


Introduction
Global agriculture faces increasing challenges driven by climate change, soil degradation, water scarcity and the growing demand for food, which together threaten crop productivity and sustainability (Roussin-Léveillée et al., Reference Roussin-Léveillée, Rossi, Castroverde and Moffett2024; Yang et al., Reference Yang, Zhang, Zhang, Wei, Zhao, Zheng and Xu2024; Ghosh et al., Reference Ghosh, Kumar and Biswas2024a). Seed germination is a critical developmental transition that determines seedling establishment and crop performance and is tightly regulated by coordinated physiological and biochemical processes occurring during the triphasic imbibition process. Seed priming has emerged as an efficient and environmentally friendly strategy to enhance germination, seedling vigour and early growth under such constraints (Fu et al., Reference Fu, Ma, Li, Hou, Zeng, Zhang and Luo2024). Traditional priming approaches, including hydropriming, osmopriming and biopriming, operate by partially activating pre-germinative metabolism, allowing seeds to initiate enzymatic activity, RNA synthesis and membrane reorganization without radicle protrusion (Song and He, Reference Song and He2021; Fu et al., Reference Fu, Ma, Li, Hou, Zeng, Zhang and Luo2024). In this context, germination is considered the process ending with radicle emergence, whereas subsequent processes involving reserve mobilization and sustained growth correspond to post-germinative seedling establishment.
In recent years, the integration of nanotechnology into seed priming has given rise to nanopriming, in which engineered or green-synthesized nanoparticles are used to modulate water uptake, redox balance and metabolic activation during the early phases of seed imbibition and metabolic reactivation (Faizan et al., Reference Faizan, Sharma, Sultan, Alam, Sehar, Rajput and Hayat2024; Ghosh et al., Reference Ghosh, Dey, Guha, Paul and Kundu2024b; Adzuan et al., Reference Adzuan, Awal Sembada, Osman, Abu Bakar, So’aib and Lenggoro2025; Durgadevi et al., Reference Durgadevi, Girigoswami and Girigoswami2025). Compared with conventional priming, nanopriming enables more precise control of physiological responses by tuning nanoparticle size, surface chemistry and dose (Cartwright et al., Reference Cartwright, Jackson, Morgan, Anderson and Britt2020; Khalaki et al., Reference Khalaki, Moameri, Lajayer and Astatkie2021; Song and He, Reference Song and He2021).
Among the different nanomaterials tested, silver nanoparticles (AgNPs) have received particular attention due to their strong redox activity, antimicrobial properties and ability to modulate plant signalling pathways (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020; Faizan et al., Reference Faizan, Sharma, Sultan, Alam, Sehar, Rajput and Hayat2024; Yang et al., Reference Yang, Zhang, Zhang, Wei, Zhao, Zheng and Xu2024). Numerous studies have demonstrated that AgNP seed priming can accelerate germination, improve seedling vigour and enhance tolerance to salinity, drought and pathogens in crops such as rice, wheat, maize, lettuce and sunflower (Song et al., Reference Song, Phenrat, Marinakos, Xiao, Liu, Wiesner and Lowry2011; Syu et al., Reference Syu, Hung, Chen and Chuang2014; Almutairi, Reference Almutairi2016; Ouyang and Sun, Reference Ouyang and Sun2016; Khan et al., Reference Khan, Raza, Awan, Shah, Rizwan, Ali and Huang2020; Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020; Sharma et al., Reference Sharma, Sidhu, Manchanda and Ahuja2022; Thongmak et al., Reference Thongmak, Ruangwong, Wongkaew, Srisonphan and Onwimol2022; Yan et al., Reference Yan, Chen, Pan, Zhao, Rui and Zhao2022; Cembrowska-Lech and Rybak, Reference Cembrowska-Lech and Rybak2023; Lopes et al., Reference Lopes, de Oliveira Silva and Courrol2023, Reference Lopes, Lopes and Courrol2024; Antunes et al., Reference Antunes, Forini, Coqueiro, Pontes, Lima, Cavalcante and Grillo2024; Kim et al., Reference Kim, Kim, Sung, Koduru, Nile, Syed and Ghodake2024; Krishnasamy et al., Reference Krishnasamy, Natesh and Obbineni2024; Lopes and Courrol, Reference Lopes and Courrol2024; Elkelish et al., Reference Elkelish, Alqudah, Alammari, Alsubeie, Hamed and Thabet2025; Handayani et al., Reference Handayani, Tanadi and Umar2025; Sahoo et al., Reference Sahoo, Sahu and Behera2025). However, these beneficial effects coexist with a narrow toxicity threshold: excessive AgNP concentrations or inappropriate surface chemistries can lead to oxidative damage, mitochondrial dysfunction and growth inhibition (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
Mechanistically, AgNPs do not act as nutrients but rather as redox-active modulators. At optimal doses, they generate controlled reactive oxygen species (ROS) through a combination of nanoparticle-specific interactions and regulated Ag⁺ release, which function as signalling molecules, activating antioxidant enzymes, mitogen-activated protein kinase (MAPK) cascades and hormonal pathways that promote metabolic activation and cellular reorganization during germination (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). When this redox signalling exceeds the buffering capacity of thiol-based systems, particularly glutathione, oxidative stress dominates and germination and growth are inhibited (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
Importantly, the response to AgNP priming is highly species-dependent, being governed by seed coat permeability, imbibition dynamics and intrinsic redox capacity (Mohamed et al., Reference Mohamed, Qayyum, Abdel-Hadi, Rehman, Ali and Rizwan2017; Chen et al., Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023; Arridho et al., Reference Arridho, Ilyas, Damayanti, Widajati and Qadir2025; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). Likewise, nanoparticle properties such as size, shape, surface charge, coating and Ag⁺ release kinetics critically determine the balance between beneficial signalling and toxicity (Liu and Hurt, Reference Liu and Hurt2010; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Savassa et al., Reference Savassa, Castillo-Michel, Pradas Del Real, Reyes-Herrera, Marques and de Carvalho2021; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023). Green-synthesized AgNPs, which carry organic capping agents and exhibit slower ion release, are, therefore, emerging as safer and more predictable platforms for seed priming (Savassa et al., Reference Savassa, Castillo-Michel, Pradas Del Real, Reyes-Herrera, Marques and de Carvalho2021; Das and Dutta, Reference Das and Dutta2022; Labeeb et al., Reference Labeeb, Badr, Haroun, Mattar and El-Kholy2022).
Rather than viewing AgNP seed priming as a purely empirical treatment, the evidence reviewed here supports a mechanistically guided framework in which redox homeostasis, glutathione buffering capacity and MAPK-dependent signalling act as central regulators of the priming response. In this view, AgNPs do not simply function as antimicrobial or stress-inducing agents but as modulators of the intrinsic redox–metabolic network of the seed, defining a narrow hormetic window in which germination and early seedling establishment are enhanced. This redox-guided perspective provides a coherent explanation for the wide variability in AgNP priming outcomes reported across species, doses and experimental systems.
Redox-based mechanisms of AgNP seed priming
Dose-dependent redox signalling and oxidative thresholds
AgNPs act as redox-active modulators rather than as nutrient sources. Their biological activity is primarily driven by the controlled release of Ag⁺ ions and the associated generation of ROS at the seed surface and within internal tissues (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). Although part of this activity derives from dissolved Ag⁺, nanoparticle-specific effects such as interactions with the seed coat and localized ROS generation may also contribute to the observed physiological responses. At low to moderate concentrations, this redox activity induces a transient oxidative signal that functions as a developmental cue, activating early metabolic processes associated with germination and accelerating radicle emergence (Fig. 1) (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). These effects arise from the combined contribution of nanoparticle-specific interactions and the controlled release of Ag⁺ ions, which together determine the resulting redox signalling.
Dose-dependent effects of silver nanoparticles (AgNPs) on seed germination. Low AgNP doses promote beneficial redox signalling and antioxidant activation, resulting in improved germination and seedling vigour, whereas high doses induce oxidative stress, lipid peroxidation and cellular damage, leading to growth inhibition. The curve illustrates the non-linear (hormetic) response of germination to increasing AgNP concentration.

Figure 1 Long description
The illustration shows two sections of a seed, each depicting the effects of silver nanoparticles (AgNPs) at different doses. On the left, labeled 'Low AgNP dose', AgNPs are shown entering the seed, leading to 'ROS signaling' and 'Antioxidant activation' with enzymes SOD, CAT and APX, resulting in 'Improved germination and vigor'. On the right, labeled 'High AgNP dose', AgNPs cause 'Oxidative stress', 'Lipid peroxidation (MDA)' and 'Cellular damage', leading to 'Growth inhibition'. Above, a graph illustrates the germination rate against AgNP concentration, indicating an optimal dose for germination. The diagram highlights the contrasting effects of AgNPs on seed germination and growth depending on the concentration used.
As summarized in Table 1, during the imbibition phase, AgNPs modulate water uptake and membrane reorganization, partly by altering seed coat permeability and aquaporin activity (Mohamed et al., Reference Mohamed, Qayyum, Abdel-Hadi, Rehman, Ali and Rizwan2017; Chen et al., Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). This early redox signal stimulates antioxidant and signalling pathways, including superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX) and MAPK cascades, allowing ROS to be maintained within a signalling window rather than reaching damaging levels (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
Physiological and molecular effects of silver nanoparticle (AgNP) priming during seed germination and early seedling development

Table 1 Long description
The table compares physiological and molecular effects during seed germination and early seedling development with and without silver nanoparticle priming. Key findings include enhanced water uptake and increased seed coat permeability during imbibition with AgNP priming. Controlled ROS generation and antioxidant activation occur during the lag phase, improving metabolic processes. Radicle emergence is accelerated due to enhanced metabolism and hormone signaling. Seedling establishment shows improved stress tolerance and synchronized growth, while post-germination growth benefits from persistent stress memory and enhanced resilience. These effects suggest that AgNP priming positively influences seed germination and development by modulating key physiological and molecular processes.
Notes:
a Trzcińska-Wencel et al. (Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025); Mohamed et al. (Reference Mohamed, Qayyum, Abdel-Hadi, Rehman, Ali and Rizwan2017); Chen et al. (Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023).
b Trzcińska-Wencel et al. (Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025); Abou-Zeid and Moustafa (Reference Abou-Zeid and Moustafa2014); Abdel-Aziz and Rizwan (Reference Abdel-Aziz and Rizwan2019); Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023).
c Prażak et al. (Reference Prażak, Święciło, Krzepiłko, Michałek and Arczewska2020); Abou-Zeid and Moustafa (Reference Abou-Zeid and Moustafa2014).
d Kumar et al. (Reference Kumar, Muthukrishnan and Rajalakshmi2020); Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023); Trzcińska-Wencel et al. (Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
e Chandrasekaran et al. (Reference Chandrasekaran, Luo, Wang and Shu2020); Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023); Trzcińska-Wencel et al. (Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
However, this beneficial effect operates within a narrow dose range. When AgNP concentration exceeds the buffering capacity of cellular thiols and antioxidant systems, ROS accumulate uncontrollably, leading to lipid peroxidation, protein oxidation and mitochondrial dysfunction (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). In addition, Ag⁺ ions can directly interact with thiol (–SH) groups in glutathione and cysteine residues of proteins, potentially disrupting enzyme activity and redox homeostasis. In this context, direct binding of Ag⁺ to thiol groups in proteins may represent an additional mechanism of toxicity, independent of ROS accumulation.
This transition from redox signalling to oxidative stress is reflected by increased malondialdehyde (MDA) levels and suppressed radicle elongation, resulting in growth inhibition (Fig. 2) (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). The existence of this dose–response window explains the frequent observation that low AgNP doses promote germination and vigour, whereas higher doses are phytotoxic.
Dose-dependent redox–MAPK–glutathione model during AgNP seed priming. Low AgNP doses promote MAPK activation, NADPH-dependent glutathione reduction and balanced redox homeostasis, supporting metabolic activity and early seedling growth. High AgNP doses lead to excessive ROS accumulation, glutathione oxidation and mitochondrial dysfunction, resulting in oxidative damage and growth inhibition. The central panel summarizes the core redox–metabolic network linking MAPK signalling, carbon metabolism and the GSH/GSSG system.

Figure 2 Long description
The diagram illustrates the effects of silver nanoparticles (AgNPs) exposure on the redox-MAPK-glutathione hub and the resulting outcomes. On the left, AgNPs are shown entering a cell, leading to reactive oxygen species (ROS) generation. The central pathway includes ROS, MAPK, NADPH and the conversion between GSH and GSSG. On the right, outcomes are divided into adaptive response and oxidative imbalance. Adaptive response includes antioxidant activation, stress tolerance and seedling establishment. Oxidative imbalance includes excess ROS, lipid peroxidation (MDA) and growth inhibition.
Although the concept of a narrow hormetic window is consistently reported, quantitative ranges can be approximated from experimental studies. Beneficial effects of AgNP seed priming are most frequently observed at low concentrations, typically below 10–50 mg L−1, depending on species and nanoparticle properties (Prażak et al., Reference Prażak, Święciło, Krzepiłko, Michałek and Arczewska2020; Savassa et al., Reference Savassa, Castillo-Michel, Pradas Del Real, Reyes-Herrera, Marques and de Carvalho2021; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023). In contrast, concentrations above 50–100 mg L−1 are commonly associated with the onset of oxidative stress, growth inhibition and metabolic disruption (Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020). This biphasic response is consistent with a hormetic dose–response pattern widely described for nanomaterials, where low doses promote redox signalling and metabolic activation, while higher doses overwhelm antioxidant defences and induce toxicity (Agathokleous et al., Reference Agathokleous, Feng, Iavicoli and Calabrese2019). Importantly, the threshold between stimulation and inhibition is highly dependent on plant species, nanoparticle size and physicochemical properties, indicating that no universal cut-off can be defined but rather a system-specific redox window.
Redox–MAPK–glutathione coupling as a central regulatory node
The cellular response to AgNP priming is orchestrated through the tight coupling between ROS, MAPK signalling and the glutathione redox system (Fig. 2) (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). Low levels of AgNP-derived ROS activate MAPK cascades, including MAPKs such as MPK3 and MPK6, which, in turn, regulate the expression of antioxidant enzymes, stress-responsive genes and hormone-related pathways (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023). These MAPK-mediated responses illustrate how moderate ROS signals generated during nanopriming are transduced into coordinated transcriptional and metabolic adjustments that support processes associated with seed germination. This signalling network integrates redox cues with transcriptional and metabolic reprogramming required for germination.
Glutathione (GSH) plays a pivotal role in maintaining this balance. The ratio between reduced and oxidized glutathione (GSH/GSSG) determines whether ROS function as signals or as toxic agents (Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020). At optimal AgNP doses, NADPH-dependent glutathione reductase sustains a high GSH/GSSG ratio, enabling efficient scavenging of excess ROS while preserving redox signalling (Fig. 3) (Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023). In addition to ROS-mediated signalling, silver ions (Ag⁺) released from AgNPs may interact directly with thiol (–SH) groups present in glutathione and other cysteine-containing biomolecules. Silver ions are well-known thiol-reactive species that form stable Ag–S complexes with reduced sulphur groups, potentially altering the cellular redox buffering capacity and modulating oxidative stress signalling pathways (Liu et al., Reference Liu, Sonshine, Shervani and Hurt2010; Wilkinson et al., Reference Wilkinson, White and Chipman2011; Marchioni et al., Reference Marchioni, Gallon, Worms, Jouneau, Lebrun, Veronesi, Truffier-Boutry, Mintz, Delangle, Deniaud and Michaud-Soret2018). This mechanism may contribute to toxicity independently of ROS overproduction, highlighting an additional non-redox pathway primarily associated with Ag⁺-induced cellular disruption.
Effect of seed coat thickness on the penetration and distribution of silver nanoparticles (AgNPs) during seed priming. In seeds with thin or highly permeable seed coats, AgNPs and released Ag+ readily cross the testa and reach the endosperm and embryo. In contrast, thick or lignified seed coats restrict nanoparticle penetration, resulting in the accumulation of AgNPs mainly in the outer layers and reduced delivery of Ag+ to internal tissues.

Figure 3 Long description
The diagram consists of three sections: AgNP exposure, metabolic regulation and outcomes. The left section shows a seed with AgNPs labeled, indicating controlled ROS signaling. The middle section illustrates metabolic regulation, including ROS, central carbon metabolism, glycolysis, TCA cycle and ATP production. NADH and FADH subscript 2 are noted for electron transport and ATP synthesis. The right section lists outcomes: adaptive metabolic activation with increased ATP production, enhanced carbohydrate and lipid metabolism and improved seedling vigor. It also describes metabolic disruption at high AgNP doses, leading to excess ROS accumulation, mitochondrial dysfunction and reduced ATP production.
When AgNP-induced ROS production overwhelms this system, glutathione becomes oxidized, MAPK signalling is disrupted and oxidative damage accumulates, leading to impaired germination and seedling growth (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Ma et al., Reference Ma, Liu, Chen, Zhao, Guo, Minocha and Dhankher2020; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). Thus, the balance between ROS production, MAPK signalling and glutathione redox buffering defines a central regulatory module determining whether AgNP exposure promotes adaptive signalling or triggers oxidative toxicity.
Distinguishing nanoparticle-specific and ion-mediated effects
In addition to dose-dependent redox effects and intracellular signalling, an essential aspect of AgNP-mediated seed priming lies in distinguishing nanoparticle-specific processes from ion-mediated mechanisms. In most biological systems, AgNPs act as dynamic reservoirs of silver ions (Ag⁺), and a substantial fraction of their physiological impact is mediated by ion release (Liu et al., Reference Liu, Sonshine, Shervani and Hurt2010). However, increasing evidence indicates that AgNPs and Ag⁺ differ fundamentally in their uptake, transport and biological effects.
Silver ions (Ag⁺) can rapidly diffuse into seed tissues and interact with cellular components without physical constraints, whereas nanoparticle uptake is size-dependent and limited by structural barriers such as cell wall pores and apoplastic restrictions. Studies on plant–nanoparticle interactions indicate that uptake, translocation and accumulation depend strongly on particle size, surface properties and plant species (Rico et al., Reference Rico, Majumdar, Duarte-Gardea, Peralta-Videa and Gardea-Torresdey2011). Furthermore, AgNP exposure often results in higher accumulation levels than equivalent ionic treatments, supporting the existence of particle-specific transport pathways (Geisler-Lee et al., Reference Geisler-Lee, Wang, Yao, Zhang, Geisler, Li and Ma2013).
At the molecular level, Ag⁺ readily interacts with thiol-containing molecules, including glutathione and cysteine residues in proteins, leading to disruptions in redox homeostasis, enzyme activity and membrane integrity. These interactions are closely associated with the generation of ROS and the activation of antioxidant systems and stress-related signalling pathways. While many of these responses are shared between AgNP and Ag⁺ treatments, comparative studies demonstrate that only a fraction of gene expression changes overlap, indicating the coexistence of ion-mediated and nanoparticle-specific signalling mechanisms (Kaveh et al., Reference Kaveh, Li, Ranjbar, Tehrani, Brueck and Van Aken2013).
However, AgNPs cannot be considered merely passive sources of ions. Their particulate nature introduces additional mechanisms that are not replicated by dissolved silver alone. These include direct interactions with biological interfaces, localized surface reactivity and the formation of spatially heterogeneous redox microenvironments. Experimental evidence shows that nanoparticle exposure can induce distinct proteomic and cellular responses compared to ionic silver, including effects on intracellular compartments and membrane-associated processes (Vannini et al., Reference Vannini, Domingo, Onelli, Prinsi, Marsoni, Espen and Bracale2013). Moreover, inhibition of early growth processes and tissue damage have been observed under AgNP exposure even when equivalent Ag⁺ treatments produce weaker or different effects, further supporting nanoparticle-specific toxicity pathways (Yin et al., Reference Yin, Cheng, Espinasse, Colman, Auffan, Wiesner, Rose, Liu and Bernhardt2011).
Importantly, nanoparticle-specific and ion-mediated effects are tightly coupled and cannot be fully disentangled under physiological conditions. AgNPs undergo partial dissolution during imbibition while simultaneously maintaining particle-specific interactions with seed surfaces and internal tissues. As a result, the biological response to AgNP priming emerges from the interplay between controlled Ag⁺ release and nanoparticle-driven processes such as size-dependent uptake, surface reactivity and localized signalling. As a consequence, similar nominal silver concentrations may lead to markedly different physiological outcomes depending on nanoparticle formulation, dissolution behaviour and seed-specific interface processes.
Within this framework, AgNPs function not only as sources of reactive silver species but also as modulators of their spatial and temporal delivery, defining the boundary between beneficial redox signalling and oxidative toxicity. This distinction is particularly relevant when considering seed coat-mediated bioavailability and the establishment of effective dose windows during priming.
Redox priming, signal transduction and establishment of stress memory
Once the origin and nature of AgNP-derived redox signals are characterized, their downstream integration facilitates the establishment of a persistent stress memory in primed seeds. Within this mechanistic framework, AgNP-mediated seed priming is conceptualized as a biphasic process: an initial triggering phase (characterized by transient redox signalling) followed by an adaptive phase where a sustained physiological state is maintained (Chen and Arora, Reference Chen and Arora2013; Paparella et al., Reference Paparella, Araújo, Rossi, Wijayasinghe, Carbonera and Balestrazzi2015).
During seed imbibition, AgNPs interact with the seed coat and penetrate internal tissues, modulating the biochemical ontogeny of germination. A pivotal mechanism in this interaction is the calibrated generation of ROS, specifically hydrogen peroxide (H2O2), which functions as a key signalling cue (Bailly, Reference Bailly2019). Controlled ROS accumulation triggers metabolic shifts that enhance enzymatic activity and initiate reserve mobilization processes contributing to embryo growth, leading to radicle emergence, while simultaneously upregulating antioxidant defence hubs, including CAT and various peroxidases (Mahakham et al., Reference Mahakham, Sarmah, Maensiri and Theerakulpisut2017). This ‘oxidative window’ ensures that the transient burst operates within a homeostatic range, acting as a developmental trigger rather than a cytotoxic agent (Méndez-Andrade et al., Reference Méndez-Andrade, Ruiz-Torres, Wang, García-Cerda and Vera-Reyes2024).
This early redox signal is mechanistically coupled to the redox–MAPK–glutathione regulatory node. ROS serve as secondary messengers that activate MAPK cascades and redox-sensitive transcription factors, effectively bridging oxidative stimuli with transcriptional reprogramming (Colcombet and Hirt, Reference Colcombet and Hirt2008). Consequently, stress-responsive genes enter a ‘primed’ state, allowing for rapid recruitment upon subsequent challenges. This state is defined by an enhanced sensitivity to stimuli rather than the metabolic cost of constitutive defence activation (Martinez-Medina et al., Reference Martinez-Medina, Flors, Heil, Mauch-Mani, Pieterse, Pozo, Ton, van Dam and Conrath2016).
The implications of this signalling phase extend into the early metabolic reprogramming of the seed. Redox-mediated pathways stimulate central carbon metabolism and mitochondrial bioenergetics, providing the ATP and reducing power (NADPH) necessary for embryo growth and the transition from quiescence to radicle protrusion (Lahuta et al., Reference Lahuta, Szablińska-Piernik, Głowacka, Stałanowska, Railean-Plugaru, Horbowicz, Pomastowski and Buszewski2022). Concurrently, the robust antioxidant buffering prevents oxidative drift, maintaining the ‘signalling tone’ required for persistent physiological modification.
The longevity of this stress memory relies on integrated redox-regulated mechanisms. Primed seeds exhibit a superior metabolic preparedness, characterized by the accumulation of protective osmolytes and secondary metabolites (Khan et al., Reference Khan, Zhang, Luo, Liu, Rizwan, Fahad, Xu and Hu2019). Furthermore, emerging evidence suggests that redox signalling may induce long-term regulatory adjustments through epigenetic modifications, such as DNA methylation or histone acetylation, although the specific epigenetic footprint of AgNP-priming remains an active area of investigation (Stałanowska et al., Reference Stałanowska, Głowacka, Buszewski and Lahuta2025).
These processes enable primed seeds to retain a functional ‘memory’, resulting in a reduced response latency and optimized redox homeostasis when facing environmental constraints. Unlike non-primed seeds, which must initiate de novo stress perception, AgNP-primed individuals demonstrate superior seedling establishment and enhanced tolerance to abiotic stressors.
Metabolic reprogramming during AgNP-mediated priming
AgNP seed priming has been shown to modify amino acid pools, phenolic and flavonoid content, soluble sugars, osmolytes and antioxidant metabolites, reflecting coordinated metabolic reprogramming initiated during germination and further developed during early seedling establishment (Pintos et al., Reference Pintos, de Diego and Gomez-Garay2024).
At the biochemical level, successful seed germination depends on the timely mobilization of stored reserves. Experimental evidence supports the role of AgNP priming in accelerating these processes. For instance, AgNP nanopriming has been shown to enhance α-amylase activity, leading to increased soluble sugar availability during germination (Mahakham et al., Reference Mahakham, Sarmah, Maensiri and Theerakulpisut2017). Comparable nanoparticle-based priming systems have also been reported to stimulate both α-amylase and protease activities, correlating with decreased starch and protein reserves and supporting embryo growth (Saharan et al., Reference Saharan, Kumaraswamy, Choudhary, Kumari, Pal, Raliya and Biswas2016). In parallel, enhanced antioxidative enzyme activity under AgNP treatment suggests that redox-mediated regulation may contribute to metabolic activation and reserve remobilization (Gupta et al., Reference Gupta, Agarwal and Pradhan2018). These biochemical changes are consistent with the reported stimulation of radicle elongation and early seedling establishment under AgNP priming conditions (Méndez-Andrade et al., Reference Méndez-Andrade, Ruiz-Torres, Wang, García-Cerda and Vera-Reyes2024).
Redox signalling induced by AgNPs is directly linked to metabolic activation in germinating seeds (Fig. 4). Low AgNP doses stimulate glycolysis and the tricarboxylic acid (TCA) cycle, increasing the production of NADH and FADH₂, which fuel mitochondrial oxidative phosphorylation and ATP synthesis (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Prażak et al., Reference Prażak, Święciło, Krzepiłko, Michałek and Arczewska2020; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023). This enhanced energy supply supports membrane repair and cell division associated with radicle emergence, while sustaining reserve mobilization and growth during early seedling establishment.
Concurrently, amino acid, lipid and secondary metabolite pathways are upregulated, providing structural components and protective molecules required for rapid growth (Kumar et al., Reference Kumar, Muthukrishnan and Rajalakshmi2020; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). This coordinated metabolic reprogramming explains the improved vigour, synchronized germination and increased stress tolerance observed in AgNP-primed seeds (Table 1) (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Kumar et al., Reference Kumar, Muthukrishnan and Rajalakshmi2020; Prażak et al., Reference Prażak, Święciło, Krzepiłko, Michałek and Arczewska2020; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025).
At high AgNP doses, excessive ROS disrupt mitochondrial function and oxidative phosphorylation, leading to decreased ATP production and altered NAD(P)H homeostasis (Fig. 4) (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011). This metabolic collapse, together with membrane lipid peroxidation (MDA accumulation), results in delayed germination and growth inhibition (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014).
Dose-dependent modulation of seed metabolism by AgNPs. Low AgNP doses stimulate central carbon metabolism and mitochondrial energy production, whereas high doses disrupt oxidative phosphorylation and redox balance, leading to metabolic dysfunction and growth inhibition. The central panel depicts the core metabolic network (glycolysis, the TCA cycle and NAD(P)H-linked redox reactions) underlying these contrasting responses.

Figure 4 Long description
The diagram illustrates the effect of seed coat thickness on the bioavailability of silver nanoparticles. Image A shows a thin or permeable seed coat allowing both silver nanoparticles and ions to penetrate and reach the embryo. Image B depicts a thick or lignified seed coat, which blocks silver nanoparticles, resulting in silver ion dominance and limited penetration to the embryo.
Seed coat and nanoparticle properties as determinants of AgNP bioavailability
Seed coat structure and composition vary widely across plant taxa, encompassing differences in layer number, tissue specialization and biochemical composition, from multi-layered coats in model and legume species to highly reduced structures in others (Radchuk and Borisjuk, Reference Radchuk and Borisjuk2014; Lee and Yeung, Reference Lee and Yeung2023). These include lignified testa layers, mucilage-producing epidermal cells and phenolic-rich tissues such as proanthocyanidin-accumulating endothelia. Mucilage composition also differs substantially among species, ranging from pectin-rich matrices such as rhamnogalacturonan I to hemicellulose-based polysaccharides including heteroxylans and arabinoxylans (North et al., Reference North, Berger, Saez-Aguayo and Ralet2014). In addition, seed coats may accumulate phenolics (Briggs et al., Reference Briggs, Morris and Ashford2005) and other defensive or toxic metabolites (Radchuk and Borisjuk, Reference Radchuk and Borisjuk2014).
These structural and chemical features determine key functional properties of the seed coat, including mechanical resistance through lignification and sclerenchyma (Briggs et al., Reference Briggs, Morris and Ashford2005; Radchuk and Borisjuk, Reference Radchuk and Borisjuk2014), permeability and water regulation via cuticles and permeability barriers (Briggs et al., Reference Briggs, Morris and Ashford2005; Debeaujon et al., Reference Debeaujon, Lepiniec, Pourcel and Routaboul2007), mucilage swelling properties (North et al., Reference North, Berger, Saez-Aguayo and Ralet2014), and control of solute diffusion and gas exchange (Debeaujon et al., Reference Debeaujon, Lepiniec, Pourcel and Routaboul2007). Importantly, these properties are also expected to modulate the interaction between AgNPs and the seed surface, influencing nanoparticle retention, penetration and Ag⁺ release dynamics.
The magnitude and spatial distribution of AgNP-induced redox signalling are strongly influenced by seed coat characteristics and nanoparticle physicochemical properties, which together regulate the relative contribution of nanoparticle-derived versus ionic silver species (Fig. 3). Seed coats are not merely passive barriers: their permeability is governed by both structural (thickness, porosity) and biochemical features, including lignin, suberin and mucilage polysaccharides, which can interact with nanoparticle surface coatings and influence Ag⁺ release kinetics (Liu et al., Reference Liu, Liu, Zhu, Hu, Zhang, Guo, Yada and Cui2021).
Seeds with thin or highly permeable coats allow partial penetration of AgNPs into the endosperm and embryo, generating localized ROS and Ag⁺ signals that efficiently activate metabolic and signalling pathways (Mohamed et al., Reference Mohamed, Qayyum, Abdel-Hadi, Rehman, Ali and Rizwan2017; Chen et al., Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023; Arridho et al., Reference Arridho, Ilyas, Damayanti, Widajati and Qadir2025; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). In this context, both nanoparticle-specific effects (e.g., surface reactivity or physical interactions with cell walls) and ion-mediated effects may contribute to redox signalling. Released Ag⁺ ions readily interact with thiol-containing biomolecules such as glutathione and metallothioneins, and thiol-rich environments can further promote AgNP dissolution (Veronesi et al., Reference Veronesi, Deniaud, Gallon, Jouneau, Villanova, Delangle, Carrière, Kieffer, Charbonnier, Mintz and Michaud-Soret2016; Marchioni et al., Reference Marchioni, Gallon, Worms, Jouneau, Lebrun, Veronesi, Truffier-Boutry, Mintz, Delangle, Deniaud and Michaud-Soret2018).
In contrast, thick or lignified coats restrict nanoparticle entry, so that mainly Ag⁺ ions diffuse through the testa, often producing weaker or more heterogeneous responses, reflecting a shift towards predominantly ion-driven signalling. This distinction is consistent with plant uptake studies showing that the relative contribution of particulate versus ionic silver depends on dissolution dynamics and interface processes (Dang et al., Reference Dang, Wang, Cai, Zhou and Xing2020).
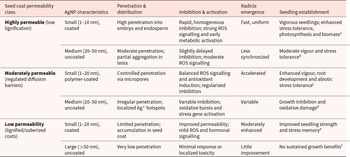
Nanoparticle size, shape, surface charge and coating further modulate this interaction (Table 2). Smaller AgNPs and those with less steric hindrance penetrate more easily, whereas green-synthesized and polymer-coated particles exhibit slower Ag⁺ release and reduced toxicity (Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Savassa et al., Reference Savassa, Castillo-Michel, Pradas Del Real, Reyes-Herrera, Marques and de Carvalho2021; Das and Dutta, Reference Das and Dutta2022; Labeeb et al., Reference Labeeb, Badr, Haroun, Mattar and El-Kholy2022; Thongmak et al., Reference Thongmak, Ruangwong, Wongkaew, Srisonphan and Onwimol2022; Elkelish et al., Reference Elkelish, Alqudah, Alammari, Alsubeie, Hamed and Thabet2025). In particular, surface coatings play a mechanistic role by binding Ag⁺ or creating diffusion barriers that modulate dissolution rates and spatial distribution of released ions (Liu et al., Reference Liu, Liu, Zhu, Hu, Zhang, Guo, Yada and Cui2021).
AgNP characteristics are presented according to the dominant physicochemical drivers of seed–nanoparticle interactions

Table 2 Long description
The table examines how different seed coat permeability classes interact with various AgNP characteristics, affecting seed and seedling development. Highly permeable seed coats with small, coated nanoparticles result in high penetration and vigorous seedlings with enhanced stress tolerance. In contrast, low permeability coats with large, uncoated nanoparticles show very low penetration and minimal growth benefits. Moderately permeable coats with small, polymer-coated nanoparticles lead to controlled penetration and enhanced vigor, while medium, uncoated nanoparticles cause variable growth outcomes. The data suggest that nanoparticle size, coating, and seed coat permeability significantly influence seedling establishment and stress responses.
Notes:
a Abou-Zeid and Moustafa (Reference Abou-Zeid and Moustafa2014); Elkelish et al. (Reference Elkelish, Alqudah, Alammari, Alsubeie, Hamed and Thabet2025); Thongmak et al. (Reference Thongmak, Ruangwong, Wongkaew, Srisonphan and Onwimol2022).
b Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023); Thongmak et al. (Reference Thongmak, Ruangwong, Wongkaew, Srisonphan and Onwimol2022).
c Elkelish et al. (Reference Elkelish, Alqudah, Alammari, Alsubeie, Hamed and Thabet2025); Sahoo et al. (Reference Sahoo, Sahu and Behera2025); Handayani et al. (Reference Handayani, Tanadi and Umar2025).
d Abou-Zeid and Moustafa (Reference Abou-Zeid and Moustafa2014); Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023).
e Das and Dutta (Reference Das and Dutta2022); Thongmak et al. (Reference Thongmak, Ruangwong, Wongkaew, Srisonphan and Onwimol2022); Lopes et al. (Reference Lopes, de Oliveira Silva and Courrol2023).
f Alhammad et al. (Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023); Sahoo et al. (Reference Sahoo, Sahu and Behera2025).
These parameters together define the effective redox dose perceived by the seed, which emerges from the dynamic interplay between seed coat composition and nanoparticle surface chemistry, explaining species- and formulation-specific responses to nanopriming and highlighting the need to distinguish nanoparticle-mediated from ion-mediated effects.
A redox-guided framework for AGNP seed priming
The experimental evidence accumulated in recent years indicates that the effects of AgNP seed priming are not random or species-independent but instead follow reproducible redox- and metabolism-driven patterns across crops. Studies in rice, wheat, maize, pea, watermelon and cucumber have consistently shown that low AgNP doses accelerate germination, enhance seedling vigour and improve tolerance to abiotic stresses, whereas higher doses lead to oxidative damage and growth inhibition (Nair and Chung, Reference Nair and Chung2014; Acharya et al., Reference Acharya, Jayaprakasha, Semper and Patil2020; Savassa et al., Reference Savassa, Castillo-Michel, Pradas Del Real, Reyes-Herrera, Marques and de Carvalho2021; Das and Dutta, Reference Das and Dutta2022; Chen et al., Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023; Rahman et al., Reference Rahman, Chakraborty, Kibria and Hossain2023; Mickky, Reference Mickky2025).
These outcomes are governed by the interaction between seed biological traits, nanoparticle physicochemical properties and priming conditions. Seed coat permeability, imbibition rate and intrinsic antioxidant capacity determine how much AgNP-derived Ag⁺ and ROS reach internal tissues during hydration (Mazumdar and Ahmed, Reference Mazumdar and Ahmed2011; Abou-Zeid and Moustafa, Reference Abou-Zeid and Moustafa2014; Prażak et al., Reference Prażak, Święciło, Krzepiłko, Michałek and Arczewska2020; Trzcińska-Wencel et al., Reference Trzcińska-Wencel, Mucha, Rai, Tyburski and Golińska2025). In parallel, nanoparticle size, coating, oxidation state and release kinetics control the intensity and spatial distribution of redox signals (Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Mawale et al., Reference Mawale, Nandini and Giridhar2024; Mickky, Reference Mickky2025). Green-synthesized and organically coated AgNPs, which display slower ion release and higher colloidal stability, therefore tend to produce broader beneficial windows than chemically synthesized particles (Abdel-Aziz and Rizwan, Reference Abdel-Aziz and Rizwan2019; Alhammad et al., Reference Alhammad, Abdel-Aziz, Seleiman and Tourky2023; Chen et al., Reference Chen, Liu, Yangzong, Gardea-Torresdey, White and Zhao2023; Mawale et al., Reference Mawale, Nandini and Giridhar2024).
Importantly, these responses arise from the combined contribution of nanoparticle-specific interactions and the controlled release of Ag⁺ ions, which together define the effective redox signal perceived by the seed. The resulting redox environment modulates MAPK signalling, glutathione homeostasis, mitochondrial metabolism and the activation of hydrolytic enzymes involved in starch and protein reserve remobilization, thereby determining whether metabolic reprogramming supports reserve mobilization, cell expansion, embryo growth, radicle protrusion and stress priming or instead collapses into oxidative injury (Figs 3 and 4) (Acharya et al., Reference Acharya, Jayaprakasha, Semper and Patil2020; Kumar et al., Reference Kumar, Muthukrishnan and Rajalakshmi2020; Méndez-Andrade et al., Reference Méndez-Andrade, Ruiz-Torres, Wang, García-Cerda and Vera-Reyes2024; Pintos et al., Reference Pintos, de Diego and Gomez-Garay2024; Mickky, Reference Mickky2025). This redox-centred control explains why apparently similar AgNP formulations can produce contrasting physiological outcomes in different species and experimental settings.
Taken together, this framework (Fig. 5) can be interpreted as a predictive model in which the outcome of AgNP seed priming emerges from the balance between redox signal intensity and the intrinsic buffering capacity of the seed. When nanoparticle-derived ROS and Ag⁺ signals remain within the detoxification and redox-regulatory capacity of the seed (largely determined by glutathione availability, antioxidant enzyme activity and seed coat permeability), priming promotes metabolic activation, improved germination and the establishment of a primed physiological state. Conversely, when this balance is exceeded, oxidative damage, metabolic disruption and growth inhibition prevail. This balance-based perspective provides a mechanistic basis for defining species-specific optimal dose windows and for guiding the rational design of nanopriming strategies based on seed traits and nanoparticle properties.
Schematic of the predictive framework integrating seed traits, nanoparticle properties and priming conditions within a multiscale model linking redox signalling to germination outcomes and stress memory establishment.

Figure 5 Long description
A diagram illustrating the relationship between seed traits, nanoparticle properties and priming conditions on redox balance. The central oval labeled 'Redox balance (ROS plus Ag superscript plus left right arrow buffering capacity)' connects to two outcomes. The left box, labeled 'Within buffering capacity', lists 'Controlled ROS', 'Metabolic activation' and 'Primed state', leading to 'Enhanced germination, stress tolerance and stress memory'. The right box, labeled 'Exceeds buffering capacity', lists 'Excess ROS', 'GSH depletion' and 'Mitochondrial dysfunction', leading to 'Impaired germination and growth inhibition'.
Acknowledgements
The authors thank colleagues for helpful discussions and feedback during the preparation of this manuscript.
Funding statement
This research received no specific grant from any funding agency, commercial or not-for-profit sectors. Open access publication was supported by the institutional open access agreement.
Competing interests
The authors declare no conflicts of interest.







 Open access
Open access