were between 1.27 and 2.54 cm (0.5 and 1 inch) in diameter and planted on 0.6, 0.9, and 1.2 m (2, 3, and 4 ft) spacings. However, we believe that this cutting size and planting density would not be effective in gaps where H. japonicus is prevalent in the UMR. Following a 2017 field trial using P. deltoides cuttings, it was determined that the densest planting attempted (using a 1.2-m spacing) provided H. japonicus with an added advantage, as it created a trellis that allowed H. japonicus to quickly overtake the treatment site. Additionally, trees less than 2.54 cm in diameter were easily pulled over by H. japonicus, consistent with an earlier study using container trees as planting stock (Guyon and Cosgriff Reference Guyon and Cosgriff2022). We hypothesized that Salix cuttings greater than 2.54 cm in diameter planted on a minimum 1.2 by 2.4 m (4 by 8 ft) spacing would provide the best cultural treatment to control shade-intolerant invasive plant species in the UMR.
This project was designed to evaluate the effectiveness of large-diameter Salix cuttings and P. deltoides and P. occidentalis container stock in shading out invasive plant species at four sites within the UMR. Two different planting densities were evaluated for Salix cuttings, and a third for container stock. Salix easily establishes from cuttings, and container stock provides saplings with excellent root reserves for rapid growth within the first growing season. Additionally, we also incorporated maintenance activities to evaluate the effects of standard management practices on invasives species control and tree survival and growth. We believe that fast-growing trees, planted at the right density, have the potential to shade out invasive plant species. The development of effective and novel management approaches to reforestation and invasive species control that require minimal long-term maintenance resources is an important consideration for resource managers, as much of the UMR floodplain forest resource is located on islands and other difficult to access areas.
The overarching goal of this project is therefore to identify effective reforestation methods, with and without traditional maintenance practices (e.g., herbicides, mowing), for use as a cultural treatment in the control and management of invasive plant species in UMR floodplain forest habitats. Specifically, this study addresses the following questions:
-
1. Can artificial reforestation with early successional species be used as a technique to fill canopy gaps and reduce the abundance of invasive plant species such as P. arundinacea and H. japonicus?
-
2. What are the most effective planting densities to achieve expedient canopy closure with minimal maintenance requirements?
-
3. Can successful reforestation be achieved without the use of standard maintenance techniques (e.g., mowing, herbicide application)?
This project will broadly inform river restoration and management by investigating the impacts of active forest management practices on invasive species and forest regeneration. Specifically, it will inform decisions regarding species selection, planting densities, and maintenance regimens necessary to control invasive species and rapidly establish floodplain forest canopy cover in impacted areas.
Materials and Methods
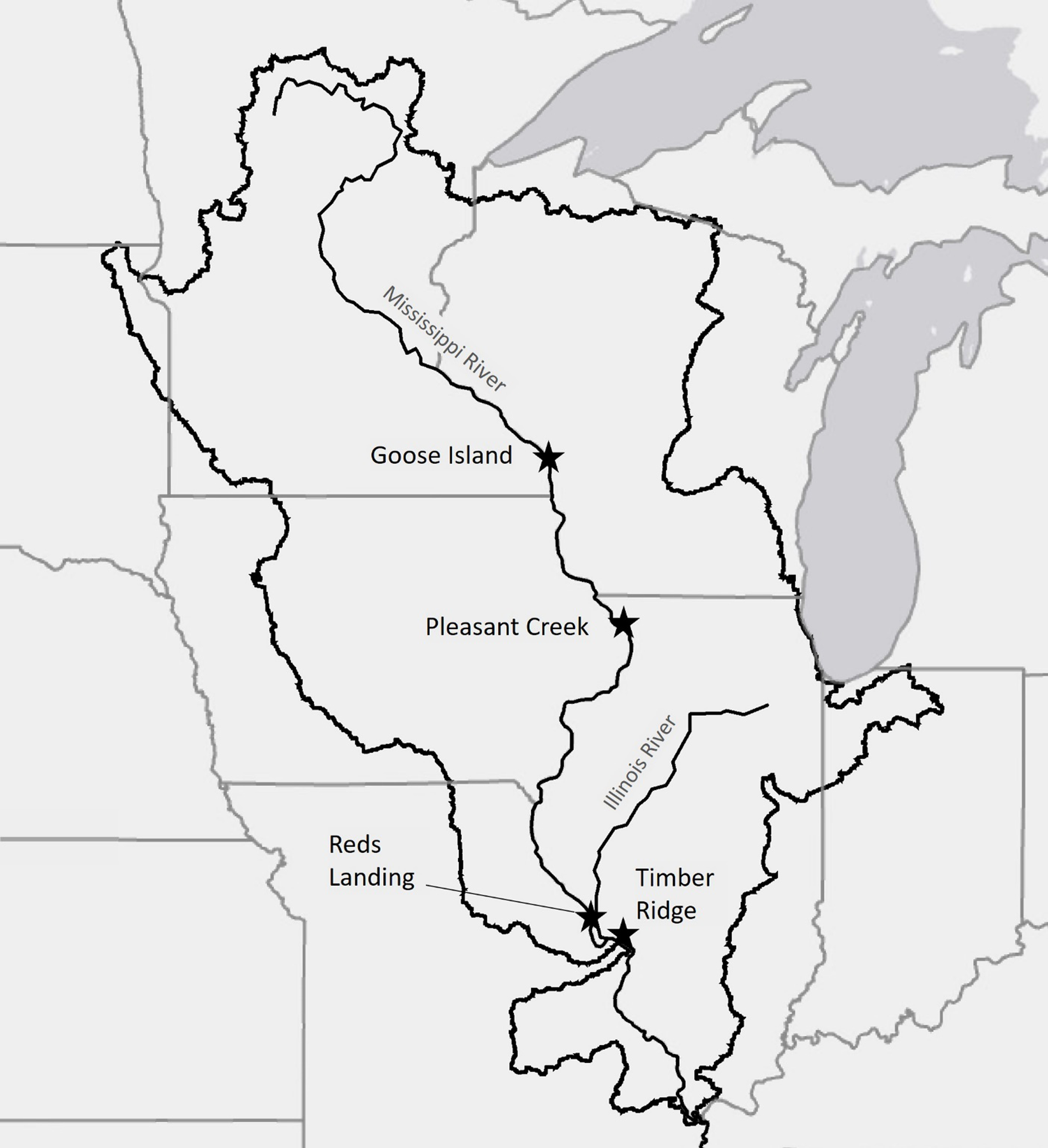
The project was implemented at four different study sites along the UMR, ranging from southern Wisconsin to just north of St Louis, MO (Figure 1; Table 1). Each study site has a different disturbance history and is under a slightly different resource management regime but is significantly impacted by invasive species. The northernmost site is located just south of La Crosse, WI, at Goose Island County Park (43.750, −91.227). This site has a dense population of P. arundinacea, and although H. japonicus has been documented in the general vicinity, it had not yet invaded this site at the time of the study. The Pleasant Creek study site is located near Bellevue, IA, adjacent to the confluence of the Maquoketa and Mississippi rivers (42.185, −90.315). This site had a dense population of P. arundinacea that was recently colonized by and replaced in some areas with H. japonicus. Farther south near Hamburg, IL, Reds Landing (39.175, −90.705) had significant P. arundinacea and minimal H. japonicus present until a flooding event in 2014 brought in additional H. japonicus seed. Currently, the site has a dense population of both H. japonicus and P. arundinacea that is affecting survivorship and growth of tree seedlings and saplings. Finally, Timber Ridge is a wetland restoration and reforestation site located along lower Piasa Creek, a tributary of the Mississippi River near Godfrey, IL (38.962, −90.268). This site has dense populations of H. japonicus, P. arundinacea and Johnsongrass [Sorghum halepense (L.) Pers.], another invasive grass species common to the region. Natural tree regeneration is generally suppressed at all four sites due to the prevalence of invasive plant species.
Study site locations along the Upper Mississippi River, USA. Map credits: Esri, HERE, Garmin, © OpenStreetMap contributors, and the GIS user community.

Study site characteristics.

a Average days per growing season.
To minimize potentially confounding effects of hydrological variation, we selected study sites that are all on the higher end of relative elevational and corresponding UMR floodplain inundation zones (Van Appledorn et al. Reference Van Appledorn, De Jager and Rohweder2023, Reference Van Appledorn, Rohweder and De Jager2024). Goose Island is generally the wettest and most variable site, with plots located in inundation zones flooded on average from 0 to 40 d per growing season, mostly between 10 and 30 d. At Pleasant Creek, plots were located in zones flooded between 0 and 20 d on average, with most flooded fewer than 10 d. At Reds Landing, plots were located in inundation zones flooded 10 to 30 d on average, with most flooded fewer than 20 d. Inundation zone maps were not available for the Timber Ridge site, which lies in a floodplain zone associated with Piasa Creek, a tributary of the Mississippi River with a more unpredictable flood pattern. However, observational evidence suggests that Timber Ridge is likely the driest study site.
For reference, De Jager et al. (Reference De Jager, Thomsen and Yin2012) found that UMR floodplain sites flooded less than 40% of the growing season (72 d) were generally associated with more diverse UMR floodplain forest communities with a wider range of species composition, and De Jager et al. (Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024) further identified growing season inundation durations of approximately 100 d to be an important hydrologic driver of forest loss in areas where P. arundinacea was prevalent. Thus, all of our study sites were well under that threshold and are not in elevational zones where hydrologic stress on its own would be expected to negatively affect floodplain forest composition or forest loss.
There were three replicates at each study site, with each replicate containing four 0.04 hectare (20 by 20 m) tree-planting treatment plots, including one control plot with no planted trees. Half of each planting treatment plot (0.02 hectare) received invasive suppression (hereafter “maintenance”) treatments twice annually and the other half did not. In the tree-planting treatment plots, one contained large-diameter Salix cuttings planted at a 1.2 by 2.4 m (4 by 8 ft) spacing, one contained large-diameter Salix cuttings planted at a 2.4 by 2.4 m (8 by 8 ft) spacing, one contained 13-L (3-gallon) RPM (Root Production Method®) containerized P. deltoides and P. occidentalis saplings planted at a 4.9 by 4.9 m (16 by 16 ft) spacing, and one contained no trees (the control plot). The protocol for the RPM plantings called for planting equal numbers of each species in plots, with random species selection at each specific planting location. Following an unsuccessful field trial using 1.2 by 1.2 m spacings for Salix cuttings, it was determined that the 1.2 by 2.4 m spacing was likely the densest that could still adequately provide room for invasives control. The larger 2.4 by 2.4 m spacing was incorporated to determine whether less dense Salix cutting plantings could still achieve similar results. Although the Salix cuttings were the primary focus of this effort, containerized saplings are commonly used in tree plantings across the UMR, and we felt it was important to include them as well. Planting containerized trees at scale at densities similar to the Salix plantings would not be realistic from a cost-basis perspective, so we accepted the difference in densities between the two types of plantings as unavoidable. The 4.9 by 4.9 m spacing used for them is generally the densest still regularly employed by foresters in the UMR, so that is what we incorporated into this study. The control treatment plots were included to provide a baseline measure of invasive species cover, with and without maintenance treatments, independent of the planted trees. The control plots also provide a basis of comparison for analyzing potential effects of planted trees on reducing invasives cover by shading them out. Long-term survivorship of planted trees is at least in part contingent on their ability to grow taller than competing vegetation, and we surmised that large cuttings should provide an initial size and growth advantage. Cuttings were sourced from local populations of Salix saplings as close to the study sites as practical, which were harvested and cut again into two or three manageable sections to be used for planting. They were transported to the study site locations and planted as soon as practical, generally within a day or two. Planted cuttings were a minimum of 2.54 cm in diameter and 2 m in length, planted to a depth of approximately 0.5 to 1.0 m using a planting auger, and most did not retain apical buds, as they originated from stem sections.
Annual maintenance treatments included two mowing and postemergence herbicide applications (glyphosate at an application rate of 2% v/v using backpack sprayers). Mowing and herbicide applications occurred at the same time, with the first treatment in early summer (e.g., early June) just after the first round of seasonal vegetation sampling, and the second in late summer before the final seasonal sampling period (e.g., late August). Each tree-planting treatment plot was also mowed around the outer edge to reduce the potential for lateral growth of H. japonicus into the plots. At Reds Landing and Timber Ridge, Salix cuttings were planted in the early spring of 2018, and container trees were planted in the late fall of 2019. At Pleasant Creek, container trees and Salix cuttings were planted in the late fall and early spring of 2019/2020. Initially, trees at this site were planted a year previously, but those were all killed by record Mississippi River flooding in 2019 and subsequently replanted a year later. All trees at Goose Island were planted in late fall and early spring of 2020/2021 and were similarly delayed by flooding and additional logistical issues (see Table 1).
Invasive species, other naturally occurring ground-layer vegetation, and planted trees within each study plot were monitored twice a year over the course of the study. Initial sampling occurred in late spring/early summer before the first maintenance treatment each year to get a baseline assessment of invasive species and vegetation cover at the sites. All of the trees planted in each treatment were measured at this time to determine overwintering survivorship. An additional monitoring effort occurred 2 to 4 wk after the second maintenance treatment in early fall to determine seasonal invasive species and vegetation cover and maintenance treatment effects. All of the trees planted in each plot were measured again at this time to determine seasonal survivorship and growth.
Quantifiable ground-layer vegetation data were collected from ten 0.25-m2 quadrats randomly located within each 0.02 hectare split plot using x,y coordinates and a random number generator. Quadrat locations were not changed between spring and fall sampling periods, but were relocated randomly each year. Percent cover of invasives and all other herbaceous layer vegetation was recorded by species. The diameter at breast height (dbh) and height of all planted trees at each study site were recorded by species. The Salix cuttings were planted in rows and were easily identifiable. The RPM trees were planted with stem guards and were also easily identifiable. Any other naturally occurring tree regeneration greater than 1.37 m (4.5 ft) in height was also tallied and recorded by species, dbh, and height if it occurred in the sampling quadrats.
Data analysis tested for the effects of planting density and maintenance treatments on several quantifiable measures of vegetation response, including survivorship and growth of planted trees, invasive species cover, and total vegetation species diversity. We tested the effects of planting type and maintenance on these metrics using generalized linear mixed models. We included planting and treatment and their interaction as main effects, and we included study site as a random effect for each variable modeled. The site random effect also largely accounts for differences in planting date, because planting years were specific to each site in nearly all cases. For this reason, planting date was not included as a separate effect in models. Survivorship was quantified as the percentage of living stems in the fall of 2022 out of the total number originally planted. Growth was assessed in terms of both the average annual change in height and dbh using the following calculation:
 $$\scriptsize {{\rm{Average}}\;{\rm{annual}}\;{\rm{growth}} = {{{\rm{last}}\;{\rm{observed}}\;{\rm{height}}\;{\rm{or}}\;{\rm{dbh}}\; - \;{\rm{first}}\;{\rm{observed}}\;{\rm{height}}\;{\rm{or}}\;{\rm{dbh}}} \over {{\rm{year}}\;{\rm{of}}\;{\rm{final}}\;{\rm{observation}}\; - \;{\rm{year}}\;{\rm{of}}\;{\rm{first}}\;{\rm{observation}}}}}$$
$$\scriptsize {{\rm{Average}}\;{\rm{annual}}\;{\rm{growth}} = {{{\rm{last}}\;{\rm{observed}}\;{\rm{height}}\;{\rm{or}}\;{\rm{dbh}}\; - \;{\rm{first}}\;{\rm{observed}}\;{\rm{height}}\;{\rm{or}}\;{\rm{dbh}}} \over {{\rm{year}}\;{\rm{of}}\;{\rm{final}}\;{\rm{observation}}\; - \;{\rm{year}}\;{\rm{of}}\;{\rm{first}}\;{\rm{observation}}}}}$$
Control plots were not included in analyses of tree survivorship and growth, because they contained no trees. For invasives cover and diversity, we assessed change since first measurement to account for any plot-specific differences in those metrics that may have influenced the final values.
Analyses were run using R version 4.4.0 (R Core Team Reference Core Team2024). Mixed models were run using the lme4 package (Bates et al. Reference Bates, Maechler, Bolker and Walker2015), and assumptions were assessed using plots and statistics generated using the DHARMa package (Hartig Reference Hartig2024). The DHARMa package takes a simulation-based approach to create scaled and standardized residuals from a given model, which are expected to follow a uniform distribution if the model is correctly specified. For all models, simulated residuals were not found to be significantly different from a uniform distribution using Kolmogorov-Smirnov (KS) tests (P > 0.05), and we found no evidence of over- or underdispersion or problematic outliers, so no transformations of variables were found to be necessary. We followed up fitting each model with pairwise Tukey’s honestly significant difference (HSD) post hoc comparison tests with the emmeans package (Lenth Reference Lenth2024).
We used nonmetric multidimensional scaling (NMDS) to represent the vegetation community similarities and differences in a reduced number of dimensions and examined how these communities were positioned relative to a gradient of invasives cover (percent cover of H. japonicus and P. arundinacea combined) and plant community diversity (Shannon H′ values). We used relativized species percent cover estimates as inputs and performed a NMDS ordination analysis for a range of dimensions (2 to 5) using the vegan package (Oksanen et al. Reference Oksanen, Simpson, Blanchet, Kindt, Legendre, Minchin, O’Hara, Solymos, Stevens, Szoecs, Wagner, Barbour, Bedward, Bolker and Borcard2022) and compared stress values, assessing the minimum number of dimensions we could use to best represent the data and allow for reliable interpretations of ordination distances.
Results and Discussion
Tree Survival
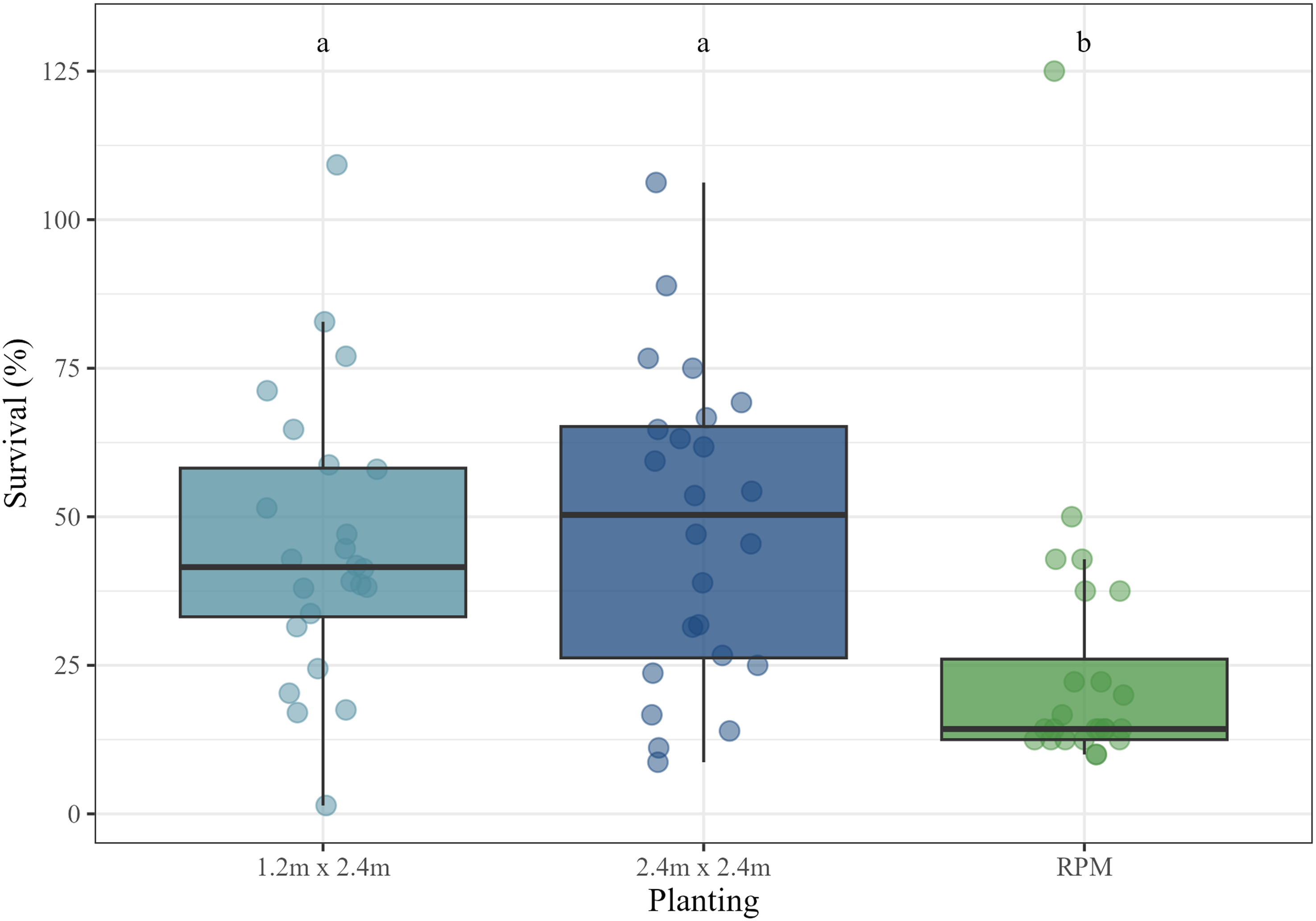
We ran a mixed model of cumulative percent tree survival (from the Fall 2022 sampling period) by maintenance treatment (maintained vs. not maintained), planting treatment (1.2 by 2.4 m spacing, 2.4 by 2.4 m spacing, and RPM container stock), and the interaction between maintenance and planting with study site included as random effect and found that both the maintenance treatments and planting type had a significant effect on survival (P < 0.05 and P < 0.01, respectively). Diagnostic plots and results of the KS test and Levene’s test for homogeneity of variance indicated that assumptions were satisfied and no data transformations were necessary. Post hoc comparison tests revealed that maintained plots had higher survival (mean = 45.5%) than unmaintained plots (mean = 33.7%) and both the 1.2 by 2.4 m (mean = 45.4%) and 2.4 by 2.4 m (mean = 48.3%) plantings had significantly higher rates of survival than the RPM plots (mean = 25.0%; P < 0.05 and P < 0.01, respectively) (Figure 2).
Tree survival by planting treatment. Letters correspond to Tukey’s honestly significant difference (HSD) post hoc comparison significance tests. RPM, Root Production Method®.

We hypothesized that the large-diameter Salix cuttings would fare better than the RPM trees under intense competition from invasives, leading to better long-term survival and growth, which would eventually lead to suppression of the shade-intolerant invasive plant species by shading them out. We did in fact find that the Salix cuttings overall had significantly better survival (45% and 48% in the 1.2 by 2.4 m and 2.4 by 2.4 m plantings, respectively) than the containerized RPM trees (25%). Survival of Salix cuttings was also much better than the 20% overall survival rate reported in a previous study that looked at survival of containerized RPM trees planted in H. japonicus patches (Guyon and Cosgriff Reference Guyon and Cosgriff2022). Also noteworthy was that trees had significantly higher survival rates in plots that were maintained using mowing and herbicide treatments. This indicates that although pursuing planting methods that minimize the need for costly maintenance activities remains worthwhile, implementing some maintenance activities in the early years following plantings continues to have a significant benefit to overall survival rates and therefore planting success.
Hydrology is an extremely important driver of floodplain vegetation community dynamics, and we controlled for differential effects of elevational and related hydrological variation to the best of our ability by selecting sites and plot distributions at the upper (drier) end of documented UMR floodplain inundation zones (Van Appledorn et al. Reference Van Appledorn, Rohweder and De Jager2024). However, an additional extraneous factor that could have influenced survival rates was the fact that Salix cuttings at Reds Landing and Timber Ridge were planted before near-record river flooding that occurred in 2019. This event killed all planted trees at the Pleasant Creek site, which was replanted the following year. Although the majority of trees at Reds Landing and Timber Ridge survived that initial event, it still could have influenced overall mortality, particularly at Reds Landing, which had the lowest overall survival rate and is adjacent to the main channel and open to river flooding. Overstory mortality in the immediate vicinity of the study sites was negligible following the 2019 flood, but other lower-lying areas near the Pleasant Creek site in particular, and the Goose Island site to a lesser degree, were observed to experience high levels of overstory mortality related to that event. As previously noted, the Goose Island site lies in an area generally characterized by relatively sparse overstory cover and long-term colonization by heavy P. arundinacea cover. This is not uncommon in the northern part of the river system, and over larger areas is related to the interacting drivers of hydrology and invasive species resulting in loss of trees and suppressed regeneration (De Jager et al. Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024). However, the relatively higher elevations at our study site places our study plots there well above growing season inundation zone durations where hydrology would be expected to be the main driver of forest loss (approximately 100 growing season days), indicating that P. arundinacea pressure is likely the main driver of forest loss here and that reforestation is an achievable outcome (De Jager et al. Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024; Van Appledorn et al. Reference Van Appledorn, Rohweder and De Jager2024).
Tree Height and Diameter Growth
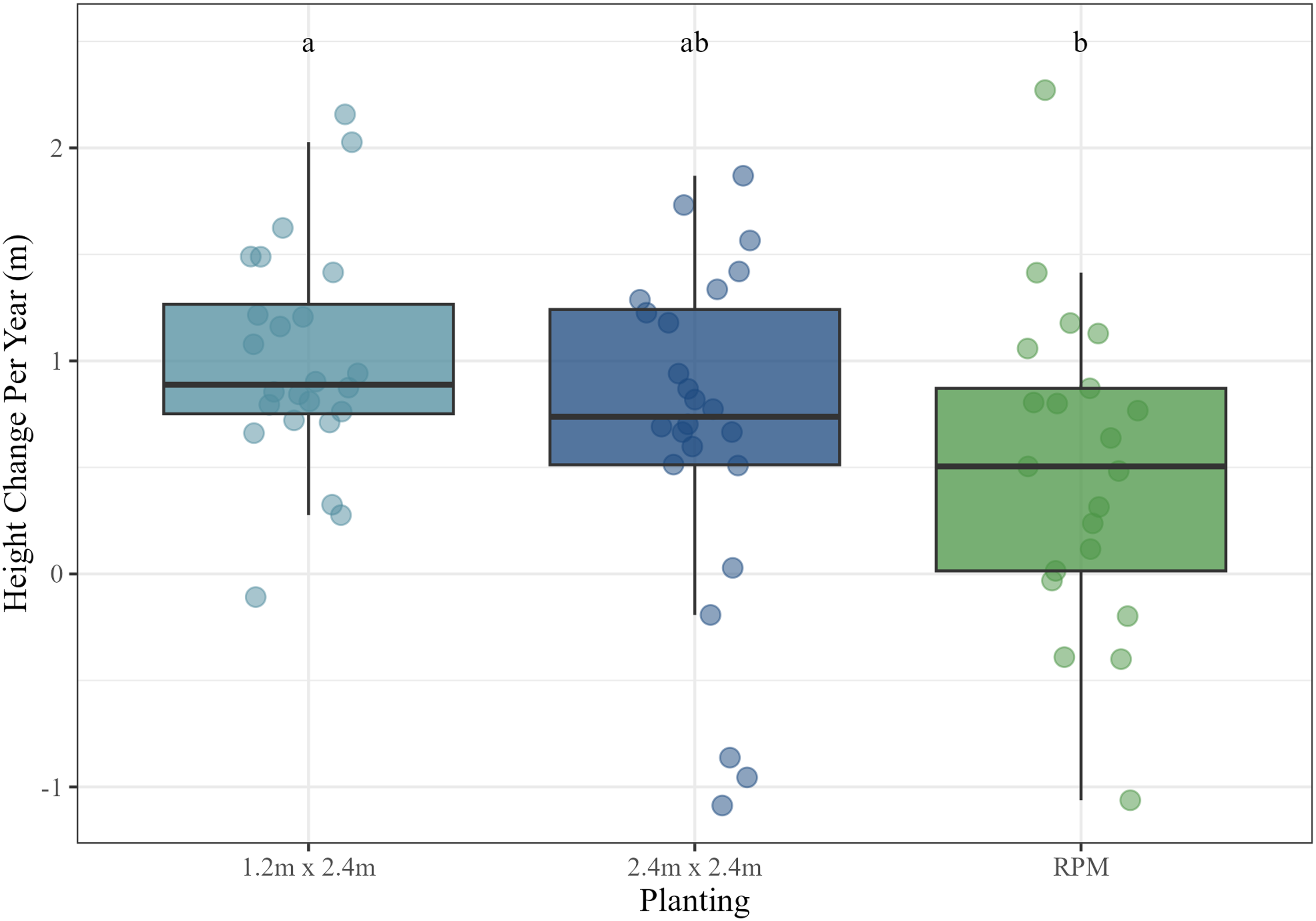
We ran a mixed model of annual height growth by maintenance and planting, as well as their interaction, with site as a random effect. Assumptions of normality and homogeneity of variance were satisfied, and no transformations were required. Results indicated that planting treatment had a significant effect on annual height growth (P < 0.05). The 1.2 by 2.4 m plantings (mean = 1.01 m yr−1) had a significantly higher growth rate than the RPM plantings (mean = 0.53 m yr−1) (Figure 3). For comparison, the 2.4 by 2.4 m plantings had an intermediate mean annual growth rate of 0.68 m yr−1, but no other pairwise comparisons among plantings were significant (Figure 3).
Annual height growth (m) by planting treatment. Letters correspond to Tukey’s Tukey’s honestly significant difference (HSD) post hoc comparison significance tests. RPM, Root Production Method®.

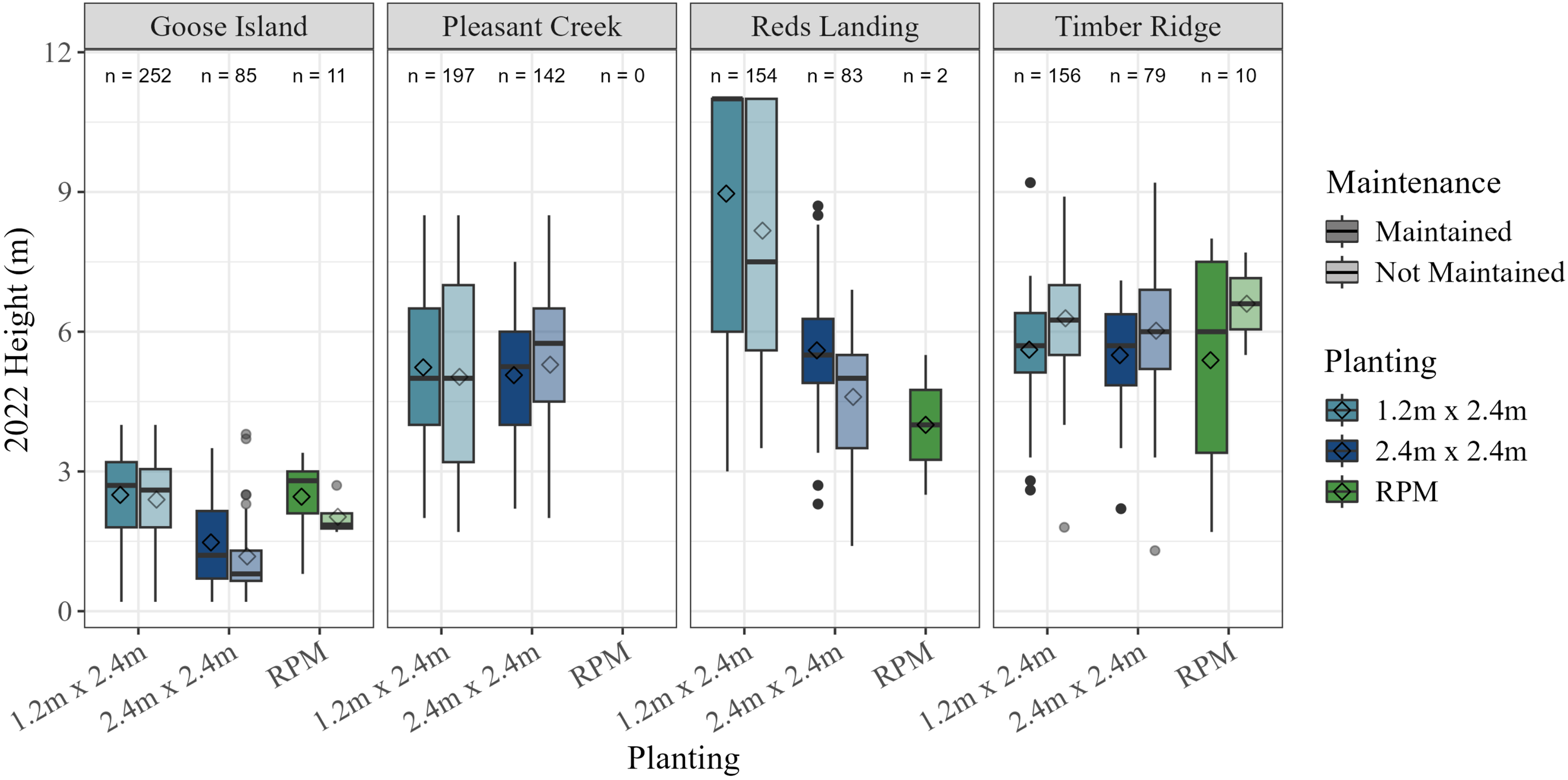
We also summarized cumulative tree heights at the end of the 2022 field season, although we elected not to report statistical tests on this parameter due to the disparity in ages of the planted trees at the different sites, which were all planted at different times (Figure 4; Table 1). Cumulative tree heights were markedly different across sites, although again this was expected. Reds Landing had the tallest trees (mean = 7.5 m), followed by Timber Ridge (mean = 5.9 m), Pleasant Creek (mean = 5.2 m), and Goose Island (mean = 2.2 m) (Figure 4).
Cumulative tree heights (m) by study site, planting treatment, and maintenance treatment. RPM, Root Production Method®. Means are indicated by diamond symbols.

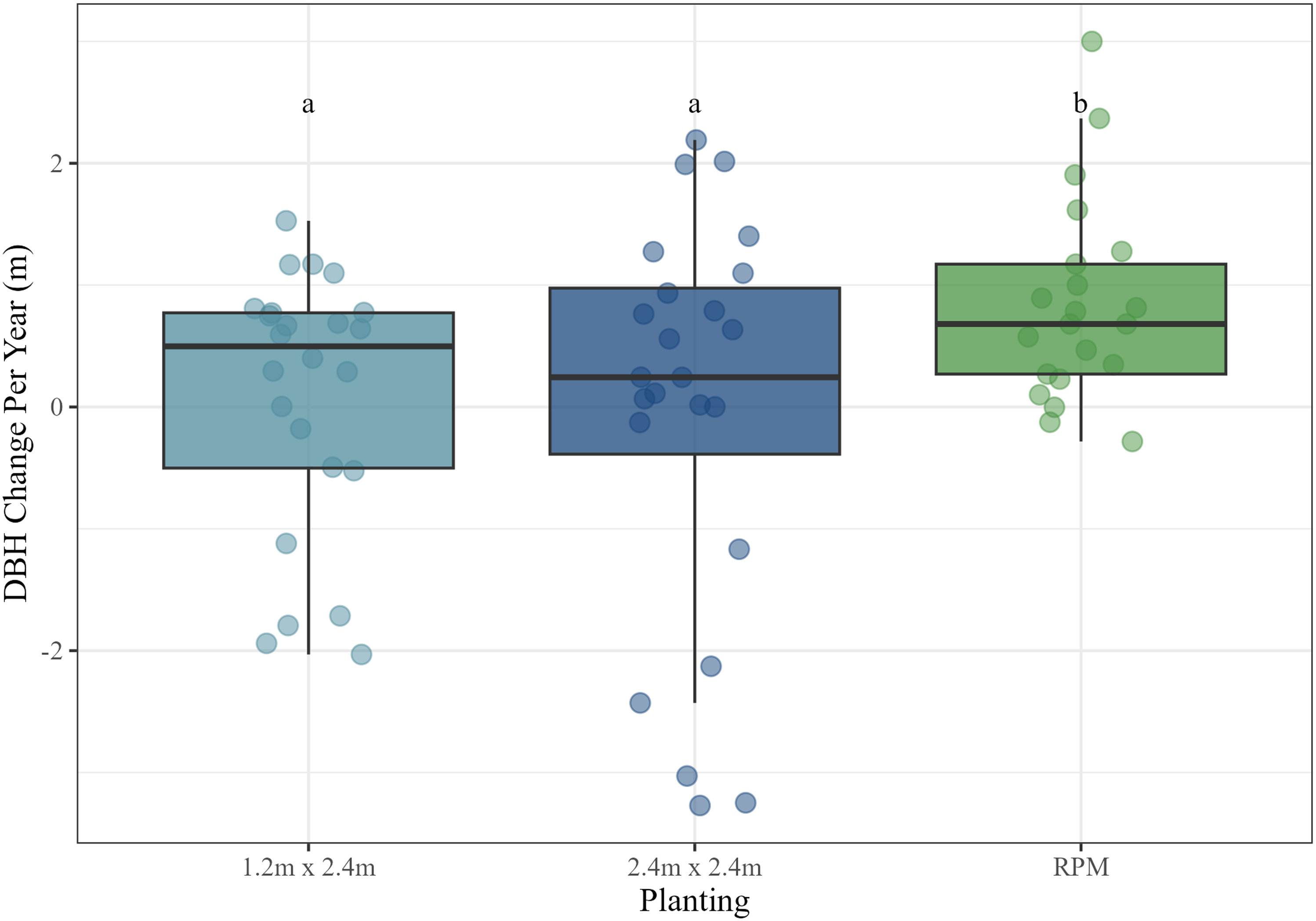
We then ran a mixed model of annual tree diameter (dbh) growth by maintenance treatment, planting treatment, and their interaction, again using site as a random effect. We only examined annual diameter growth rates (and not total or cumulative diameter), given the wide range of initial diameters of Salix cuttings when they were planted. No transformations were required to satisfy assumptions of normally distributed residuals and homogeneity of their variance. Results indicated that maintenance did not have a significant effect on annual diameter growth (P = 0.57), but planting was significant (P < 0.01). Post hoc tests revealed that both 1.2 by 2.4 m (mean = 0.08 cm yr−1) and 2.4 by 2.4 m plantings (mean = −0.04 cm yr−1) had significantly lower annual diameter growth rates than the RPM plantings (mean = 0.92 cm yr−1; P < 0.05 and P < 0.01, respectively) (Figure 5).
Annual diameter growth (cm) by planting treatment. Letters correspond to Tukey’s honestly significant difference (HSD) post hoc comparison significance tests. RPM, Root Production Method®. DBH = diameter at breast height.

Results regarding tree height and diameter growth were nuanced. Salix cuttings had higher annual height growth rates but lower annual diameter growth rates than RPM containerized trees, although annual height growth was only significantly greater in the 1.2 by 2.4 m plantings. The poorer height growth of the 2.4 by 2.4 m plantings was influenced by the performance of those trees at the Goose Island site, where growth was actually negative. This was largely a result of Castor canadensis (North American beaver) activity at that site, which topped a good number of cuttings in several plots about a half meter off the ground. Many of these trees ultimately survived and sprouted new growth, but the effect was to reduce heights and diameters. In contrast to height growth, annual diameter growth of surviving RPM trees was significantly greater than that of the Salix cuttings. Negative growth rates of Salix cuttings at Goose Island again clearly influenced these results, also an effect of the beaver activity mentioned. The relatively large diameter of the Salix cuttings also likely moderated diameter growth by reducing incremental growth rates. However, it is noteworthy that although RPM trees had relatively poor survival compared with Salix cuttings, those that did survive generally maintained competitive growth rates overall. Given that all tree species planted (Salix, P. deltoides, and P. occidentalis) were selected for their rapid potential growth rates, it is not entirely unexpected that their effective maximum growth rates could be somewhat similar. Finally, the overall heights achieved by the majority of Salix cuttings over such a short time period was impressive. Mean cumulative heights greater than 5 m, with many individuals reaching heights greater than 6 to 7 m in three to five growing seasons at the Pleasant Creek, Reds Landing, and Timber Ridge sites is promising. Trees of this height are already likely tall enough to prevent H. japonicus from overtopping them and are therefore also likely able to withstand competitive pressure from that invasive species moving forward.
Change in Invasive Species Cover
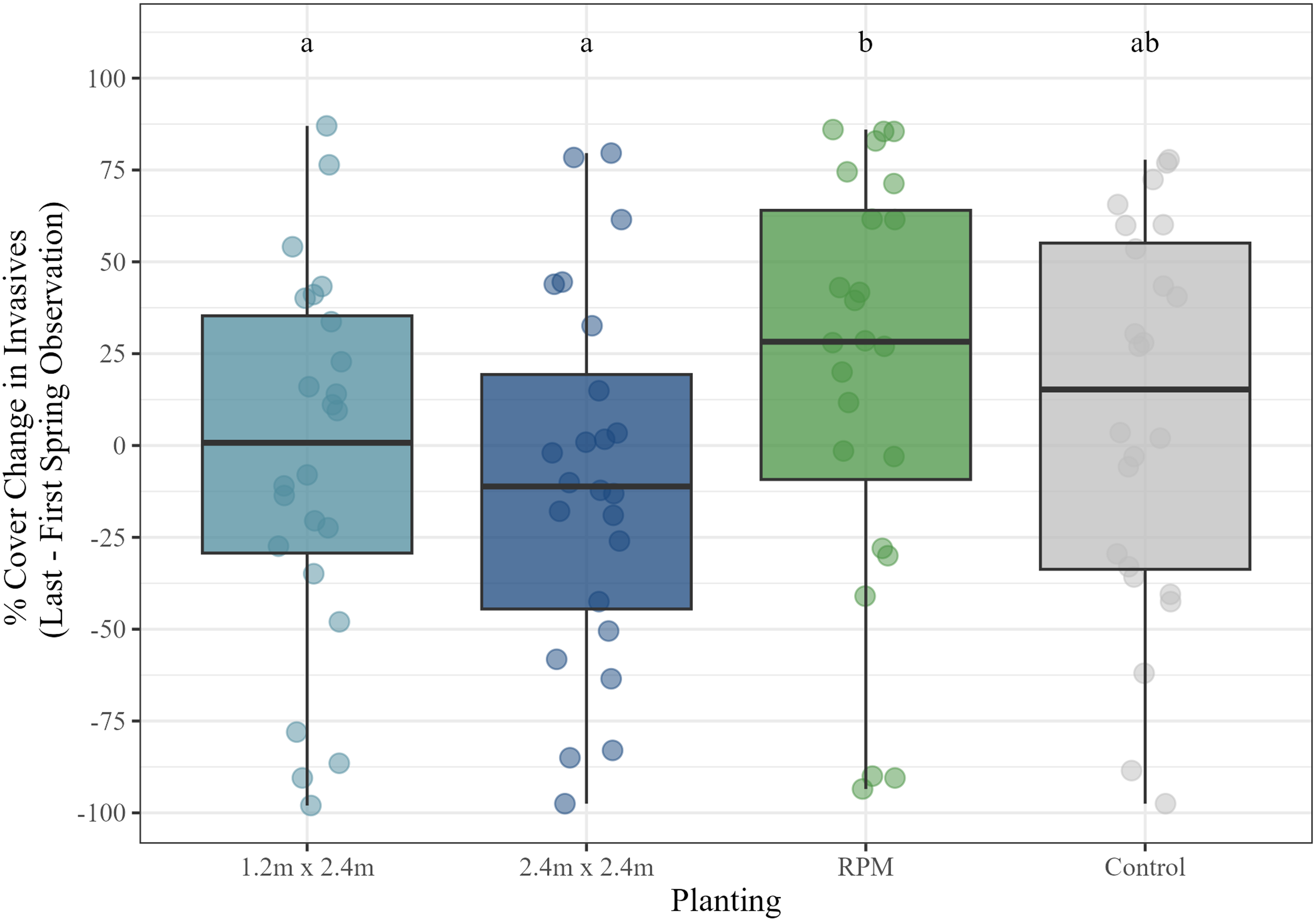
Next, we ran a mixed model of the change (between the first and last spring measurements) in invasive species percent cover by maintenance, planting, and their interaction with site as a random effect. We used data from the spring 2022 (preseason maintenance) sampling period under the assumption that it would be more reflective of baseline vegetation conditions than fall vegetation data, which was more likely to be directly influenced by late-season maintenance activities. Model residuals were normally distributed and did not violate assumptions of homogeneity of variance, and data were left untransformed. Invasive species cover for these series of analyses was limited to the two predominant invasives—H. japonicus and P. arundinacea. Results including both of these species combined indicated that maintenance (P < 0.001) and planting (P < 0.01) both had significant effects on invasive species cover change. The interaction term maintenance:site was not significant (P = 0.97). Invasive species cover declined on average in maintained plots (mean = −10.8%), which was significantly lower than unmaintained plots, which increased in invasives cover (mean = 18.4%). In the plantings, invasives cover declined on average in the 1.2 by 2.4 m (mean = −3.7%) and 2.4 by 2.4 m (mean = −9.1%) plantings, but increased in the RPM plantings (mean = 19.6%) and control group (mean = 8.45%) (Figure 6). The 1.2 by 2.4 m and 2.4 by 2.4 m plantings were both found to be significantly different in invasives cover change than the RPM treatment (P < 0.05 and P < 0.01, respectively). However, the control group was not significantly different from the other planting types (P-values were between 0.1 and 0.5 in these cases).
Invasive species percent cover change by planting treatment during spring observations at the start and end of the study. Letters correspond to Tukey’s honestly significant difference (HSD) post hoc comparison significance tests. RPM, Root Production Method®.

Invasive species cover declined significantly in maintained plots compared with unmaintained plots, indicating that maintenance activities were successful in suppressing invasives over the course of the study. A closer within-site examination of invasives cover indicated that this effect was most pronounced where P. arundinacea was present. We suspect this is likely primarily related to the perennial growth habit of P. arundinacea versus the annual growth habit of H. japonicus, with herbicide treatments potentially having a more lasting effect on the former. Because we used data from the Spring 2022 sampling period, which preceded any annual maintenance activities that year, we are reasonably confident that this reflects a contemporary baseline status of the vegetation communities at the end of the study rather than the more immediate and possibly transitory effects of recent invasive species control actions. By comparison, the difference between invasives cover in maintained and unmaintained plots was much more pronounced at the end of the growing season after annual late-season maintenance activities. Another noteworthy finding is that invasives cover also declined significantly in the Salix plantings compared with the more open RPM plantings (Figure 6), indicating that we may already be starting to see the initial effects of these denser plantings in shading out and suppressing the shade-intolerant invasives. The RPM plantings were generally characterized by more open conditions due to the wider planting spacings used in those plots, which likely also contributed to this effect, but RPM survival was also very low, creating even more open conditions conducive to invasive plant colonization and growth.
Vegetation Community
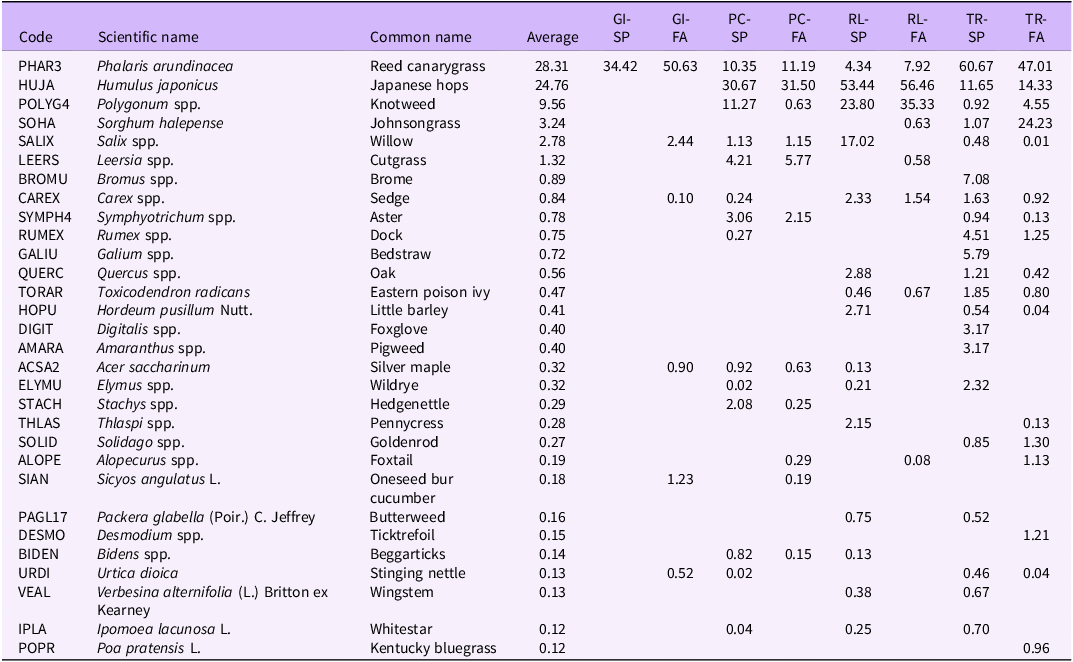
The average percent cover of the top 30 species by site and sampling period is listed in Table 2. The two focal invasive species of this study, P. arundinacea and H. japonicus, were by far the most dominant overall, with average respective percent covers of 28% and 25% in 2022. Of note is that P. arundinacea was so prominent at the Goose Island site that it was the only top 30 species recorded during the spring sampling period that year, and only a handful of additional top 30 species were recorded during the fall sampling period. Knotweed (Polygonum), a native species, was the third most common species overall, followed by S. halepense, another non-native invasive grass species that was most prevalent during the fall sampling period at the Timber Ridge site. Other notable general characteristics of the vegetation community are that it was generally dominated by a relatively small number of species, with most exhibiting very low percent cover overall; there was considerable compositional variability across sites; and seasonal effects were also apparent.
Average percent cover of vegetation by species (top 30), site (GI = Goose Island; PC = Pleasant Creek; RL = Reds Landing; TR = Timber Ridge), and sampling period (SP = spring; FA = fall) in 2022.

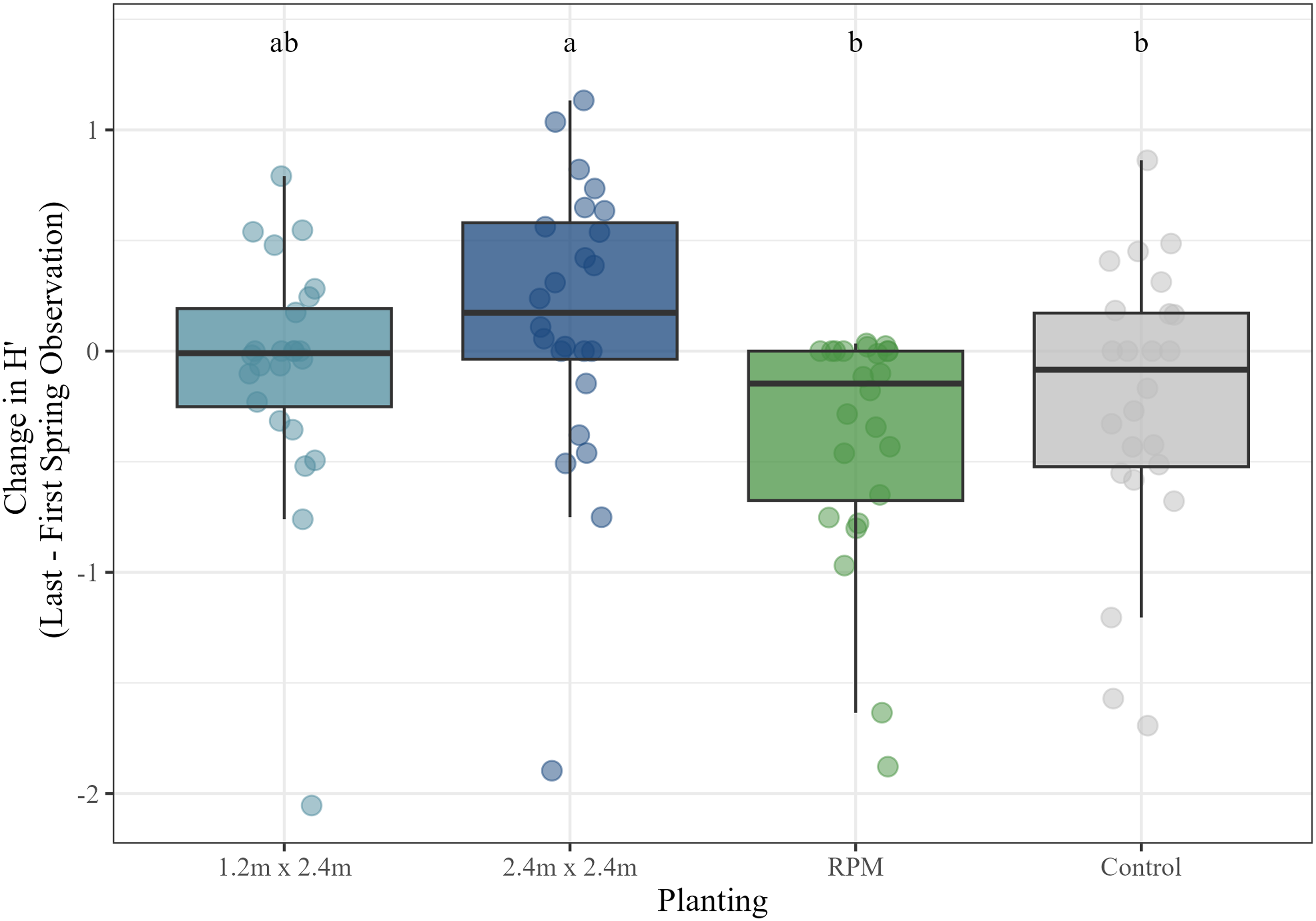
To characterize the overall diversity of the vegetation community, we calculated Shannon (H′) diversity indices for the first and last measurements taken using data from the spring sampling periods and compared the change in diversity across treatments, plantings, and the interaction between treatment and planting using a mixed model with study site included as a random effect. Results indicated that maintenance (P < 0.001) and planting (P < 0.01) had significant effects on diversity. Maintained plots on average increased in diversity (mean H′ change = 0.06), while non-maintained plots declined in diversity (mean H′ change = −0.33). These treatments were significantly different from one another (P < 0.001). Regarding plantings, mean diversity change was greater in the 1.2 by 2.4 m (mean H′ change = −0.08) and 2.4 by 2.4 m (mean H′ change = 0.15) plantings than the RPM (mean H′ change = −0.39) and control plantings (mean H′ change = −0.22), although it should be noted that only the 2.4 by 2.4 m plantings resulted in a mean increase in diversity (Figure 7). Tukey HSD pairwise comparison tests indicated significant differences between the RPM and 2.4 by 2.4 m plantings (P < 0.001), as well as between the control and 2.4 by 2.4 m plantings (P < 0.05).
Change in plant community diversity (H′) by planting treatment during spring observations at the start and end of the study. Letters correspond to Tukey’s honestly significant difference (HSD) post hoc comparison significance tests. RPM, Root Production Method®.

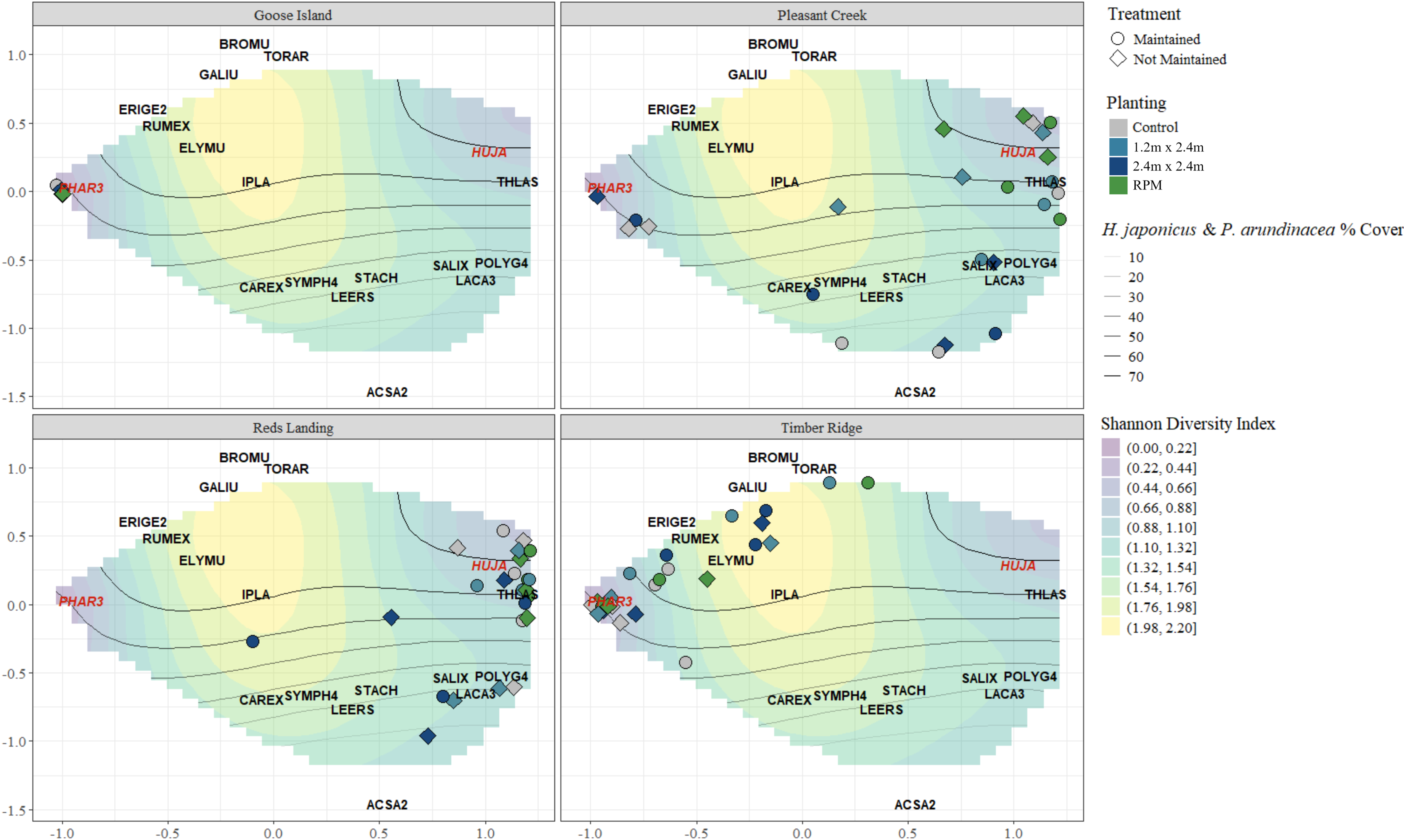
To further examine the broader plant community response to maintenance and tree plantings, we ran an NMDS multivariate analysis using vegetation cover data from the spring 2022 sampling period to generate centroids for maintained and unmaintained plots by planting treatment. After running the analysis and comparing stress values for a range of dimensions, we moved forward with a three-dimensional solution, which converged after 88 iterations and achieved a stress value of 0.06. We graphed the first two dimensions and overlaid species occurrences on the resulting ordination, as well as gradients of invasive species cover (percent cover of H. japonicus and P. arundinacea combined) and plant community diversity (Shannon H′ values) in multivariate space (Figure 8). The x axis in these panels is strongly influenced by the relative dominance of P. arundinacea (PHAR3; negative values) and H. japonicus (HUJA; positive values). Also notable is that the lowest diversity index scores are associated with high P. arundinacea (negative x axis values) and H. japonicus cover (positive x axis values) in this figure. The y axis, in turn, is strongly influenced by combined invasive species percent cover (highest values in the center, lower values toward the positive and negative ends). Goose Island, and to a lesser extent Timber Ridge, are clearly impacted by heavy P. arundinacea cover in this analysis. By contrast, Reds Landing and Pleasant Creek are more strongly impacted by H. japonicus.
Nonmetric multidimensional scaling (NMDS) analyses of the plant community, with vegetation plot attributes on species occurrences, gradients of invasive species cover, and Shannon (H′) diversity. Panels correspond to the four different study sites. RPM, Root Production Method®. See Table 2 for species codes (codes for the invasive species are highlighted in red on the figure). Stress = 0.06; dimensions = 3; iterations = 88.

The change in diversity (H′) of the plant communities followed a pattern similar to the change in invasive species cover. Diversity increased significantly in maintained plots compared with unmaintained plots, in which it actually decreased. Diversity also declined in the RPM and control plots compared with the Salix cutting plots, but this difference was only significant in the 2.4 by 2.4 m plantings, in which diversity increased. If we can safely assume that overall plant diversity is often negatively correlated with invasive species cover due to the effects of suppression (e.g., Guyon and Cosgriff Reference Guyon and Cosgriff2022), then this relationship would be expected based on the lower invasives cover in the Salix cutting plots. Even though the plant communities were generally dominated by relatively few species across the study, with the invasives still at the top of the list (see Table 1), this phenomenon was even more striking at the Goose Island site, where P. arundinacea has established a near monoculture and overall diversity was extremely low (H′ = 0.08). Other studies have also noted that the UMR floodplain vegetation community is generally dominated by relatively few species, with most exhibiting low percent cover, and has considerable compositional variability across sites (Guyon and Battaglia Reference Guyon and Battaglia2018; Guyon and Cosgriff Reference Guyon and Cosgriff2022). For comparison, the most common UMR ground-layer vegetation species previously recorded at a series of reference sites not significantly impacted by invasives include Canadian woodnettle [Laportea canadensis (L.) Weddell], false nettle [Boehmeria cylindrica (L.) Sw.], stinging nettle (Urtica dioica L.), Virginia wildrye (Elymus virginicus L.), eastern poison ivy [Toxicodendron radicans (L.) Kuntze], calico aster [Symphyotrichum lateriflorum (L.) Á. Löve & D. Löve], and whitegrass (Leersia virginica Willd.) (Guyon and Battaglia Reference Guyon and Battaglia2018; LJ Guyon and LL Battaglia unpublished data). Shannon (H′) diversity indices of ground-layer vegetation at these sites ranged from 0.81 to 3.46 (Guyon and Battaglia Reference Guyon and Battaglia2018; LJ Guyon and LL Battaglia, unpublished data).
The inverse correlation between invasives cover and the diversity of the overall plant community is also apparent from the NMDS analysis (Figure 8), as is the heavy influence of the invasives on the positioning of study plots in multivariate space. Ground-layer vegetation is notably variable across the UMR floodplain (e.g., Guyon and Battaglia Reference Guyon and Battaglia2018), and we feel that the NMDS analysis was able to capture this variability while simultaneously providing insight into underlying trends in vegetation community dynamics.
In conclusion, forest health and overall forest cover have declined in the UMR due to the cumulative effects of altered hydrology, flood disturbances, and ever-increasing competitive pressure from invasive species (De Jager et al. Reference De Jager, Rohweder, Van Appledorn, Hlavacek and Meier2024; Guyon and Weiss Reference Guyon and Weiss2023). Without forest management actions, these trends are likely to continue unabated into the future, compounded even further by the impending loss of F. pennsylvanica from the system due to EAB (De Jager et al. Reference De Jager, Van Appledorn, Fox, Rohweder, Guyon, Meier, Cosgriff and Vandermyde2019). Reforesting areas that have been heavily colonized by invasive species presents even more difficulties than planting trees under favorable conditions in the UMR, which still often meets with limited success. The results of this experimental tree planting indicate that large-diameter Salix cuttings planted at high densities had significantly better survival than containerized trees in areas heavily colonized by invasive species. We found few differences between the 1.2 by 2.4 m and the 2.4 by 2.4 m planting densities, so we recommend the 2.4 by 2.4 m planting density in most cases to save costs. Height growth of the Salix cuttings was also impressive, with most cuttings achieving mean annual height growth of about a meter per year, reaching mean cumulative heights of more than 5 m within 3 to 5 yr at most sites. Also noteworthy was that invasives cover was significantly lower and ground-layer vegetation diversity was higher in the Salix plantings than in the containerized plantings. Salix is a good fit for reforesting lower-lying flood-prone areas, given its high tolerance for flooded conditions compared with most other UMR floodplain tree species, but sourcing and transporting cuttings may be a concern in locations without viable stock nearby. Given the significant effects of maintenance activities, we would also recommend maintaining plantings with a combination of mowing and herbicide application two times per year for at least a few years following planting. Damage due to beaver and other wildlife can also be significant, but using tree-planting guards can minimize that to some degree. We believe that an overarching goal of forest restoration activities in the UMR should be to stop and hopefully reverse the trend toward increasing loss of forest cover, health, and diversity—increasing the availability of a diverse set of viable reforestation methods such as those implemented in this study is an important component of that effort.
Funding statement
Funding for this work was provided by the Long-Term Resource Monitoring element of the Upper Mississippi River Restoration Program. AT was partially supported by NSF REU Site award “Wetland Science in the Modern World” (DEB 2050400).
Competing interests
The authors declare no conflicts of interest.

 Open access
Open access