



Summations
-
Longitudinal design enables the study of developmental stages of behaviour.
-
Deeply phenotyped highly representative birth cohort samples help to detect reliable biomarkers and understand their relationships with adaptive strategies.
-
Gene × environment interactions differ gender-wise.
Perspectives
-
Longitudinal design is setting limits to sample size, and hence to more granular investigation of distinct developmental curves. Pooling data of similar studies and replication testing are important for validation of the findings.
-
Environmental variability is impeding accurate reproducibility, unless the salient features in ‘enviromics’ are identified.
-
Adaptive strategies and gender differences are better interpreted in the broader socio-cultural context.
Introduction
Longitudinal studies with deep phenotyping have provided valuable insights into human development and etiopathogenesis of disease, perhaps most importantly for cardiovascular health (Dawber et al., Reference Dawber, Kannel and Lyell1963; Caruana et al., Reference Caruana, Roman, Hernández-Sánchez and Solli2015; Andersson et al., Reference Andersson, Johnson, Benjamin, Levy and Vasan2019). While mental and physical health are often treated as separate categories, the impact of mental health on systemic health and vice versa is difficult to underestimate (Prince et al., Reference Prince, Patel, Saxena, Maj, Maselko, Phillips and Rahman2007). Because of common risk factors, the impacts of health behaviour and shared diathesis mechanisms are indeed inseparable. More recently, several longitudinal studies on patient as well as community cohorts have provided important data on depression and other mental health conditions, their co-morbidities and association with physiological and behavioural risk factors (Luppino et al., Reference Luppino, de Wit, Bouvy, Stijnen, Cuijpers, Penninx and Zitman2010; McKay et al., Reference McKay, Cannon, Chambers, Conroy, Coughlan, Dodd, Healy, O.’Donnell and Clarke2021; Palmese et al., Reference Palmese, Remelli, Dekhtyar, Grande, Marengoni, Calderón-Larrañaga, Domenicali, Volpato, Vetrano and Triolo2025). Herewith we review the procedure and neuropsychiatrically relevant findings of the Estonian Children Personality Behaviour and Health Study (ECPBHS; EstChild), an investigation of highly representative birth cohort samples conducted since 1998. While some results of this study have been reported since 2001 (the list of ECPBHS journal articles and book chapters is available at https://www.tai.ee/et/teadustoo/eliktu-artiklid), the breadth of this study has not yet been comprehensively described. The ECPBHS has collected a highly multidisciplinary dataset, but it is the neuropsychiatric dimension that has received most attention, unveiling several provocative findings.
How the ECPBHS has been conducted: methods and the database
The sample through the study waves
The ECPBHS originates from the European Youth Heart Study (EYHS; e.g., Andersen et al., Reference Andersen, Harro, Sardinha, Froberg, Ekelund, Brage and Anderssen2006); in Estonia it was conducted in 1998 and 1999. Each country participating in the EYHS aimed to include 1000 children, 500 aged 9 and 500 aged 15, but to account for incomplete data, the Estonian team decided to sample 600 + 600 children. The selected age groups reflected key stages of sexual maturation: 9-year-olds were typically prepubertal, while 15-year-olds were in the final stages of puberty (Harro M et al., Reference Harro, Eensoo, Kiive, Merenäkk, Alep, Oreland and Harro2001). The Estonian sample involved a homogeneous sample of Caucasian participants representing both sexes and the main ethnic groups in Estonia (Estonian and Russian), as well as urban and rural populations. The number of children in each category was determined based on the distribution of 9- and 15-year-olds in Tartu County, as per local statistics.
The school was used as the primary sampling unit. Headmasters from 54 of the 56 schools in Tartu County, which included 9- and 15-year-old students, agreed to participate. The minimum expected number of children per age group per school was set at 25. A random sample of 25 schools was drawn using cluster sampling, stratified by location (urban or rural), age group, and language of instruction (Estonian or Russian), with probability proportional to the number of students of the respective age groups in the school. All students in grades 3 and 9 at the selected schools were invited to participate. Of the 1486 invited participants, 79.1% (n = 1176) consented and were enrolled in the study. In 2001, the decision was made to conduct a follow-up of the older cohort, until they still attend their schools. The short preparation period together with the reluctance of schools to grant free time in spring just before exams reduced the number of participants, therefore an additional 62 individuals from other schools were included in the study.
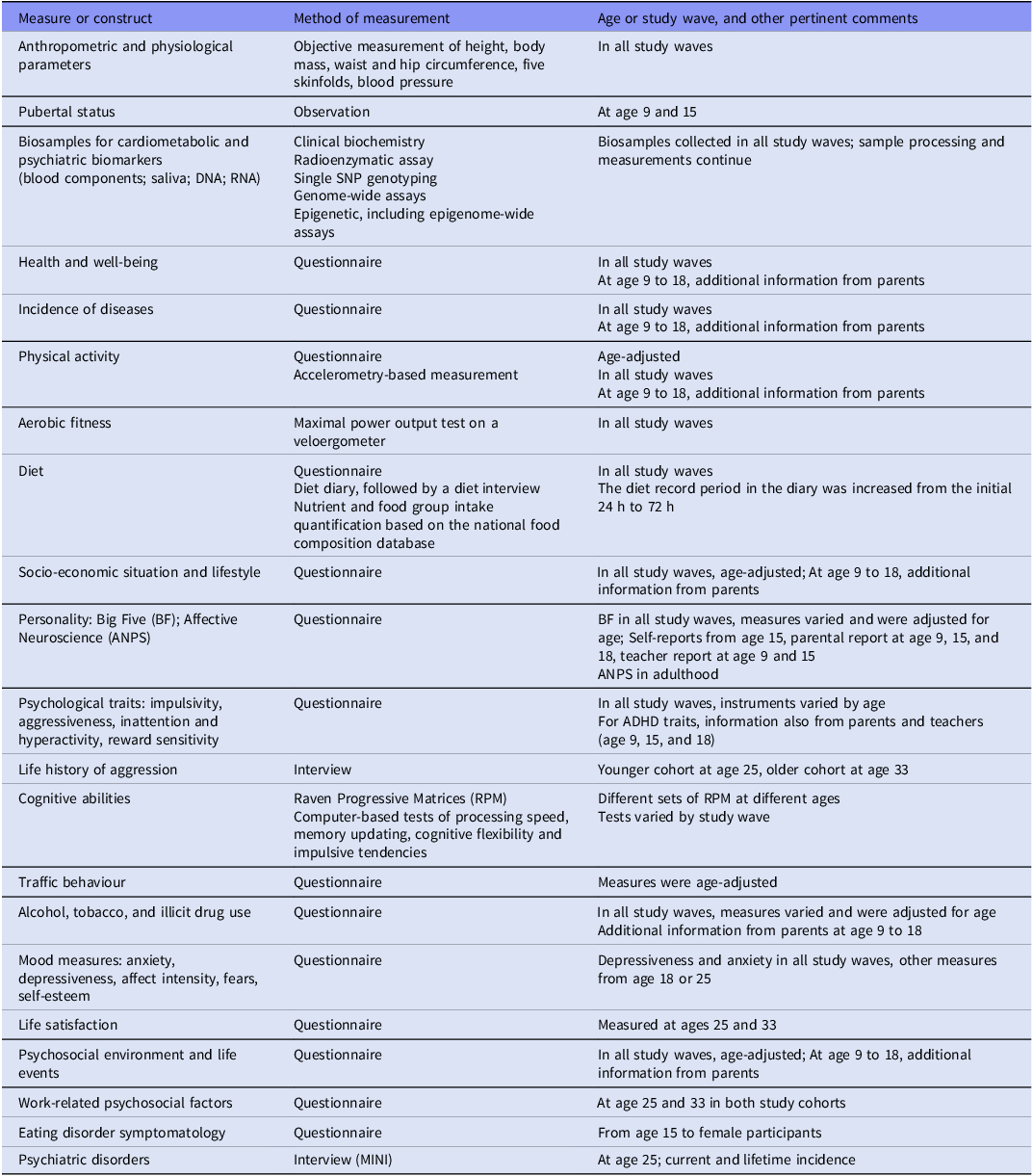
This original sample, consisting of two birth cohorts, was followed longitudinally; all participants have been assessed at ages 15, 18, 25, and 33 (Figure 1).
Timelines of the Estonian Children Personality Behaviour and Health Study portrayed schematically. For more detail, see Table 1.

At ages 15 and 18, participants were mostly still found at the schools they had attended during the first study wave. Those no longer there, an invitation letter was sent to their home address, which was obtained from other participants in the study or the population register, or they were contacted by phone. At ages 25 and 33 (and in the parents’ study wave), home addresses were requested from the Estonian National Registry. Invitation letters were sent to the participants’ electronic and home addresses, collected in former study waves, or from their parents or from other ECPBHS participants. If the letter garnered no response, the participants received a call on their personal mobile phones. Non-responders were subsequently contacted via social media (Orkut or Facebook) or work phone and, before year 2016, home visits were also conducted. These procedures helped to achieve low attrition (Table 1).
The parents of the birth cohort participants were engaged from the outset by completing various questionnaires about their child and family. However, between 2011 and 2013, mothers and fathers were invited to join the study and data were collected about the parents themselves. In total, 1416 parents (581 fathers and 835 mothers, with mean age at the laboratory visit 54.1 ± 6.5 and 52.0 ± 5.8, respectively) were included.
Participation in the data collection waves of the Estonian Children Personality Behaviour and Health Study by birth cohort and gender

n, number of participants; M, male; F, female; % - proportion of the total sample at follow-ups; S.D., standard deviation.
The total number of the ECPBHS parents is 1416: six fathers and five mothers have a child in both birth cohort samples. The numbers in the table represent the child-associated information on parents themselves.
Data collection procedure
Children were picked up at their school in the morning and taken to the laboratory. Adults came on their own. Typically, fasting participants arrived between 8:00 and 9:00 a.m. They were instructed not to eat or drink that morning. First, blood samples were collected. After a light breakfast blood pressure was measured (before drinking coffee), anthropometric measurements were taken and an aerobic fitness test was conducted. Participants completed questionnaires, attended the diet interview and performed computer tasks in a variable order.
When the participants were 9, 15 or 18 years old, a parental questionnaire consisting of three sections (A, B, and C) was administered. Section A was completed by a parent with information on the child’s lifestyle, health and risk behaviours. Section B was completed by the mother, and Section C by the father. These sections included questions concerning the parents’ health, education, occupation, and living conditions, as well as the child’s health and personality. A few psychometric instruments were also administered to class teachers. In 2011–2013, the parents themselves completed the core set of the questionnaires administered to their children as adults. They also took part in the dietary assessment, measurement of anthropometric and physiological parameters and physical activity, and biosamples were collected.
At all study waves, participants completed a self-administered questionnaire concerning their living conditions, mental and physical health, socioeconomic background, and trusted persons. The questionnaire also included sections on risk behaviours and physical activity. Several additional instruments were used to assess personality and behavioural characteristics.
The set of instruments used for data collection expanded over the years and, although data on key indicators were gathered in every wave, some variation of content exists between the waves, owing to involvement in European research consortia with their specific objectives, or the conceptual evolution among the EstChild team. The volume of questionnaires thus became burdensome to be completed during the laboratory session. Consequently, the material was divided into three packages A, B, and C: Package A was mailed to the participants and completed prior to the laboratory visit, B during the visit, and C was taken home and returned subsequently together with the accelerometer, that had been distributed during the laboratory visit.
Measures and biosamples collected from the ECPBHS target subjects are listed in Table 2, and detailed information on instruments is provided in Supplementary Tables 1 and 2. Parents of the ECPBHS children provided material on all the main constructs, with the exception that aerobic fitness was not assessed (Supplementary Table 3).
Data collected in the Estonian Children Personality Behaviour and Health Study since 1998. More detail on the measures, instruments and equipment used in each study wave can be found in Supplementary Tables 1 and 2. Most of these measures and constructs were also addressed in the parents’ data collection wave (Supplementary Table 3)
Participation in the study was voluntary in each wave, and participants were allowed to refuse any procedure they found uncomfortable. Written informed consent was obtained from all participants prior to enrolment; in the case of minors, consent was also provided by their parents. The study was conducted in accordance with the Declaration of Helsinki. Ethical approval for each study wave was granted by the Ethics Review Committee on Human Research of the University of Tartu.
Anthropometry, and physical health objectively and subjectively
Height was measured with a stadiometer (Tanita HR 001; Amsterdam, TANITA Europe B.V.) to the nearest 0.5 cm, body mass was initially measured using a calibrated beam balance, later with an electronic scale with the subject in light clothing to the nearest 0.1 kg, and body composition assessed (Omron BF300, Kyoto, OMRON Matsusaka Co. Ltd., or BC-420MA; Amsterdam, TANITA Europe B.V.). Waist and hip circumferences were measured with a metal anthropometric tape. Waist circumference was measured midway between the lower rib margin and the iliac crest at the end of gentle expiration. Hip circumference was measured as the largest circumference of the buttocks. Skinfolds were measured on biceps brachii, triceps brachii, subscapular, suprailiac, and medial calf with a Harpenden calliper (Baty International, Burguess Hill, U.K.) (Lohman et al., Reference Lohman, Roche and Martorell1991). All measurements were taken twice, and the mean value was calculated (Ortega et al., Reference Ortega, Ruiz and Sjöstrom2007). Based on anthropometric indicators, body mass index (BMI = body weight/height2 [kg/m2], waist-to-hip ratio (WHR) and the sum of five skinfold thicknesses were calculated.
Resting blood pressure was measured on the upper arm with an automatic blood pressure monitor (Dinamap Compact; Critikon L.L.C. Tampa, FL, U.S.A.) five times at 2-min intervals, from which the average systolic and diastolic blood pressure values were calculated. In the original data collection wave, lung function was assessed with the Vitalograph 2170 (Vitalograph Ltd. Maides Moreton, Buckingham, England) spirometer.
The pubertal stage was assessed in a private room by an experienced medical researcher of respective gender, following a brief observation, in 9- and 15-year-old subjects (Tanner & Whitehouse, Reference Tanner and Whitehouse1976).
As an indicator of subjective physical and psychological health, and general well-being, a measure of self-rated health was taken on a 5-point scale (Pärna & Ringmets, Reference Pärna2011) health questionnaire also recorded the incidence of acute diseases, accidents, injuries, and health complaints, as well as lifetime chronic conditions.
Physical activity and aerobic fitness
Physical activity was assessed both by structured questionnaires and objectively by using accelerometry (Kiive et al., Reference Kiive, Katus, Eensoo, Villa, Mäestu, Veidebaum and Harro2025). Participants wore the accelerometer (Caltrac, Hemokinetics, Madison, WI, U.S.A. or ActiGraph GT1M or GT3X, Monrovia, CA, U.S.A.) on their right hip. Reliable accelerometer data required at least eight hours of daily recording for a minimum of four days, including two weekend days. The average data from the recorded days was calculated for total physical activity (counts per minute), sedentary time, and light, moderate, and vigorous activity. Additional information concerning the training status, participating in training groups and attitude towards training, was also recorded. In addition to self-assessment, parents also evaluated their child´s physical activity via questionnaire.
Aerobic fitness was assessed using a cycle ergometer with progressively increasing load until exhaustion (Muntaner-Mas et al., Reference Muntaner-Mas, Ortega, Femia, Kiive, Eensoo, Mäestu, Franke, Reif, Faraone and Harro2021). In the earlier study waves, the Monark 839E (Monark Exercise, Vansbro, Sweden) was used, while later waves utilised the Tunturi T8 (Tunturi OY, Turku, Finland) or Ergoselect 100 (Ergoline Gmbh, Bitz, Germany). Each test began with a 3-min warm-up. In the initial study wave, the starting work rate was set at either 20 or 25 W, depending on whether the subjects weighed less than or more than 30 kg, respectively. In subsequent waves, the starting work rate for females was set at 40 W with subsequent increases of 40 W, while for males, the initial workload was 50 W, increasing by 50 W. Throughout all study waves, each workload was maintained for three minutes. The bike ergometer allowed participants to choose a comfortable pedalling cadence within a range of 60 to 80 revolutions per minute (rpm). Heart rate (HR) was continuously monitored during the test using a Polar Vantage HR monitor (Polar Electro, Kempele, Finland). Criteria for exhaustion were defined as either a heart rate exceeding 185 beats per minute, the ability to maintain a pedalling rate of at least 30 rpm, or the subjective judgement of the observer. Aerobic fitness was calculated as maximal work output (Wmax) and further adjusted for body weight (Wmax/kg). Maximal power output was determined for each individual using the formula: W1 + (W2 × t/180), where W1 is the work rate at the fully completed stage, W2 is the work rate increased at the final incomplete stage, and t is the time in seconds at the final incomplete stage. After exercise, participants rated their perceived exertion (Borg, Reference Borg1998).
Diet diary and interview
Dietary assessment included questionnaires, diary and an interview (Matrov et al., Reference Matrov, Kurrikoff, Villa, Sakala, Pulver, Veidebaum, Shimmo and Harro2022). Participants were instructed to complete a food record at home prior to the study day (covering a period from 24 to 72 h, as the study progressed). For participants in the younger cohort, who were initially 9 years old, parental assistance was provided when necessary to ensure accurate reporting.
On the study day, a face-to-face interactive interview was conducted. The food record data were validated against the interview responses, and any discrepancies were discussed with the participants. Portion sizes not specified in the food records were estimated using pictures of portion sizes (Haapa et al., Reference Haapa, Toponen, Pietinen and Räsänen1985). Dietary intake over several days was averaged to calculate the mean food consumption.
Nutrient and food group intake was analysed using the Finnish Micro-Nutrica Nutritional Analysis platform, adapted to include Estonian foods, and the evidence-based food composition Estonian database NutriData, maintained by the National Institute for Health Development (Tallinn, Estonia). A food frequency questionnaire was completed by the participants at home beforehand and brought with them on the study day. This questionnaire addressed the frequency of consumption of certain food groups and food items, including e.g., caffeine.
Questionnaires on socio-economic situation and lifestyle
Questions on socioeconomic background, living conditions and lifestyle varied across study waves; they were age-appropriate and became more specific as participants grew older. For participants aged 9–18, parents helped to provide information. For example, the questionnaire for participants aged 9–18 included sections on school-related topics, such as the relationships with teachers and classmates, academic progress, and the overall school climate (Vaht et al., Reference Vaht, Kurrikoff, Laas, Veidebaum and Harro2016). It included questions about eating habits and attitudes, physical activity, sports participation, and commuting (e.g., to school). Several items addressed hobbies, friends and pets, television viewing and video gaming habits, and, in later waves, internet use. Parental education level, household income, living conditions, and perceived socioeconomic status (SES) compared to peers (ranging from ‘poor’ to ‘among the wealthiest in the country’) were queried (excluded at age 9).
At age 25 and 33 the cohorts faced questions about the level of education, total household income, and self-reported SES assessment compared to peers, living conditions, parenthood, and occupational classification.
Personality assessment: comprehensive models
Personality data based on the Five-Factor Model (Neuroticism, Extraversion, Openness, Agreeableness, and Conscientiousness) were reported at ages 9, 15, and 18 by the mother and the father, and from age 15 also self-reported, using a variety of instruments: the Estonian Brief Big Five Inventory (Laidra et al., Reference Laidra, Allik, Harro, Merenäkk and Harro2006), the Estonian version of the NEO-PI-R (Kallasmaa et al., Reference Kallasmaa, Allik, Realo and McCrae2000), the Short Five (Konstabel et al., Reference Konstabel, Lönnqvist, Walkowitz, Konstabel and Verkasalo2012) or, most commonly and in the most recent waves, the semantically simplified EE.PIP-NEO (Mõttus et al., Reference Mõttus, Pullmann and Allik2006).
The more recent data collection waves used an adaptation (Harro et al., Reference Harro, Laas, Eensoo, Kurrikoff, Sakala, Vaht, Parik, Mäestu and Veidebaum2019) of the short version of the Affective Neuroscience Personality Scale ANPS (Davis et al., Reference Davis, Panksepp and Normansell2003), which is a self-report instrument constructed bottom up to correspond to the activity in neural circuits underlying basic emotive systems as defined in animal research (Panksepp, Reference Panksepp1998). It comprises scales termed ANGER, FEAR, SADNESS, SEEKING, CARE, and PLAY.
Psychological traits: impulsivity, aggressiveness, attention deficit, hyperactivity, reward sensitivity
Hyperactive behaviour and concentration difficulties were assessed from age 9 by class teachers and parents (Kiive et al., Reference Kiive, Kurrikoff, Mäestu and Harro2010) using the 7-point Hyperactivity Scale (af Klinteberg & Oreland, Reference af Klinteberg and Oreland1995). In addition, data on ADHD symptoms in adolescents were collected using teacher- and parent-rated Swanson, Nolan and Pelham Questionnaire IV (SNAP-IV; Swanson, Reference Swanson1992). As young adults, participants filled in the World Health Organisation Adult ADHD Self-Report Scale symptom checklist (ASRS; Kessler et al., Reference Kessler, Adler, Ames, Demler, Faraone, Hiripi, Howes, Jin, Secnik, Spencer, Ustun and Walters2005). Because the wording of the ASRS items is linguistically complex, the ADHD symptomatology of both cohorts at age 33 was also assessed using a measure specifically constructed for this purpose, featuring simple and easily understandable statements (Simple ADHD). Furthermore, at age 33, inattention and concentration difficulties were assessed using the Attention Control Scale (ATTC; Derryberry & Reed, Reference Derryberry and Reed2002), a self-report measure evaluating two key components of attention: focusing and shifting.
Facets of impulsivity (Fast Decision Making, Excitement Seeking, Disinhibition, and Thoughtlessness) were assessed by self-report from ages 15 to 33 using the Adaptive and Maladaptive Impulsivity Scale (AMIS). The AMIS is an original composite instrument based on the Dickman Impulsivity Inventory (Dickman, Reference Dickman1990) and impulsivity-related subscales of the NEO Personality Inventory. Fast Decision Making and Excitement Seeking can be combined to form the Adaptive Impulsivity score, whereas Disinhibition and Thoughtlessness comprise Maladaptive Impulsivity (Paaver et al., Reference Paaver, Eensoo, Pulver and Harro2006). In addition, the Barratt Impulsiveness Scale, 11th version (BIS-11; Patton et al., Reference Patton, Stanford and Barratt1995), was employed.
Aggressive behaviour was assessed across the study. At ages 9, 15, and 18, class teachers who had known the child for at least three years rated aggression using the Aggressiveness subscale of the 7-point Hyperactivity Scale (af Klinteberg & Oreland, Reference af Klinteberg and Oreland1995). In adulthood, aggression was self-reported using the Buss-Perry Aggression Questionnaire (BPAQ; Buss & Perry, Reference Buss and Perry1992). BPAQ specifies Physical Aggression, Verbal Aggression, Anger, and Hostility. Additionally, the Aggressive Provocation Questionnaire (APQ; O’Connor et al., Reference O’Connor, Archer and Wu2001), based on Frijda’s modular theory of emotions (Reference Frijda1988; Reference Frijda2017), was used to assess aggressive tendencies in provided real-life situations.
The Illinois Bully Scale (IBS; Espelage & Holt, Reference Espelage and Holt2001) comprises three dimensions (Bully, Fight, and Victim) and, although originally designed for 8–18-year-olds, was used here as a retrospective measure. Participants also completed peer ratings of classmates they remembered well; scores were averaged when multiple ratings were available. One original instrument (KVLS-KAKK) was designed for the purpose of research on types of aggressiveness less studied in the general population, but it has not yet been properly described.
Reward sensitivity was assessed using the original Reward Openness and Insatiability Scale (ROIS; Pulver et al., Reference Pulver, Kiive and Harro2020a) that will be discussed below.
Cognitive abilities and computer-based tasks
As a paper and pencil test, Raven’s Standard Progressive Matrices (RPM) (Raven, Reference Raven, Raven and Court1998), standardised on the Estonian population (Lynn et al., Reference Lynn, Allik, Pullmann and Laidra2002), was used to measure the nonverbal intellectual abilities. No time limit was applied.
In specific study waves, a variety of computer-based experiments were conducted to measure processing speed, memory updating, cognitive flexibility and impulsive tendencies in the participants. These included the Visual Comparison Test (VCT; based on Dickman & Meyer, Reference Dickman and Meyer1988; Paaver et al., Reference Paaver, Nordquist, Parik, Harro, Oreland and Harro2007), the Wisconsin Card Sorting Test (WCST; Grant & Berg, Reference Grant and Berg1948), the Stop Signal Task (SST; based on Logan et al., Reference Logan, Schachar and Tannock1997; Reif et al., Reference Reif, Kiive, Kurrikoff, Paaver, Herterich, Konstabel, Tulviste, Lesch and Harro2011), a 2-back working-memory updating task using simple schematic facial expressions (Tamm et al., Reference Tamm, Kreegipuu and Harro2021), the CogShift task, originally based on the attention-switching paradigm described by Ravizza and Ciranni (Reference Ravizza and Ciranni2002), and a visual discrimination task modelled on Forster and Lavie’s low-load paradigm (Forster & Lavie, Reference Forster and Lavie2015; Tuvi et al., Reference Tuvi, Harro, Kiive, Vaht and Bachmann2020).
Behaviour in traffic
The traffic safety questionnaire (Eensoo et al., Reference Eensoo, Harro, Pullmann, Allik and Harro2007; Luht et al., Reference Luht, Eensoo, Tooding and Harro2018) was introduced in year 2004 and was age adjusted (e.g., schoolchildren were questioned about ‘frequency of using pedestrian crossings on the way to school’, and young adults about ‘using a mobile phone while driving’).
Participants who had driven a car in the year prior to the study have completed the Driver Behaviour Questionnaire (DBQ, Reason et al., Reference Reason, Manstead, Stradling, Baxter and Campbell1990; Tokko et al., Reference Tokko, Miškinyte, Eensoo and Harro2022), Driver Anger Scale (DAS, Deffenbacher et al., Reference Deffenbacher, Oetting and Lynch1994; Tokko et al., Reference Tokko, Eensoo, Harro, Martin, Preedy and Patel2023) and Driver Skill Inventory (DSI, Lajunen & Summala, Reference Lajunen and Summala1995; Eensoo et al., Reference Eensoo, Paaver and Harro2010). The majority of participants have consented to anonymized accession to their objective traffic records.
Substance use
Alcohol and tobacco use were self-reported in all study waves. The questionnaires addressed, for example, the age of initiation, quantity, and frequency of alcohol and tobacco use, as well as the preferred type of alcohol or tobacco product, and were adapted to be age-appropriate. At age 33, the Alcohol Use Disorders Identification Test (AUDIT, Saunders et al., Reference Saunders, Aasland, Babor, De La Fuente and Grant1993) was applied, and the Fagerström test (Heatherton, Reference Heatherton, Kozlowski, Frecker and Fagerström1991) to detect possible nicotine dependence.
Mood measures
The State–Trait Anxiety Inventory subscales (STAI-S and STAI-T; Spielberger et al., Reference Spielberger, Gorsuch, Lushene, Vagg and Jacobs1983) has been administered since age 15. Depressive symptoms were assessed using the Montgomery-Åsberg Depression Rating Scale (Montgomery & Åsberg, Reference Montgomery and Åsberg1979) and the Beck Depression Inventory (Beck et al., Reference Beck, Ward, Mendelson, Mock and Erbaugh1961).
We compiled a list of fears that included items known to be common sources of phobias and items that had been described as the most common fears in adolescents and youth from previous literature. The respondents were asked to assess how much they fear 18 items on a 5-point scale ranging from 0 = not scared to 4 = very scared (Tulviste et al., Reference Tulviste, Kiive, Akkermann and Harro2015).
The Rosenberg Self-Esteem Scale (Rosenberg, Reference Rosenberg1965; Pullmann & Allik, Reference Pullmann and Allik2000) assessed an overall positive or negative attitude toward oneself. Global cognitive evaluation of life satisfaction was made with the Satisfaction with Life Scale (Diener et al., Reference Diener, Emmons, Larsen and Griffin1985).
Psychosocial environment and life events
At its beginnings, ECPBHS applied a questionnaire used by sociologists of childhood. This served as the source of the development of the Tartu Family Relationships Scale (TFRS; Paaver et al., Reference Paaver, Kurrikoff, Nordquist, Oreland and Harro2008). The TFRS measures four dimensions of family functioning: closeness, support, misprise, and abuse, which can be combined into two higher-order scales: warmth (Closeness and Support) and maltreatment (Misprise and Abuse). In two study waves, the Parental Acceptance–Rejection/Control Questionnaire (PARQ/Control; Tulviste & Rohner, Reference Tulviste and Rohner2010) was used for mother–child relationship, and when the adolescents reached adulthood, at ages 25 and 33, they were asked about their current relationships with their mothers and fathers. In addition, 25- and 33-year-olds who reported being in a stable cohabiting romantic relationship, either currently or in the past, described their relationship with their partner. For this purpose, an instrument based on the TFRS was used, with the questions adapted to assess a partner relationship.
History of stressful life events (SLE) was self-reported by participants (Laas et al., Reference Laas, Reif, Kiive, Domschke, Lesch, Veidebaum and Harro2014b). The list of adverse life events varied across study waves and consisted of 10–17 (dependent on the study wave) stressful events including parental death, parental divorce or separation, absence of both parents, familial unemployment, financial difficulties and poverty, poor living conditions, poor health and chronic diseases, serious illness of a family member, death of a close relative, trauma, fear of school, bullying at school and humiliation at home. At ages 25 and 33, recent stressful life events encountered in the five preceding years were self-assessed. The checklist consisted of 14 personal SLE-s involving divorce/termination of any romantic relationship, separation from a significant other, serious conflicts with husband/wife/partner, having been assaulted, major financial trouble, difficulties with residence, serious illness or injury, being laid off or sacked from work, trouble with the law, having been robbed, serious problems at work, birth of a child, traffic accident, or other accident considered serious. Data were also collected on SLE-s related to significant other/family members, such as conflicts with significant other/family member and personal crisis, death or serious illness of significant other/family member. The events were recorded as dichotomous variables (present or absent) and then counted to determine the number of experienced adverse life events.
In adulthood, the Copenhagen Psychosocial Questionnaire (COPSOQ; Kristensen et al., Reference Kristensen, Hannerz, Høgh and Borg2005) medium-length version was used to assess psychosocial factors at work, stress, and well-being.
Disordered eating
Eating behaviour and attitudes were assessed in female participants from age 15 onward by the Eating Disorders Inventory-2 (EDI-2; Garner, Reference Garner1991; Podar et al., Reference Podar, Hannus and Allik1999), a self-report questionnaire designed to assess eating disorder psychopathology.
Clinical interview for lifetime incidence of psychiatric disorders and aggressiveness
The Mini-International Neuropsychiatric Interview (M.I.N.I.5.0.0; Sheehan et al., Reference Sheehan, Lecrubier, Sheehan, Amorim, Janavs, Weiller, Hergueta, Baker and Dunbar1998; Estonian version Shlik et al., Reference Shlik, Aluoja and Kihl1999) was used to screen for current and lifetime psychiatric disorders. M.I.N.I. 5.0.0 is a structured psychiatric interview developed to diagnose DSM-IV and ICD-10 mental disorders. A diagnostic assessment was conducted by experienced clinical psychologists in both study cohorts at the age of 25.
An interview of Life History of Aggression (LHA) was conducted by an experienced clinical psychologist. The aggression subscale of this measurement tool for lifetime aggressive behaviour (Coccaro et al., Reference Coccaro, Berman and Kavoussi1997) was used to assess dimensions of aggression, including adult frequency of temper tantrums, general fighting, specific physical assault, specific property assault, and verbal assault.
Biosamples, biochemical measures and genotyping
Biosamples were collected at each study wave, prepared for a variety of assays, and have been stored in a biobank. Cardiovascular risk indicators, including triglycerides, cholesterol, HDL-cholesterol, and LDL-cholesterol, have been measured in an accredited laboratory. Platelet monoamine oxidase has been measured radioenzymatically (Harro M et al., Reference Harro, Eensoo, Kiive, Merenäkk, Alep, Oreland and Harro2001). Several hypothesis-based and hypothesis-free genetic and DNA methylation analyses have been conducted (see the following sections). As expected from such a sample, each genotype has been in Hardy–Weinberg equilibrium.
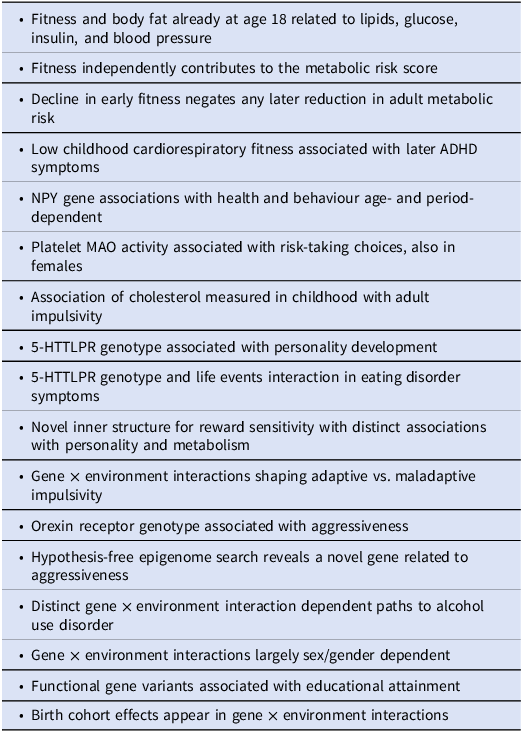
The perspective-opening findings that have energed from the ECPBHS are summarized in Table 3. Below these results have been put into context.
A selection of perspective-opening findings that have emerged in the ECPBHS/EstChild and are briefly discussed in the following sections
Cardiovascular risk factors in ECPBHS and their relevance to neuroscience
Physical fitness, physical activity, and metabolic health are closely related. Low cardiorespiratory fitness is associated with insulin resistance, poor lipid metabolism, and excess body fat (Ortega et al., Reference Ortega, Cadenas-Sanchez, Lee, Ruiz, Blair and Sui2018). Conversely, metabolic dysfunction can impair exercise capacity and diminish the body’s ability to sustain cardiovascular fitness. Cardiorespiratory fitness refers to the capability of the circulatory and respiratory systems to supply oxygen to skeletal muscles for energy production during physical activity (Raghuveer et al., Reference Raghuveer, Hartz, Lubans, Takken, Wiltz, Mietus-Snyder, Perak, Baker-Smith, Pietris and Edwards2020). Research indicates that metabolic risk factors can track from childhood and adolescence into adulthood (Juhola et al., Reference Juhola, Magnussen, Viikari, Kähönen, Hutri-Kähönen, Jula, Lehtimäki, Åkerblom, Pietikäinen, Laitinen, Jokinen, Taittonen, Raitakari and Juonala2011). Furthermore, evidence suggests that being physically fit, regardless of body fat levels, can positively influence metabolic markers in young adults (Sacheck et al., Reference Sacheck, Kuder and Economos2010; Tur-Boned et al., Reference Tur-Boned, Andersen, López-Bueno, Núñez-Cortés, Cruz-Montecinos, Suso-Martí, Polo-López and Calatayud2025).
In the ECPBHS birth cohorts, both fitness and body fat were related already at age 18 to blood lipids, glucose levels, insulin, and blood pressure, and fitness independently contributed to the metabolic risk score (Lätt et al., Reference Lätt, Jürimäe, Harro, Loit and Mäestu2018). However, high cardiorespiratory fitness in adolescence, if followed by a decline in fitness during adulthood, does not yield benefits compared to consistently low cardiovascular fitness from age 15 to 33, while improving cardiorespiratory fitness from adolescence to adulthood can lower metabolic risk (Mäestu et al., Reference Mäestu, Harro, Veidebaum, Kurrikoff, Jürimäe and Mäestu2020).
Metabolic disturbances are characteristic of a variety of neuropsychiatric disorders, and physical activity has successfully served as an adjunctive treatment (Vancampfort et al., Reference Vancampfort, Firth, Stubbs, Schuch, Rosenbaum, Hallgren, Deenik, Ward, Mugisha, Van Damme and Werneck2025). Attention-deficit/hyperactivity disorder (ADHD) is a neurodevelopmental disorder that begins in childhood and often persists into adulthood. It has been associated with obesity (Cortese & Tessari, Reference Cortese and Tessari2017), diabetes and hypertension (Chen et al., Reference Chen, Hartman, Haavik, Harro, Klungsøyr, Hegvik, Wanders, Ottosen, Dalsgaard, Faraone and Larsson2018). Different aspects of physical fitness are associated with ADHD in young adults (Jeoung, Reference Jeoung2014) and interventions aimed at increasing physical activity have been shown to improve attention and reduce impulsivity in individuals with ADHD (Vancampfort et al., Reference Vancampfort, Firth, Stubbs, Schuch, Rosenbaum, Hallgren, Deenik, Ward, Mugisha, Van Damme and Werneck2025). The ECPBHS was the first study showing with longitudinal design that lower cardiorespiratory fitness in childhood is associated with increased, indeed doubling, ADHD symptoms during adolescence, even after adjusting for baseline ADHD symptoms (Muntaner-Mas et al., Reference Muntaner-Mas, Ortega, Femia, Kiive, Eensoo, Mäestu, Franke, Reif, Faraone and Harro2021). This association has recently been confirmed at the diagnostic level in a large national sample of 1.9 million participants in Taiwan (Chiang et al., Reference Chiang, Chuang, Chen, Hsu, Ho, Hsu, Sheu, Gau and Liang2024).
An intriguing case of apparent non-association with body composition and metabolic function is presented by the neuropeptide Y (NPY) gene. NPY, a 36-amino acid neuropeptide (Tatemoto et al., Reference Tatemoto, Carlquist and Mutt1982), much studied in the context of anxiety, addiction and depression (Heilig & Widerlöw, Reference Heilig and Widerlöv1995; Wu et al., Reference Wu, Feder, Wegener, Bailey, Saxena, Charney and Mathé2011; Thorsell & Mathé, Reference Thorsell and Mathé2017), is a most potent orexigenic stimulus (Kokot & Ficek, Reference Kokot and Ficek1999) that leads to an increase in fat mass and body weight (Stanley et al., Reference Stanley, Kyrkouli, Lampert and Leibowitz1986). Also, NPY expression is increased in adipose subjects (Sitticharoon et al., Reference Sitticharoon, Chatree and Churintaraphan2013), and physical activity has an impact on NPY gene expression that may relate to its antidepressant properties (Melas et al., Reference Melas, Lennartsson, Vakifahmetoglu-Norberg, Wei, Åberg, Werme, Rogdaki, Mannervik, Wegener, Brené, Mathé and Lavebratt2013). Nevertheless, hypothesis-free searches in the genome have not identified NPY as a risk gene for any of the plausible candidate conditions, and the evidence for association of in vitro functional variations (Kallio et al., Reference Kallio, Pesonen, Kaipio, Karvonen, Jaakkola, Heinonen, Uusitupa and Koulu2001; Zhou et al., Reference Zhou, Zhu, Hariri, Enoch, Scott, Sinha, Virkkunen, Mash, Lipsky, Hu, Hodgkinson, Xu, Buzas, Yuan, Shen, Ferrell, Manuck, Brown, Hauger, Stohler, Zubieta and Goldman2008) in body composition is controversial. When the ECPBHS cohorts were analysed longitudinally for NPY variants, all examined genotypes were found as associated with changes in metabolic markers and with blood pressure (Katus et al., Reference Katus, Villa, Ringmets, Veidebaum and Harro2021). Age appeared as a modifying factor, possibly owing to lifestyle changes in transition from childhood to adulthood. Indeed, animal experiments have suggested that the effect of the orexigenic stimulus by NPY on feeding behaviour can be strongly dependent on a complex sequence of preliminary behaviours (Woods et al., Reference Woods, Figlewicz, Madden, Porte, Sipols and Seeley1998), and thus conceivably contingent on environmental factors.
Classic psychiatric risk markers: resurrection of platelet MAO and new thoughts about low cholesterol
Human platelets express the monoamine oxidase B isoenzyme (MAO-B, platelet MAO), that was long ago proposed as a marker of general psychiatric vulnerability (Buchsbaum et al., Reference Buchsbaum, Coursey and Murphy1976). The finding that tobacco smoking has a limited but dose-dependent inhibitory effect on MAO activity (Fowler et al., Reference Fowler, Volkow, Wang, Pappas, Logan, MacGregor, Alexoff, Shea, Schlyer, Wolf, Warner, Zezulkova and Cilento1996) depressed interest in this measure. (The measurement is also technically demanding.) Nevertheless, the body of evidence has indirectly suggested that, despite smoking being prevalent in patients and in socially deviant groups, platelet MAO activity is the most reliable biological correlate of impulsivity-related behaviours, especially if coexistent with other, independent risk factors (Oreland, Reference Oreland2004). For example, platelet MAO activity was lower in passenger car drivers who had been caught driving drunk (Eensoo et al., Reference Eensoo, Paaver, Pulver, Harro and Harro2004) and in patients with traumatic spinal cord injury (Sabre et al., Reference Sabre, Harro, Eensoo, Vaht, Kabel, Pakkanen, Asser and Kõrv2016); low platelet MAO was particularly prevalent in youngsters who had repeatedly committed criminal acts if this co-occurred with high psychopathy scores (Alm et al., Reference Alm, af Klinteberg, Humble, Leppert, Sörensen, Thorell, Lidberg and Oreland1996). Incidentally, the initial ECPBHS paper reported on platelet MAO activity, confirming for the first time in a birth cohort representative sample that males have, on average, lower enzyme activity than females (Harro et al., Reference Harro, Eensoo, Kiive, Merenäkk, Alep, Oreland and Harro2001). An important finding made in longitudinal observation was that platelet MAO activity is predictive of initiation of smoking, whereas both lower and higher than average enzyme activity related to smoking (Harro et al., Reference Harro, Fischer, Vansteelandt and Harro2004). Indeed, the work of Alm et al. (Reference Alm, af Klinteberg, Humble, Leppert, Sörensen, Thorell, Lidberg and Oreland1996) had shown the overrepresentation of subjects with both low and high platelet MAO activity among juvenile delinquents, highlighting the point that both low and high platelet MAO levels, reflective of lower and higher central serotonergic capacity (Fahlke et al., Reference Fahlke, Garpenstrand, Oreland, Suomi and Higley2002; Eriksson et al., Reference Eriksson, Berggren, Fahlke, Engel and Balldin2006; Harro et al., Reference Harro, Eensoo, Suvi, Timpmann and Ööpik2025), are risk factors for suboptimal behavioural choices, possibly owing to early developmental impact of serotonin (Harro & Oreland, Reference Harro and Oreland2016; Shah et al., Reference Shah, Courtiol, Castellanos and Teixeira2018). A twin study has revealed that when the analysis is corrected for smoking, platelet MAO activity can be positively associated with neuroticism (Kirk et al., Reference Kirk, Whitfield, Pang, Heath and Martin2001), which is also a risk factor not only for commencing smoking (Byrne et al., Reference Byrne, Byrne and Reinhart1995) but more generally of deviant brain responses and behaviour under pressure (Zhu et al., Reference Zhu, Wang, Chen, Lei, Yan, Yang, Yang and Wang2023; Ziegler et al., Reference Ziegler, Groß, Boreatti, Heine, McNeill, Kranz, Romanos, Jacob, Reif, Kittel-Schneider and Lesch2024; Fioravanti et al., Reference Fioravanti, Bocci Benucci and Ghinassi2025).
One controversial aspect in this regard has always been the very unreliable association of low platelet MAO activity with substance use disorders (apart from Type 2 alcoholism), given their link to risk-taking and antisocial conduct. In the ECPBHS males, low platelet MAO activity was indeed associated with higher prevalence of illicit drug experience, and with earlier debut, but this association was even stronger in subjects who had tried the substance only once (Sakala et al., Reference Sakala, Kasearu, Katus, Veidebaum and Harro2022). This observation explains the missing link between illicit drug use disorders and low platelet MAO activity, but the underlying mechanism of why the low MAO subjects less often progress from initiation to regular use remains to be clarified. This could relate, e.g., to higher novelty-driven behavioural strategies, less pleasurable (early) drug experience or lower sensitisation of the brain salience-attributing system by the drugs.
Low serum levels of cholesterol is another much-studied biomarker in psychiatry, often associated with impulsive action and suicide (e.g., Engelberg, Reference Engelberg1992; Wu et al., Reference Wu, Ding, Wu, Xie, Hou and Mao2015; Khosravani et al., Reference Khosravani, Samimi Ardestani, Berk, Aghaeimazraji and Sharifibastan2025), but not previously investigated with longitudinal design. Taking advantage of the ECPBHS database, we have assessed the link between impulsivity measured in young adulthood (age 25) and cholesterol levels measured at four time points, starting from age 9. In this sample, the correlation between impulsivity self-reports and cholesterol levels was stronger, the earlier the serum sample was collected (Tomson-Johanson et al., Reference Tomson-Johanson, Kaart, Kiivet, Veidebaum and Harro2020). This unexpected finding suggests that the association between cholesterol levels and impulsivity is the reflection of developmental processes at early age.
The 5-HTTLPR in the ECPBHS
5-HTTLPR, denoting the serotonin transporter gene linked polymorphic region (Heils et al., Reference Heils, Teufel, Petri, Stöber, Riederer, Bengel and Lesch1996), is almost a notorious gene variation, as it has become the most studied target in the ‘candidate gene’ studies of psychiatric disorders and a variety of behaviours, and brain structure and function. The very strong association of the short (s) versus long (l) allele (meaning the extent of tandem repeats) in the putative promoter region with neuroticism, harm avoidance and anxiety disorders in the original study (Lesch et al., Reference Lesch, Bengel, Heils, Sabol, Greenberg, Petri, Benjamin, Müller, Hamer and Murphy1996), however, immediately raised the question of what is the evolutionary meaning of such a high frequency of the allegedly neuroticism-promoting s-allele, present in the majority of Europeans and in the vast majority of Japanese. Several subsequent studies have not replicated the original association. While the findings on the Dunedin Birth Cohort demonstrated that the s-allele, and especially the s/s-homozygocity, contributed strongly to depression and suicidality only in the context of stressful life events (Caspi et al., Reference Caspi, Sugden, Moffitt, Taylor, Craig, Harrington, McClay, Mill, Martin, Braithwaite and Poulton2003), even these results were not easily replicable. The sheer number of research papers reporting statistically significant associations of 5-HTTLPR with a wide variety of behaviours and measures suggests that this genetic variation may bear important functional consequences at a very deep layer of neural processes, with huge dependency of the observable phenomena on the multiple developmental steps. The ECPBHS sample was explored for 5-HTTLPR after successful replications of the original finding (Hariri et al., Reference Hariri, Mattay, Tessitore, Kolachana, Fera, Goldman, Egan and Weinberger2002) that the amygdala activation is higher in carriers of the 5-HTTLPR s-allele. Importantly, an analogous polymorphism, present in rhesus macaques, was found associated with higher frequency or longer duration of exhibiting fear and anxiety related behaviours in several non-human primate populations (Bennett et al., Reference Bennett, Lesch, Heils, Long, Lorenz, Shoaf, Champoux, Suomi, Linnoila and Higley2002; Howarth et al., Reference Howarth, Szott, Witham, Wilding and Bethell2023). We observed higher Big Five neuroticism in the 5-HTTLPR s/s-homozygous participants, but also lower openness to experience, agreeableness and conscientiousness at age 9 as reported by parents; extraversion was not associated with the 5-HTTLPR (Harro et al., Reference Harro, Merenäkk, Nordquist, Konstabel, Comasco and Oreland2009). At age 15, parental reports no longer differed by genotype, but the s/s-homozygous subjects self-reported higher neuroticism. By age 18, this difference had also disappeared. In contrast, as the children aged, the 5-HTTLPR genotype effect emerged in terms of experimenting with intoxicants: this measure increasing with age, the 5-HTTLPR s/s-homozygocity more frequently associated with smoking, alcohol intake and use of illicit drugs (Merenäkk et al., Reference Merenäkk, Mäestu, Nordquist, Parik, Oreland, Loit and Harro2011; Vaht et al., Reference Vaht, Merenäkk, Mäestu, Veidebaum and Harro2014). Hence, in a representative sample, the 5-HTTLPR genotype is associated with personality and behaviour, while the associations follow the developmental curve. More recently, ongoing brain imaging studies have shed further light on the impact of the 5-HTTLPR to structure and function of limbic networks, and shown it to be complex (e.g., Hariri et al., Reference Hariri, Drabant and Weinberger2006; Kirchner et al., Reference Kirchner, Garvert, Wittfeld, Ameling, Bülow, Meyer Zu Schwabedissen, Nauck, Völzke, Grabe and Van der Auwera2023: Klöbl et al., Reference Klöbl, Murgaš, Reed, Silberbauer, Hartmann, Godbersen, Gryglewski, Nics, Hahn, Rujescu, Hacker and Lanzenberger2025) and experience-dependent (Ancelin et al., Reference Ancelin, Carrière, Artero, Maller, Meslin, Dupuy, Ritchie, Ryan and Chaudieu2021), which may lead to variable results between studies on highly divergent samples. Nevertheless, response of the neurohormonal stress axis diverges by genotype (Sun et al., Reference Sun, Li, Zhong, Dong, Ming, Gao, Xiong, Cheng, Zhao, Wang and Yao2020; Kuhn et al., Reference Kuhn, Noack, Skoluda, Wagels, Röhr, Schulte, Eisenkolb, Nieratschker, Derntl and Habel2021). The 5-HTTLPR s-allele carriers appear as having a higher sensitivity to stress, but also higher capability to recover, adding a time dimension to the gene × environment (G×E) interaction studies (Delli Colli et al., Reference Delli Colli, Borgi, Poggini, Chiarotti, Cirulli, Penninx, Benedetti, Vai and Branchi2022). Thus, with regard to compromising mental health, the 5-HTTLPR s-allele may serve as an amplifier of more causal factors, such as, in the ECPBHS sample, enhancing anxiety and bulimia in subjects with binge eating (Akkermann et al., Reference Akkermann, Nordquist, Oreland and Harro2010). Fittingly with this example, the l/l-homozygocity of the 5-HTTLPR is associated with stronger responses to high-calorie food in cognitive control brain areas, indicating better capacity to resist the food reward (Markus and Keulers, Reference Markus and Keulers2025).
Gene × environment interactions: from family relationships to birth cohort effects
Gene variants interact with environmental factors in the development of the nervous system, in the formation of behavioural traits, and in the pathogenesis of psychiatric disorders – but how? We know something about the structure of heredity, but not much about the interrelationship of all things that can be interpreted as environmental impacts. What is defined as adverse life events may differ in important ways with regard to their power, duration, and timing. ECPBHS has a rich collection of environmental data, which indicate that common gene variants often interact with the sum of adverse life events quite differently in comparison with the perceived family environment.
While not the first functional polymorphism shown to interact with environmental measures, 5-HTTLPR was made the spearhead case by the report of Caspi et al. (Reference Caspi, Sugden, Moffitt, Taylor, Craig, Harrington, McClay, Mill, Martin, Braithwaite and Poulton2003). Also the monkey variant of 5-HTTLPR is the subject of G×E interactions, the s-allele carriers being more sensitive to adverse rearing conditions in terms of brain development (Schoenfeld et al., Reference Schoenfeld, Gupta, Syed, Rozenboym, Fulton, Jackowski, Perera and Coplan2021). In humans, 5-HTTLPR has most consistently been associated with the diagnosis or symptoms of eating disorders, the s-allele carriers having a higher vulnerability to physical or sexual abuse (Rozenblat et al., Reference Rozenblat, Ong, Fuller-Tyszkiewicz, Akkermann, Collier, Engels, Fernandez-Aranda, Harro, Homberg, Karwautz, Kiive, Klump, Larson, Racine, Richardson, Steiger, Stoltenberg, van Strien, Wagner, Treasure and Krug2017). This cited meta-analysis included data of the ECPBHS that had reported overall similar findings (Akkermann et al., Reference Akkermann, Kaasik, Kiive, Nordquist, Oreland and Harro2012). Adverse life events did not increase the drive for thinness and bulimia in the 5-HTTLPR l/l-homozygous participants, but such an increase was observed in the s-allele carriers, with a larger increase in subjects who had experienced more adversities. The single most impactful life event in this regard was sexual abuse. Interestingly, the s-allele carriers with no reported adverse life events had the lowest symptom scores of all groups, supporting the higher plasticity in the 5-HTTLPR s-allele carriers, also observable in the bidirectional impact of parenting styles on decision making under risk (Rehn et al., Reference Rehn, Nilsson, Hultman, Vadlin and Åslund2025).
Life events can thus have a different impact, and the meaning of each type of life event may vary individually owing to genetic and developmental neural factors, and learning. Therefore, offering a list of events and forming a score may be the safest while not always the optimal method for G×E. A more instructive inquiry into G×E might arise from the comparison of birth cohorts. Subjects in a birth cohort are exposed to a more similar total environment as compared to others, especially the social environment (Wedow et al., Reference Wedow, Zacher, Huibregtse, Mullan Harris, Domingue and Boardman2018; Virtanen et al., Reference Virtanen, Kaprio, Viken, Rose and Latvala2019), and this has consequences to e.g., psychological distress (Keyes et al., Reference Keyes, Nicholson, Kinley, Raposo, Stein, Goldner and Sareen2014), depressive symptoms (Yoo, Reference Yoo2023), drinking alcohol (Pabst et al., Reference Pabst, Kraus, Piontek and Mueller2010; Radaev & Roshchina, Reference Radaev and Roshchina2019), and overweight and obesity (Allman-Farinelli et al., Reference Allman-Farinelli, Chey, Bauman, Gill and James2008; Opazo Breton & Gray, Reference Opazo Breton and Gray2023). Studies on G×E interactions in terms of allelic variants and birth cohorts are, however, limited. With regard to drinking alcohol, the longitudinal Older Finnish Twin Cohort study detected variation by birth cohort of the contribution of genetic and environmental factors on alcohol consumption in the Finnish population (Virtanen et al., Reference Virtanen, Kaprio, Viken, Rose and Latvala2019). The ECPBHS birth cohorts were found to differ in their alcohol use debute: as expected, the younger cohort started to experiment younger, and the gender gap diminished, which are results expected on the grounds of research in other countries (Keyes et al., Reference Keyes, Li and Hasin2011). The most interesting finding, however, was the three-way interaction between birth cohort, gender and the 5-HTTLPR genotype (Vaht et al., Reference Vaht, Merenäkk, Mäestu, Veidebaum and Harro2014). The most prominent difference emerged in the comparison of s/s-homozygous females of the two cohorts: While in the older cohort this group was the last to taste alcohol, it was the earliest in the younger cohort, even surpassing boys. The age gap between these two groups was on average three years, which is sufficiently large to not be trivial: It can make a difference, if one starts drinking not in the 9th grade, but in the 6th.
But could social desirability itself, and its underlying neurobiology, be subject to birth cohort effect? Agreeableness, the social adjustment-oriented Big Five personality dimension, has been found to be sensitive to birth cohort effects, and uniquely even changeable in a cohort-dependent manner (Brandt et al., Reference Brandt, Drewelies, Willis, Schaie, Ram, Gerstorf and Wagner2022). Agreeableness correlates negatively with the dimension ANGER of the Affective Neuroscience Personality Scale (ANPS; Davis & Panksepp, Reference Davis and Panksepp2011; Montag et al., Reference Montag, Elhai and Davis2021) and positively with CARE, and is indeed higher in the younger cohort of the ECPBHS than in the older (Kiive et al., Reference Kiive, Kanarik, Veidebaum and Harro2024a). Interestingly, Agreeableness associated in a complex manner with anxiety-related gene variants. NPY strongly attenuates anxiety in animal experiments (Heilig & Widerlöv, Reference Heilig and Widerlöv1995) including social anxiety, and controls the stress axis by attenuating excessive, anxiety-provoking stimulation of corticotropin-releasing factor type 1 receptors (Kask et al., Reference Kask, Rägo and Harro1997), yet the NPY variants are not known to relate to anxious traits. Neither were the body composition and metabolism related NPY variations rs16147 and rs5574 (see above) associated with anxious personality, but in either case, homozygous subjects for one variant strongly differed by birth cohort in Agreeableness, whereas this interaction was amplified by the 5-HTTLPR genotype (Kiive et al., Reference Kiive, Kanarik, Veidebaum and Harro2024a). If the interval of the ECPBHS birth cohorts would appear small, it must be recalled that the study area was a theatre for the fastest transition amongst the Countries of Central and Eastern Europe from central planning to free market economy (Allaste & Bennett, Reference Allaste, Bennett and Allaste2013), and that major economic shifts precipitate cohort effects (Sutin et al., Reference Sutin, Terracciano, Milaneschi, An, Ferrucci and Zonderman2013). That major societal changes with individualism taking the helm from collectivism could place a heavy burden on the neurobiology that has evolved to support prosocial behaviour is conceivable. Social fear conditioning has been reported to produce the most prominent alterations in the expression of NPY and serotonin-related genes throughout the brain (Hamann et al., Reference Hamann, Bankmann, Mora Maza, Kornhuber, Zoicas and Schmitt-Böhrer2022), thus supporting the notion that these neurochemical systems interact in social adaptive strategies, hypothetically through their interaction in orbitofrontal cortex to support behavioural flexibility (Longo et al., Reference Longo, Fadda, Brasso, Mele, Palanza, Nanavaty, Bertocchi, Oberto and Eva2018). A telling case has recently been provided in the nascent research on the role of RNA m6A methylation in neuropsychiatry (Malovic & Pandey, Reference Malovic and Pandey2023). Levels of RNA m6A methylation and expression of METTL3, a RNA methyl transferase and FTO, a RNA demethylase, have been linked to depressive disorder and suicide risk (Roy & Dwivedi, Reference Roy and Dwivedi2025), and pharmacological activation of METTL3 has anxiolytic-like properties (Kanarik et al., Reference Kanarik, Liiver, Norden, Teino, Org, Laugus, Shimmo, Karelson, Saarma and Harro2025). Cai et al. (Reference Cai, Forsell, Lavebratt and Melas2024) have reported on the association of the FTO rs1421085 T-allele with higher neuroticism, and we have confirmed this finding in a longitudinal analysis of the ECPBHS personality data (Kurrikoff et al., Reference Kurrikoff, Eensoo and Harro2026). Interestingly, this association was apparent in the older cohort at age 25, data having been collected during the impact of the Great Recession of 2007–2009, when the subjects of the older cohort also reported much more socio-economic difficulties (Kurrikoff et al., Reference Kurrikoff, Eensoo and Harro2026). This cohort effect extends to changes in dietary intake, body composition, and glucose metabolism (Katus et al., Reference Katus, Villa, Ringmets, Eensoo, Mäestu, Veidebaum and Harro2026). These findings agree well with the conclusion that the impact of total environment on brain health and behavioural outcomes, recently termed as ‘exposome’ should be taken seriously in the study of development of brain health (Robinson et al., Reference Robinson, Dave, Barzilay, Wagner, Kells and Keller2026).
Why impulsivity?
Impulsivity is the most common symptom across the whole DSM-5, possibly owing to its potentially devastating impact on social adaptation. Yet it appears that the meaning of premature action is in its eventual outcome, as preference of speed over accuracy, may confer benefits. The delicate balance in the multifaceted regulation of impulse control has been searched in the serotonin system (Evenden, Reference Evenden1999), and in the ECPBHS, an intriguing interaction between two markers of the system was revealed. Thus, impulsivity as measured with either a computer-based discrimination task or the Barratt Impulsivity Scale correlated negatively with platelet MAO activity in the s-allele carriers of the 5-HTTLPR, while among 5-HTTLPR l/l-homozygotes this association rather took an inverted U-shape, as participants with low enzyme activity exhibited lower impulsivity (Paaver et al., Reference Paaver, Nordquist, Parik, Harro, Oreland and Harro2007). In the visual discrimination-based computer task, participants with the l/l-genotype and high platelet MAO activity had the lowest tendency of impulsive action; interestingly, this combination of biomarkers had, in females, a synergistic effect of increasing drive for thinness, a compulsivity measure (Akkermann et al., Reference Akkermann, Paaver, Nordquist, Oreland and Harro2008). Which relationship aspects of impulsivity and compulsivity take, appears to depend on the inner structure of these two constructs, sometimes possible to be considered as opposite poles of one dimension, and in other instances sharing the substrate and acting synergistically (Grant & Kim, Reference Grant and Kim2014).
Where impulsivity is being taken as such a negative trait, which mechanisms maintain it in the ‘survival of the fittest’? Amongst the variety of impulsivity measures collected in the ECPBHS, the favourite has been the Adaptive and Maladaptive Impulsivity Scale (AMIS), which has drawn on the concept of functional and dysfunctional impulsivity of Dickman (Reference Dickman1990). This concept contrasts a tendency to act with little forethought when it is optimal against the tendency for thoughtlessness and inability to plan, leading to negative consequences. It appears likely that fast decision-making and action are sufficiently important to be selected for, that its other, maladaptive side can be tolerated by evolution, given that environmental contingencies vary. In the ECPBHS sample, a functional polymorphism in the neuronal nitric oxide synthase gene (NOS1) provided an illuminating example of G×E in impulsivity. Deletion of this gene, responsible for the glutamate-responsive production of the gaseous neurotransmitter in the CNS (Zhang & Snyder, Reference Zhang and Snyder1993), in mice results in reduction of frontal-cortical serotonergic neurotransmission (Chiavegatto et al., Reference Chiavegatto, Dawson, Mamounas, Koliatsos, Dawson and Nelson2001) and impulsive-aggressive behaviour, that is, however, moderated by environmental factors and develops only if living in social isolation (Chiavegatto & Nelson, Reference Chiavegatto and Nelson2003). Homozygocity for short (s) alleles of the dinucleotide repeat polymorphism in the promoter region of the alternative first exon 1f of human NOS1 (NOS1 ex1f-VNTR) was found to be associated with impulsivity and aggressiveness in diverse samples by a variety of measures, and with hypoactivation of the anterior cingulate cortex (Reif et al., Reference Reif, Jacob, Rujescu, Herterich, Lang, Gutknecht, Baehne, Strobel, Freitag, Giegling, Romanos, Hartmann, Rösler, Renner, Fallgatter, Retz, Ehlis and Lesch2009). In the ECPBHS, the NOS1 ex1f-VNTR genotype was confirmed to be related to all analysed impulsivity constructs, and in a G×E manner that illustrated the multifaceted role of ‘impulsivity alleles’ in behaviour, and their persistence (Reif et al., Reference Reif, Kiive, Kurrikoff, Paaver, Herterich, Konstabel, Tulviste, Lesch and Harro2011). In brief, self-reports indicated higher adaptive impulsivity in the s-allele carriers, while behavioural tests revealed a larger number of errors; they were more prone to an increase of maladaptive impulsivity if the family relationships had been inferior, but this dysfunctional measure did not increase with adverse life events, which was the case with l-allele homozygotes. Overall, when generalising across measures and conditions, heterozygotes demonstrated the best adaptive performance.
Another telling case of heterosis in impulsivity, and related apects of mental health, was observed with the NPSR1 Asn107Ile (A > T; rs324981) polymorphism. Neuropeptide S (NPS) is an evolutionarily conserved peptide with restricted localisation in the CNS, and a role in arousal and anxiety (Xu et al., Reference Xu, Reinscheid, Huitron-Resendiz, Clark, Wang, Lin, Brucher, Zeng, Ly, Henriksen, de Lecea and Civelli2004). The appearance of NPS and its receptor NPSR1 in the lungfish has been suggested to underlie anxiety regulation in the water-to-land transition (Wang et al., Reference Wang, Wang, Zhu, Yang, Ren, Ruan, Fan, Hu, Xu, Bi, Zhu, Song, Chen, Ma, Zhao, Jiang, Zhang, Feng, Yuan, Gan, Li, Zeng, Liu, Zhang, Shao, Hao, Zhang, Xu, Liu, Wang, Zhu, Zhang, Zhao, Qiu, He and Wang2021). The rs324981 polymorphism in the human gene encoding the NPS receptor leads to exchange of an amino acid and results in a more effective signal mediation with the minor allele (Reinscheid et al., Reference Reinscheid, Xu, Okamura, Zeng, Chung, Pai, Wang and Civelli2005). This corresponds to the higher prevalence of panic disorder in females and elevated anxiety sensitivity (Domschke et al., Reference Domschke, Reif, Weber, Richter, Hohoff, Ohrmann, Pedersen, Bauer, Suslow, Kugel, Heindel, Baumann, Klauke, Jacob, Maier, Fritze, Bandelow, Krakowitzky, Rothermundt, Erhardt, Binder, Holsboer, Gerlach, Kircher, Lang, Alpers, Ströhle, Fehm, Gloster, Wittchen, Arolt, Pauli, Hamm and Deckert2011). Responsiveness of the basolateral amygdala to fear-relevant stimuli was increased by each NPSR1 Asn107Ile T-allele (Dannlowski et al., Reference Dannlowski, Kugel, Franke, Stuhrmann, Hohoff, Zwanzger, Lenzen, Grotegerd, Suslow, Arolt, Heindel and Domschke2011); also suggestive of higher arousability, the T/T-homozygous subjects had less sleep (Spada et al., Reference Spada, Sander, Burkhardt, Häntzsch, Mergl, Scholz, Hegerl and Hensch2014), that is compatible with the role of NPS to promote wakefulness through histamin- and orexinergic neurons in animal experiments (Zhao et al., Reference Zhao, Shao, Zhang, Fan, Kong, Wang and Hou2012). Interestingly, the T/T-homozygotes had higher startle response to neutral stimuli, but lower startle in response to unpleasant stimuli in caffeine vs. placebo condition (Domschke et al., Reference Domschke, Klauke, Winter, Gajewska, Herrmann, Warrings, Mühlberger, Wosnitza, Dlugos, Naunin, Nienhaus, Fobker, Jacob, Arolt, Pauli, Reif, Zwanzger and Deckert2012). The NPSR1 Asn107Ile allele homozygous participants had highly distinct gender-dependent developmental profiles of impulsivity and mental health-related symptoms in the ECPBHS. Thus, homozygocity for the T-allele, previously associated with psychiatric vulnerability, was indeed associated with ADHD-related traits and, if combined with a higher score of stressful life events, with an increase of maladaptive as well as adaptive impulsivity (Laas et al., Reference Laas, Reif, Kiive, Domschke, Lesch, Veidebaum and Harro2014b). The T/T-homozygous participants, however, did not appear as highly sensitive to inferior family environment. In contrast, the A/A-homozygous subjects had higher adaptive impulsivity and openness to experience but, if subjected to less favourable family relations, their adaptive impulsivity and extraversion decreased, whereas maladaptive impulsivity and neuroticism increased. The A/A homozygous females exposed to environmental adversities had affective and anxiety disorders more frequently; they also exhibited higher anxiety and depressiveness and lower self-esteem, and reported more frequently the experience of suicidal thoughts/behaviour (Laas et al., Reference Laas, Reif, Akkermann, Kiive, Domschke, Lesch, Veidebaum and Harro2014a). Both environmental adversities and early alcohol use debut, as expected, increased the probability of alcohol use disorder, but its development included G×E by gender (Laas et al., Reference Laas, Reif, Akkermann, Kiive, Domschke, Lesch, Veidebaum and Harro2015). Namely, alcohol use disorder was more common in female carriers of the A-allele and male carriers of the T-allele, with the tendency of homozygocity in either case being the most vulnerable. In summary, one can imagine two different NPSR1 Asn107Ile-related pathways to the disorder: In females, environmental adversities together with the A-allele promoted neuroticism, followed by early alcohol use likely related to perceived social pressures, and in male T-allele carriers, through even earlier alcohol debut in association with impulsive and ADHD-like traits. Of note, in the population-representative ECPBHS sample, the T-allele is not the ‘minor’ allele, but the allele frequency is equal. Addictions can involve disturbances in multiple neurobiological networks, such as those responsible for the regulation of hedonic values, associative learning, formation of habits, stress response, attribution of salience, physiological withdrawal response, changes of mood and impulsive decision-making (Baler & Volkow, Reference Baler and Volkow2006). Individuals may vary a great deal in the relative importance of these networks in their addiction. Hence the large number of genes involved in substance use disorders, and their potential to contribute along more than one pathway.
Aggressiveness, agonistic and antisocial behaviour
A prominent cross-diagnostic trait in psychiatry is impulsivity-related aggressiveness (Vuijk et al., Reference Vuijk, Martin, Braaten, Genovese, Capawana, O’Keefe, Lee, Lind, Smoller, Faraone, Perlis and Doyle2020). In modern society, inter-personal aggression is considered unacceptable, and it is disruptive and detrimental to families and the whole communities (Dai & Lin, Reference Dai and Lin2025). Aggressiveness is, however, a prevalent aspect of innate social behaviour across the animal kingdom, and can also be interpreted as an important component in the coordinated adaptive response to challenges presented by the environment (van Kampen, Reference van Kampen2015). But, again both in mice and in men, aggression is increased by social deprivation in early age, and can be mitigated by stable care and reintegration (Calado et al., Reference Calado, Manhães-de-Castro, Gouveia, Nogueira, da Silva Souza, Soares Martins de Lira, de Araújo Merencio, Lima, de Oliveira MV and Toscano2025).
The ECPBHS database was mined with machine-learning techniques to reveal predictors of antisocial behaviour leading to police contact in adulthood, including more than 554 behavioural and environmental measures in the hypothesis-free analysis. As expected, antisocial behaviour was more common in males; apart from this, past substance use disorder, aggressive mode of action upon provocation, and teacher-rated concentration difficulties and physical fighting in school at age 15 years appeared as the strongest risk factors (Schoenmacker et al., Reference Schoenmacker, Sakala, Franke, Buitelaar, Veidebaum, Harro, Heskes, Claassen and Alejandro2020). At first glance, such an outcome is not surprising, but interestingly, stress-related measures were not predictive of antisocial behaviour, unless accompanied by substance abuse. Thus, exposure to stress does not, in most instances, derail socially well-adjusted development, but sometimes it does. Among the neurobiological vulnerabilities that could predict the failure to adapt without outright aggression, the role of the suboptimal serotonergic system has received the most attention, owing to its well-documented role in impulsive acts (Coccaro et al., Reference Coccaro, Fanning, Phan and Lee2015).
The impact of early age serotonin levels on the development of CNS and the consequent adult behavioural integrity is, in terms of functional long-term outcome, of inverted U-shape, both low and excessive levels of serotonin leading to suboptimal performance (Shah et al., Reference Shah, Courtiol, Castellanos and Teixeira2018). The enzyme responsible for the synthesis of serotonin in the brain is tryptophan hydroxylase 2 (TPH2). The TPH2 gene has a promoter polymorphism −703G/T (rs4570625), the minor T-allele associated with reduced serotonin levels, and with a larger mean connectivity in the ‘rich club’, the heavily interconnected hubs (Markett et al., Reference Markett, de Reus, Reuter, Montag, Weber, Schoene-Bake and van den Heuvel2017). Commonly all T-allele carriers of TPH2 −703G/T are kept together in data analysis, but in the ECPBHS it was the small (<5%) group of T/T homozygous participants that stood out with regard to their remarkably low aggressiveness and maladaptive impulsivity, and with almost total absence of anxiety disorders (Laas et al., Reference Laas, Kiive, Mäestu, Vaht, Veidebaum and Harro2017). This was particularly clear for male subjects, the male T/T group reporting – and retrospectively reported by their classmates – levels of aggressiveness typical for females. It is not clear yet to which degree common TPH2 variants can reduce the availability of serotonin, but genetic modification of TPH2 levels in mice profoundly changes expression of aggressiveness, in a sex-dependent manner (Kästner et al., Reference Kästner, Richter, Urbanik, Kunert, Waider, Lesch, Kaiser and Sachser2019).
To explore beyond serotonin, a genome-wide search of differentially methylated DNA loci was performed in peripheral blood collected from a subsample of the ECPBHS male subjects at age 15 and 25 (Pishva et al., Reference Pishva, van den Hove, Laroche, Lvovs, Roy, Ortega, Burrage, Veidebaum, Kanarik, Mill, Lesch and Harro2023). The LHA total score was associated with methylation of the PDLIM5, a gene with a role in dendrite branching (Srivastava et al., Reference Srivastava, Donta, Mireles, Paulucci-Holthauzen, Waxham and McCrea2024) and previously associated with affective disorders and schizophrenia (e.g., Kato et al., Reference Kato, Iwayama, Kakiuchi, Iwamoto, Yamada, Minabe, Nakamura, Mori, Fujii, Nanko and Yoshikawa2005). A subset of differentially methylated positions at age 25 were already, at age 15, predictive of later aggressiveness. Top differentially methylated positions colocalized with genetic variants previously associated, not surprisingly, with general cognitive function and educational attainment, but also with cholesterol levels. The latter finding appears highly relevant from the viewpoint of the role of low cholesterol levels in impulsive tendencies, to which a novel perspective was added in the ECPBHS (see above).
Neurotransmission by ‘classic’ neurotransmitters such as serotonin is fine-tuned at higher activity levels by a host of neuropeptides (Hökfelt et al., Reference Hökfelt, Barde, Xu, Kuteeva, Rüegg, Le Maitre, Risling, Kehr, Ihnatko, Theodorsson, Palkovits, Deakin, Bagdy, Juhasz, Prud’homme, Mechawar, Diaz-Heijtz and Ögren2018), including orexins (Schöne & Burdakov, Reference Schöne and Burdakov2017). A unique study on epileptic patients monitored the release of orexin A in the amygdala and found it increased while subjects reported higher levels of anger (Blouin et al., Reference Blouin, Fried, Wilson, Staba, Behnke, Lam, Maidment, Karlsson, Lapierre and Siegel2013). Orexins, alternatively called hypocretins, are hypothalamic neuropeptides that maintain wakefulness and regulate emotional arousal as well as autonomic functions in a coordinated defence response (Johnson et al., Reference Johnson, Molosh, Fitz, Truitt and Shekhar2012; Kuwaki, Reference Kuwaki2021). Orexin-expressing neurons are activated by a large variety of stressors, and hence it has been proposed that orexin neurotransmission becomes salient during highly arousing aversive conditions (Berridge et al., Reference Berridge, Espana and Vittoz2010). A variant of the HCRTR1 gene (rs2271933, G1222A) in exon 7 leads to amino acid substitution (Ile408Val) (Meerabux et al., Reference Meerabux, Iwayama, Sakurai, Ohba, Toyota, Yamada, Nagata, Irukayama-Tomobe, Shimizu, Yoshitsugu, Ohta and Yoshikawa2005). In both younger and older cohorts of the ECPBHS, the HCRTR1 Ile408Val genotype was significantly associated with scores of the BPAQ and the LHA interview, the A/A homozygous participants reporting higher aggression (Harro et al., Reference Harro, Laas, Eensoo, Kurrikoff, Sakala, Vaht, Parik, Mäestu and Veidebaum2019). This was the first demonstration of the involvement of the orexin system in aggressiveness, whereas further study on the genotype revealed a variable relationship of aggressiveness with reward sensitivity (Pulver et al., Reference Pulver, Kiive, Kanarik and Harro2020b; see below), illuminating genotype-based differences in the psychological background of aggression.
Components of reward sensitivity?
Reward sensitivity is emerging as a major transdiagnostic component of psychiatric disorders (Oka et al., Reference Oka, Sasaki and Kobayashi2025). Reward sensitivity is a reflection of the activity in the behavioural approach system, one of the three basic motivational systems (Gray & McNaughton, Reference Gray and McNaughton2000), that has been applied to make predictions for depression (Hazell, Reference Hazell2021), attention deficit-hyperactivity disorder (ADHD; Brooker et al., Reference Brooker, Ostojic and Miller2018), substance use (Liu & Filbey, Reference Liu and Filbey2024), psychopathy (De Pascalis et al., Reference De Pascalis, Scacchia, Sommer and Checcucci2019), aggressiveness (Parker et al., Reference Parker, Farrell and Walker2022), antisocial behaviour (Hoppenbrouwers et al., Reference Hoppenbrouwers, Neumann, Lewis and Johansson2015), and depressive/hypomanic symptoms in daily life (Sperry et al., Reference Sperry, Stromberg, Roberts and Prisciandaro2025). Multiple questionnaires and computer tasks can assess reward sensitivity, and the relative inconsistency in linking reward sensitivity with several mental health conditions may derive from this multiplicity, but also from reward sensitivity being a multidimensional construct (Corr, Reference Corr2016). Using the pool of item data collected in the ECPBHS, a novel scale was developed that distinguished two independently variable components of reward sensitivity, one reflecting the striving towards a multiplicity of rewards (Openness to Rewards), and the other, an excessive fixation on a particular reward (Insatiability by Reward). These two components of reward sensitivity had highly divergent relationships with personality traits in the Affective Neuroscience taxonomy: SEEKING and PLAY (and, to a lesser extent, CARE) co-varied with Openness to Rewards, while FEAR, SADNESS, and ANGER were, less strongly related to Insatiability to Rewards (Pulver et al., Reference Pulver, Kiive and Harro2020a). Insatiability by Reward, but not Openness to Rewards, was linked to symptoms of ADHD and aggressiveness. The dissociation between these two facets of reward sensitivity extended to metabolic measures: Insatiability by Reward was related to body mass index, sum of skinfolds, and serum triglyceride levels, but negatively with body mass adjusted energy intake and cardiorespiratory fitness (Kiive et al., Reference Kiive, Katus, Eensoo, Villa, Mäestu, Veidebaum and Harro2025). Openness to Rewards was, again, not related to these measures, but positively associated with physical activity and negatively with blood pressure and serum levels of glucose, insulin and cholesterol levels.
Structuring reward sensitivity this way may explain the variable findings in the literature, but of higher salience is the derived hypothesis that individual differences in reward sensitivity are part of a complex physiological variability, including energy expenditure profiles, and behavioural solutions to these inherent differences in metabolism. Reward sensitivity is evolutionarily conserved: many mammalian species show reward-related behavioural patterns similar to humans (Spear, Reference Spear2011) and share its neurobiological substrate (Panksepp, Reference Panksepp1998). It is plausible that variability within species of energy use aids survival in changeable environments and facilitates habitation of environments with different demands. This in turn has the potential to interfere with inter-individual variability of multiple other survival-related traits, not the least aggressive or agonistic behaviour.
The above-mentioned case of association of aggressiveness with the HCRTR1 Ile408Val genotype (Harro et al., Reference Harro, Laas, Eensoo, Kurrikoff, Sakala, Vaht, Parik, Mäestu and Veidebaum2019) provides a tool for probing the complex relationship between reward sensitivity and aggression. The orexinergic system has been proposed to serve as a link between physical activity, metabolism, and cognition, integrating energy balance and behavioural responses (Nigro et al., Reference Nigro, Argentino, Musumeci and Daniele2025), and possibly also moderating aggressiveness. Not surprisingly, in the whole sample, aggressiveness was in positive correlation with Insatiability by Reward and not with Openness to Rewards. Intriguingly, the more aggressive genotype, the HCRTR1 Ile408Val A/A homozygous group that represents 15% of the sample, had a different pattern of association (Pulver et al., Reference Pulver, Kiive, Kanarik and Harro2020b). Namely, in the A/A homozygous subjects, the correlation between aggressiveness and Insatiability by Reward was substantially lower than in the G-allele carriers (r = 0.27 vs. 0.53), while the correlation between aggressiveness and Openness to Rewards, virtually zero in the G-allele carriers, was small but statistically significant in the A/A group. Thus, the higher aggressiveness of the HCRTR1 Ile408Val A/A homozygous participants may relate to a distinct reward sensitivity profile. Speculatively, the A/A subjects may be more likely to get into conflict-provoking situations owing to their pursuit of multiple rewards. Such individual differences in psychological traits underlying aggression may guide improvement of individualised treatments (Blair et al., Reference Blair, Habel, Votinov, Wagels and Denson2025).
Coping strategies
Searching only for highly+specific statistically+significant differences or associations in group comparisons may obscure variability within the groups: If, say, low serotonergic capacity can predict the risk of impulsive behaviours and the consequent failure of social adjustment, then what are the resources of many subjects with genetically or developmentally low serotonergic capacity who thrive despite a presence of the measurable risk factor(s)? Such resources may range from interactions in the brain at the molecular level to action patterns at the level of community or society. The direction of causality, however, mostly remains unexplained. For example, serotonin release capacity is lower in people who reside in census tracts of lower socio-economic status (Manuck et al., Reference Manuck, Bleil, Petersen, Flory, Mann, Ferrell and Muldoon2005), but at present, how this difference develops begs further inquiry.
The discovery of a rare point mutation in the gene encoding the MAO-A isoenzyme that rendered the enzyme functionally ineffective and caused the development of weak impulse control and predisposition to violent outbursts in response to unexpected stressful stimuli (Brunner et al., Reference Brunner, Nelen, Breakefield, Ropers and van Oost1993) led to MAOA often being labelled as a “gene for aggression”. Subsequently, an upstream 30 base pair variable number of tandem repeats (uVNTR) polymorphism in the promoter region of the MAO-A gene was described (Sabol et al.,Reference Sabol, Hu and Hamer1998), and it was found that the alleles leading to low activity MAO-A (MAOA-L) in males predict diminished responses of prefrontal cortex to angry and fearful faces, whereas responses of the amygdala are increased (Meyer-Lindenberg et al., Reference Meyer-Lindenberg, Buckholtz, Kolachana, Hariri, Pezawas, Blasi, Wabnitz, Honea, Verchinski, Callicott, Egan, Mattay and Weinberger2006). In males, the MAOA-L genotype has been associated with higher aggressiveness or more violent behaviour, but not consistently with traits of common personality models (Harro & Oreland, Reference Harro and Oreland2016). Importantly, male MAOA-L subjects are more likely to present an aggressive/antisocial profile if having been maltreated in childhood (Caspi et al., Reference Caspi, McClay, Moffitt, Mill, Martin, Craig, Taylor and Poulton2002; Byrd & Manuck, Reference Byrd and Manuck2014). No association of the MAOA genotype with aggressiveness was noted in the ECPBHS sample. However, consist with the concept of ‘plasticity genes’ in children from supportive environments, the MAOA-L genotype is protective against delinquency (Nilsson et al., Reference Nilsson, Comasco, Hodgins, Oreland and Åslund2014). Serious maltreatment is not common in the ECPBHS sample. Instead, we revealed significantly higher frequency of university education among the MAOA-L males (Kiive et al., Reference Kiive, Laas, Akkermann, Comasco, Oreland, Veidebaum and Harro2014), and it may be relevant that the study is conducted in a traditionally highly educated region. The experiment by Frydman et al. (Reference Frydman, Camerer, Bossaerts and Rangel2011) that addressed financial decision-making under pressure revealed that students with the MAOA-L genotype did make riskier, but also more successful choices. Again, it may be relevant that the present subjects seem to have represent a socioeconomically and intellectually advantaged background.
At variance with MAO-B, regulation of MAO-A activity appears as highly prone to epigenetic control by G×E (Naoi et al., Reference Naoi, Maruyama, Shamoto-Nagai and Riederer2025). High perceived parental care could mitigate the increase in impulsivity by stress in female MAOA-L allele carriers (Kinnally et al., Reference Kinnally, Huang, Haverly, Burke, Galfalvy, Brent, Oquendo and Mann2009), and it is conceivable that its effect was epigenetically mediated, as MAOA methylation is sensitive to psychotherapy (Schiele et al., Reference Schiele, Ziegler, Kollert, Katzorke, Schartner, Busch, Gromer, Reif, Pauli, Deckert, Herrmann and Domschke2018). In the ECPBHS males, methylation at an MAOA locus that was associated with antisocial behaviour correlated with impulsivity in an age, stress and MAOA genotype related manner. Thus, the correlation between the adaptive aspect of impulsivity and MAOA methylation in MAOA-L males from adverse home environments increased from 15 to 18 to 25 years of age (Kanarik et al., Reference Kanarik, Sakala, Matrov, Kaart, Roy, Ziegler, Veidebaum, Lesch and Harro2024). In contrast, maladaptive impulsivity had at age 18 a positive correlation of MAOA-L genotype with inferior family relationships and a negative correlation of MAOA-H with superior home environment, while both of these associations disappeared by age 25. Altogether these dynamics of MAOA methylation spur an hypothesis that it may serve as a marker for adaptive developmental neuroplasticity in the MAOA-L genotype, prone to aggression in less favourable environments.
Another functional genetic polymorphism is responsible for synaptic dopamine: in the gene encoding catechol-O-methyl transferase (COMT), an SNP results in the Val158Met (rs4680) variation, resulting in higher enzymatic activity and correspondingly lower dopamine levels with each Val-allele (Chen et al., Reference Chen, Lipska, Halim, Ma, Matsumoto, Melhem, Kolachana, Hyde, Herman, Apud, Egan, Kleinman and Weinberger2004). Activation by unpleasant stimuli in the prefrontal cortex, amygdala and dorsal hippocampus increased with each Met-allele (Smolka et al., Reference Smolka, Schumann, Wrase, Grüsser, Flor, Mann, Braus, Goldman, Büchel and Heinz2005), but the COMT Val158Met alleles have opposite effects in neural activation during executive and emotional tasks (Mier et al., Reference Mier, Kirsch and Meyer-Lindenberg2010). Studies on global brain functional connectivity have yielded heterogeneous outcomes, wherein association of the Val-allele trends with greater resting-state connectivity and the Met-allele with greater task-based connectivity (Morris et al., Reference Morris, Grace, Woods, Dean and Rossell2020). Attempts to link the COMT rs4680 to distinct psychopathologies or traits have yielded some statistically significant findings, but again, overall the associations are weak, dependent perhaps on the environments of a particular investigation, and possibly on the coping strategies available there. The COMT genotype may relate to the preferable strategy, if this hypothesis is correct (Morris et al., Reference Morris, Grace, Woods, Dean and Rossell2020): that the Val-allele is associated with improved cognitive flexibility allowing integration of novel relevant stimuli, and the Met-allele allows improved sustained attention and targeted neural processing, particularly between limbic regions and prefrontal cortex. In the ECPBHS, either allele was related to psychiatric conditions by age 25: the Met-allele homozygous participants had a higher occurrence of alcohol use and substance use disorders while Met-allele carriers had a significantly higher occurrence of agoraphobia; the occurrence of panic disorder was higher in female Met-allele carriers than in Val/Val homozygote females (Kiive et al., Reference Kiive, Kurrikoff, Veidebaum and Harro2024b). In contrast, the occurrence of generalised anxiety disorder was higher in Val/Val females when compared to Met-allele carriers, and Val/Val homozygous females had a higher occurrence of eating disorders than the Met-allele carriers. While interesting, this dissociation of vulnerability by the COMT genotype remains to be explained, whereas two other associations point at genotype-dependent coping strategies. COMT Val158Met heterozygocity, presumably representing the optimal prefrontal dopaminergic capacity, was associated in a gender-dependent manner with university education, heterozygous females being the most and heterozygous males the least educated group (Lehto et al., Reference Lehto, Akkermann, Parik, Veidebaum and Harro2013). This finding was similar in the parents of the ECPBHS target subjects (Kurrikoff et al., Reference Kurrikoff, Kaarma, Tooding, Vaht, Tulviste, Veidebaum and Harro2018). Quite consistently with the ‘warrior-worrier-hypothesis’ (Stein et al., Reference Stein, Newman, Savitz and Ramesar2006), COMT Val158Met Val/Val homozygotes were overrepresented among enterprising occupations and the Val-allele carriers among self-classified managers, while several factors associated with this type of career selection were different in males and females (Kurrikoff et al., Reference Kurrikoff, Kaarma, Tooding, Vaht, Tulviste, Veidebaum and Harro2018).
A commentary of sex and gender
Evolutionary models for sex differences in disease risk have posited that alleles conferring higher risk in one sex may be protective in the other, and several such polymorphisms have been identified (Harper et al., Reference Harper, Janicke and Morrow2021). The collective findings in the selection of ECPBHS reports presented above, and others in the full bibliography (https://www.tai.ee/et/teadustoo/eliktu-artiklid), suggest a vast difference between males and females in G×E. This is likely to derive from differences in their respective biopsychosocial developmental curves. The highly representative sample, with minimal gender-related selection bias, may have helped reveal the contrasts. It may also be relevant that Estonia has the largest gender gaps in Europe with regard to education level (in favour of females; OECD, 2025) and income (in favour of males; Boll & Lagemann, Reference Boll and Lagemann2018).
Some of the gender differences in G×E may result from exposure. For example, higher aggressiveness of the A/A homozygous subjects of the HCRTR1 Ile408Val genotype was observed in females with higher experience of stressful life events, while in males, the A/A group was more aggressive irrespective of life event scores (Harro et al., Reference Harro, Laas, Eensoo, Kurrikoff, Sakala, Vaht, Parik, Mäestu and Veidebaum2019). Yet, male A/A subjects who become aggressive may be more sensitive to life events that they do not report, or remain out of the scope of the traditional wording of questionnaires.
A consistent finding in studies that have measured platelet MAO activity is the higher average enzyme activity in females. This ∼10% difference has been confirmed in the first pertinent study on birth cohort representative samples, the ECPBHS (Harro et al., Reference Harro, Eensoo, Kiive, Merenäkk, Alep, Oreland and Harro2001), while the distribution curves in males and females largely overlap (Harro & Sakala, Reference Harro, Sakala, Martin, preedy, Patel and rajendram2025). Low platelet MAO activity has been associated with risky and law-breaking behaviours only in males, even though the meta-analysis of the association of low platelet MAO activity with the Zuckerman construct of sensation seeking revealed similar effect size for males and females (Winfield et al., Reference Winfield, Mendez and Frietze2025). Seriousness of the breach of law may confound gender comparisons (Oreland et al., Reference Oreland, Nilsson, Damberg and Hallman2007), and quantification of expression of antisocial behaviour can reveal low platelet MAO activity in females (Sakala et al., Reference Sakala, Katus, Kiive, Veidebaum and Harro2023). Thus, regression analysis with antisocial behaviour at two levels of frequency plus consideration of self-reported use of illicit drugs (that for most individuals remains unknown to police) either in a single occasion or repeatedly, demonstrated ‘dose-dependency’ in the relationship of antisocial behaviour and platelet MAO activity. Females, although few, with repeated police contacts had low platelet MAO activity, and even lower if multiple police contacts occurred together with repeated illicit drug use. The serotonin system in females may be more resilient to early stressors, but potentially vulnerable to additional psychosocial adversities at later stages such as adolescence. For example, specifically in female adolescents, the risk of alcohol-related problem behaviour was higher if platelet MAO activity in the lowest quartile was associated with unfavourable environment (Nilsson et al., Reference Nilsson, Wargelius, Sjöberg, Leppert and Oreland2008), and in the ECPBHS, low platelet MAO activity associated with high-risk traffic behaviour during adolescence specifically in girls (Eensoo et al., Reference Eensoo, Harro, Pullmann, Allik and Harro2007; Luht et al., Reference Luht, Eensoo, Tooding and Harro2018). Such a notion also agrees with the association of early-age low cholesterol levels with maladaptive impulsivity specifically in males (Tomson-Johanson et al., Reference Tomson-Johanson, Kaart, Kiivet, Veidebaum and Harro2020). Low cholesterol levels may in turn contribute to a decrease in brain serotonergic neurotransmission via altered microviscosity (Engelberg, Reference Engelberg1992). Also, mouse studies suggest that females become atypically aggressive if the loss of serotonin is profound (Kästner et al., Reference Kästner, Richter, Urbanik, Kunert, Waider, Lesch, Kaiser and Sachser2019).
For maladaptive impulsivity measures, girls appeared to be more sensitive to low levels of warmth in the family, while this was restricted to the 5-HTTLPR s-allele carriers (Paaver et al., Reference Paaver, Kurrikoff, Nordquist, Oreland and Harro2008). In males, the s-allele related to higher maladaptive impulsivity regardless of family atmosphere. This finding resembles the sexually dimorphic response of the hormonal stress axis among rhesus macaques, where male s-allele carriers of the rh5-HTTLPR had a higher response to stress, but in case of females, experience of early adversity was additionally required (Barr et al., Reference Barr, Newman, Schwandt, Shannon, Dvoskin, Lindell, Taubman, Thompson, Champoux, Lesch, Goldman, Suomi and Higley2004).
Several further candidate genes had a rather gender-selective association with the phenotypes of interest even without consideration of life events, such as the link between NOS1 ex1f-VNTR and aggressiveness (O‘Leary et al., Reference O’Leary, Laas, Vaht, Kiive, Veidebaum, Reif and Harro2020) in males, or CCKB receptor gene (rs2941026) and anxiety-related measures (Lvovs et al., Reference Lvovs, Matrov, Kurrikoff, Veidebaum and Harro2022) in females. Since development and genotype may impact the perception, processing and interpretation of environments then any genotype effect is a G×E effect. This notwithstanding, sex/gender differences appeared substantively in representative samples. Thus the road toward precision medicine should earnestly examine these differences in epidemiological factors and pathogenesis (Shi et al., Reference Shi, Ma, Li, Liu, Liu, Chen, Liu, Liu and Huang2024).
The alternative sensitivity hypothesis
The notion of ‘vulnerability genes’ soon received a competitor concept of ‘plasticity genes’ (Belsky et al., Reference Belsky, Jonassaint, Pluess, Stanton, Brummett and Williams2009), according to which the much studied common gene variants such as the s-allele of the 5-HTTLPR would convey higher sensitivity to the environment (Delli Colli et al., Reference Delli Colli, Borgi, Poggini, Chiarotti, Cirulli, Penninx, Benedetti, Vai and Branchi2022). Some support for this concept has emerged from animal studies focusing on the serotonin transporter, that reveal certain advantages of partial deficit of its function in more beneficial environments (Kästner et al., Reference Kästner, Richter, Lesch, Schreiber, Kaiser and Sachser2015). While it is plausible that certain functional variants of genes encoding key proteins make the person more malleable by some form of impactful stimuli, the concept appears to require refinement. Indeed, to say that some gene variants convey sensitivity to environment implicitly suggests that the other corresponding variants confer insensitivity to environment, which, in a Darwinian sense, may not be realistic for highly prevalent alleles. An alternative or rather progressing hypothesis to explain the contradictory findings would suggest the alternative alleles at a functionally important locus are responsive to different types of impacts in the same total environment. In one of the first two-generation analyses of the ECPBHS, risky behaviour in the 5-HTTLPR s’-allele carriers was sensitive to parental support while behaviour of the l’/l’ homozygoous participants was predicted by their fathers’ higher adaptive impulsivity, which is related to risky traffic behaviour, suggesting a possible mediation by role modelling (Tokko et al., Reference Tokko, Eensoo and Harro2024). Thus, the case of the 5-HTTLPR genotype serves as an example of variable sensitivity to different environmental factors within the family that impact on traffic behaviour in future, and more generally, of how functional variants of a gene can promote similar gross behaviours via different mediating pathways. Indeed, for the 5-HTTLPR, which allele is associated with resilience across divergent resilience definitions, depends on the age of subjects (Cahill et al., Reference Cahill, Chandola and Hager2022). Whether the chronological age itself, or the role that the humans play in their community in an age-dependent manner, is of importance, deserves testing in future studies. The alternative sensitivity model also permits the role of agency. Cybernetic, self-regulatory models of complex behaviour that interpret temperament/personality traits acting together (Van Egeren, Reference van Egeren2009) require that their mode of response to the environment included agency, an active engagement with external stimuli. Agency here is the ability to act, including the individual acting differently than predicted by gross models. Experimental studies in animals, using models such as social defeat have demonstrated that differences both in behaviour before a social conflict as well as during the social conflict itself can influence subsequent learned responses and brain activation patterns (Walker et al., Reference Walker, Masters, Dielenberg and Day2009). Hence the gene×environment interaction models appear insufficient for precision medicine, and gene×environment×action interaction models will be required (Harro, Reference Harro2010; Harro & Sakala, Reference Harro, Sakala, Martin, preedy, Patel and rajendram2025).
Strengths, limitations and opportunities
ECPBHS/EstChild has collected through five study waves a multidisciplinary database, including biosamples. Its strengths include high representation of the eligible cohorts, low attrition, and rigorous data collection. The recruitment procedure allows comparison of birth cohorts, born and developing during rapid socioeconomic transition. The large gender gaps unveiled in education and income – in opposite directions – in the country of study invite respective analyses and further scrutiny. Involvement of parents in the study enables two-generation models. The data of the ECPBHS are available to other researchers upon any reasonable request. As this is an ongoing longitudinal study, the data are maintained in a pseudonymised form; for external researchers, datasets are released in an anonymised format to ensure full protection of confidentiality. Access to the ECPBHS data requires submitting a formal data request, and requires valid approvals from the relevant Research Ethics Committee(s). Researchers must provide documentation confirming that their planned analyses meet ethical and data-protection regulations.
An obvious limitation here is that the longitudinal design with laboratory assessments has restricted the sample size. Pooling data of similar studies and replication testing are important for validation of the findings, but given the complexity of environmental variables, true reproducibility testing requires strict attention to the salience of critical features in ’enviromics’. The caveat that causality can not be inferred from statistical associations, even if made with longitudinal design, should also be borne in mind.
Concluding notes
The ECPBHS/EstChild study has helped to interpret several controversial findings in biological psychiatry with its multidisciplinary deep phenotyping, consideration of brain-body connection and the developmental stages of behaviour. It has appeared that common biomarker effects can be understood in the context of adaptive strategies, that are subject to gene×environment interactions that are gender dependent. Further study of these birth cohorts will need to harness family aggregation effects in behaviour and mental health.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/neu.2026.10072.
Acknowledgements
ECPBHS/EstChild has been made possible by contributions from a large number of people and several institutions. Research grants from the Estonian Ministry of Education and Science, the Estonian Science Foundation and the Estonian Research Council are gratefully acknowledged. ECPBHS also received support through the Estonian Centre of Behavioural and Health Sciences. Partial funding came via partnerships in the EU 7th Framework Programme project Aggressotype (No 602805) and the Horizon 2020 projects CoCA and Eat2beNICE (No 667302 and 728018, respectively). We are particularly grateful to the participants of the ECPBHS and their parents, to the whole ECPBHS Team, to our many collaborators, and to the many colleagues who have advised us on specific aspects of materials and methods. Professor Illimar Altosaar kindly reviewed this manuscript for language and style. The ECPBHS would not have happened without the initiative, energy and dedication of Dr Maarike Harro (1960–2006).
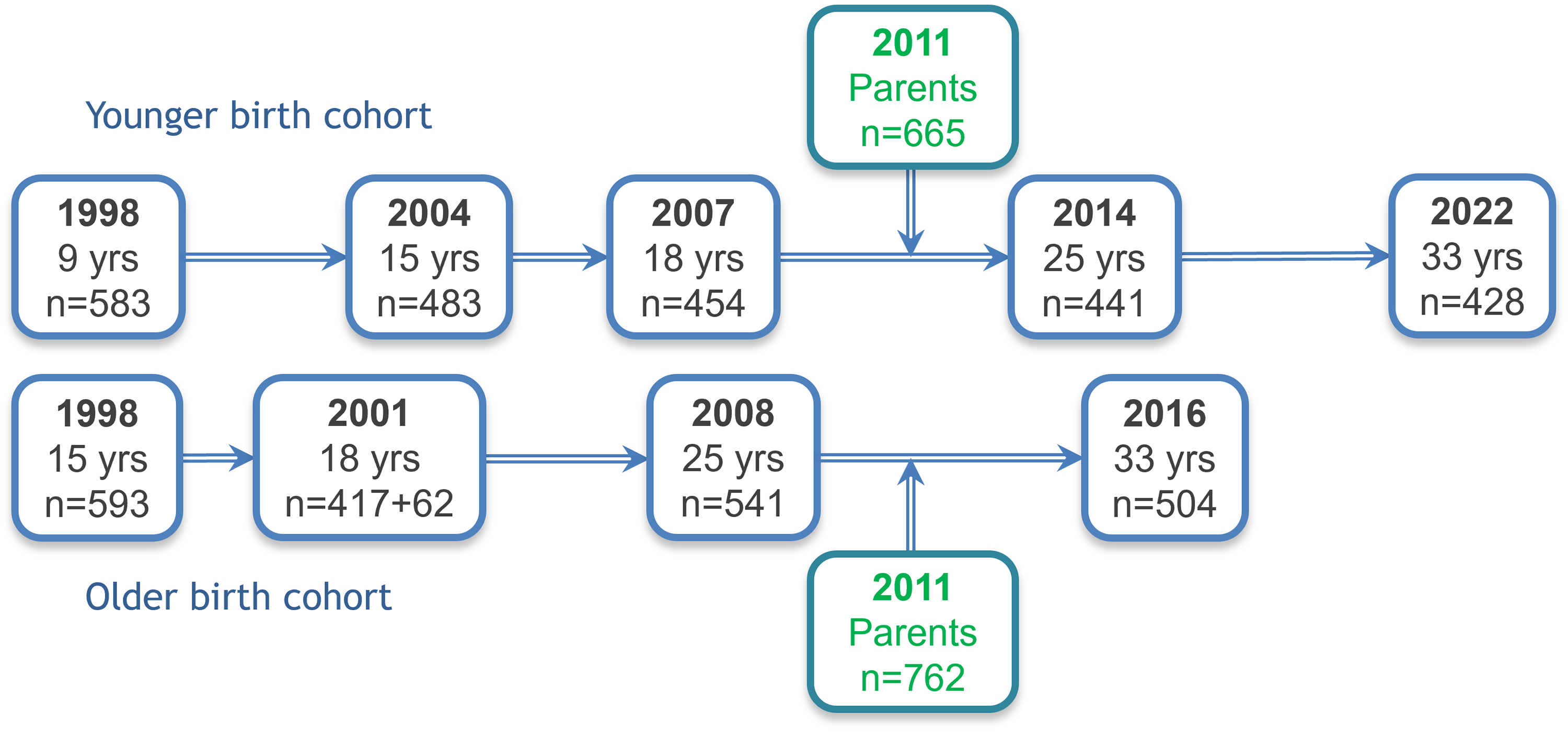
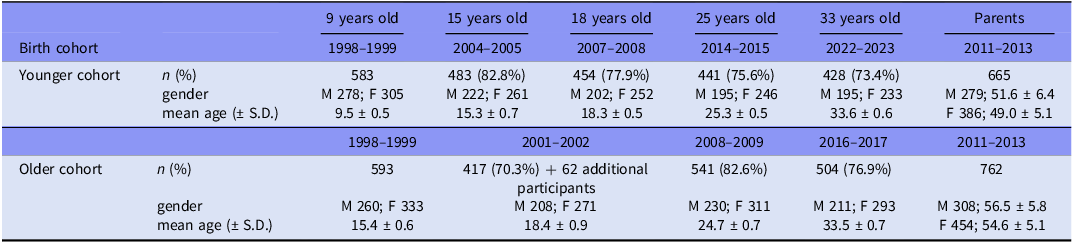
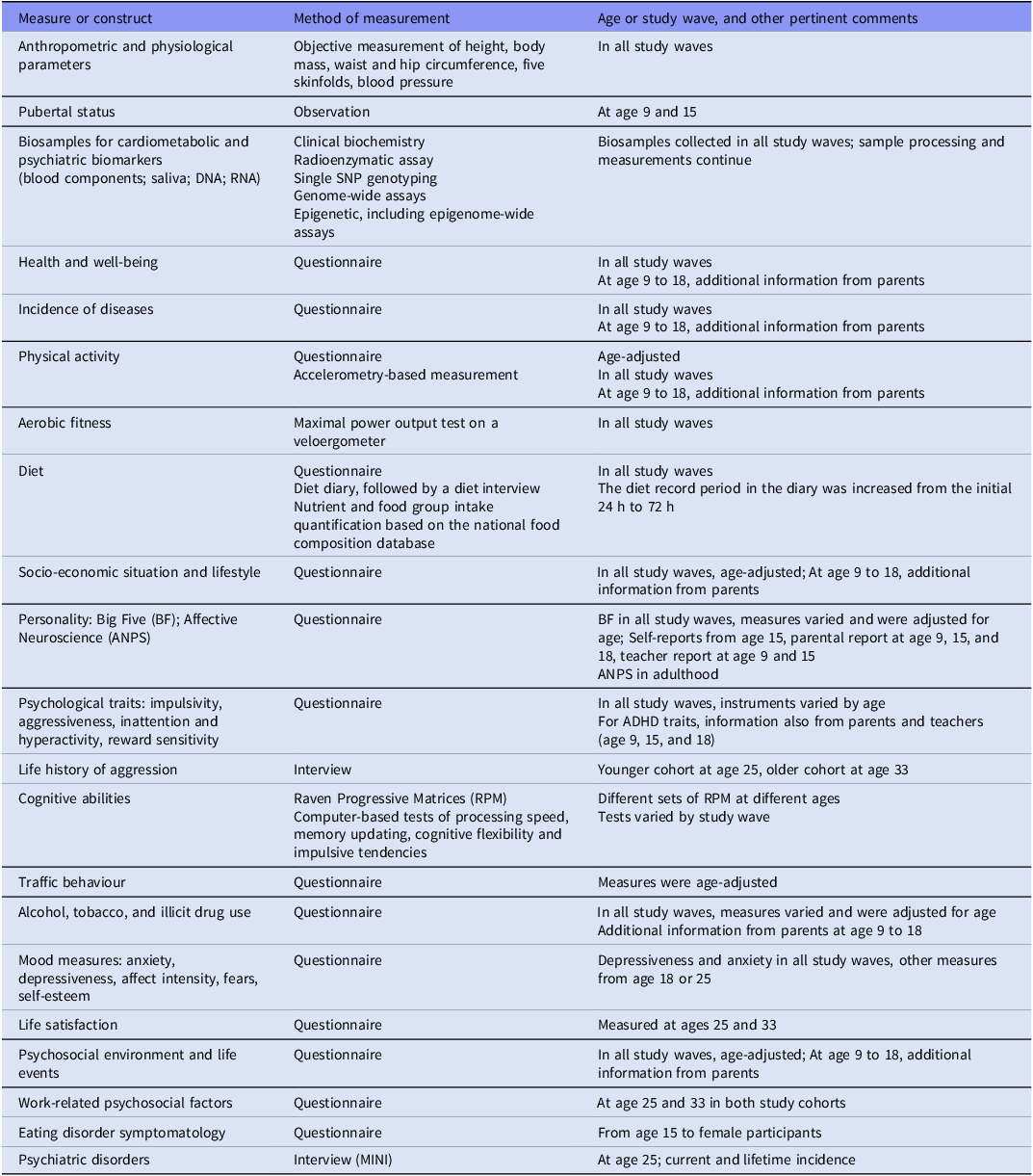
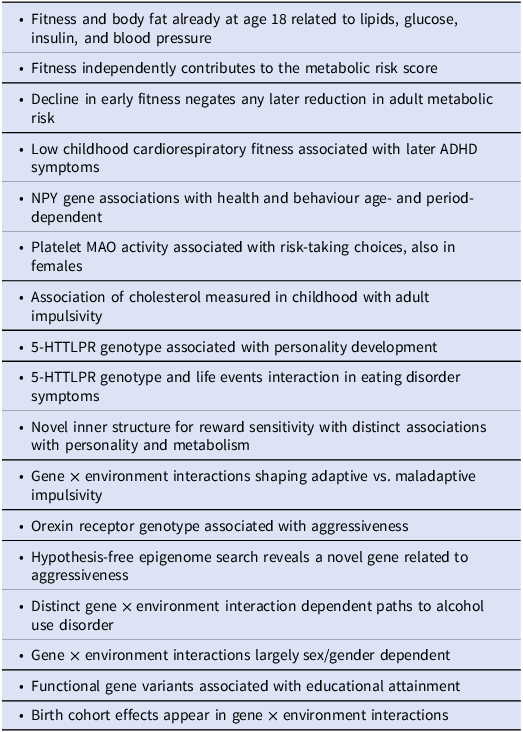



 Open access
Open access