


1. Introduction
Scalidophora, occasionally also referred to as Cephalorhyncha (Nielsen, Reference Nielsen1995), is a small cycloneuralian clade of strictly marine benthic ecdysozoans, containing the vermiform phyla Priapulida, Kinorhyncha and Loricifera (Lemburg, Reference Lemburg1995; Maas, Reference Maas and Schmidt-Rhaesa2013; Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013a, b; Giribet and Edgecombe, Reference Giribet and Edgecombe2017; Brusca et al. Reference Brusca, Giribet and Moore2019; Giribet and Edgecombe, Reference Giribet and Edgecombe2020; Howard et al. Reference Howard, Edgecombe, Shi, Hou and Ma2020). While kinorhynchs are relatively speciose, with currently some 361 recognized extant species and more awaiting description (Yamasaki, Reference Yamasaki2022), priapulids and loriciferans are considerably less diverse, being represented by 23 (Yamasaki, Reference Yamasaki2018a) and 47 (Yamasaki, Reference Yamasaki2018b) described living species, respectively.
Although the validity of Scalidophora as a natural clade has found wide acceptance, it is based on the shared possession of a fully retractable introvert with radially arranged scalids and a ring-shaped brain surrounding the anterior part of the pharynx – anatomical characters which may, in fact, be plesiomorphic with respect to the cycloneuralian stem (Maas, Reference Maas and Schmidt-Rhaesa2013; Giribet and Edgecombe, Reference Giribet and Edgecombe2017; Giribet and Edgecombe, Reference Giribet and Edgecombe2020; Howard et al. Reference Howard, Edgecombe, Shi, Hou and Ma2020), or possibly even the Ecdysozoan stem, considering that multiple molecular studies recover Panarthropoda within a paraphyletic Cycloneuralia (Garey, Reference Garey2001; Mallatt and Giribet, Reference Mallatt and Giribet2006; Park et al. Reference Park, Hyun, Kristensen, Kim and Giribet2006; Dunn et al. Reference Dunn, Hejnol, Matus, Pang, Browne, Smith, Seaver, Rouse, Obst, Edgecombe, Sørensen, Haddock, Schmidt-Rhaesa, Okusu, Kristensen, Wheeler, Martindale and Giribet2008; Sørensen et al. Reference Sørensen, Hebsgaard, Heiner, Glenner, Willerslev and Kristensen2008; Rota-Stabelli et al. Reference Rota-Stabelli, Campbell, Brinkmann, Edgecombe, Longhorn, Peterson, Pisani, Philippe and Telford2011, Reference Rota-Stabelli, Daley and Pisani2013). The concept of Scalidophora has so far eluded rigorous molecular testing, in large part due to a lack of relevant extensive molecular datasets for the group (Giribet and Edgecombe, Reference Giribet and Edgecombe2017; Giribet and Edgecombe, Reference Giribet and Edgecombe2020; Howard et al. Reference Howard, Edgecombe, Shi, Hou and Ma2020). However, while the few molecular studies that have been conducted for the most part support a close link between Priapulida and Kinorhyncha (Mallatt and Giribet, Reference Mallatt and Giribet2006; Dunn et al. Reference Dunn, Hejnol, Matus, Pang, Browne, Smith, Seaver, Rouse, Obst, Edgecombe, Sørensen, Haddock, Schmidt-Rhaesa, Okusu, Kristensen, Wheeler, Martindale and Giribet2008; Mallatt et al. Reference Mallatt, Craig and Yoder2012; Borner et al. Reference Borner, Rehm, Schill, Ebersberger and Burmester2014; Laumer et al. Reference Laumer, Bekkouche, Kerbl, Goetz, Neves, Sørensen, Kristensen, Hejnol, Dunn, Giribet and Worsaae2015), the position of Loricifera has proven more problematic to establish (Park et al. Reference Park, Hyun, Kristensen, Kim and Giribet2006; Sørensen et al. Reference Sørensen, Hebsgaard, Heiner, Glenner, Willerslev and Kristensen2008; Yamasaki et al. Reference Yamasaki, Fujimoto and Miyazaki2015; Laumer et al. Reference Laumer, Bekkouche, Kerbl, Goetz, Neves, Sørensen, Kristensen, Hejnol, Dunn, Giribet and Worsaae2015, Reference Laumer, Fernández, Lemer, Combosch, Kocot, Riesgo, Andrade, Sterrer, Sørensen and Giribet2019).
The fossil record of scalidophorans is extremely patchy and unevenly distributed through time. Treptichnid burrows, which mark the base of the Cambrian (Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955)), are generally considered to have been made by a priapulid-like organism (Vannier et al. Reference Vannier, Calandra, Gaillard and Zylińska2010; Kesidis et al. Reference Kesidis, Slater, Jensen and Budd2019), with the oldest unequivocal scalidophoran body fossils also dating back to the Fortunian (Liu et al. Reference Liu, Xiao, Shao, Broce and Zhang2014; Zhang et al. Reference Zhang, Xiao, Liu, Yuan, Wan, Muscente, Shao, Gong and Cao2015; Shao et al. Reference Shao, Liu, Wang, Zhang, Tang and Li2016; Zhang et al. Reference Zhang, Maas and Waloszek2018). The group has an extensive and diverse near-global Cambrian record during the Terreneuvian–Miaolingian interval (see Maas, Reference Maas and Schmidt-Rhaesa2013 for a review), being known from both a wide array of exceptionally preserved body fossils and disarticulated microfossils (scalids, pharyngeal teeth, e.g. Smith et al. Reference Smith, Harvey and Butterfield2015; Shan et al. Reference Shan, Harvey, Yan, Li, Zhang and Servais2023; Wernström et al. Reference Wernström, Slater, Sørensen, Crampton and Altenburger2023, Mussini and Butterfield Reference Mussini and Butterfield2025a, b, and palaeoscolecid sclerites). Included among the Cambrian scalidophoran record are multiple stem priapulids (see Wang et al. Reference Wang, Vannier, Sun, Yu and Han2023 for a discussion on the complexities and inconsistencies of assigning fossils to the stem or crown of Priapulida based on morphological characters). The only known loriciferan fossils include the giant probable stem-loriciferan Sirilorica carlsbergi Peel, Reference Peel2010 from the Cambrian Series 2, Stage 3 Sirius Passet Fauna, and the highly miniaturized Eolorica deadwoodensis Harvey and Butterfield, Reference Harvey and Butterfield2017 from the Fortunian Deadwood Formation of Canada. Eokinorhynchus rarus Zhang et al. Reference Zhang, Xiao, Liu, Yuan, Wan, Muscente, Shao, Gong and Cao2015 from the Fortunian Dengying Formation in China was described as the first kinorhynch fossil, followed by the closely related Zhongpingscolex qinensis Shao et al. Reference Shao, Wang, Liu, Qin, Zhang, Liu, Shao, Zhao and Zhang2020 from the Fortunian Kuanchuanpu Formation in China. However, phylogenetic work by Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021, Reference Wang, Vannier, Sun, Yu and Han2023) and Wang (Reference Wang2022) has placed Eokinorhynchus within the priapulid stem, leaving the putative kinorhynch fossil record in doubt. In contrast to the extensive Cambrian record of Scalidophora, their post-Cambrian record is virtually limited to rather scarce palaeoscolecid body- and microfossils (Botting et al. Reference Botting, Muir, Van Roy, Bates and Upton2012; Howard et al. Reference Howard, Parry, Clatworthy, D’Souza and Edgecombe2024), which have an increased preservation potential due to the phosphatic mineralization of their cuticular plates: palaeoscolecids range at least up to the late Silurian (Botting et al. Reference Botting, Muir, Van Roy, Bates and Upton2012; Howard et al. Reference Howard, Parry, Clatworthy, D’Souza and Edgecombe2024), with Radnorscolex latus Howard et al. Reference Howard, Parry, Clatworthy, D’Souza and Edgecombe2024 from the lower Ludfordian of Leintwardine (Wales) being the youngest described member of the group, and may possibly have extended into the Devonian (PVR personal observation 2023). It must, however, be pointed out that, while palaeoscolecids are generally considered as either stem or crown members of Scalidophora (see e.g. Harvey et al. Reference Harvey, Dong and Donoghue2010; Shao et al. Reference Shao, Liu, Wang, Zhang, Tang and Li2016; Whitaker et al. Reference Whitaker, Jamison, Schiffbauer and Kimmig2020; Shi et al. Reference Shi, Howard, Edgecombe, Hou and Ma2021; Wang et al. Reference Wang, Vannier, Sun, Yu and Han2023), their phylogenetic position is debated, with some favouring a sister-group relationship to Cycloneuralia (Budd, Reference Budd2001) or Ecdysozoa as a whole (Conway Morris and Peel, Reference Conway Morris and Peel2010; Peel, Reference Peel2010), while other works resolve them as a grade of very basal stem-group Panarthropoda (Dzik and Krumbiegel, Reference Dzik and Krumbiegel1989; Smith and Dhungana, Reference Smith and Dhungana2022), or allied with the Nematomorpha (Hou and Bergström, Reference Hou and Bergström1994) or stem-group Nematoida more broadly (Budd, Reference Budd2001). A nematoid affinity for the group, however, is appearing increasingly unlikely (Harvey et al. Reference Harvey, Dong and Donoghue2010). Apart from Palaeoscolecida, after the Cambrian, Scalidophora is only represented by a specimen of Markuelia sp. from the lowermost Tremadocian (Lower Ordovician) Vinni Formation of Nevada, U.S.A. (Dong et al. Reference Dong, Bengtson, Gostling, Cunningham, Harvey, Kouchinsky, Val’Kov, Repetski, Stampanoni, Marone and Donoghue2010), two or three rare taxa from the late Tremadocian (Lower Ordovician) Fezouata Biota of Morocco (Van Roy et al. Reference Van Roy, Orr, Botting, Muir, Vinther, Lefebvre, Hariri and Briggs2010; Van Roy et al. Reference Van Roy, Briggs and Gaines2015; Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024), a possible specimen of Ottoia sp. from the Floian Liexi Fauna in China (Fang et al. Reference Fang, Mao, Liu, Yuan, Chen, Wu, Li, Zhang, Ma, Wang, Zhan, Peng, Zhang and Huang2022) and Priapulites konecniorum Schram, Reference Schram1973 from the late Moscovian (Pennsylvanian, Carboniferous) of Mazon Creek, Illinois, U.S.A. (Schram, Reference Schram1973). Notably, P. konecniorum represents the only supposedly unequivocal fossil of a crown-priapulid (but see Wang et al. Reference Wang, Vannier, Sun, Yu and Han2023), while fossils of the kinorhynch and loriciferan crown are entirely lacking (Maas, Reference Maas and Schmidt-Rhaesa2013; Giribet and Edgecombe, Reference Giribet and Edgecombe2020).
Within Scalidophora, the family Selkirkiidae (Conway Morris, Reference Conway Morris1977) is of particular interest due to their unique tubiculous mode of life within an annulated, sclerotized, open-ended tube with a flattened, oval cross-section, secreted by the animal (Conway Morris, Reference Conway Morris1977; Briggs et al. Reference Briggs, Erwin and Collier1994; Hou et al. Reference Hou, Bergström, Wang, Feng and Chen1999; Luo et al. Reference Luo, Hu, Chen, Zhang and Tao1999; Lan et al. Reference Lan, Yang, Hou and Zhang2015; Hou et al. Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Yang et al. Reference Yang, Vannier, Yang, Wang and Zhang2021; Wang, Reference Wang2022). While tube-dwelling lifestyles are a common strategy among a wide range of benthic metazoan taxa as diverse as cnidarians, annelids, molluscs, panarthropods, nematodes, phoronids, tentaculits and pterobranchs (see Supplementary Text for a discussion), the only other scalidophoran example is found in the minute extant priapulid genus Maccabeus (Por, Reference Por1973). The tube of both Maccabeus species is, however, constructed of algal fragments longitudinally embedded in mucus secreted by the animal (Por and Bromley, Reference Por and Bromley1974; von Salvini-Plawen, Reference von Salvini-Plawen1974; Malakhov, Reference Malakhov1978; Adrianov and Malakhov, Reference Adrianov and Malakhov1995; Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013b; Todaro et al. Reference Todaro, Dal Zotto and Rebecchi2019). Furthermore, among palaeoscolecids, Huang et al. (Reference Huang, Chen, Zhu and Zhao2014) showed that the early Cambrian Cricocosmia jinningensis Hou and Sun, Reference Hou and Sun1988 and Mafangscolex sinensis Hou and Sun, Reference Hou and Sun1988 both reinforced the walls of their burrows with secretions that agglutinated sediment grains and likely organic material, while Zhang et al. (Reference Zhang, Hou and Bergström2006) presented evidence that the early Cambrian Maotianshania cylindrica Sun and Hou, Reference Sun and Hou1987 also lined its burrows with secretions of mucus. Regardless, the flimsy tubes constructed by the extant Maccabeus and the linings secreted by the Cambrian Cricocosmia, Mafangscolex and Maotianshania are evidently very different from the rigid, sclerotized, and probably chitinous tube of selkirkiids. Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021) and Wang (Reference Wang2022) proposed that after having secreted it from the body wall, selkirkiids moulted their tube to detach it from their body, after which the animal lived freely within the tube. The animal would then discard the tube to moult its cuticle and grow, and repeat the process.
Although the tube would have considerably impaired the mobility of selkirkiids compared to other scalidophoran taxa, it did offer increased protection for their soft bodies. Selkirkiids have been proposed to have been largely epibenthic, while also being capable of at least shallow burrowing into the substrate (Maas et al. Reference Maas, Huang, Chen, Waloszek and Braun2007; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Yang et al. Reference Yang, Vannier, Yang, Wang and Zhang2021; Wang, Reference Wang2022). Based on a preferential attachment of epibionts to the rear of the tube in Chengjiang specimens, it is assumed that even when burrowing, they remained close to the sediment-water interface, with the rear part of the tube exposed on the surface to facilitate respiration and defecation (Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Yang et al. Reference Yang, Vannier, Yang, Wang and Zhang2021). Preserved gut contents suggest that they likely were generalists (Lan et al. Reference Lan, Yang, Hou and Zhang2015).
Selkirkiids were originally described as stem-group priapulids (Conway Morris, Reference Conway Morris1977) and are often still placed in this group without further reservations (Briggs et al. Reference Briggs, Erwin and Collier1994; Hou et al. Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017). However, as is the case for many other putative stem-group priapulids, unequivocally proving this assignment is challenging, because the anatomical synapomorphies uniting Priapulida (Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013b) are usually difficult if not impossible to observe and apply consistently in fossil material (Wang et al. Reference Wang, Vannier, Sun, Yu and Han2023). As a consequence, the position of selkirkiids in phylogenetic analyses has tended to be unstable and dependent on taxon sampling (Zhang et al. Reference Zhang, Xiao, Liu, Yuan, Wan, Muscente, Shao, Gong and Cao2015; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Wang, Reference Wang2022). Nevertheless, recent morphological and anatomical observations have provided increased support for a position in the priapulid stem (Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Yang et al. Reference Yang, Vannier, Yang, Wang and Zhang2021; Wang, Reference Wang2022).
Until the recent description of Selkirkia tsering Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024, from the Tremadocian Fezouata Biota of Morocco, selkirkiids were believed to be restricted to the Terreneuvian–Miaolingian interval and have been held up as typical Cambrian ‘Burgess Shale-type’ organisms (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024). Here, we describe two specimens of an even considerably younger selkirkiid from a newly discovered exceptionally preserved fauna from the Upper Pragian (Lower Devonian) La Roche Formation in Warmifontaine (Neufchâteau), Luxembourg Province, Wallonia, Belgium (Figures 1 and 2), extending the range of the group by a remarkable 70 Ma into the late Palaeozoic. While the preservational state of the specimens unfortunately precludes a formal description, and the material is consequently left under open nomenclature, the fossils are nevertheless readily identifiable as selkirkiids with preserved soft parts (see also Supplementary Text for a discussion and refutation of alternative phylogenetic affinities).
Schematic geological map of southern Belgium and adjacent countries and simplified structural setting of the Palaeozoic formations (adapted from de Béthune, Reference de Béthune and Fourmarier1954 and Candela and Mottequin, Reference Candela and Mottequin2022).

Figure 1. Long description
The map illustrates the geological features of southern Belgium and neighboring countries, focusing on Palaeozoic formations. It highlights various geological units such as the Brabant Massif, Dinant Synclinorium, Rocroi Massif, Serpont Massif, Givonne Massif, and Ardenne Allochthon. Key cities like Mons, Charleroi, Namur, Dinant, Liege, Bastogne, Neufchateau, and Lierneux are marked. The map uses different colors to represent various geological periods, including Mesozoic and Cenozoic, Carboniferous, Permian, Emsian, Lower Devonian, Lochkovian, Middle and Upper Devonian, and Pragian. Fault lines are indicated with red lines, and specific localities are marked with stars. The map provides a detailed view of the geological structure and formations in the region.
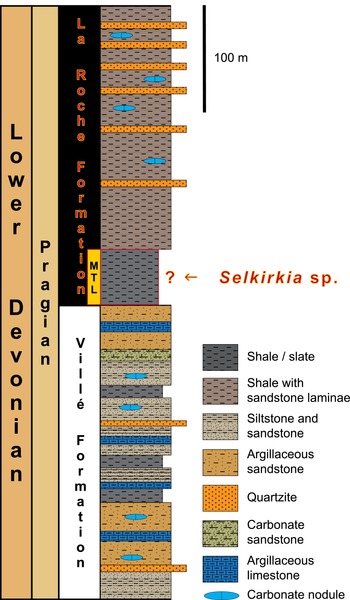
Schematic lithological column of the Pragian lithostratigraphic units (partim) encountered in the western part of the Neufchâteau–Eifel Synclinorium (modified from Ghysel, Reference Ghysel2022), The range of Selkirkia sp. within the lower part of the La Roche Formation is unknown. Abbreviation: MTL, Martelange Member.

Figure 2. Long description
The schematic lithological column illustrates the Pragian lithostratigraphic units encountered in the western part of the Neufchâteau-Eifel Synclinorium. The column is divided into two main formations: the Ville Formation and the La Roche Formation. The Ville Formation consists of various layers including shale, shale with sandstone laminae, siltstone and sandstone, argillaceous sandstone, quartzite, carbonate sandstone, and argillaceous limestone. The La Roche Formation, which is the focus of the column, contains multiple layers of shale, shale with sandstone laminae, siltstone and sandstone, argillaceous sandstone, quartzite, and carbonate nodules. The Martelange Member is indicated within the La Roche Formation. The range of Selkirkia sp. within the lower part of the La Roche Formation is unknown.
2. Geological setting
Warmifontaine is located on the northern flank of the Neufchâteau–Eifel Synclinorium, a major Variscan tectonic unit belonging to the Ardenne Allochthon (Belanger et al. Reference Belanger, Delaby, Delcambre, Ghysel, Hennebert, Laloux, Marion, Mottequin and Pingot2012; Figure 1). In this village, a subterranean quarry exploited the dark grey to black slates of the Martelange Member that corresponds to the lower part of the La Roche Formation (Upper Pragian, Lower Devonian; Ghysel Reference Ghysel2022; Figure 2). These slates are slightly older than the Lower Emsian Hunsrück slates from Bundenbach, southeastern Germany, which are famous for their highly diverse marine fauna preserved in astonishing detail through pyritization, including minute features and labile parts (Bartels and Brassel, Reference Bartels and Brassel1990; Bartels et al. Reference Bartels, Briggs and Brassel1998; Kühl et al. Reference Kühl, Bartels, Briggs and Rust2011; Kühl et al. Reference Kühl, Bartels, Briggs and Rust2012; Südkamp, Reference Südkamp2017; Jansen and Schindler, Reference Jansen, Schindler and Linnemann2025). The slates of the Martelange Member are almost totally devoid of pale laminae of siltstone–sandstone, and this member is therefore particularly uniform. The La Roche (LAR) Formation represents a relatively monotonous transgressive sequence consisting mostly of dark grey or bluish slates, slaty siltstones and shales with occasional bluish sandstone intercalations that increase in thickness and frequency towards the top of the unit (Bultynck and Dejonghe Reference Bultynck and Dejonghe2001; Denayer and Mottequin, Reference Denayer and Mottequin2024). XRD data obtained as part of the Warmifontaine project show that the slates are composed of muscovite (39.50–43.00 wt%), quartz (29.05–31.96 wt%) and chlorite (22.59–31.45 wt%). The deposits formed in fairly calm, deeper water, at the height of the post-Caledonian transgression onto the newly amalgamated continent of Laurussia to the NW, which coincided with maximal extension and deepening of the basin (Goemaere and Dejonghe, Reference Goemaere and Dejonghe2005; Dejonghe, Reference Dejonghe2013). The LAR Formation ranges in thickness from ca 215 m south of Couvin on the southern flank of the Dinant Synclinorium (Godefroid et al. Reference Godefroid, Blieck, Bultynck, Dejonghe, Gerrienne, Hance, Meilliez, Stainier and Steemans1994) to over 1600 m in the Ardenne Anticlinorium (Dejonghe, Reference Dejonghe2012a, b), although faulting complicates establishing the exact vertical extent of the formation; in the western part of the Neufchâteau–Eifel Synclinorium, the minimal thickness of the LAR Formation is estimated at 700 m (ca 300 m for the Martelange Member; Ghysel Reference Ghysel2022; Figure 1). Based on structural considerations, the deposits of the LAR Formation are dated to the late Pragian (Bultynck and Dejonghe, Reference Bultynck and Dejonghe2001; Denayer and Mottequin Reference Denayer and Mottequin2024), but south of Couvin, the occurrence of the conodont Caudicriodus celtibericus Carls and Gandl, Reference Carls and Gandl1969 in the upper part of the Formation suggests an early Emsian age for the top of this lithostratigraphic unit (Carls and Gandl, Reference Carls and Gandl1969). During that time, the basin was located in a southern low, subtropical palaeolatitude (Molyneux et al. Reference Molyneux, Delabroye, Wicander and Servais2013). Only a few biomineralized fossils have been collected from the Martelange Member at Warmifontaine as reflected by the content of the old collections curated at the RBINS, at Liège University (Dewalque, Reference Dewalque1891) and at the Centre Grégoire Fournier of Maredsous Abbey (Mottequin, Reference Mottequin2021). These include bivalves, rugose corals, trilobites and orthocone cephalopods, and indeterminate specimens.
Historically, the Martelange Member of the LAR Formation has been a major source of roof slates, being host to Belgium’s largest slate quarry, the subterranean exploitation of Sainte-Barbe (Asselberghs, Reference Asselberghs1924; Culot, Reference Culot1997; Cnudde, Reference Cnudde1998), located in the small village of Warmifontaine, part of Grapfontaine, and since 1977 subsumed by the larger commune of Neufchâteau in the Province of Luxembourg. At Sainte-Barbe, two economically productive slate levels extending over a distance of ca 900 m were exploited to a vertical depth of approximately 130 m. The main level has a thickness of ca 40 m, whereas the thinner level reaches ca 8 m, both being separated from each other by 3 to 6 m of non-productive shales. Strata have a strike of N 63° E and dip ca 50° to the SE; cleavage subtends only a small angle with the bedding, exhibiting an average direction of ca N 68° E and dip of around 53.5° towards the SE (Asselberghs, Reference Asselberghs1924).
Because the subsurface quarry is currently partially inundated and difficult to access, so far, all recent collecting had to be done from float blocks in the talus at the surface. As a result, sadly, no in situ data on the context or the occurrence of the fossils are currently available. This is particularly unfortunate, considering that a historical specimen from the old collections of Maredsous Abbey shows a preservation significantly superior to any of the recently collected fossils, suggesting that there likely are different levels yielding labile parts at Sainte-Barbe, some of which may have considerably better quality of preservation than others. Recent excavations in the talus at the surface at this locality yielded hundreds of new fossils that are generally difficult to identify. Amongst them are the selkirkiid specimens studied in this paper and two different taxa of echinoderms (Lefebvre et al. Reference Lefebvre, Cincotta, Guériau, Schöder, Olive, Goolaerts and Mottequin2025).
3. Preservation
Both scalidophoran specimens are preserved in faint relief, with the anterior region, including the introvert, surrounded by large euhedral crystals of pyrite. While the tube is essentially preserved with an elemental and mineralogical composition close to but not identical to that of the surrounding sediment, X-ray µCT data (Figs 3, 4, S12, S13) show that the anterior region, associated with the soft parts, is composed of a thin mass of mostly euhedral crystals, which are confirmed by EDS (Figs S1–S5, S9, S10), XRF (Figs S6, S7) and TIMA-X (Figs S8, S11) to be pyritic; the pyritization of IRSNB a13906 appears to be somewhat finer and more framboidal than the coarse crystals observed in IRSNB a13907, and the former is the better preserved of both specimens. This type of preservation closely resembles that of the Hunsrück Slate fossils, where soft parts have been pyritized more delicately through replacement or infilling by a groundmass of <20 µm crystals with occasional larger crystals (Briggs et al. Reference Briggs, Raiswell, Bottrell, Hatfield and Bartels1996; Bartels et al. Reference Bartels, Briggs and Brassel1998). In their investigation of the controls acting on the pyritization of Hunsrück fossils, Briggs et al. (Reference Briggs, Raiswell, Bottrell, Hatfield and Bartels1996) and Bartels et al. (Reference Bartels, Briggs and Brassel1998) postulated that the large euhedral cubes of pyrite occasionally visible on the surface of the fossils, as observed for the present scalidophoran specimens, probably grew at a later stage than the smaller crystals that replicated delicate parts. The matrix directly adjacent to the preserved soft parts is also replete with myriads of minuscule euhedral pyrite cuboids of only a few dozen µm in diameter; in addition, the sediment also contains multiple pyritized burrows and larger ovoid pyritic structures a few mm in diameter. The ubiquitous presence of finely dispersed pyrite crystals throughout the matrix surrounding the specimens may hold a clue to their rather poor preservation, as it indicates that the sediments originally contained relatively large quantities of finely dispersed organic matter that was being consumed by sulphate-reducers. This ready supply of organics in the sediment would have delayed preferential colonization of the carcasses by sulphate-reducing bacteria, precluding the rapid early formation of extensive fine pyritization over the surface of the specimens, but rather promoting the slow growth of coarse, euhedral crystals while allowing significant decay of labile parts to take place.
Complete specimen IRSNB a13906. (a–c) Part. (a) Dry; (b) Under demineralized water; (c) Coated with NH4Cl. (d–f) Counterpart. Specimen is lit from the SW and mirrored to create a false positive relief image for easy comparison with the part. (d) Dry; (e) Under demineralized water; (f) Coated with NH4Cl. (g) Interpretative drawing of IRSNB a13906 combining information from reflected visible light imaging of part and counterpart, elemental maps and X-ray µCT-scans. (h–I) µCT-scans of combined part + counterpart of IRSNB a13906, exposing coarse pyritization formed on soft parts in the anterior and faint pyritization of caudal appendages in the posterior part of the tube. (h) Orthographic projection of the µCT scan of the obverse side. (i) Orthographic projection of the µCT-scan of the reverse side. Scale bars equal 10 mm.

Long description
The image consists of multiple elements: one photo, one diagram, and two illustrations. The photo displays a fossil specimen IRSNB a13906 in different conditions: dry, under demineralized water, and coated with NH4Cl. The diagram provides an interpretative drawing of the specimen, combining information from reflected visible light imaging, elemental maps, and X-ray micro-CT scans. The two illustrations show micro-CT scans of the combined part and counterpart of the specimen, exposing coarse pyritization formed on soft parts in the anterior and faint pyritization of caudal appendages in the posterior part of the tube. The images are overlaid and side-by-side, providing a comprehensive view of the specimen’s structure and preservation.
Partial specimen IRSNB a13907, part. (a) Dry; (b) Under demineralized water; (c) Coated with NH4Cl. (d) Interpretative drawing of IRSNB a13907, part, based on reflected visible light imaging and X-ray µCT-scans. (e–f) µCT-scans of IRSNB a13907, exposing coarse pyritization formed on soft parts in the anterior. (e) Orthographic projection of the µCT scan of the obverse side. (f) Orthographic projection of the µCT-scan of the reverse side. Scale bars equal 10 mm.

Long description
The image displays a fossil specimen labeled IRSNB a13907, part, in three different conditions. The first panel (A) shows the specimen in a dry state, revealing its natural texture and structure. The second panel (B) presents the specimen under demineralized water, highlighting different features due to the water’s effect. The third panel (C) depicts the specimen coated with NH4Cl, which enhances certain details. The fourth panel (D) is an interpretative drawing of the specimen, based on reflected visible light imaging and X-ray micro-CT scans, providing a clear and detailed representation of the fossil’s structure. The drawing includes labels for different parts such as the introvert, trunk, and tube, with specific regions marked as soft parts, soft parts inside the sclerotized tube, and the sclerotized tube itself. The final two panels (E and F) are micro-CT scans of the specimen, showing the coarse pyritization formed on the soft parts in the anterior region. Panel E is an orthographic projection of the micro-CT scan of the obverse side, while Panel F is an orthographic projection of the micro-CT scan of the reverse side. These scans provide a detailed internal view of the fossil, revealing intricate structures and features.
XRF elemental mapping of the part of IRSNB a13906 (Fig. S6, S7) shows that the gross morphology of the soft body of the animal is highlighted by the presence of Pb (enhanced in Fig S7): even though the signal for Pb is very weak, it is clearly confined to the soft parts. Natural pyrites typically contain a myriad of minor and trace elements that are present in the form of inclusions or enter the pyrite lattice via substitution processes; in the case of Pb this occurs mainly in the form of inclusions of other phases, most likely as Pb-bearing minerals such as galena (Abraitis et al. Reference Abraitis, Pattrick and Vaughan2004). While Raman spectroscopy performed on several other Warmifontaine fossils further revealed the presence of graphitic carbon, indicating preservation as pyritized and carbonaceous compression fossils, the EDS data for both IRSNB a13906 and IRSNB a13907 do not show any appreciable presence of C outside of the organic stain flanking IRSNB a13906. In this respect, it must be considered that these specimens spent considerable time at the surface, and as a consequence, any original C may have been removed through weathering; however, in that case, at least some of the pyrite would likely be expected to have been transformed to Fe-oxide and Fe-hydroxide pseudomorphs, which is not observed to any considerable extent; in this respect, it is also of interest to note that the stain next to IRSNB a13906 does show elevated levels of C. If organic matter was originally preserved in these specimens as graphitic carbon, this could hold a clue to the presence and origin of Pb and Pb-bearing mineral inclusions in the fossils: decaying organic matter tends to precipitate/chelate heavy metals such as Pb onto its surface (Osés et al. Reference Osés, Petri, Voltani, Prado, Galante, Rizzutto, Rudnitzki, Da Silva, Rodrigues, Rangel, Sucerquia and Pacheco2017) through microbial action (Labrenz et al. Reference Labrenz, Druschel, Thomsen-Ebert, Gilbert, Welch, Kemner, Logan, Summons, De Stasio, Bond, Lai, Kelly and Banfield2000; Kan et al. Reference Kan, Obraztsova, Wang, Leather, Scheckel, Nealson and Arias-Thode2013), which would have concentrated and made Pb available for the precipitation of Pb-bearing mineral inclusions.
The preservation of the tube and a portion of the soft parts in silicate minerals finds a counterpart in Burgess Shale-type preservation, where the occurrence of various aluminosilicates replicating non-biomineralized morphology was originally considered to represent early diagenetic replication of anatomical features in clay minerals (Orr et al. Reference Orr, Briggs and Kearns1998); later work, however, has shown that this coating is the result of aluminosilicification of originally carbonaceous remains during late greenschist-facies metamorphism (Butterfield et al. Reference Butterfield, Balthasar and Wilson2007, Gaines, Reference Gaines2014). A similar scenario was likely involved in the preservation of the organic selkirkiid tubes and some of the soft parts at Warmifontaine.
4. Materials and methods
Both studied specimens are housed in the Palaeozoic invertebrate palaeontology collections (prefixed IRSNB a) of the Royal Belgian Institute of Natural Sciences (RBINS/KBIN/IRSNB), Brussels, Belgium. Detailed locality data are curated with the specimens.
Image stacks of specimens were photographed dry, coated with ammonium chloride and immersed in demineralized water, under cross-polarized lighting (Bengtson, Reference Bengtson2000; Crabb, Reference Crabb2001) using a Sony α 7R V (ILCE-7RM5) 35 mm full-frame digital mirrorless interchangeable lens camera. Elemental maps were constructed using SEM-EDS on a Tescan Mira 3 SEM with EDAX Element EDS system and synchrotron-µXRF at the PUMA beamline of the SOLEIL synchrotron; mineralogical phase maps were done with TIMA-X (Hrstka et al. Reference Hrstka, Gottlieb, Skála, Breiter and Motl2018) on a Tescan Mira 3 with EDAX Element EDS system. X-ray µCT scans of the specimens were prepared on a Tescan CoreTOM X-ray-µCT-scanner. A detailed discussion of the techniques and equipment employed and of the processing and post-processing of the data is provided in the Supplementary Text.
5. Systematic palaeontology
An extensive discussion and refutation of alternative phylogenetic placements of the Warmifontaine fossils is provided in the Supplementary Text. The terminology for the division of the introvert follows Conway Morris (Reference Conway Morris1977) and Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021). ‘Left’ and ‘right’ refer to these directions in the part.
Institutional abbreviations. IRSNB a, Palaeozoic invertebrate palaeontology collections of the Royal Belgian Institute of Natural Sciences (RBINS / KBIN / IRSNB), Brussels, Belgium.
ECDYSOZOA Aguinaldo et al. Reference Aguinaldo, Turbeville, Linford, Rivera, Garey, Raff and Lake1997
CYCLONEURALIA Ahlrichs, Reference Ahlrichs1995
SCALIDOPHORA Lemburg, Reference Lemburg1995 (= CEPHALORHYNCHA Nielsen, Reference Nielsen1995)
Remarks. Notwithstanding the previously mentioned uncertainty regarding the validity and composition of Scalidophora, until more definite evidence becomes available, we chose to retain this clade, considering that at least a less inclusive grouping containing priapulids and kinorhynchs seems to be well-supported by both morphological and molecular data, and morphological support for Scalidophora is generally strong (Howard et al. Reference Howard, Edgecombe, Shi, Hou and Ma2020).
?PRIAPULIDA Delage and Hérouard, Reference Delage and Hérouard1897
Remarks. Defining priapulid synapomorphies, i.e. the presence of 25 rows of rings of 9 + 8 + 8 scalids, pharyngeal teeth organized in pentagons, organization of the urogenital system, presence of a larval lorica and erythrocytes with haemocyanin (Adrianov and Malakhov, Reference Adrianov and Malakhov1995; Adrianov and Malakhov, Reference Adrianov and Malakhov2001a, b; Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013b), are often difficult or even outright impossible to recognize in fossil material, and for that reason, definite evidence of priapulid, rather than broader scalidophoran, affinities is often lacking for supposed fossil priapulids (see Wang et al. Reference Wang, Vannier, Sun, Yu and Han2023 for a discussion of the problems associated with assigning fossils to Priapulida based on morphological characters). In the case of selkirkiids, recent work by Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021) and Wang (Reference Wang2022) has shown that the scalids in both Selkirkia sinica Luo et al. Reference Luo, Hu, Chen, Zhang and Tao1999 and Selkirkia transita Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021 are arranged into quincunxes, and the introvert likely carries 25 rows of scalids; Yang et al. (Reference Yang, Vannier, Yang, Wang and Zhang2021) and Wang (Reference Wang2022) further showed that the location of gonads in Selkirkia sinica was similar to that in extant meiobenthic priapulids. These findings provide additional morphological support for a bona fide priapulid affinity of this group, and both the parsimony and constrained Bayesian phylogenetic analyses of Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021) and Wang (Reference Wang2022) also do resolve selkirkiids as basal priapulids. In contrast, the unconstrained Bayesian analysis of these authors placed selkirkiids in the scalidophoran stem, leaving open the possibility that at least some of the characters used to unite Priapulida actually are plesiomorphic within Scalidophora. Subsequent analyses including selkirkiids have done little to further solidify the position of this group. Shi et al. (Reference Shi, Howard, Edgecombe, Hou and Ma2021) found Selkirkia sinica to be a stem priapulid in their maximum likelihood and implied weights parsimony trees, but had it in an unresolved position within Ecdysozoa in their 50% majority rule Bayesian inference tree. Smith and Dhungana (Reference Smith and Dhungana2022) in their summary of posterior Bayesian trees recovered Sekirkia sinica at the base of a clade including priapulids, paleoscolecids, nematoids and panarthropods. The analyses of Wang et al. (Reference Wang, Vannier, Sun, Yu and Han2023) resolved Selkirkia sinica as a stem priapulid, except in the 50% majority rule Bayesian inference tree, where it resolved uptree outside of total priapulids, as sister taxon to Eokinorhynchus, in a grade leading up to palaeoscolecids; for low concavity values, the implied weight parsimony trees of Wang et al. (Reference Wang, Vannier, Sun, Yu and Han2023) show Selkirkia sinica as a basal sister taxon to palaeoscolecids, within total-group Priapulida. Liu and Liu (Reference Liu and Liu2025) recovered Selkirkia sinica in the priapulid stem, just below the crown. Therefore, we prefer to err on the side of caution, and until selkirkiids are unequivocally confirmed to belong within Priapulida, we only refer them to this clade provisionally.
SELKIRKIOMORPHA Adrianov and Malakhov, Reference Adrianov and Malakhov1995
Remarks. This clade was proposed as an order containing the family Selkirkiidae by Adrianov and Malakhov (Reference Adrianov and Malakhov1995). Phylogenetic analyses by Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021) and Wang (Reference Wang2022) indicate that it likely consists of selkirkiids, Markuelia Val’kov, Reference Val’kov, Khomentovsky, Yakshin and Karlova1983 and Eokinorhynchus Zhang, Xiao, Liu, Yuan, Wan, Muscente, Shao, Gong and Cao, Reference Zhang, Xiao, Liu, Yuan, Wan, Muscente, Shao, Gong and Cao2015 and is characterized by the similarities in detailed organization of scalids on the basal part of the introvert.
SELKIRKIIDAE Conway Morris, Reference Conway Morris1977
Remarks. Apart from Selkirkia Walcott, Reference Walcott1911a, and Paraselkirkia Hou et al. Reference Hou, Bergström, Wang, Feng and Chen1999, which was synonymized with Selkirkia by Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021), the Selkirkiidae may possibly also include the poorly known Sullulika broenlundi Peel and Willman, Reference Peel and Willman2018 from the upper member of the Cambrian Series 2 Buen Formation of northern Greenland. However, no soft parts are known from S. broenlundi, and some aspects of the tube morphology, like the presence of lappets at the anterior opening, are unknown in undoubted selkirkiids, making its membership of this clade ambiguous.
SELKIRKIA Walcott, Reference Walcott1911a
Selkirkia sp.
Material. IRSNB a13906, a complete specimen consisting of part and counterpart, and IRSNB a13907, an incomplete specimen consisting of the anterior half of the animal, represented only by the part.
Description of IRSNB a13906. This specimen is complete and unbroken and consists of both part and counterpart (Figure 3). The split cracks in the middle through the specimen, following largely the boundaries of coarse peripheral late pyritization present in the anterior half of the specimen. A small part of the posterior of the tube has adhered to the counterpart. The complete specimen has an axial length of ca 37.2 mm.
The shape of the animal is outlined by coarse euhedral pyrite crystals, and in the XRF elemental map of the part is highlighted by a subtle but consistent enrichment in Pb, which is virtually absent from the rest of the specimen and the surrounding matrix (Figs S8, S9); in visible wavelengths, the colour of the specimen is also slightly darker in this area, and X-ray micro-computed tomography (µCT) scans (Figs 3, S12, S13) show that this region is mostly composed of a thin mass of euhedral crystals, identified by EDS, XRF and TIMA-X as being composed of pyrite (Figs S1–S8). Total length of the animal including the introvert is ca 18.5 mm, or about half the length of the complete specimen including the tube. The introvert extends out of the tube and is slightly narrower than the tube opening at its base, having a width of around 3.0 mm. Zone Ia of the introvert is bulbous in shape and expands distally, reaching its greatest width approximately of 3.4 mm in its anterior half. The exposed length of zone Ia is roughly 2.0 mm. Zone Ib appears to be essentially absent and may be expressed by a narrow crease, separating zone Ia from zone Ic. At its base, zone Ic is 1.9 mm wide, tapering distally to a width of ca 1.0 mm. The length of zone Ic is around 0.8 mm. Zone II is roughly parallel-sided, has a length of ca 0.8 mm and a width of around 1.0 mm at its base, tapering slightly distally. The pharynx (zone III) is not everted, corresponding to eversion stage 3 of Conway Morris (Reference Conway Morris1977), which was found to be the most common stage in S. columbia. Although some darker patches on the introvert may be associated with sclerites, no recognizable scalids or spines are preserved. The trunk behind the introvert does not preserve any fine details either, but its outline is squat, with a blunt, straight posterior margin. A short, blunt protuberance emanates from the posterior end. Including this protuberance, but excluding the introvert, the trunk is around 14 mm long. Further down on the right side of the tube, some limited pyritization combined with a trace Pb signature indicates the further presence of more delicate fossilized soft parts, associated with the termination of the trunk (Fig 3, Figs S1–S13).
The tube is ca 34.1 mm in length and has a blunt termination that is around 1.1 mm wide. The anterior opening and widest point of the tube is ca 4.9 mm wide. Its shape is not strictly conical: in the lower third, the sides of the tube are almost subparallel, but over the middle third gradually transition to an opening angle with the vertical of around 8° in the upper third of the tube. No details of ornament are preserved on the surface of the tube. However, in the upper half, there is a vague hint of possible banding, the bands being roughly 0.3 – 0.5 mm wide. These putative bands are exceedingly faint and best seen on the counterpart immersed in demineralized water (Figure 3e).
Approximately halfway down, to the right side, the specimen is associated with a dark stain in the matrix.
Description of IRSNB a13907. An incomplete specimen consisting of only the part, preserving the anterior half of the animal (posterior broken away) and exhibiting extensive, coarse (?late) euhedral pyritization (Figure 4). The specimen is significantly darker than the surrounding matrix, and the highly attenuating phase observed in µCT-data (Figures 4, S13) corresponds with euhedral pyrite, as confirmed by EDS- and TIMA-X-data (Fig. S9–S11). Because of poor preservation, all measurements are approximate. The total preserved axial length is around 18.7 mm.
The introvert appears fully extended (stage 4 of Conway Morris, Reference Conway Morris1977), which was considered a stress reflex by Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021), but the state of preservation complicates recognition of the different zones. Zone Ia is roughly parallel-sided, having an exposed length of around 3.4 mm. At its base, it is ca 5.0 mm wide. Zone Ib again appears to be essentially absent. Due to the coarse pyritization, the exact shape and dimension of zone Ic are difficult to establish; it is approximately 2.0 mm long and 2.9 mm wide at its widest point. A slightly darkened line marks the division between zones II and III. Zone II has a length of around 1.6 mm and greatest width at its base of ca 2.2 mm, while zone III with around 1.7 mm is slightly narrower at its base, having a length of ca 1.2 mm. Zone III tapers to a bluntly rounded tip. No identifiable scalids or spines are preserved.
Due to the incompleteness of the tube, the only meaningful measurements that can be made are the width of the anterior opening, which is ca 5.7 mm, and the opening angle with the vertical of around 8–9°. There is no ornamentation visible on the tube.
Occurrence. Upper Pragian (Lower Devonian) Martelange Member of the La Roche Formation, subterranean Sainte-Barbe slate quarry, Warmifontaine (Grapfontaine, Neufchâteau), Luxembourg Province, Wallonia, Belgium (Figures 1 and 2).
6. Discussion
6.a. Morphology and anatomy
The morphology of the introvert of the Warmifontaine specimens, which essentially lack zone Ib, is most similar to that of Selkirkia columbia Conway Morris, Reference Conway Morris1977. The termination of the trunk in other selkirkiids is unknown, but the squat, blunt shape of the trunk as shown by IRSNB a13906 (Figs 3, S1A, B, S7) is also observed in crown-priapulids and recalls in particular the shape of the diminutive tube-dwelling Maccabeus species. The protrusion at the end of the trunk in IRSNB a13906 likely represents a caudal appendage, and the minor pyritization and Pb concentration noted further back in the tube in both part and counterpart may belong to the distal parts of a pair of appendages. Elongate caudal appendages, which likely function as respiratory organs, are present in the extant macroscopic priapulid genera Priapulopsis Koren and Danielssen, Reference Koren and Danielssen1875, Acanthopriapulus van der Land, Reference van der Land1970 and Priapulus de Lamarck, Reference de Lamarck1816 (Adrianov and Malakhov, Reference Adrianov and Malakhov1995; Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013b), and have also been reported from the early Cambrian Selkirkia sinica Luo et al. Reference Luo, Hu, Chen, Zhang and Tao1999, although they appear stiffer in the Cambrian animals than the soft caudal structures of extant species (Lan et al. Reference Lan, Yang, Hou and Zhang2015; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Wang, Reference Wang2022) and have been speculated to have functioned for anchoring the animal to the substrate (Lan et al. Reference Lan, Yang, Hou and Zhang2015).
The complete tube of IRSNB a13906 is 34.1 mm long, with an opening 4.9 mm wide; assuming the same ratios for the incomplete specimen IRSNB a13907, which has an anterior opening of ca 5.7 mm wide, its tube must have reached approximately 39.7 mm in length. These sizes are within the range recorded for Selkirkia columbia Conway Morris Reference Conway Morris1977 and Selkirkia transita Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021, with Selkirkia tsering Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024 being somewhat smaller, Selkirkia sinica Luo et al. Reference Luo, Hu, Chen, Zhang and Tao1999 and Selkirkia willoughbyi Conway Morris and Robison, Reference Conway Morris and Robison1986 both being considerably shorter, and Selkirkia spencei Resser, Reference Resser1939 coming in significantly larger (Conway Morris, Reference Conway Morris1977; Conway Morris and Robison, Reference Conway Morris and Robison1986; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024). If the very faint possible banding that may be observed in IRSNB a13906 (Figs. 3, S1A, B) is accepted as representing the surface ornament, this would mean the tube had 2–3 bands per mm. This is considerably coarser than in any previously described selkirkiid, with Selkirkia transita (Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021) and Selkirkia tsering (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024) coming closest with 5–9 bands per mm and 6 bands per mm, respectively. However, it must be stressed that the evidence for preserved surface ornament in IRSNB a13906 is tenuous at best. The Warmifontaine selkirkiid is set apart from other selkirkiids by the shape of its tube: while in other members of this clade, the tube has straight margins, in IRSNB a13906 it very gently flares out in its upper half. Considering that the animal is about half as long as the tube in IRSNB a13906, it may be surmised that the upper, widened part of the tube served to shelter the trunk, while the narrow, lower subparallel section of the tube accommodated elongated caudal appendages; this notion may be supported by the limited presence of preserved soft parts lower down in the tube, as shown by the minor pyritization and Pb signature present on the right side of IRSNB a13906 (Figs. S1–S8). It must, however, be pointed out that the strong pyritization in the anterior part and subsequent compaction may also have slightly modified the outline of what was an originally purely conical tube, in which case the minor flaring of the tube noted in the Warmifontaine material would merely represent a taphonomic artefact. The blunt termination of the tube suggests it was open-ended, as has also been shown to be the case in S. columbia, S. sinica and S. transita (Conway Morris, Reference Conway Morris1977; Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Wang, Reference Wang2022). This posterior opening would have allowed the respiratory caudal appendage to be ventilated while the worm resided in its tube and would have facilitated the evacuation of waste products.
The dark stain off the right of the middle part of the tube (Figure 3) is enriched in C, Mg and Fe (Figs. S1–S5) and mainly composed of chlorite and biotite (Fig. S8); it may have originated from organic-rich decay fluids that seeped out of the fractured tube. Some limited soft parts, possibly belonging to the caudal appendages, are spilled out of the tube just below the stain. Similar stains are regularly encountered in fossils from the Burgess Shale, and are particularly common in Marrella splendens (Walcott, Reference Walcott1912; Whittington, Reference Whittington1971).
6.b. Species discrimination in Selkirkia
Discrimination of species in extant priapulids relies for a large part on differences in the detailed arrangement and morphology of scalids and spines, and variations in the caudal region of the trunk (Adrianov and Malakhov, Reference Adrianov and Malakhov1995; Adrianov and Malakhov, Reference Adrianov and Malakhov2001a; Schmidt-Rhaesa, Reference Schmidt-Rhaesa and Schmidt-Rhaesa2013b). Since in most cases the tube in selkirkiids largely obscures any detailed morphology of the trunk, only details of scalids and spines are left as a reliable way to distinguish between different species of Selkirkia. For S. columbia, S. sinica and S. transita, Wang et al. (Reference Wang, Vannier, Aria, Sun and Han2021) and Wang (Reference Wang2022) were able to demonstrate differences in detailed scalid morphology between them, putting these species on firm ground. However, in S. spencei and S. tsering, preservation is too poor to allow any meaningful details on scalid and/or spine morphology to be documented, while S. willoughbyi is known only from isolated tubes. In S. spencei and S. willoughbyi, the authors justified their placement of the fossils in a new species mostly by referring to differences in ornament (or lack thereof) and size of the tube (Resser, Reference Resser1939; Conway Morris and Robison, Reference Conway Morris and Robison1986; Foster et al. Reference Foster, Sroka, Howells, Cothren, Dehler and Hagadorn2022); these are, however, unreliable characters: ornament of the tube may be influenced by environmental factors during life and modified by taphonomy after death, while size will in part depend on the age of individuals, and the environment they live in (e.g. food supply, oxygenation). In the case of S. tsering, the authors used tube ornament and further claimed that the opening angle of the tube was considerably smaller than that for other selkirkiids; while this observation may seem to be correct, it is important to keep in mind that Conway Morris (Reference Conway Morris1977) and Conway Morris and Robison (Reference Conway Morris and Robison1986) showed that S. columbia and S. willoughbyi tubes have a flattened, oval cross-section, resulting in the lateral aspect of the tube varying between almost parallel-sided when viewed from the narrow side, and more widely opening when exposed on the wide side (see also Foster et al. Reference Foster, Sroka, Howells, Cothren, Dehler and Hagadorn2022). In fact, it seems that in the case of S. spencei, S. willoughbyi and S. tsering, the decision of the authors to place them into separate species may have been as much motivated by geographical and temporal considerations as by significant differences in morphology – and Nanglu and Ortega-Hernández (Reference Nanglu and Ortega-Hernández2024) in fact explicitly stated as much for S. tsering. Consequently, the validity of S. spencei, S. willoughbyi and S. tsering is currently relatively poorly supported, and it cannot be excluded that at least some of these species may in fact be synonymous with any of the better documented representatives in the genus. Therefore, while our material may exhibit some differences in tube morphology, we feel that the preservational quality of the available specimens is too poor to allow the erection of a new species, notwithstanding the material’s much younger age compared to the other members of the genus; hence, both specimens from Warmifontaine are left under open nomenclature.
6.c. Clade longevity and ecology
Until the recent description of a selkirkiid from the Early Ordovician Fezouata Biota (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024), selkirkiids were believed to have been restricted to the early and middle Cambrian. While the discovery of Selkirkia tsering in the Fezouata Biota represented a not so surprising extension of the temporal range for the group by some 20 Ma, the Warmifontaine material adds a remarkable further 70 Ma to the longevity of this group. While selkirkiids were long believed to have been some relatively short-lived Cambrian experiment, the Warmifontaine specimens show that the clade existed and thrived for at least 110 Ma, well into the late Palaeozoic. This finding of prolonged ecological success and longevity requires a re-evaluation of the notion (Wang et al. Reference Wang, Vannier, Aria, Sun and Han2021; Wang, Reference Wang2022; Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024) that the penalty in mobility incurred by selkirkiids as a pay-off for the greater protection provided by their sclerotized tubes relatively quickly resulted in them being outcompeted and replaced by more motile but less protected groups. In this regard, it is of significant interest to consider the Hyalinoeciinae, an extant clade of vagrant, epibenthic onuphid polychaetes, and the genus Hyalinoecia Malmgren, Reference Malmgren1867 in particular. Members of Hyalinoecia secrete long, narrow, tapering, slightly curved tubes that are highly sclerotized and composed of onuphic acid and a mucoprotein (Defretin, Reference Defretin, Florkin and Stotz1971); the tubes are closed off at the ends by a unique system of valves. The worms inhabiting the tubes extend out of the front and possess modified anterior parapodia which allow them to engage in caterpillar-like epibenthic crawling, dragging their tubes along with them (Paxton, Reference Paxton1986; Meyer et al. Reference Meyer, Wagner, Ball, Turner, Young and Van Dover2016; Arias and Paxton, Reference Arias and Paxton2022). They have a predominantly predatory and scavenging lifestyle (Meyer et al. Reference Meyer, Wagner, Ball, Turner, Young and Van Dover2016). As such, they provide an almost perfect modern analogue for the extinct selkirkiids. If one considers that the genus Hyalinoecia has a worldwide distribution, ranges from shallow waters to depths of over 2400 m (Paxton, Reference Paxton1986) and contains 22 described species (WoRMS, 2026), it becomes clear that the penalty on mobility imposed by a rigid tube is not sufficient to prevent a vagrant clade from being widespread and highly successful, even in modern marine ecosystems. The presence of selkirkiids in the Early Devonian of Avalonia, which at the time was located at a southern subtropical palaeolatitude, also refutes the earlier suggestion (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024) that by the Early Ordovician, the group had become restricted to high polar latitudes because of increased ecological pressure. However, it was previously suggested (Hearing et al. Reference Hearing, Legg, Botting, Muir, McDermott, Faulkner, Taylor and Brasier2016) that by the Ordovician, ‘Burgess Shale-type’ animals had become displaced from warm, shallow habitats to cold water habitats, either in deeper water settings, or shallow high-latitude environments; deep marine and polar biotas are indeed known to act as refugia and therefore tend to be more conservative (Conway Morris, Reference Conway Morris1989; Wood, Reference Wood1998; Mckinney, Reference Mckinney2003; Gili et al. Reference Gili, Arntz, Palanques, Orejas, Clarke, Dayton, Isla, Teixidó, Rossi and López-González2006; Sallan et al. Reference Sallan, Friedman, Sansom, Bird and Sansom2018), and in this respect, the fact that the Warmifontaine selkirkiids come from a deeper water setting may be significant. Notwithstanding, considering their apparently continued presence at varied latitudes, and the fact that selkirkiids possessed relatively resistant sclerotized tubes, their virtual absence from the post-Miaolingian fossil record, only briefly returning in the late Tremadocian Fezouata Biota (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024) and the late Pragian of Warmifontaine, is baffling; likewise, aside from indeterminate tubes from the Aptian (Lower Cretaceous) of Alexander I Island (Antarctica) which were tentatively compared to Hyalinoecia (Taylor, Reference Taylor1969), it is interesting to note that the analogous onuphid genus Hyalinoecia appears to lack a fossil record entirely, despite the possession of similar sclerotized tubes.
Because none of the Warmifontaine fossils so far have been collected in situ, little can be said about their living environment or detailed mode of life. It seems, however, probable that they lived on and just within the bottom sediments, just like other selkirkiids, as there is nothing in their morphology that would suggest they had adopted a different lifestyle to that of older representatives of the group. Since the La Roche Formation was deposited in deeper water, at a time of maximal deepening of the basin, the Warmifontaine selkirkiids likely adopted an environment and mode of life broadly similar to that of most extant macroscopic priapulids.
6.d. The concept of ‘Cambrian Burgess Shale-type’ organisms
The concept of supposedly typically early and middle Cambrian ‘Burgess Shale-type’ organisms that are somehow clearly distinct from later taxa is pervasive and widely accepted (see e.g. Conway Morris, Reference Conway Morris1989, Reference Conway Morris1998; Gould, Reference Gould1989; Briggs et al. Reference Briggs, Erwin and Collier1994; Van Roy et al. Reference Van Roy, Orr, Botting, Muir, Vinther, Lefebvre, Hariri and Briggs2010, Reference Van Roy, Briggs and Gaines2015; Hearing et al. Reference Hearing, Legg, Botting, Muir, McDermott, Faulkner, Taylor and Brasier2016; Hou et al. Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017). This idea stems from the fact that, because exceptional fossil preservation is particularly common in the early and mid-Cambrian (Allison and Briggs, Reference Allison and Briggs1993b), non-biomineralized organisms and biotas of this age are relatively common. After the mid-Cambrian; however, most of these taxa suddenly seemed to disappear from the fossil record. While there was debate about the actual reasons for the disappearance of these supposedly ‘Cambrian’ clades from the fossil record (see e.g. Aronson, Reference Aronson1992, Reference Aronson1993, Allison and Briggs Reference Allison and Briggs1993a, b; Butterfield, Reference Butterfield1995, Gaines, Reference Gaines2014), it was generally tacitly accepted that most had likely been largely replaced or at least marginalized by more advanced forms by the end of the Cambrian.
The concept of distinct ‘Burgess Shale-type’ organisms is, however, fraught with serious problems. First, there is a basic semantic problem: there is no actual definition of what constitutes a ‘Burgess Shale-type’ organism. Conscious of this, Van Roy et al. (Reference Van Roy, Briggs and Gaines2015) attempted to bypass this issue by referring to ‘Cambrian stem organisms’ instead. This, however, does not solve the matter. Aside from the fact that the phylogenetic position of ancient taxa is often unsettled and hotly debated, whether a fossil organism is considered to belong to the stem or the crown actually depends at least in part on the systematic level one is looking at: e.g. a stem euchelicerate, is, by definition, a crown euarthropod; and a stem arachnid is, by definition, a crown euchelicerate, and hence also a crown euarthropod. The application of the ‘stem’ criterion to define a ‘Burgess Shale-type’ organism is also not applied consistently: e.g. palaeoscolecids, which in most schemes belong to either the scalidophoran or the priapulid stem and which have their greatest diversity in the Cambrian are usually not considered as ‘Burgess Shale-type’ animals, likely in part because they were first described from the Ordovician (Whittard, Reference Whittard1953) and likely in part because they are not a significant component of the actual Burgess Shale fauna (see e.g. Briggs et al. Reference Briggs, Erwin and Collier1994) – even though they are of major importance in other Cambrian biotas (see e.g. Luo et al. Reference Luo, Hu, Chen, Zhang and Tao1999; Maas et al. Reference Maas, Huang, Chen, Waloszek and Braun2007; Hou et al. Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017). Recognizing that it is impossible to formulate a consistent definition for a ‘Burgess Shale-type’ organism, as a corollary, it also becomes problematic to adequately define a ‘Burgess Shale-type’ fauna, although this issue may at least partially be resolved by focusing on ecology and community structure, rather than taxonomic composition in the definition of such biotas.
Second, the discovery of the Fezouata Biota in the Early Ordovician of Morocco (Van Roy, Reference Van Roy2006; Van Roy et al. Reference Van Roy, Orr, Botting, Muir, Vinther, Lefebvre, Hariri and Briggs2010; Van Roy et al. Reference Van Roy, Briggs and Gaines2015; Lefebvre et al. Reference Lefebvre, El Hariri, Lerosey-Aubril, Servais and Van Roy2016), which contains a complete array of ‘Burgess Shale-type’ taxa in co-existence with more ‘modern’ forms (e.g. Van Roy and Tetlie, Reference Van Roy and Tetlie2006; Vinther et al. Reference Vinther, Van Roy and Briggs2008; Van Roy and Briggs, Reference Van Roy and Briggs2011; Van Roy et al. Reference Van Roy, Daley and Briggs2015; Legg, Reference Legg2016a; Ortega-Hernández et al. Reference Ortega-Hernández, Van Roy and Lerosey-Aubril2016; Parry et al. Reference Parry, Edgecombe, Sykes and Vinther2019; Pérez-Peris et al. Reference Pérez-Peris, Laibl, Lustri, Gueriau, Antcliffe, Bath Enright and Daley2021; Potin et al. Reference Potin, Gueriau and Daley2023; Lustri et al. Reference Lustri, Gueriau and Daley2024; Van Roy et al. Reference Van Roy, Richards and Ortega-Hernández2025; Vinther et al. Reference Vinther, Parry, Briggs and Van Roy2017), has incontrovertibly shown that several of these organisms are not limited to the early and mid-Cambrian, but continued to thrive long afterwards. As more and more discoveries are being made in the Ordovician around the globe (e.g. Botting et al. Reference Botting, Muir, Jordan and Upton2015, Reference Botting, Muir, Pates, McCobb, Wallet, Willman, Zhang and Ma2023; Legg and Hearing, Reference Legg and Hearing2015; Legg, Reference Legg2016b; Aris et al. Reference Aris, Corronca, Quinteros and Pardo2017; Pates et al. Reference Pates, Botting, Mccobb and Muir2020, Reference Pates, Botting, Muir and Wolfe2022; Fang et al. Reference Fang, Mao, Liu, Yuan, Chen, Wu, Li, Zhang, Ma, Wang, Zhan, Peng, Zhang and Huang2022; Parry et al. Reference Parry, Briggs, Ran, O’Flynn, Mai, Clark and Liu2024; Liu et al. Reference Liu, Tu, Fan, Xu, Hu, Zong and Gong2025; Van Roy et al. Reference Van Roy, Oumouhou, Zicha, Gaines, Bruthansová and Kraft2025), it becomes ever more evident that ‘Burgess Shale-type’ organisms are a significant and representative part of faunal assemblages at least during the early Palaeozoic and not just part of the Cambrian.
Third, even long before the Fezouata Biota was discovered, it was well known that many ‘Burgess Shale-type’ groups extended well past the middle Cambrian, and indeed, well beyond the Ordovician: hallucigeniid lobopodians occur at least until the Homerian (Thanahita distos Siveter et al. Reference Siveter, Briggs, Siveter, Sutton and Legg2018, Coalbrookdale Formation, Herefordshire Lagerstätte, U.K.) or possibly even the Middle Pennsylvanian (Carbotubulus waloszeki Haug et al. Reference Haug, Mayer, Haug and Briggs2012), Carbondale Formation, Mazon Creek, Illinois, U.S.A.); naraoiids continue until the Přídolí (Naraoia bertiensis Caron et al. Reference Caron, Rudkin and Milliken2004, Fiddlers Green Formation, Canada; Bicknell et al. Reference Bicknell, Goodman, Laibl and Amati2025); the youngest marrellomorphs (Mimetaster hexagonalis Gürich, Reference Gürich1931 and Vachonisia rogeri Lehmann, Reference Lehmann1955) are known from the lower Emsian Hunsrück Slate, Germany (Stürmer and Bergström, Reference Stürmer and Bergström1976; Kühl et al. Reference Kühl, Bergström and Rust2008; Kühl and Rust, Reference Kühl and Rust2010); eldonioids occur up to the upper Frasnian (Paropsonema cryptophya Clarke, Reference Clarke1900, Nunda Sandstone, New York, U.S.A.; MacGabhann, Reference MacGabhann2012; MacGabhann et al. Reference MacGabhann, Schiffbauer, Hagadorn, Van Roy, Lynch, Morrison and Murray2019); and protospongiid and choiid demosponges are even present in the lower Tithonian (Ammonella quadrata Walther, Reference Walther1904, Pfalzpaint Plattenkalk, Germany and Neochoiaella frattigianii Keupp and Schweigert, Reference Keupp and Schweigert2012, Langenaltheim Plattenkalk, Germany). Further Emsian Hunsrück euarthropods of ‘Cambrian’ aspect include Captopodus poschmanni Kühl and Rust, Reference Kühl and Rust2012, Cambronatus brasseli Briggs and Bartels, Reference Briggs and Bartels2001, Wingertshellicus backesi Briggs and Bartels, Reference Briggs and Bartels2001, Magnoculus blindi Briggs and Bartels, Reference Briggs and Bartels2001 and Schinderhannes bartelsi Kühl et al. Reference Kühl, Briggs and Rust2009. If Enaliktidae are accepted as megacheirans (Siveter et al. Reference Siveter, Briggs, Siveter, Sutton, Legg and Joomun2014, but see Aria, Reference Aria2022), this would mean this ‘Cambrian’ clade also extends into the Emsian (Bundenbachiellus giganteus Broili, Reference Broili1930, Hunsrück Slate, Germany; Moore et al. Reference Moore, Briggs, Haven and Bartels2008). With the discoveries from Warmifontaine, selkirkiids can now be added to this already extensive list of long-term ‘Cambrian survivors’.
The idea about the Cambrian uniqueness of ‘Burgess Shale-type’ organisms is an artefact that sprang from the nexus of taphonomy and history. It has long been recognized that the early and mid-Cambrian are unusually rich in exceptional fossil preservation compared to the remainder of the Palaeozoic (Allison and Briggs, Reference Allison and Briggs1993a, Gaines, Reference Gaines2014), a condition which, in part at least, is related to the low sulphate concentrations and more alkaline nature of Cambrian oceans being more conducive to the preservation of labile parts (Gaines et al. Reference Gaines, Hammarlund, Hou, Qi, Gabbott, Zhao, Peng and Canfield2012). As a result of this, the first major exceptionally preserved fauna to be discovered and to receive international acclaim was the Drumian Burgess Shale (Walcott, Reference Walcott1911a, b; Walcott, Reference Walcott1912), which in time lend its name as a general descriptor to similar assemblages and animals from other localities. Because of the uneven temporal distribution of exceptional preservation, the large majority of the exceptionally preserved Palaeozoic organisms discovered in the decades after Walcott’s discovery of the Burgess Shale was also Cambrian in age. Because of a dearth in post-Cambrian exceptionally preserved faunas, this created the artificial impression that the organisms from the early and mid-Cambrian were in some way unique to this time interval. As demonstrated above, multiple discoveries over the past 20 years – and before – have now unequivocally shown this idea to be largely false. As exemplified by the Fezouata Biota (Van Roy et al. Reference Van Roy, Orr, Botting, Muir, Vinther, Lefebvre, Hariri and Briggs2010; Van Roy et al. Reference Van Roy, Briggs and Gaines2015; Richards et al. Reference Richards, Nanglu and Ortega-Hernández2024) and multiple other recent discoveries (Botting et al. Reference Botting, Muir, Jordan and Upton2015, Reference Botting, Muir, Pates, McCobb, Wallet, Willman, Zhang and Ma2023; Legg and Hearing, Reference Legg and Hearing2015; Legg, Reference Legg2016b; Aris et al. Reference Aris, Corronca, Quinteros and Pardo2017; Pates et al. Reference Pates, Botting, Mccobb and Muir2020, Reference Pates, Botting, Muir and Wolfe2022; Fang et al. Reference Fang, Mao, Liu, Yuan, Chen, Wu, Li, Zhang, Ma, Wang, Zhan, Peng, Zhang and Huang2022; Parry et al. Reference Parry, Briggs, Ran, O’Flynn, Mai, Clark and Liu2024; Liu et al. Reference Liu, Tu, Fan, Xu, Hu, Zong and Gong2025; Van Roy et al. Reference Van Roy, Oumouhou, Zicha, Gaines, Bruthansová and Kraft2025; possibly also Saleh et al. Reference Saleh, Lustri, Gueriau, Potin, Pérez-Peris, Laibl, Jamart, Vite, Antcliffe, Daley, Nohejlová, Dupichaud, Schöder, Bérard, Lynch, Drage, Vaucher, Vidal, Monceret, Monceret and Lefebvre2024, but see Muir and Botting, Reference Muir and Botting2024), a significant number of the taxa that first appeared in the early and mid-Cambrian continued to be major constituents of marine biotas into the Ordovician – and at least for some of them, well beyond. They were gradually complemented and replaced by more ‘modern’ forms, but, in a testimony to their long-term evolutionary and ecological success, several of these lineages persisted for over 100 Ma into the Devonian, or even the Jurassic in the case of sponges. Indeed, at this point in time, rather than talking about ‘survival’ or ‘persistence’ of ‘Cambrian Burgess Shale-type’ organisms into the later Palaeozoic, one might dwell with equal merit on the ‘extension’ of ‘Ordovician Fezouata-type’ animals into the Cambrian instead. The concept of uniquely Cambrian ‘Burgess Shale-type’ organisms has historical merit, but it has long since been overtaken by the facts, and it is well past retirement – as again is firmly underscored by the selkirkiids from the Devonian of Warmifontaine.
7. Conclusions
-
• The new discoveries at Warmifontaine add to the almost non-existent post-Miaolingian fossil record of Scalidophora, which is almost exclusively limited to palaeoscolecid scleritomes and sclerites.
-
• The specimens from Warmifontaine show that selkirkiids, the only scalidophoran clade to have developed a tubiculous lifestyle within a strongly sclerotized tube, existed at least until the Pragian, 70 myr longer than hitherto realized. This means Selkirkiidae persisted for at least 110 myr, a testimony to their prolonged evolutionary success.
-
• The subtropical palaeolatitude of Warmifontaine during the Pragian shows that selkirkiids had not become restricted to high polar paleolatitudes by the Early Ordovician as suggested previously (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024), but retained a wider distribution.
-
• Considering their longevity, distribution and relatively easily fossilizable sclerotized tube, it is remarkable that selkirkiids left virtually no record after the Miaolingian (middle Cambrian) save for brief appearances in the Tremadocian (Early Ordovician) subpolar Fezouata Biota of Morocco (Nanglu and Ortega-Hernández, Reference Nanglu and Ortega-Hernández2024) and the Pragian (Early Devonian) subtropical deposits of Warmifontaine, Belgium.
-
• The selkirkiids from Warmifontaine join a long list of supposedly typically Cambrian ‘Burgess Shale-type’ organisms that ‘survived’ into later times. In reality, the concept of ‘Burgess Shale-type’ organisms is semantically unsound as it is impossible to define in any consistent way, and consequently, it is also not applied consistently. More importantly, the entire flawed idea of the ‘uniqueness’ of Cambrian ‘Burgess Shale-type’ organisms stems from the fact that exceptional fossil preservation is considerably more common in the Cambrian than in later periods: as more and more post-Cambrian exceptionally preserved faunas are discovered, it becomes increasingly clear that ‘Burgess Shale-type’ organisms are just a regular component of Palaeozoic marine ecosystems and not unique to the Cambrian in any way. Multiple discoveries over the past decades have shown that the idea of uniquely Cambrian ‘Burgess Shale-type’ organisms is untenable, and it is well past time that this misguided concept is discarded.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0016756826100752.
Repository package
X-ray µCT-reconstructed slices and specimen 3D .STL files are available in the MorphoSource Digital Repository: https://www.morphosource.org/projects/000801048; Synchrotron-µXRF-data are available in the Zenodo Digital Repository: https://zenodo.org/records/20037340; DOI: https://doi.org/10.5281/zenodo.20037340.
Acknowledgements
We are greatly indebted to the Warmifontaine field teams, Mr Herman Borré, the owner of the quarry, who allowed the excavations and Mr Guy Conard, the neighbour of the quarry who provided assistance. As ever, PVR is highly grateful to the staff of the Natural Museum, National Museum in Prague, in particular J. Bruthansová, Petr Daneš, J. Kvaček, V. Turek, M. Valent and J. Wagner for their help, support and generous hospitality during his stays at the museum and use of the outstanding photographic facilities. We thank A. Folie and J. Lalanne (IRSNB) for curatorial support, T. Goevaerts (IRSNB) for X-ray diffraction analyses of the slates and geological preparation, N. Vallée-Gillette (IRSNB) for fossil preparation of Warmifontaine material and E. Goemaere (IRSNB) for fruitful discussions. J. Jurceka (UGent) cut down specimen IRSNB a13907 for µCT scanning. This paper benefitted from constructive reviews by L.A. Parry (Oxford) and an anonymous referee.
Financial support
This work benefited from funding supplied by the Belgian Science Policy Office (Belspo), and in particular of the Belspo-funded B2/202/P1/VERTIGO (AC, SO and BM) project, of which this paper is an official outcome and also is a contribution to the Strategic Research Plan of the Czech Geological Survey (DKRVO/ČGS 2023–2027). The Ghent University Special Research Fund (BOF-UGent) is acknowledged for their support to the UGent Core facility UGCT (BOF.COR.2022.008) and the Research Foundation – Flanders (Fonds Wetenschappelijk Onderzoek, FWO) for the infrastructure grant I013118N to fund the SEM instrumentation. Beamtime at the SOLEIL synchrotron was provided under proposal 20220721. PVR acknowledges funding from Synthesys + grant CZ-TAF-8307 during his stay at the National Museum in Prague in July 2023.
Competing interests
The authors declare that they have no competing interests.






 Open access
Open access