


Introduction
Antarctica is the only continent in the world without an Indigenous human population, and the parts of the continent permanently occupied by humans are limited to a small number of year-round research stations and logistics hubs. Despite the relatively limited nature of human presence on the continent, studies have shown that even minor human activity can have negative impacts on the Antarctic environment, particularly its rare and biologically diverse ice-free areas. These impacts range from chemical contamination (Kennicutt II et al. Reference Kennicutt, Klein, Montagna, Sweet, Wade and Palmer2010) and trampling and compaction of soils (Ayres et al. Reference Ayres, Nkem, Wall, Adams, Barrett and Broos2008, O’Neill et al. Reference O’Neill, Balks, López-Martínez and McWhirter2012) to the introduction of invasive species (Duffy & Lee Reference Duffy and Lee2019) and the deposition of black carbon on the landscape (Khan et al. Reference Khan, Jaffé, Ding and McKnight2016). Such impacts are also often seen as threatening Antarctica’s wilderness values, resulting in efforts to map the spatial coverage of the human footprint (Hughes et al. Reference Hughes, Fretwell, Rae, Holmes and Fleming2011, Brooks et al. Reference Brooks, Jabour, van den Hoff and Bergstrom2019, Leihy et al. Reference Leihy, Coetzee, Morgan, Raymond, Shaw and Terauds2020). A key challenge facing both environmental stewardship and ecological research in Antarctica is distinguishing contemporary environmental change from the lingering biological legacies of past human activity.
There is an implied temporal dimension to much of the human impact research in Antarctica. This reflects an underlying assumption that the environmental conditions of Antarctica may slow ecological recovery following disturbance from human impacts (e.g. very little water available to leach chemicals from the soils). However, few studies have sought to assess the temporal dimensions of human activities and their associated disturbances in this region. This lack of research is probably due to several factors that make it difficult for such research to be carried out. First is that, once begun in a particular place, human activities often tend to continue, meaning that it is difficult to attribute impact to a particular historical disturbance. Other sites of human activity have undergone extensive ‘cleanup’ operations upon decommissioning, which makes an assessment of the temporal dimensions of human impact more difficult (Taylor et al. Reference Taylor, Webster-Brown and Hawes2016). In contrast, structures built before the 1950s are often left in place as Antarctic ‘heritage’, which makes assessing the underlying soils difficult.
The McMurdo Dry Valleys (MDVs) region in East Antarctica offers a useful location for overcoming these barriers and assessing the legacy of the impact of human activities. The MDVs are a relatively bounded space - the largest ice-free area on the continent - and have a long history of human activity (Howkins et al. Reference Howkins, Chignell, Gullett, Fountain, Brett and Preciado2020, Chignell et al. Reference Chignell, Howkins, Gullett and Fountain2022). MDV soils are sensitive indicators of the ecological consequences of human activity (Campbell et al. Reference Campbell, Claridge and Balks1998a, Balks et al. Reference Balks, Paetzold, Kimble, Aislabie and Campbell2002, Aislabie et al. Reference Aislabie, Chhour, Saul, Miyauchi, Ayton, Paetzold and Balks2006, Ayres et al. Reference Ayres, Nkem, Wall, Adams, Barrett and Broos2008, O’Neill et al. Reference O’Neill, Balks and López-Martínez2013). Extant communities that have not been disturbed often serve as a baseline for evaluating the type and magnitude of change in response to disturbance (e.g. foot traffic or climate variation; Campbell et al. Reference Campbell, Balks and Claridge1993, Reference Campbell, Claridge, Campbell, Balks and Priscu1998b, Barrett et al. Reference Barrett, Virginia, Wall and Adams2008, Ayres et al. Reference Ayres, Nkem, Wall, Adams, Barrett and Broos2008, Andriuzzi et al. Reference Andriuzzi, Adams, Barrett, Virginia and Wall2018). The nature of human activities in the MDVs ranges from discrete disturbance events (walking) to prolonged settlement (occupying a camp over multiple research seasons). Human activities can compact soils, disrupt sediments, increase the movement of materials around the valleys and inadvertently promote the introduction of nutrients, chemicals, and non-native biota to the MDVs. Soils therefore offer a potential record of the impacts of these past activities.
Human activity began in the MDVs in December 1903 when a British sledge party led by Captain Robert Falcon Scott encountered a predominantly ice-free valley on their return from a reconnaissance expedition to the East Antarctic Ice Sheet (Scott Reference Scott1905). Since the International Geophysical Year (IGY) of the late 1950s, the MDVs have been an important location for scientific research. Over the last several decades, 16 semi-permanent scientific field camps have been constructed in the region by the USA and New Zealand (Priscu & Howkins Reference Priscu and Howkins2016, Chignell et al. Reference Chignell, Myers, Howkins and Fountain2021). Seven of these semi-permanent field camps have since been removed, and five of these remain accessible (two have since been inundated by rising lake levels). Additionally, there are at least three sites associated with the Dry Valley Drilling Project (DVDP) of the early to mid-1970s (see McGinnis Reference McGinnis1981, Torii Reference Torii1981), which are currently accessible by foot and provide examples of short-term but extensive disturbance.
In Antarctic terrestrial environments, assessments of disturbance and recovery have often relied on visual indicators of surface condition. While these approaches provide rapid assessments of environmental impact, they may not capture longer-term biological responses within soil ecosystems. Soil invertebrates, particularly nematodes, integrate multiple environmental stressors and therefore offer a sensitive biological signal of ecological change. Examining their responses to historical disturbances provides an opportunity to evaluate whether the apparent recovery of surface features corresponds to recovery of underlying soil communities.
In this study, we combine approaches from the fields of environmental history and soil ecology to examine the interactions between human activity, scientific research and environmental change in the MDVs over the past 50 years. Given the truncated food web, delicate soil structure and low biomass of the system, we hypothesize that research facilities constructed in the region will leave an imprint on the landscape long after the facilities have been removed. Specifically, we explore the following research questions: 1) Is there evidence of enduring ecological impacts on soils at sites of former research facilities? 2) Is there evidence of recovery from historical human disturbances in soil ecosystems of the MDVs? 3) Does recovery differ as a result of the type and duration of human disturbance? We conclude by offering ecological explanations for these patterns and discussing their implications for environmental management and conservation in the MDVs and other ice-free areas of Antarctica.
Methods
Site identification
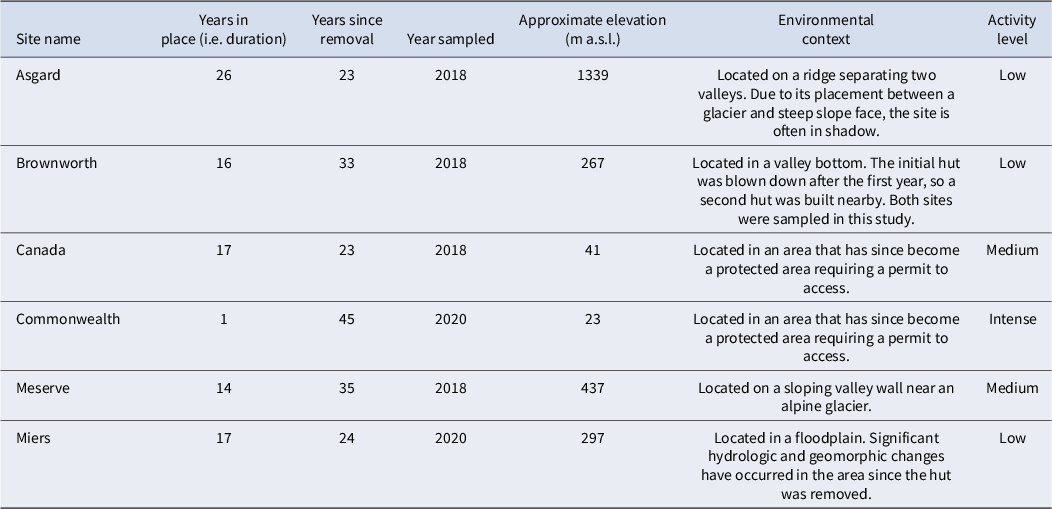
Historical data provide valuable information on the location, duration and intensity of human presence in the MDVs. We used archival sources and historical photographs to field-locate the sites of all historical huts that have been removed from the MDVs (see Howkins et al. Reference Howkins, Chignell, Gullett, Fountain, Brett and Preciado2020, Chignell et al. Reference Chignell, Myers, Howkins and Fountain2021). We then sampled the Asgard, Meserve and Brownworth sites in Wright Valley, the Canada (also known as Fryxell Site of Special Scientific Interest (SSSI)) and Commonwealth sites in Taylor Valley and the Miers site in Miers Valley (Fig. 1). We did not sample the old Bonney Hut or Vanda Station because rising lake levels in the MDVs have fully or partially submerged these sites, and because human impacts on the soils of the latter have been previously studied (O’Neill et al. Reference O’Neill, Balks, López-Martínez and McWhirter2012, Reference O’Neill, Balks and López-Martínez2013, Taylor et al. Reference Taylor, Webster-Brown and Hawes2016, Hawes et al. Reference Hawes, Howard-Williams, Webster-Brown and Poirot2025). The sites we sampled were in place for anywhere between 1 and 26 years and collectively experienced a variety of disturbances including huts, outhouses, helicopter landing pads, a borehole and drilling structure, a campsite and an experimental greenhouse. They are located in a diversity of environmental settings, and as a result our study does not include a true control site. Rather, our sites cover the natural gradient of environments (e.g. Bockheim et al. Reference Bockheim, Prentice and McLeod2008) and range of historical human activities that occur across the MDVs (Table I).
Map of the McMurdo Dry Valleys with locations and historical photographs of the six sites sampled. Photographs used with permission from the Antarctica New Zealand Image Library (https://adam.antarcticanz.govt.nz/).

Names and characteristics of the sites sampled.

Activity level is based on Wharton & Doran (Reference Wharton and Doran1999, p. 18): Low = used sporadically, less than 10 overnight occupations per year; Medium = occupied only for 10–60 days during the field season by groups of < 10; Intense = short period of high-disturbance activity with groups of > 20 (Dry Valley Drilling Project sites).
m a.s.l. = metres above sea level.
Sampling protocol
We carried out field sampling during the 2017–2018 and 2019–2020 summer field seasons. At each site, we identified two to three areas (‘plots’ hereafter) of past human activity (e.g. hut, outhouse, helicopter landing pad, etc.). After sampling the centre point of the plot, we set up transects in three directions (0°, 120° and 240° compass angles). To prevent disturbance of the soils through the process of setting up the plot, we walked adjacent to the sampling zone while unrolling the measuring tape and subsequently moved the tape to align with the correct angle. For each transect, we collected soil samples at positions located 5, 15 and 45 m away from the centre point. We chose these distances in order to capture the gradient of potential change in soil characteristics that would be expected as one moves from areas of greater disturbance to areas with less disturbance. We chose 45 m as the end point (and not further) as a balance between capturing areas considerably far away from the site’s centre, keeping the protocol manageable given limited time in the field and helping to avoid transects intersecting to adjacent unsampleable features such as glaciers or lakes (see later). We took a cluster of three samples at each position along the transect, one at each point of a 1 m-sided equilateral triangle (Fig. 2). We did this in order to capture the natural variability at each position and to mitigate the effects of outliers on subsequent analyses.
Diagram of the study sampling design. ‘Site’ refers to the larger area where the disturbances occurred (e.g. Asgard, Brownworth, etc.); ‘Plot’ refers to each disturbance within a site (e.g. footprint of a removed hut, represented as a grey rectangle in the figure); ‘Plot Position’ refers to the triangular areas along each transect where we collected samples.

We collected samples (0–10 cm depth) using clean scoops (200 cc) and placed them in Whirl-Pak® bags for invertebrate analysis. The total mass of each invertebrate sample was ~750 g, and these were returned to the Crary Laboratory at McMurdo Station in insulated coolers for extraction. We photographed the location of each sample and recorded GPS coordinates with a handheld unit (Garmin GPSMap 78s). We also photographed and marked the coordinates at the centre of each plot. This resulted in 30 samples per plot (N = 420), with the exception of the Meserve outhouse, where field conditions prevented collection of a third transect, and Miers hut, where an extra transect was collected in order to better quantify the variation of the surrounding environment.
We made ad hoc modifications to the sampling design based on specific field conditions. These included adding an additional transect at Miers Hut (at 60°) due to unexpected extra field time, as well as collecting only along the 0° (north) and 120° transects for the Meserve outhouse plot due to less-than-expected field time.
Soil invertebrate extraction and identification
We extracted invertebrates from the soil using a modified sugar centrifugation protocol developed specifically for Antarctic soils (Freckman & Virginia Reference Freckman and Virginia1993). We identified and enumerated invertebrates via light microscopy (Olympus CKX41SF). We identified nematodes to genus (Scottnema, Eudorylaimus, Plectus), by life stage (juveniles or adults), sex and as either living or dead. We measured gravimetric soil moisture by weighing 50 g of soil before and after drying in an oven at 105°C for 24 h. We calculated invertebrate abundance as the number of individual animals per kilogram of dry soil. We determined electrical conductivity (index of ion concentration) of soils in a 1:5 sediment-to-deionized (DI) H2O slurry using a YSI conductivity meter and soil pH in a 1:2 sediment-to-DI H2O slurry using an Orion pH meter.
Statistical modelling
We analysed the data using R statistical software (R Foundation for Statistical Computing 2018). We focused our analyses on Scottnema because it integrates multiple stressors, including human disturbance, soil chemistry, microclimate and geological legacy (Barrett et al. Reference Barrett, Virginia, Lyons, McKnight, Priscu and Doran2007, Poage et al. Reference Poage, Barrett, Virginia and Wall2008). Scottnema abundance, distribution and life-history structure therefore reflect both local environmental conditions (Xue et al. Reference Xue, Thompson and Adams2024) and broader climatic forcing, making Scottnema a powerful bioindicator of both legacy impacts and potential resilience. We plotted Scottnema abundance (living organisms), mortality (dead/live ratio) and fecundity (live juveniles/live females; see Poage et al. Reference Poage, Barrett, Virginia and Wall2008) against distance from plot centre. However, soil invertebrates are sparsely distributed in the MDVs, leading to a prevalence of zero counts across samples. The presence of excess zeroes complicates the application of standard distributions such as the Gaussian or Poisson distributions, resulting in inadequate representation of the data and unreliable inferences. Furthermore, these excess zeroes pose distributional challenges that cannot be normalized through transformations. To address this issue, various statistical techniques have been explored in the literature to handle count data exhibiting excess zeroes. One prominent approach is the zero-inflated model, which is a two-component mixture model that enables the simultaneous evaluation of the impacts of explanatory variables on both the count response and the probability of a zero count occurrence. Zero-inflated models such as the zero-inflated binomial, zero-inflated negative binomial and zero-inflated Poisson models have been successfully used to address the issue of excessive zeroes encountered in count responses in ecological data (Welsh et al. Reference Welsh, Cunningham, Donnelly and Lindenmayer1996, Arab et al. Reference Arab, Wildhaber, Wikle and Gentry2008, Larsen et al. Reference Larsen, Bissonette, Flinders and Robinson2011, Pierce et al. Reference Pierce, Larsen, Flinders and Whiting2011, Martin et al. Reference Martin, Cooper, Price, Turnbull and Roberts2015). The bias in parameter estimates arising from numerous zero counts can be reduced through these approaches. Model selection (Akaike Reference Akaike, Parzen, Tanabe and Kitagawa1998, Burnham et al. Reference Burnham, Anderson and Burnham2002) testing was used in determining the most suitable distribution for the observed counts. For our analyses, we used generalized linear mixed models (glmmADMB package; Fournier et al. Reference Fournier, Skaug, Ancheta, Ianelli, Magnusson and Maunder2012) and the MASS library (Venables & Ripley Reference Venables and Ripley2002) within R (version 4.0.3; www.r-project.org). Hereafter, we refer to the values produced by the zero-inflated model as ‘predicted’ values (e.g. ‘predicted abundance’), which we used in our subsequent plots and statistical analyses.
To examine whether the duration of a disturbance is related to impact, we computed the difference between the mean predicted value of Scottnema abundance at the 45 m (i.e. outermost) plots and the mean value at the 5 m (i.e. innermost) plots at each site, and then we plotted this against the number of years over which each camp was in place (i.e. duration of disturbance at each site). In doing so, we assume that the greater the difference in abundance between the 45 and 5 m plots at the time of sampling, the greater the long-term impact at that site.
Similarly, to explore whether time since camp removal is related to recovery from a disturbance, we again computed the difference between the mean predicted value of Scottnema abundance at the 45 m plots and the mean value at the 5 m plots, and then we plotted this against the number of years since each camp was removed. In doing so, we assume that the longer the time since the camp was removed and disturbances ceased, the more time invertebrate populations would have to recover, and this would be reflected in a smaller difference in mean predicted Scottnema abundance between a site’s outermost and innermost plots.
Geospatial analysis
We converted all GPS data to Esri shapefiles (WGS84 South Pole Lambert Azimuthal Equal Area projection) and imported them into Esri ArcGIS Pro geographical information system (GIS) software (version 2.9). For each soil characteristic, we computed the mean ‘predicted’ value (from the zero-inflated model) of the three samples at each triangular plot position and then joined the resulting values to the corresponding geographical coordinates recorded by the GPS.
To characterize the variation of invertebrates within each plot, we used the ‘Empirical Bayesian Kriging’ tool in ArcGIS Pro Geostatistical Analyst to interpolate new values for the plot based on the averaged sample points. We focused this analysis on the predicted abundance, fecundity and mortality values for Scottnema, as Scottnema is understood to be a key indicator species of change in the low-diversity soils of the MDVs (Freckman & Virginia Reference Freckman and Virginia1997). Empirical Bayesian kriging has been shown to outperform other interpolation methods, even when variation is changing rapidly and dissimilarly in different parts of the data extent (Gribov & Krivoruchko Reference Gribov and Krivoruchko2020). We used the default kriging model and parameters (i.e. data transformation = ‘none’; semivariogram = ‘power’), which generally produce reliable outputs (Krivoruchko & Gribov Reference Krivoruchko and Gribov2019). To prevent extrapolation outside the areas sampled, we constrained the interpolation to areas within a convex hull polygon created by the outer positions of each plot. We set the grid size of the resulting raster map to 5 m, which is the recommended grid resolution for our data based on the positional accuracy of our GPS unit (recorded in the field as ± 2.74 m; Hengl Reference Hengl2006).
Results
Soil pH (6.8–10.9) and electrical conductivity (7–3550 μS/cm) were typical of the range of values reported for MDV soils. Soil moisture content ranged from 0.44% to 19.6% by weight and skewed to wetter soils than typical for the MDVs for all sites except for the Commonwealth sites. These higher-than-expected soil moisture values may be related to the preferred locations and landscapes selected for field camp placement in the past.
We did not identify any non-native invertebrates in the samples. Soil invertebrate communities consisted of three commonly observed nematode taxa (Scottnema lindsayae, Eudorylaimus spp. and Plectus spp.), rotifers and tardigrades, which were only identified to phylum. Scottnema was the most abundant and widely distributed soil animal, as has been previously reported, ranging in abundance from 0 to 5319 individuals per kg dry soil, and occurring in 59% of samples. Eudorylaimus, Plectus and tardigrades were present in less than 10% of samples, with abundances ranging from 0 to 500 individuals per kg dry soil (i.e. approximately an order of magnitude less dense than Scottnema). Rotifers were also patchily distributed, generally co-occurring with Plectus and tardigrades, but with population densities of up to 6230 individuals per kg dry soil.
Ten of the 14 plots have the highest predicted Scottnema abundance at the most distant plot position (45 m; Fig. 3). The results of the geospatial analysis show a pattern of lower predicted Scottnema abundance in disturbed areas, with this being the case for 12 of the 14 plots (Fig. 4). The exceptions are the Meserve Helicopter Pad, which may be in a favourable location for nematode habitat (i.e. adjacent to a glacier and in a topographical depression, and thus slightly more sheltered), and the Miers Hut, which may have been significantly altered due to its location in a flood plain (i.e. dynamic changes in water, sediment and soil structure). Contrary to abundance, we did not observe any consistent patterns in fecundity (Figs S1 & S2) or mortality (Figs S3 & S4) across sites. Previous studies have used the combination of high fecundity, low mortality and high abundance as an indicator of a ‘healthy’ nematode population. Our results show that two of these characteristics are missing at every site, and that fecundity is low at all sites.
Predicted abundance by distance from plot centre for Scottnema at each site. Helo = helicopter pad.

Map of predicted abundance values for Scottnema at each site, overlaid (30% transparency) on a digital elevation model derived from Fountain et al. (Reference Fountain, Fernandez-Diaz, Obryk, Levy, Gooseff and Van Horn2017), provided by the Polar Geospatial Center (PGC) and National Center for Airborne Laser Mapping (NCALM) via ArcGIS Online (with the exception of Asgard, which was not covered by this dataset and is therefore overlaid on a simple grey background). Any negative values predicted by the interpolation method have been displayed as a value of 0 in the maps.

The difference of mean predicted Scottnema abundance between outermost and innermost plots had a weak positive relationship with duration of disturbance at the site level (Fig. 5). This relationship holds when the Meserve and Miers sites are considered as outliers and removed, as well as when the data are disaggregated to the plot level (Fig. S5).
Difference of mean predicted Scottnema abundance values between outermost (45 m) and innermost (5 m) plots at each site vs duration of disturbance. Dotted blue line represents the linear trend line fit to the data.

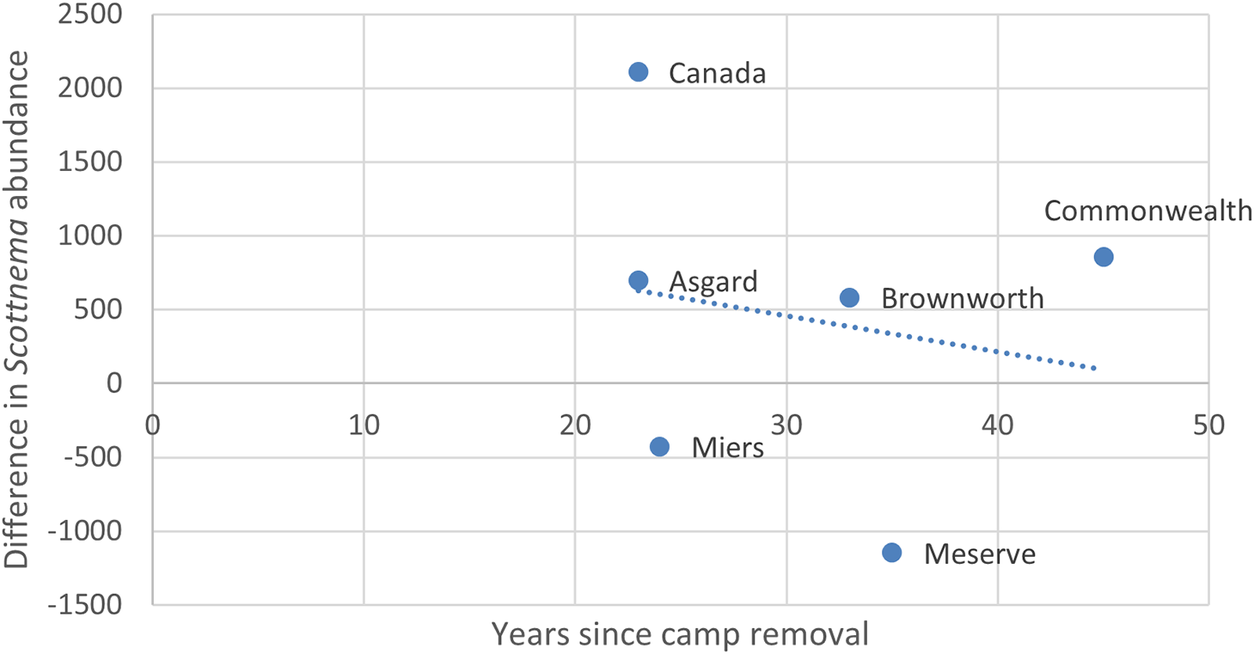
The difference of mean predicted Scottnema abundance between outermost and innermost plots had a strong negative relationship with years since camp removal (Fig. 6). Again, this relationship holds when the Meserve and Miers sites are considered as outliers and removed, as well as when the data are disaggregated to the plot level (Fig. S6).
Difference of mean predicted Scottnema abundance values between outermost (45 m) and innermost (5 m) plots at each site vs years since camp removal. Dotted blue line represents the linear trend line fit to the data.

Discussion
Our results show signs of invertebrate recovery at many sites, with the clearest examples being the Canada Hut and Helicopter Pad as well as the Commonwealth Campsite. This could be due to the specific types of disturbances at these sites, as well as their respective locations in the valleys. Brownworth also shows potential invertebrate recovery, although the differences are small. Interestingly, our results do not show a major difference in abundance, fecundity or mortality between the two huts at the Brownworth site. This is surprising given that the first hut was only in place for 1 year, and the second was in place for 15 years.
However, both the Canada Greenhouse and Commonwealth Borehole were plots with unique and particularly intense disturbances, and they show no signs of recovery of nematode populations after 23 and 45 years, respectively. Additionally, the Asgard and Meserve sites have low invertebrate abundance, low fecundity and low mortality, which do not quite match what we expect as signs of recovery. This may be related to their specific environmental factors, as both sites are located higher up in their respective valleys, are often in shadow, and they are in close proximity to a glacier.
Disturbance legacies in the MDVs persist longer than surface evidence alone would suggest. Across the sites we examined, nematode populations were consistently lower near areas of historical infrastructure, even decades after disturbance ceased. These results indicate that human activities associated with field camps can leave detectable ecological signatures in soil communities long after structures have been removed and landscapes appear visually recovered.
These patterns also indicate that not all disturbances are equal. Previous work on trampling, vehicle tracks and field camp disturbance has shown that recovery trajectories vary widely depending on disturbance intensity, substrate characteristics and geomorphological context (Campbell et al. Reference Campbell, Balks and Claridge1993, Reference Campbell, Claridge and Balks1998a, Ayres et al. Reference Ayres, Nkem, Wall, Adams, Barrett and Broos2008, O’Neill et al. Reference O’Neill, Balks and López-Martínez2013, Reference O’Neill, Aislabie, Balks and Bockheim2015). Our results extend this literature by demonstrating that even after structures have been removed and sites appear recovered, nematode populations may remain depressed for decades. Visual assessments have long been used as rapid indicators of environmental impact in Antarctic terrestrial environments (Campbell et al. Reference Campbell, Balks and Claridge1993), yet surface morphology may recover on timescales that differ from underlying soil physical and biological processes (Campbell et al. Reference Campbell, Claridge and Balks1998a, O’Neill et al. Reference O’Neill, Aislabie, Balks and Bockheim2015). Our results show that sites where historical infrastructure is no longer visibly apparent may still exhibit substantially reduced nematode populations decades after disturbance has ceased. The biological signal recorded by nematode communities therefore provides information that is independent of aesthetic restoration of the landscape. The sites we examined illustrate how soils can appear stable and undisturbed while nematode populations remain substantially reduced relative to surrounding areas. This decoupling of surface appearance and ecological structure and functioning has direct implications for environmental monitoring and management decisions that rely heavily on visual inspection.
Although our disturbance chronosequence does not include sites disturbed within the past 2 decades, this reflects both the availability of appropriate legacy sites and the specific objective of this study. Previous research in the MDVs has already examined the short-term ecological consequences of disturbance, including trampling and vehicle tracks, and has demonstrated measurable impacts on soil structure and biological communities within years to decades following disturbance (Campbell et al. Reference Campbell, Balks and Claridge1993, Reference Campbell, Claridge, Campbell, Balks and Priscu1998b, Ayres et al. Reference Ayres, Nkem, Wall, Adams, Barrett and Broos2008, O’Neill et al. Reference O’Neill, Balks and López-Martínez2013). Our study instead focuses on sites where disturbance has ceased and infrastructure has been removed, allowing us to examine the persistence of ecological legacies several decades later. Including more recent disturbances would have introduced an active recovery signal that could obscure the long-term legacy patterns we sought to isolate. By focusing on sites where human activity ended more than 2 decades ago, we are able to evaluate whether biological systems return to baseline conditions once physical structures are removed and surface disturbance appears visually recovered. This perspective is particularly relevant for environmental management because infrastructure removal is often treated as the endpoint of disturbance, yet our results suggest that ecological recovery may operate on substantially longer timescales.
In lower-intensity disturbances such as temporary huts or helicopter pads, patterns at the Canada and Commonwealth sites suggest gradual recolonization from adjacent undisturbed soils, consistent with metapopulation-like dynamics described for Antarctic soil fauna (Sokol et al. Reference Sokol, Brown and Barrett2017). In contrast, the borehole and greenhouse plots represent disturbances that probably altered soil structure and geochemistry sufficiently to push systems beyond ecological thresholds that allow rapid ecological recovery. Similar substrate- and disturbance-specific recovery dynamics have been documented for hydrocarbon remediation sites (Roura Reference Roura2004) and for foot traffic impacts in Wright Valley soils (McLeod Reference McLeod2012). These comparisons further suggest that disturbance type and environmental context, rather than duration alone, shape the persistence of ecological legacy effects. Populations can recover, but this resilience is highly context-dependent, being filtered through site-specific geomorphology, hydrological regimes, microclimates and geological and ecological legacies (Barrett et al. Reference Barrett, Virginia, Lyons, McKnight, Priscu and Doran2007, Xue et al. Reference Xue, Adhikari, Ball, Barrett, Miao and Perkes2023). These observations reinforce earlier work showing that surface morphology can recover on timescales that differ from soil physical and biological processes.
The strong negative relationship between recovery and time since camp removal indicates conditional resilience within soils of the MDVs. Given sufficient time and moderate disturbance intensity, our results show that Scottnema populations can partially rebound. However, resilience is a function of geomorphology, hydrological regime, microclimate, geological legacy and time (Barrett et al. Reference Barrett, Virginia, Lyons, McKnight, Priscu and Doran2007). High-elevation, glacier-adjacent sites such as Asgard and Meserve appear less capable of recovery, suggesting that background environmental stress interacts with anthropogenic disturbance to further constrain recolonization. Ecological resilience in this system, therefore, is not a simple function of elapsed time but an interaction between disturbance history and environmental context.
These findings also highlight the value of nematodes, particularly S. lindsayae, as sensitive sentinel taxa in Antarctic terrestrial ecosystems. Nematode responses provide not just a record of past human activity but also an early-warning signal of ecosystem trajectories in the face of future disturbance, including climate change. In landscapes where vascular plants and higher trophic levels are absent, Scottnema occupies a central functional role in regulating microbial communities and nutrient cycling. Persistent suppression of their populations there signals not merely demographic change but also altered ecosystem processes (Barrett et al. Reference Barrett, Virginia, Wall and Adams2008).
Our work has direct consequences for interpreting long-term monitoring datasets. Legacy effects of past human activity may obscure or mimic climate-driven trends. Without explicit recognition of disturbance history, shifts in nematode abundance could be misattributed to contemporary climatic change rather than residual anthropogenic impact. In this sense, nematodes function both as indicators of ecological recovery and as early-warning signals of future environmental change.
Implications for environmental management
Our study provides evidence of enduring impacts on soil ecosystems at sites of former research facilities. This points to the need for Antarctic environmental managers and scientists to think carefully about where and whether to build a new hut or campsite. Our results suggest that seasonal and temporary campsites may be as impactful to soil invertebrates as semi-permanent structures, and that aesthetic recovery through the removal of structures does not necessarily result in ecosystem recovery at the site. Thus, the potential for impacts to Antarctic soil ecosystems will be reduced and avoided by (re)using existing sites and structures when possible. The Brownworth data suggest that even short-duration occupation can produce impacts comparable to longer-term use. From a management perspective, this reinforces the principle that establishing new sites, even temporarily, may impose ecological costs similar to those of semi-permanent infrastructure. Reuse of previously disturbed areas is therefore likely to minimize cumulative biological impact across the landscape.
The persistence of nematode suppression decades after disturbance also raises concerns regarding cumulative impacts. In a landscape where suitable habitats are patchily distributed, repeated occupation of new sites may progressively reduce the availability of undisturbed refugia necessary for recolonization. Over time, such fragmentation could reduce the resilience of soil communities at broader spatial scales. In the MDVs, where soil ecosystems are sparse yet functionally complex, disturbance legacies can persist far longer than the surface evidence suggests. Our findings indicate that visual restoration or removal of infrastructure does not necessarily equate to ecological recovery. Sites that appear geomorphically stable may retain depressed nematode populations decades after disturbance. Management frameworks that rely primarily on visual inspection of surface conditions therefore risk underestimating the persistence of ecological impacts. Incorporating biological indicators into environmental monitoring would provide a more accurate assessment of ecosystem status and recovery trajectory.
It is important to reiterate that natural variation of the material environment plays a role in the initial populations at each site, their resilience to disturbance and their ability to recover from different types of disturbance (Poage et al. Reference Poage, Barrett, Virginia and Wall2008). In cases where a new site is required, it may be beneficial to conduct soil surveys such as ours prior to the establishment of a campsite or hut, followed by monitoring after it has been removed. Such assessments could be integrated into the environmental impact assessments and management protocols of proposed facilities sites. Given their sensitivity and central role in ecosystem function, nematodes provide a practical and informative bioindicator for inclusion in environmental impact assessments and post-disturbance monitoring. Sampling protocols such as those employed here are logistically feasible within existing field operations and can be integrated into long-term monitoring programmes to detect both legacy disturbance and emerging climate-driven change.
Our findings imply that legacy effects of human activity can confound climate change signal detection in these soils/this ecosystem. Long-term monitoring programmes may therefore be interpreting climate-driven changes against a noisy backdrop of historical disturbance legacies. Recognizing and accounting for the signal among the noise (or ‘ghost’ signals) is essential if these ecosystems are to remain useful sentinels of climate change. Failure to account for historical disturbance may lead to misinterpretation of biological trends as climate-driven when they instead reflect unresolved anthropogenic legacies. Explicit documentation of disturbance history and biological baseline surveys prior to site establishment would reduce this risk and improve the interpretive power of long-term ecological datasets.
Our results show some recovery of nematode populations in the decades following an anthropogenic disturbance. This suggests that soil organisms in the MDVs experiencing human impacts may recover with time, depending on the type of disturbance. Whether this is also true of ecosystem function is an open question and potential avenue for future research. The lack of recovery at particularly intense disturbances such as the greenhouse at the Canada site and borehole at the Commonwealth site underscores the potential longer-term impacts on soil ecosystems that these types of human activities may cause. This is an open-ended question and could be characterized and confirmed through sampling of other sites of intense disturbance in the MDVs, such as the boreholes at other DVDP sites.
Our study reveals the soil ecosystems of the MDVs as both fragile and resilient. They carry the evidence of human activity for decades, but they also demonstrate conditional recovery depending on disturbance type, environmental setting and time. Management strategies that ignore these legacy effects risk repeating the mistakes of the past. Conversely, incorporating this knowledge into protected area design, camp siting and monitoring protocols could safeguard soil ecosystems in Antarctica’s ice-free areas while preserving their role as vital sentinels of environmental change.
Measuring the impacts of human activities in places such as the MDVs is important for scientific research and environmental management because such impacts may confound our ability to accurately interpret the subtle but significant effects of climate change in the region and ecosystem responses in general. While the MDVs are often seen as unique and distinct from the rest of Antarctica, the research presented here may be informative for environmental management and conservation planning in other ice-free areas with considerable human footprint and activity. For example, there is increasing attention being given to developing a more systematic and representative system of protected areas in ice-free Antarctica (Coetzee et al. Reference Coetzee, Convey and Chown2017, Hughes & Grant Reference Hughes and Grant2017, Wright et al. Reference Wright, Brooks, Gooseff, Howkins and Chignell2024). If, as our results suggest, there is potential for long-term impacts and also recovery of soil organisms in ice-free areas impacted by human activities, then the usual cost-benefit analysis of environmental impact to scientific output (and the recommendations to mitigate the former) may need revisiting. The design and monitoring of new and existing protected areas in ice-free regions of Antarctica should therefore involve the collection and consideration of data on each site’s environmental characteristics, its past and expected future human activities and how these may interact over time. The approach developed in this study offers a starting point for doing so and could be adapted to studying other key regions of the southern continent. These results further indicate that visual restoration of a site following infrastructure removal should not be assumed to reflect ecological recovery, underscoring the value of incorporating biological indicators such as nematodes into environmental monitoring programmes.
Supplementary material
To view supplementary material for this article, please visit http://doi.org/10.1017/S0954102026100674.
Author contributions
AH, SMC, JEB, MNG, CT-V and BJA conceptualized the research study design and interpreted results. AH, SMC, DR, SP and BJA conducted fieldwork. ER, CT-V, DR, JEB, SP and BJA processed soil samples. SP conducted statistical modelling. SMC conducted the geospatial analysis. AH and SMC led the writing of the manuscript, integrating text and feedback from the other authors. Geospatial support for this work provided by the Polar Geospatial Center under NSF-OPP awards 1043681, 1559691, 2129685 and 2434541
Competing interests
The authors declare none.








 Open access
Open access