


Introduction
Maritrema gratiosum Nicoll, Reference Nicoll1907 (Digenea: Microphallidae) is a microphallid trematode with a heteroxenous life cycle involving intertidal invertebrates as first and second intermediate hosts and marine coastal birds as definitive hosts. First described from British shorebirds in the early 20th century (Nicoll Reference Nicoll1907), the species has been known under various synonyms, including Cercaria balani Lebour, Reference Lebour1908, Cercaria littorinae saxatilis V Popiel, 1976, and Maritrema arenaria Hadley & Castle, Reference Hadley and Castle1940 (Deblock and Stam Reference Deblock and Stam1960; Deblock and Tran Van Ky Reference Deblock and Tran Van Ky1966; Irwin et al. Reference Irwin, Maguire and Saville1990). The species is primarily reported from the temperate waters of western Europe (Northeast Atlantic), where its life cycle has been characterised and where it has been the subject of detailed morphological, experimental, and, more recently, molecular work (Chuang Reference Chuang2021; Chuang et al Reference Chuang, Shinn and Bron2025; Deblock and Tran Van Ky Reference Deblock and Tran Van Ky1966; Hadley and Castle Reference Hadley and Castle1940; Irwin Reference Irwin1983; Irwin et al. Reference Irwin, Maguire and Saville1990; Nicoll Reference Nicoll1907; Popiel Reference Popiel1976a, Reference Popielb, Reference Popiel1978; Tkach et al Reference Tkach, DTJ, Olson, Kinsella and Swiderski2003).
The life cycle of M. gratiosum begins when the miracidia hatch from eggs in the intestine of its first intermediate host (a marine gastropod). Within the reproductive system of the gastropod, miracidia develop into sporocysts and undergo asexual reproduction to produce daughter sporocysts (Carrol et al. Reference Carrol, Montgomery and Hanna1990; Popiel, Reference Popiel1978). In turn, daughter sporocysts give rise to clonal cercariae, which are ultimately released into the water column (Carrol et al. Reference Carrol, Montgomery and Hanna1990). Although motile, cercariae exhibit limited swimming ability (Hadley and Castle, Reference Hadley and Castle1940; Williams et al. Reference Williams, Ellis and Cross1981). The free cercariae infect the second intermediate host (a barnacle). Cercariae enter the body of the barnacle via the cirri during feeding and encyst as metacercariae embedded in the intestinal wall (Irwin et al. Reference Irwin, Maguire and Saville1990).
Metacercariae remain encysted until the second intermediate host (the barnacle) is ingested by the definitive host, which is typically a shorebird (Charadriiformes) or a waterfowl (Anseriformes), and less commonly a passerine (Passeriformes) or a grebe (Podicipediformes) (Ching Reference Ching1978; Deblock and Rausch Reference Deblock and Rausch1972; Hadley and Castle Reference Hadley and Castle1940; Nicoll Reference Nicoll1907; Presswell et al. Reference Presswell, Blasco-Costa and Kostadinova2014). Within the digestive tract of the avian host, metacercariae excyst and develop into sexually mature adults. Subsequently, eggs are produced and, eventually, expelled into the marine environment through defecation, where they are ingested by the first intermediate (molluscan) host, completing the life cycle of M. gratiosum.
Beyond the known hosts distributed in the Northeast Atlantic, M. gratiosum has been reported from a wide range of other hosts and regions. Yet, knowledge about its global distribution and life cycle remains fragmented, with many records based on morphology and incomplete identification of intermediate or definitive hosts. To synthesise this information, we consolidated published occurrence records to reconstruct the species’ global distribution and identify regions where its life cycle remains unresolved. Further, genetic evidence in support of species identity remains geographically limited for such a widespread trematode species. In particular, all known genetic sequences of M. gratiosum are from trematodes collected in the Northeast Atlantic (Tkach et al. Reference Tkach, DTJ, Olson, Kinsella and Swiderski2003; Chuang Reference Chuang2021). Here, we employed genetic methods to confirm the taxonomic identity of metacercariae of M. gratiosum recovered from barnacles in the Northeast Pacific (British Columbia), supporting previous identifications in this region (Ching Reference Ching1978, Reference Ching1991; MacLeod et al. Reference MacLeod, Armstrong and Wang2021). Through this case study and a comprehensive review of published records, we aim to clarify the known distribution of this trematode and to assess the extent to which its life cycle and host associations have been described across its range.
Material and methods
Collection of animals
In December 2020, individuals of Balanus glandula Darwin, 1854 were collected from Point Grey Park (49°16′26.0″N; 123°09′52.2″W), along the southern shore of Burrard Inlet in Vancouver, British Columbia, Canada – a site previously reported to exhibit a high prevalence of metacercarial infection (sensu MacLeod et al. Reference MacLeod, Armstrong and Wang2021). Collections targeted individuals of B. glandula inhabiting the intertidal zone. In the laboratory, somatic tissues of B. glandula were extracted using dissection forceps and examined under a light microscope for the presence of putative metacercariae of Maritrema gratiosum (Figure 1). Infected tissues from two individuals were preserved in 95% ethanol and stored at −20 °C until extraction of deoxyribonucleic acid (DNA).
(A) Encysted metacercariae of Maritrema gratiosum in the somatic tissue of Balanus glandula. (B) Close-up view of a metacercaria (black arrow). Scale bars represent 300 μm.

Genetic analysis
Genomic DNA was extracted from preserved trematode cyst tissues in B. glandula using the DNeasy Blood & Tissue Kit (QIAGEN) following the manufacturer’s protocols, with the exception that samples were eluted in a final volume of 50 μL. DNA concentrations were assessed using a Qubit fluorometer (Thermo Fisher Scientific).
Amplification via polymerase chain reaction (PCR) was used to target a partial region (~750 bp) of the 28S ribosomal RNA gene (28S) using the primers T16: 5′-GAG ACC GAT AGC GAA ACA AGT AC-3′ and T30: 5′-TGT TAG ACT CCT TGG TCC GTG-3′ (Harper and Saunders Reference Harper and Saunders2001). Additional amplification was used to target a partial region of the cytochrome c oxidase subunit I (COI) gene using the primers JB3: 5′-TTT TTT GGG CAT CCT GAG GTT TAT-3′ and Trem.cox1.rrnl: 5′-AAT CAT GAT GCA AAA GGT A-3′ (Bowles and McManus Reference Bowles and McManus1993; Král’ová et al. Reference Král’ová, Hanzelová, Scholz, Gerdeaux and Spakulová2001).
PCRs were prepared with 12.5 μL FroggaBio 2×Taq, 0.6 μL of each forward and reverse primer, 3.55 μL of Bovine Serum Albumin (New England BioLabs), 2.55 μL of UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific), and 5 μL of genomic DNA for a final volume of 25 μL. PCR conditions for samples prepared with the T16/T30 and JB3/Trem.cox1.rrnl primer sets included the polymerase activation of 94 °C for three minutes, followed by 40 cycles of denaturation and amplification at 94 °C for 30 seconds, 58 °C for 45 seconds, and 72 °C for one minute, then followed by a final elongation step of 72 °C for eight minutes. PCR amplification products were visualised on a 1.5% agarose gel stained with Red Safe (FroggaBio).
Non-purified PCR product was submitted to the Centre d’expertise et de services Génome Québec (Montréal, Canada) for Sanger sequencing. Sequences were obtained in both forward and reverse directions and edited using Geneious (version 11.0.4). A consensus sequence was formed by merging the forward and reverse sequences via de novo assembly.
Taxonomic assignment was carried out using BLASTn searches via the Basic Local Alignment Search Tool (BLAST) against the nucleotide database (nr/nt) of the National Center for Biotechnology Information (NCBI). Further taxonomic validation was carried out by placing our unknown sequences into a neighbour-joining tree with previously published Maritrema spp. sequences obtained from NCBI. This tree was constructed in Geneious v11.1.5 with 100 bootstrap replicates and all sequences trimmed to equal length (468 bp).
Results and discussion
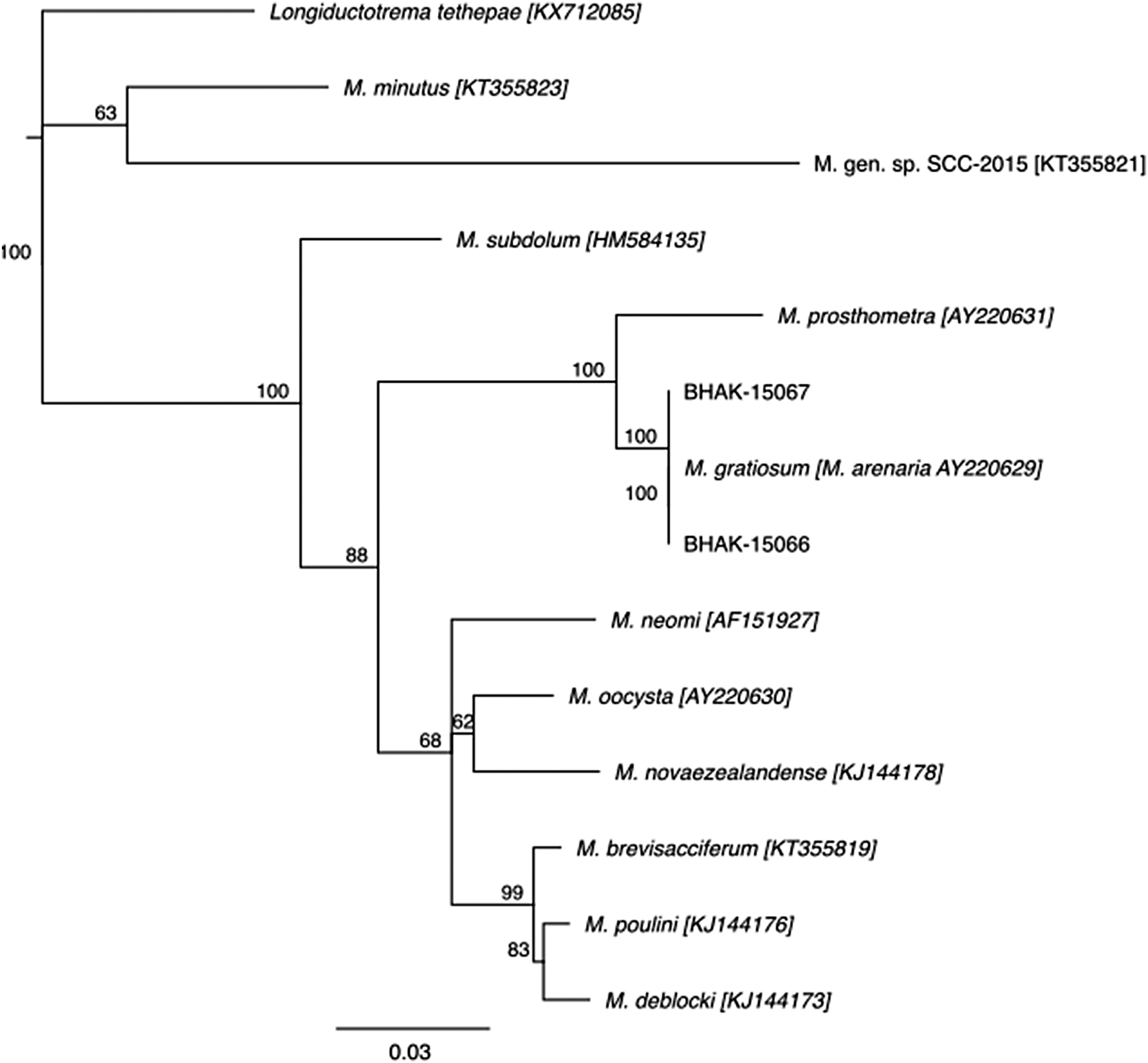
In this study, we used molecular tools to support the identity of the trematode parasitising the barnacle Balanus glandula in British Columbia, Canada, as Maritrema gratiosum. This represents the first molecular confirmation of M. gratiosum outside western Europe. Amplification using T16 / T30 (28S) was successful in yielding parasite DNA sequences from two samples (Barcode of Life Data System [BOLD]; https://doi.org/10.5883/DS-TREM). These sequences were >99% identical to Maritrema gratiosum (previously Maritrema arenaria) from Northern Ireland (NCBI Accession: AY220629.1; Tkach et al. Reference Tkach, DTJ, Olson, Kinsella and Swiderski2003). Tree-based analysis also placed our sequences into a clade with this species with zero nucleotide differences (Figure 2). The next closest clade was Maritrema prosthometra Deblock & Heard, 1969, isolated from rodents in Florida, with 96% similarity (20 nucleotide differences; AY220631.1; Galaktionov et al. Reference Galaktionov, Blasco-Costa and Olson2012; Tkach et al. Reference Tkach, DTJ, Olson, Kinsella and Swiderski2003). In a separate analysis, sequences isolated from Semibalanus balanoides (Linnaeus 1767) collected from Scotland were identical to those from Northern Ireland (Chuang, Reference Chuang2021). We also obtained sequences using the COI primers (JB3/Trem.cox1.rrnl) for the same specimens. However, these sequences had low similarity to publicly available reference sequences in BOLD and NCBI, and were not informative for inferring species identity. All DNA sequence data from both markers are publicly accessible in BOLD (https://doi.org/10.5883/DS-TREM) and NCBI (COI: PZ385258–PZ385259; 28S: PZ385594–PZ385595).
Neighbour-joining tree combining Maritrema sp. sequences from this study (BHAK-15066 and BHAK-15067) with previously published sequences of Maritrema spp. and Microphallus spp. (family Microphallidae) obtained from NCBI. Branch labels are consensus support (%), and tip labels for public records include the corresponding GenBank accession numbers.

Figure 2. Long description
The tree originates from a root on the left labeled with a consensus support value of 100%.
* The uppermost branch leads to the outgroup Longiductotrema tethepae [K X7 12 08 5].
* Below this, a clade with a consensus support value of 63% splits into M. minutus [K T3 55 82 3] and M. gen. sp. SCC-2015 [K T3 55 82 1].
* A major lower clade starts with a consensus support value of 100%. The first branch off this clade leads to M. subdolum [H M5 84 13 5].
* The remaining branches descend from a node with a consensus support value of 88%.
* A sub-clade with a consensus support value of 100% contains M. prosthometra [A Y2 20 63 1] and a cluster of three closely related sequences: BHAK-15067, M. gratiosum [M. arenaria AY2 20 62 9], and BHAK-15066, all joined with consensus support values of 100%.
* Further down, a node with a consensus support value of 68% leads to a group containing M. neomi [A F1 51 92 7], M. oocysta [A Y2 20 63 0], and M. novaezealandense [K J 1 4 4 1 7 8] with a consensus support of 62%.
* The bottom-most clade, with consensus support value of 99%, splits into M. brevisacciferum [K T3 55 81 9] and a pair consisting of M. poulini [K J1 44 17 6] and M. deblocki [K J1 44 17 3] with a consensus support value of 83%.
* A scale bar at the bottom indicates a genetic distance of 0.03.
Based on published occurrence records derived from both morphological and molecular identifications, M. gratiosum has a broad distribution spanning subtropical to arctic waters across both the Northern and Southern Hemispheres (Figure 3). In subtropical waters, the species has been detected in the South China Sea and the Gulf of Mexico (Figure 3). In temperate waters, records extend across western Europe, the Mediterranean Sea, British Columbia, the eastern United States, and New Zealand, while occurrences in subarctic waters span the Kuril Islands, the Bering Sea, and the Gulf of Alaska (Figure 3). At the northernmost extent of its range, the species has been documented in the arctic waters of the Barents Sea (Figure 3). One published record was excluded from this review. Hassan et al. (Reference Hassan, Al-Solami and Hassan2018) reported a trematode from marine gastropods in the Red Sea, attributing it to Cercaria littorinae saxatilis V (a synonym of M. gratiosum; Irwin et al. Reference Irwin, Maguire and Saville1990). Although the tail length was comparable to prior descriptions, the daughter sporocysts, cercariae, and stylets were substantially smaller than those reported for M. gratiosum, and the ventral sucker was described as absent rather than present (Irwin et al. Reference Irwin, Maguire and Saville1990; Popiel Reference Popiel1976b). These discrepancies suggest a likely misidentification.
Global distribution of the microphallid trematode Maritrema gratiosum based on occurrence records reported in the literature, spanning subtropical (South China Sea, Gulf of Mexico), temperate (western Europe, Mediterranean Sea, British Columbia, eastern United States, New Zealand), subarctic (Kuril Islands, Bering Sea, Gulf of Alaska), and arctic waters (Barents Sea). A subset of these records was supported by molecular data, including records from temperate waters in British Columbia (this study), Northern Ireland (Tkach et al Reference Tkach, DTJ, Olson, Kinsella and Swiderski2003), and Scotland (Chuang Reference Chuang2021). Sources of distributional data are provided in Table 1.

Figure 3. Long description
A world map displays the distribution of the microphallid trematode. A legend at the bottom left identifies red circles as ‘All occurrence records’ and green squares as ‘Records based on molecular data’.
* In the northwest, red circles are located in the Gulf of Alaska and the eastern United States. A green square and red circle are paired in British Columbia. Two red circles are in the Gulf of Mexico.
* In the north, two red circles are positioned in the Barents Sea.
* In the northeast, red circles mark the Bering Sea, Kuril Islands, and South China Sea.
* In the central region, western Europe contains a cluster of red circles and two green squares. A single red circle is located in the Mediterranean Sea.
* In the southeast, a single red circle is located off the coast of New Zealand.
The map includes latitude and longitude grid lines, with the equator and prime meridian clearly visible.
A review of the literature indicates that only in western Europe and the Barents Sea is the complete life cycle of M. gratiosum resolved, and the identity of associated hosts (or at least one complete set of hosts required to complete the life cycle) is known. From these regions, the hosts consisted of the periwinkle Littorina saxatilis (Olivi 1792) as its first intermediate host, S. balanoides and other barnacle species as its second intermediate host, and various species of birds as its definitive host (Table 1). In contrast, the life cycle of M. gratiosum remains incompletely known in other parts of its range. In British Columbia and the eastern United States, only two of three hosts (namely, the second intermediate and definitive hosts) have been identified (Table 1). To date, only the second intermediate host has been identified from the Bering Sea, and only the definitive host from the South China Sea, the Gulf of Mexico, New Zealand, and the Gulf of Alaska (Table 1). Host identity was not specified for the records of M. gratiosum from the Kuril Islands (Table 1). The molecular approach used in this study could be employed to identify new hosts (including paratenic hosts), which can contribute to a more complete understanding of the trematode’s life cycle for regions outside of western Europe and the Barents Sea.
Hosts in the life cycle of the microphallid trematode Maritrema gratiosum that have been identified in the literature. Names of host species were updated to their current valid nomenclature. Names of regions correspond to those shown in Figure 3

Table 1. Long description
The table is organized into four columns: Climate and region, Identified hosts in the life cycle of the trematode, Host species, and Sources.
* Subtropical Zone:
- South China Sea: Definitive hosts include Arenaria interpres interpres, Charadrius alexandrinus dealbatus, and Tringa nebularia.
- Gulf of Mexico: Definitive hosts include Limnodromus griseus and Numenius americanus.
* Temperate Zone:
- Western Europe: First intermediate host is Littorina saxatilis; Second intermediate hosts include Austrominius modestus, Chthamalus montagui, Perforatus perforatus, and Semibalanus balanoides; Paratenic host is Nucella lapillus; Definitive hosts include Calidris alpina, Charadrius hiaticula, and several Larus species.
- Mediterranean Sea: Definitive host is Calidris minuta.
- British Columbia: Second intermediate host is Balanus glandula; Definitive host is Bucephala islandica.
- Eastern United States: Second intermediate host is Semibalanus balanoides; Definitive host is Arenaria interpres morinella.
- New Zealand: Definitive hosts include Haematopus finschi and Limosa lapponica baueri.
* Subarctic Zone:
- Kuril Islands and eastern Siberia: No host data specified.
- Bering Sea: Second intermediate host is Semibalanus cariosus.
- Gulf of Alaska: Definitive hosts include Corvus brachyrhynchos and Haematopus bachmani.
* Arctic Zone:
- Barents Sea: First intermediate host is Littorina saxatilis; Second intermediate host is Semibalanus balanoides; Definitive hosts include Larus argentatus, Larus hyperboreus, and Larus marinus.
a Most likely to be M. gratiosum.
b Most likely referring to localities on the Kuril Islands or in the Bering Sea.
The survey of the literature revealed that L. saxatilis is the only first intermediate host species in which M. gratiosum has been identified to date (Galaktionov and Bustnes Reference Galaktionov and Bustnes1999; Irwin Reference Irwin1983; McCarthy et al. Reference McCarthy, Fitzpatrick and Irwin2002; Popiel Reference Popiel1976b), a host association further supported by experimental transmission studies (Irwin et al. Reference Irwin, Maguire and Saville1990). Beyond B. glandula and S. balanoides, known second intermediate host species include the barnacles Austrominius modestus (Darwin, 1854), Chthamalus montagui Southward, 1976, Perforatus perforatus (Bruguière, 1789), and Semibalanus cariosus (Pallas, 1788) (Ching Reference Ching1978; Chuang Reference Chuang2021; Sari and Malek Reference Sari and Malek2000; Williams et al. Reference Williams, Ellis and Cross1981). The most commonly reported definitive hosts are shorebirds, including sandpipers (genus Calidris), plovers (Charadrius), gulls (Chroicocephalus and Larus), oystercatchers (Haematopus), turnstones (Arenaria), terns (Sterna), curlews (Numenius), godwits (Limosa), and greenshanks (Tringa) (Table 2). Infections have also been described in several waterfowl – a goldeneye (Bucephala), a scoter (Melanitta), and an eider (Somateria) – as well as in a passerine (Corvus) and a grebe (Podiceps) (Table 2). In addition to these obligate hosts, the marine gastropod Nucella lapillus (Linnaeus 1758) has been identified as a paratenic host of M. gratiosum (McCarthy et al. Reference McCarthy, Irwin and Fitzpatrick1999). This gastropod is a predator of the barnacle S. balanoides, a known second intermediate host (Irwin et al. Reference Irwin, Maguire and Saville1990), and is itself consumed by birds, the definitive hosts (McCarthy et al. Reference McCarthy, Irwin and Fitzpatrick1999). The detection of metacercarial cysts in N. lapillus suggests that infection occurs through the consumption of infected barnacles. Paratenic hosts are thought to augment and maintain transmission between the second intermediate and avian hosts (Marcogliese Reference Marcogliese2007).
Documented definitive hosts of the microphallid trematode Maritrema gratiosum. Names of host species were updated to their current valid nomenclature. Names of regions correspond to those shown in Figure 3

Table 2. Long description
The table is organized into four columns: Host species, Common name, Regions, and Sources. It is divided into four taxonomic sections:
* Charadriiformes: Includes 21 entries such as the Ruddy turnstone (South China Sea and eastern United States), Dunlin (western Europe), Little stint (Mediterranean Sea), various Plovers (South China Sea and western Europe), Black-headed gull (western Europe), Oystercatchers (New Zealand, Gulf of Alaska, and western Europe), several Gull species (western Europe, Barents Sea, and unspecified regions), Short-billed dowitcher (Gulf of Mexico), Bar-tailed godwit (New Zealand and western Europe), Long-billed curlew (Gulf of Mexico), Arctic tern (unspecified), and Common greenshank (South China Sea).
* Anseriformes: Includes Barrow’s goldeneye (British Columbia), Common scoter (western Europe), and Common eider (western Europe).
* Passeriformes: Includes the American crow (Gulf of Alaska).
* Podicipediformes: Includes the Red-necked grebe (unspecified).
Sources cited include Fischthal and Kuntz (Reference Fischthal and Kuntz1976), Hadley and Castle (Reference Hadley and Castle1940), Nicoll (Reference Nicoll1907), Sonin (Reference Sonin1985), and Kuklin (Reference Kuklin2022), among others.
Building on earlier records from British Columbia (Ching Reference Ching1978, Reference Ching1991; MacLeod et al. Reference MacLeod, Armstrong and Wang2021), this study provides the first molecular evidence confirming the presence of M. gratiosum in this region and extends its molecularly confirmed range from the North Atlantic to the Northeast Pacific. Confirming the presence of this trematode species in the barnacle B. glandula will pave the way to a better understanding of host–parasite dynamics, given suggestions that M. gratiosum may reduce the survival of its second intermediate host and influence the barnacle’s distribution on intertidal rocky shores (Carrol et al. Reference Carrol, Montgomery and Hanna1990; Mouritsen and Poulin Reference Mouritsen and Poulin2002). Yet, a substantial gap remains in our understanding of the regional life cycle in British Columbia, most notably, the identity of the first intermediate host (MacLeod et al. Reference MacLeod, Armstrong and Wang2021). A similar gap was highlighted for the eastern United States, where the first intermediate host of M. gratiosum likewise remains unidentified (Hadley and Castle Reference Hadley and Castle1940). Future work to fully characterise these regional life cycles and assess the effects of M. gratiosum on host species can help clarify the broader ecological consequences of trematode infection for marine ecosystems (Bennett et al. Reference Bennett, Presswell and Poulin2023).
Acknowledgements
We wish to extend our thanks to the staff at the Inter-Library Loan Service for their assistance in locating and obtaining literature that was not available through the University of British Columbia Library. This work was supported by the Sentinels of Change Alliance, an Alliance Grant co-sponsored by the Tula Foundation and the Natural Sciences and Engineering Research Council of Canada.
Competing interests
The authors declare that they have no competing interests.





 Open access
Open access