


Introduction
Today, giant lacewings (Neuroptera: Polystoechotidae) have four species restricted to the New World (Oswald Reference Oswald1998). In the early Eocene, however, they had four species in Denmark alone, and their fossils are plentiful and diverse in the Okanagan Highlands series of lacustrine shales of British Columbia, Canada, and northern Washington, United States of America, a series of mid- to late Ypresian (Villeneuve and Mathewes Reference Villeneuve and Mathewes2005; Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021) lake basins that extended across about 1400 km, from northern British Columbia, Canada, to northern Washington, United States of America (Archibald et al. Reference Archibald, Greenwood, Smith, Mathewes and Basinger2011). These species were described based on 13 fossil specimens from Okanagan Highlands localities at Quilchena, British Columbia (four species, two of which are named), Horsefly River, British Columbia (one named species), and Republic, Washington (six species, five of which are named; Makarkin and Archibald Reference Makarkin and Archibald2003; Archibald and Makarkin Reference Archibald and Makarkin2006). Here, we describe two new species from two specimens bearing dark wings with a pale fascia from Republic, and we describe a third, based on a less distinctly preserved forewing with similar colouration from the Okanagan Highlands locality McAbee in south–central British Columbia, Canada, treating it as Palaeopsychops sp. indet., the first reported occurrence of the family at that locality.
Material and methods
We examined three fossil Polystoechotidae wings in lacustrine shale: SR 15-001-001 and SR 13-002-001 (counterparts not known) from Republic, Washington, United States of America, and RBCM.EH2017.050.3020 (part) and RBCM.EH2017.050.3043 (counterpart) from McAbee, British Columbia, Canada.
We generally follow the wing terminology of Breitkreuz et al. (Reference Breitkreuz, Winterton and Engel2017), except for details (e.g., spaces, veinlets, traces), the terminology for which follows Oswald (Reference Oswald1993).
Abbreviations of wing characters are as follows: 1r–m, first radius–media crossvein; A1–A3, first to third anal veins; CuA, anterior cubitus; CuP, posterior cubitus; hp, humeral plate; hv, humeral veinlet; M, media; MA, anterior media; MP, posterior media; n, distal nygma; RA, anterior radius; RP, posterior radius; RP1, proximal-most branch of RP; RP2, branch of RP distad RP1; Sc, subcosta.
Contrary character states of compared taxa are noted in square brackets.
Institutional abbreviations are as follows: SR, the Stonerose Interpretive Center, Republic, Washington; and RBCM, the Royal British Columbia Museum, Victoria, British Columbia.
The Republic specimens are housed in the collection of the Stonerose Interpretive Center. The McAbee fossil is housed in the collections of the Royal British Columbia Museum.
Systematic paleontology
Genus Palaeopsychops Andersen, 2001
Palaeopsychops goodwini sp. nov
Zoobank Registration number: urn:lsid:zoobank.org:act:9061A8E9-DA33-4C55-85F3-00506E1031E4 Figure 1
Palaeopsychops goodwini sp. nov. holotype hind wing SR 13-002-001 (only part known): A, photograph, wetted with ethanol; B, drawing, n = expected position of nygma, as indicated by the characteristic curving of RP branches at this position. Both to scale = 5 mm.

Type material. Holotype hind wing SR 13-002-001, found at exposure B4131 of the Klondike Mountain Formation in Republic, Washington by Steven Goodwin, 27 April 2013, and housed in the collection of the Stonerose Interpretive Center.
Diagnosis. HOLOTYPE. Distinguished from all other Palaeopsychops species except P. barthae by distinctive hind wing colouration: dark with pale, broad fascia mid-wing; hind wing length 27.3 mm [P. barthae: 28 mm; all other species about 40–42 mm]. Distinct from the holotype hind wing of P. barthae by fascia slightly angled to anterior margin [P. barthae: perpendicular]; without possible spots in posterior region of wing [P. barthae: with spots]; and A1 dichotomous [P. barthae: pectinate].
Description. Hind wing 27.3 mm long, 12.5 mm wide. Humeral plate well developed but partly preserved. Costal space broad, strongly broadened distad presumed fusion of Sc, RA (fusion not visibly preserved, within fascia). Humeral veinlet probably strongly recurrent, only its basal part preserved. Subcostal veinlets closely spaced, simple in basal portion, then mostly very shallowly forked 1–2 times proximad pale fascia. Subcostal space moderately broad, with no discernable crossveins. Presumed Sc+RA terminating at margin slightly before wing apex; its veinlets mostly dichotomously forked (2–3 times). RA space broadest basally, slightly more than subcostal space; 2–3 oblique crossveins detected basad fascia, other crossveins possible but poorly discernable. RP with approximately 25 branches, none forked before outer gradate series, all forked after outer series, mostly with three forks (rarely 1–2). Two gradate series of crossveins in radial space (inner, outer) nearly parallel, both complete in all preserved areas. Distal nygma clearly indicated by characteristic strongly curved basal-most branches of RP in expected positions proximad inner gradate series. 1r–m long, sinuous, connecting stem of RP near origin of RP1 and base of M. M forked slightly proximad origin of RP1. MA, MP not preserved distally (probably with few branches). Cu divided into CuA, CuP near wing base. CuA pectinate, with 15 long branches, most forked once. CuP dichotomously branched. A1 profusely dichotomously branched. A2 pectinately branched, with five branches, each forked once or twice distally). A3 pectinate, with few branches (two preserved: one shallowly forked, the other deeply forked). Trichosors distinct along entire wing margin (poorly preserved along costal margin). Colouration dark throughout, except pale, broad fascia just distad mid-wing, not perpendicular to RA (anterior end slightly closer to apex than posterior).
Etymology. The specific epithet is formed from the surname of Steven Goodwin, who found the holotype and donated it to the Stonerose Interpretive Center.
Range and age. Exposure B4131 of the Tom Thumb Tuff Member of the Klondike Mountain Formation in the village of Republic, Washington, United States of America; mid-Ypresian, 51.18 ± 0.09 Ma (Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021).
Palaeopsychops barthae sp. nov
Zoobank Registration number: urn:lsid:zoobank.org:act:6141865C-182C-4AC3-816D-3CFBDE0205DA Figure 2
Palaeopsychops barthae sp. nov. holotype hind wing SR 15-001-001 (part only known). A, photograph, wetted with ethanol; B, drawing, n = expected position of nygma. Black spots in parts of the wing are discolouration of the rock. Both to scale = 5 mm.

Type and other material. Holotype hind wing SR 15-001-001, found at exposure B4131 of the Klondike Mountain Formation in Republic, Washington by Kathrynn Barth, 8 April 2015, housed in the collection of the Stonerose Interpretive Center.
Diagnosis. Distinguished from all Palaeopsychops species except P. goodwini by the broad pale fascia, distinguished from the latter by the contrasting characters in its diagnosis; see above description.
Description. HOLOTYPE. Hind wing 28 mm long, 13 mm wide. Costal space broad as in P. goodwini, strongly broadened distad fusion of Sc, RA. Humeral veinlet not preserved. Subcostal veinlets poorly preserved, closely spaced; some forked once. Subcostal space moderately broad; no crossveins detected. Sc, RA fused distally; Sc+RA probably terminating at margin slightly before wing apex; its veinlets mostly dichotomously forked (2–3 times). RA space broadest basally, then nearly as broad as subcostal space; one preserved crossvein in basal part. RP with approximately 25 branches or slightly more; none apparently forked before outer gradate series, forked 1–3 times distad outer series. Two gradate series of crossveins in radial space (inner, outer), few crossveins preserved in both. Distal nygma probably located proximad inner gradate series of crossveins judging from curvature of RP2. Basal oblique crossvein between RP1, MA. M forked at level of origin of RP1. MA, MP poorly preserved distally probably with few branches. Cu divided to CuA, CuP apparently near wing base. CuA pectinate, with four long preserved branches. CuP dichotomously branched. Four crossveins detected between CuA, CuP. Two crossveins detected between CuP, A1. A1 pectinately branched, with six long branches, each forked 1–3 times. A2 pectinately branched, with at least three branches, proximal two forked once; distal branch forked three times. A3 not preserved. Trichosors distinct along entire wing margin. Colouration dark throughout, except: pale, rather broad fascia mid-wing, perpendicular to RA; possible large pale spot in apical region (maybe an artefact?), presumed smaller spots in posterior portion of wing.
Etymology. The specific epithet is formed from the surname of Kathrynn Barth, who found the holotype and donated it to the Stonerose Interpretive Center.
Range and age. Exposure B4131 of the Tom Thumb Tuff Member of the Klondike Mountain Formation, in the village of Republic, Washington, United States of America, U–Pb maximum likelihood age as above (Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021).
Palaeopsychops sp. indet
Figure 3
Venation of Palaeopsychops sp. indet. forewing RBCM.EH2017.050.3020/3043 (part and counterpart). A, photograph of RBCM.EH2017.050.3043 (counterpart), dry; B, drawing from part and counterpart, n = expected position of nygma. Both to scale = 5 mm.
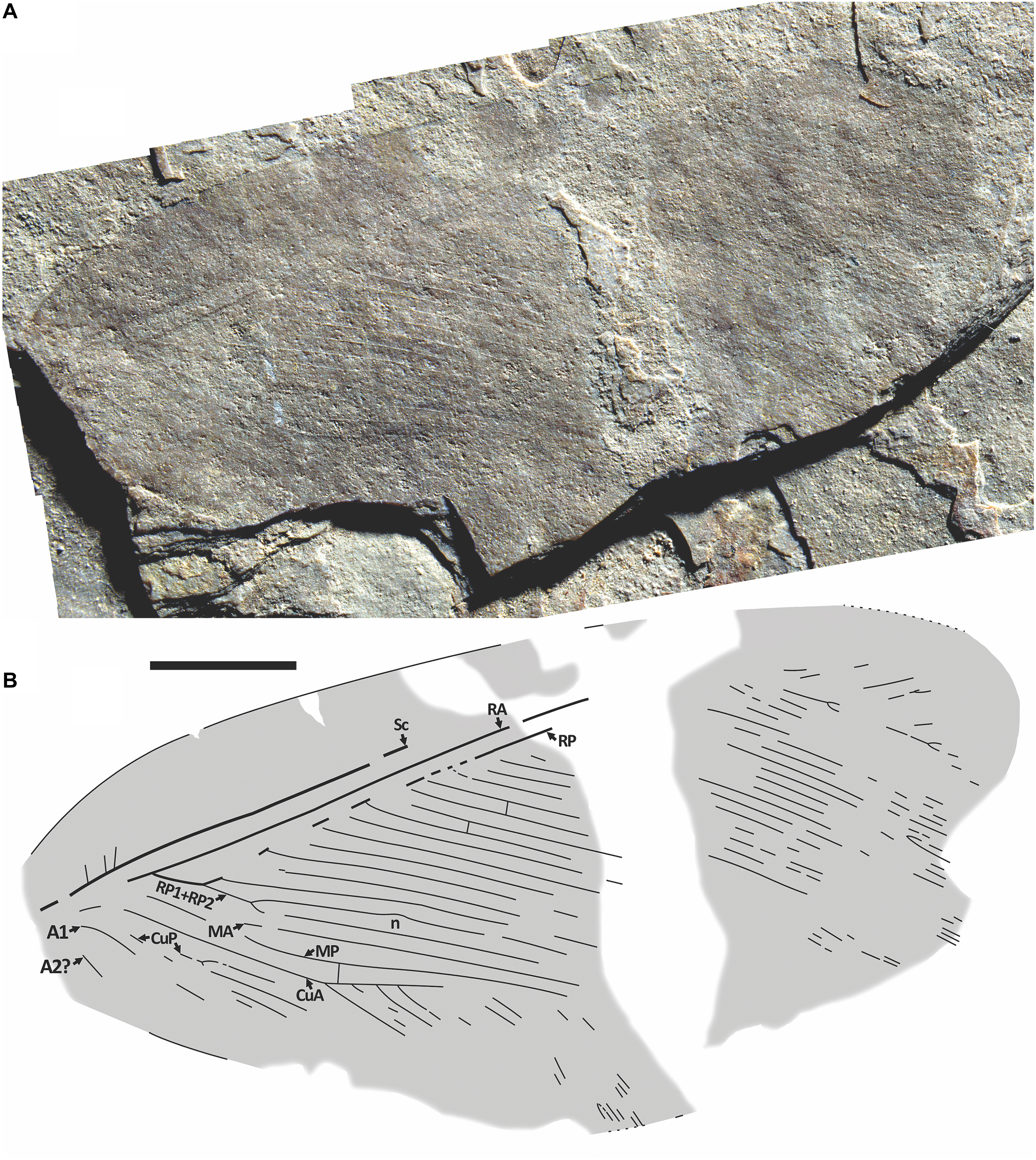
Material. Specimen RBCM.EH2017.050.3020/3043, a rather complete forewing with much of its venation indistinctly preserved, found by John Leahy, date unknown, at the McAbee location near the village of Cache Creek, British Columbia, Canada. A U–Pb maximum likelihood age from zircons for McAbee is 52.10 ± 0.26 Ma (Rubino et al. Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021).
Description. Forewing 34 mm as preserved (estimate complete length ∼35 mm), 16.9 mm wide. Costal space broadest at ca. 1/5 wing length, smoothly narrowed basad, slightly narrowed towards apex, then strongly broadened distad possible fusion of Sc, RA, not preserved, within fascia. Presumed humeral veinlet, subcostal veinlets very poorly preserved, practically indiscernible. Subcostal space moderately broad; no crossveins detected. No distal parts of Sc, RA, and possible Sc+RA discernible by preservation. RA space relatively narrow, more so than subcostal space; no crossveins detected. RP with approximately 24–25 branches, at least some forked distally (details not preserved, indicated by narrower spacing); RP1, RP2 fused basally, not far from origins. Only one crossvein preserved in radial space. Distal nygma apparently located proximad presumed inner gradate series of crossveins as indicated by characteristic curvature of RP2. M forked distad origin of RP1 + RP2. MA, MP not preserved distally. One crossvein between MP, CuA rather well preserved, possibly belonging to inner gradate series. Cu divided to CuA, CuP probably near wing base. CuA pectinate, with four long preserved branches. CuP probably dichotomously branched. Basal parts of A1, A2 preserved. A3 not preserved. Colouration dark throughout, except pale, relatively narrow fascia about mid-wing, nearly perpendicular to RA.
Remarks. The colouration of forewing RBCM.EH2017.050.3020/3043 including the placement and orientation of its pale fasciae is similar to that of the P. barthae holotype hind wing, suggesting the possibility that they might be conspecific. The 0.80 ratio of forewing (35 mm) to hind wing (28 mm) lengths is a little less than the 0.88–0.90 ratio of extant Polystoechotidae specimens (V.N.M., unpublished data), although this might be explained by the McAbee specimen being slightly smaller than the Republic holotype due to intraspecific variation or sexual dimorphism. Alternatively, they might be regional variations of the same species, closely related species, or similar by convergence. New specimens may clarify this.
In the early Eocene, Republic and McAbee had similar forests and upper microthermal climates with mild winters (Greenwood et al. Reference Greenwood, Archibald, Mathewes and Moss2005; Moss et al. Reference Moss, Greenwood and Archibald2005; Gushulak et al. Reference Gushulak, West and Greenwood2016). An increasing number of insect species are known to be shared between them, for example, Okanagrion hobani Archibald and Cannings, 2021 (Odonata, Dysagrionidae; Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021); an unnamed cercopoid (Hemiptera, Auchenorrhyncha; Archibald et al. Reference Archibald, Greenwood and Mathewes2013a); and Eorpa ypsipeda Archibald et al., 2013 (Mecoptera, Eorpidae; Archibald et al. Reference Archibald, Mathewes and Greenwood2013b). March flies (Diptera, Bibionidae, Plecia) are the most common insects at both sites (and throughout the Okanagan Highlands), although their Okanagan Highlands species are in need of revision.
Discussion
We assign these two new species to the early Eocene genus Palaeopsychops because their hind wing venation is very similar to that of all described species of the genus: P. maculatus Andersen, 2001 and one of the paratypes of P. abruptus Andersen, 2001, both from the Fur Formation of Denmark; P. setosus Archibald and Makarkin, 2006 from Horsefly, British Columbia, Canada; and P. douglasae Archibald and Makarkin, 2006 from Quilchena, British Columbia, Canada.
The colour patterning of the new species and the McAbee forewing described here is unique in Palaeopsychops. Species from the Danish Fur Formation tend to have narrow dark fascia on medium-dark-to-somewhat-lighter wings, whereas those of the Okanagan Highlands usually bear bold patterning, dark throughout or with different pale areas, broad fascia or spots; see, for example, P. douglasae, P. setosus, P. timmi Archibald and Makarkin, 2006, and P. marringerae Archibald and Makarkin, 2006.
The new species are further distinct by their hind wings, which are much shorter (27–28 mm long) than those of the previously known species (∼40–42 mm; estimated for P. abruptus).
The P. goodwin and P. barthae specimens provide new information about the hind wing venation of Palaeopsychops. Their basal parts are well preserved, and the configuration of anal veins in Palaeopsychops, which was little known in previous fossil hind wings of the genus, is now clearly discernible: A1 is multi-branched (either dichotomous or pectinate); A2 is pectinate; and A3 is probably pectinate. The presence of a distal nygma is indicated in both new species, most distinctly in the hind wing of P. goodwini, but has not been detected in the hind wing of any previously known Palaeopsychops species, presumably due to preservation. The distal nygma is present in the hind wing of the Chilean Fortecillia Navás, 1932 but is absent in those of all other extant Polystoechotidae. The only other fossil polystoechotid species with a distinct hind wing distal nygma is the Late Jurassic Osmyloides distinctus Panfilov, 1980 from Karatau (Kazakhstan; V.N.M., unpublished data).
The genus Palaeopsychops was described in Psychopsidae (Andersen Reference Andersen2001), but we later argued that it belongs to Polystoechotidae (Archibald and Makarkin Reference Archibald and Makarkin2006). The status of this family is, however, currently unresolved. Winterton and Makarkin (Reference Winterton and Makarkin2010) synonymised it with Ithonidae, the fossils of which, sensu lato, are numerous and diverse, especially in the Jurassic and Cretaceous Periods. Zheng et al. (Reference Zheng, Ren and Wang2016) divided the Ithonidae s.l. into the ithonid genus-group, polystoechotid genus-group, and rapismatid genus-group, in effect reviving the previous families Ithonidae s. str., Polystoechotidae and Rapismatidae. Xu et al. (Reference Xu, Zheng, Fang, Shih, Ren and Wang2021) renamed these as the Ithone genus-group, Polystoechotes genus-group, and Rapisma genus group. Makarkin and Archibald (Reference Makarkin and Archibald2009) had earlier established the Principiala genus-group in Ithonidae s. str. These four genus groups of Ithonidae s.l. are currently widely accepted (e.g., Lu et al. Reference Lu, Zhang, Ohl and Liu2017; Gao et al. Reference Gao, Xu, Shih, Ren and Wang2024). Additionally, the Jurassic family Epigambriidae is currently considered to constitute the subfamily Epigambrinae in Ithonidae s.l., most closely related to the polystoechotid genus-group (Makarkin et al. Reference Makarkin, Ansorge and Khramov2021). The classification of Ithonoidea then remains unresolved, and we treat the Polystoechotes genus-group as a family as we have earlier (e.g., Makarkin and Archibald Reference Makarkin and Archibald2003; Archibald and Makarkin Reference Archibald and Makarkin2006) until clear evidence for a different status appears.
Acknowledgements
We thank John Leahy for donating the McAbee specimen to the Royal British Columbia Museum and museum Curator of Fossils Victoria Arbour and Collections Manager Derek Larson for making it available to us. Likewise, we thank Kathrynn Barth, finder of SR 15-001-001 and Steven Goodwin, finder of SR 13-002-001, for donating these to the Stonerose Interpretive Center and centre President Mary Rourke and Managing Director Travis Wellman for making them available to us. We also thank Marlow Pellatt of Parks Canada for use of the microscope and camera in his laboratory. S.B.A. thanks Rolf Mathewes, Simon Fraser University, Burnaby, British Columbia, for support from his Natural Sciences and Engineering Research Council Discovery Grant 2020-05026. The research of V.N.M. was carried out within the framework of the state assignment of Ministry of Science and Higher Education of the Russian Federation (theme No. 124012400285-7).
Competing interests
The authors declare that they have no competing interests.



 Open access
Open access