


Introduction
After double fertilization, coenocytic proliferation and subsequent cellularization of the triploid endosperm are necessary for normal growth and development of the embryo through to maturity (Yadegari and Drews, Reference Yadegari and Drews2004; Lafon-Placette and Köhler 2014). Epigenetic repression by polycomb repressive complex 2 (PRC2) is indispensable for heterochromatin formation in the central cell of the female gametophyte and for normal gene expression from the maternally derived genome in the endosperm during seed development (Grossniklaus and Paro, Reference Grossniklaus and Paro2014). In Arabidopsis thaliana, the FERTILIZATION-INDEPENDENT SEED–PRC2 (FIS–PRC2) variant, composed of MEDEA, FERTILIZATION-INDEPENDENT SEED 2, FERTILIZATION-INDEPENDENT ENDOSPERM and MULTICOPY SUPPRESSOR OF IRA1, catalyses H3K27 trimethylation at key loci to drive endosperm cellularization and embryo viability, and maternal loss of any of these components induces seed abortion (Grossniklaus et al., Reference Grossniklaus, Vielle-Calzada, Hoeppner and Gagliano1998; Luo et al., Reference Luo, Bilodeau, Dennis, Dennis, Peacock and Chaudhury1999; Ohad et al., Reference Ohad, Margossian, Hsu, Hannon, Michaeli, Harada, Goldberg and Fischer1999; Köhler et al., Reference Köhler, Henning, Bouveret, Gheyselinck, Grossniklaus and Gruissem2003).
Beyond serving as a nutritive tissue, the endosperm exerts a pivotal regulatory influence on seed dormancy. The endosperm synthesizes hormones, including abscisic acid (ABA) and gibberellins, to either enforce or alleviate dormancy in response to environmental stimuli (Lee et al., Reference Lee, Piskurewicz, Turečková, Carat, Chappuis, Strnad, Fankhauser and Lopez-Molina2012; Piskurewicz et al., Reference Piskurewicz, Sentandreu, Iwasaki, Glauser and Lopez-Molina2023). This is influenced by the epigenetic state of the endosperm, with disruption of repressive chromatin leading to production of non-dormant seeds under conditions that normally cause high dormancy levels in Col-0 (Iwasaki et al., Reference Iwasaki, Hyvärinen, Piskurewicz and Lopez-Molina2019; Sato et al., Reference Sato, Santos-González and Köhler2021; Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023).
PRC2 complexes interact with accessory factors that are necessary for stable chromatin association (Holoch and Margueron, Reference Holoch and Margueron2017). The VERNALIZATION INSENSITIVE 3-LIKE (VEL) family of PRC2 accessory proteins includes VIN3, VRN5, VEL1 and VEL2, which harbour plant homeodomain (PHD) and fibronectin type III domains that mediate their interaction with PRC2 and chromatin (De Lucia et al., Reference De Lucia, Crevillén, Jones, Greb and Dean2008; Gao and He, Reference Gao and He2024). VEL1, VIN3 and VRN5 recruit PRC2 to FLOWERING LOCUS C (FLC) during vernalization, via polymerization and phase separation driven by the VEL domain (Schulten et al., Reference Schulten, Jang, Payne-Dwyer, Fiedler, Nielsen, Mateo-Bonmatí, Bienz, Leake and Dean2025). VEL2 and VEL3 are both expressed in gametophytes and seeds, and VEL3 encodes an atypical truncated VEL protein that lacks the PHD present in other VEL proteins. We recently showed that VEL3 is required for H3K27me3 deposition in the endosperm and regulates dormancy via association with histone deacetylase (HDAC) complexes (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). It remains unclear if this dormancy regulation also requires FIS–PRC2 function. VEL1 and VRN5 also interact with HDACs (De Lucia et al., Reference De Lucia, Crevillén, Jones, Greb and Dean2008) and are necessary for H3K27me3 deposition at FLC during vernalization, but it is unclear whether they also play a role in H3K27me3 deposition in heterochromatin in the maternally derived genome of the endosperm.
Here, we use CRISPR-Cas9 gene editing of the VEL family to overcome any redundancy in function and assess their contribution to seed development and primary seed dormancy. Our results show that VEL2 does not contribute to seed dormancy or abortion. Only the loss of VEL3 affects seed dormancy, while VEL1 and VRN5 are necessary alongside VEL3 to prevent seed abortion that may be associated with hyperactivation of the maternally derived genome in the endosperm. These findings demonstrate that the two effects are genetically separable.
Materials and methods
Plant materials and growth conditions
Arabidopsis thaliana mutants used in this study have been previously described: vel3-1 (SALK_052041; Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023), vel3-1 gVEL3-GFP (VEL3-GFP) (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023), vil2-1 (vel1-1) (SAIL_323_A03; Kim and Sung, Reference Kim and Sung2010) and vrn5-8 (SALK_136506; Greb et al., Reference Greb, Mylne, Crevillén, Geraldo, An, Gendall and Dean2007). Three transgenic lines expressing 35S::GFP-VEL2 were generated via Agrobacterium tumefaciens-mediated floral dipping. The VEL2 coding sequence was PCR-amplified (forward: ttGGTCTCtTTCGATGGCTTCTTTTCATAAAGGAGCTGC; reverse: ttGGTCTCtAAGCTCAATCCTGCAGCAGAATCAAAGACAC) and subsequently assembled with eGFP using Golden Gate cloning (see Supplementary Table 1 for details of plasmids). The resulting constructs were introduced into Col-0 plants. Other mutant lines used here were generated by CRISPR-Cas9: vel2-1, vel2-2, vel2-3, vel2-4 vel3-1, vel1-2 vrn5-9, vel2-5 vin3-4 vrn5-10 (velT) and vel1-3 vel2-5 vin3-5 vrn5-10 (velQ). The vel2-4 vel3-1 double mutant was created by CRISPR-Cas9 knock out of VEL2 in the vel3-1 background. Arabidopsis plants were directly grown in long days (16 h light and 8 h darkness at 22°C) until bolting. In the vernalization experiment, 2-week-old seedlings were subjected to a 4-week period at 4°C, followed by a transfer to 22°C until bolting occurred. After bolting, plants were either maintained at 22°C or transferred to 15°C for seed set. All mutants and wild type (WT) were grown side-by-side for each experiment.
Seed germination
For dormancy assays, freshly matured seeds were sown onto 0.9% water agar plates and placed in long days (16 h light and 8 h darkness) at 22°C. Stratification was performed at 4°C in the dark for 3 days as indicated. For ABA sensitivity, seeds were sown onto 0.9% water agar plates with or without exogenous ABA (1, 5 or 10 µM), stratified and then placed in long days (16 h light and 8 h darkness) at 22°C. Seed batches from individual mother plants were used as biological replicates; at least 6 biological replicates of ∼50 seeds were used for each experiment. Seed germination was scored by radicle emergence at 7 days after sowing.
Microscopy
Arabidopsis thaliana 35S::VEL2 seeds carrying GFP reporter constructs were germinated and seedlings were grown under long-day conditions (16 h light/8 h dark, 22°C). Hypocotyls from 5- to 7-day-old seedlings were mounted in phosphate-buffered saline (pH 7.4) on glass slides. Fluorescence imaging was performed with a Leica SP8X confocal microscope using a white light laser, with GFP excited at 488 nm and emission collected between 500 and 550 nm. All samples were imaged under identical acquisition settings to ensure comparability across biological replicates. Images were processed in Leica LAS X software, applying only linear adjustments to brightness and contrast without altering raw fluorescence signals. Representative images were selected from at least three independent experiments.
VEL gene family editing by CRISPR-Cas9
CRISPR-Cas9 gene editing was performed as described in Lawrenson et al. (Reference Lawrenson, Shorinola, Stacey, Li, Østergaard, Patron, Uauy and Harwood2015). In short, target sequences for VEL1, VEL2, VIN3 and VRN5 were selected for guide RNAs (gRNAs) using a web-based bioinformatics tool (CHOPCHOP – https://chopchop.cbu.uib.no/), with four guides designed for each gene. Separately, we performed CRISPR-Cas9 targeting the beginning of the first exon of the VEL2 gene (see Supplementary Table 1 for details of gRNAs). Plasmids were constructed (see Supplementary Table 1 for details of plasmids and gRNAs) using the Golden Gate system following a previously described assembly strategy (Lawrenson et al. Reference Lawrenson, Shorinola, Stacey, Li, Østergaard, Patron, Uauy and Harwood2015). We used the Discosoma Red (DsRED) fluorescent protein marker to screen for putative transgene-free CRISPR-Cas9 edited plants of Arabidopsis (Aliaga-Franco et al., Reference Aliaga-Franco, Zhang, Presa, Srivastava, Granell, Alabadí, Sadanandom, Blázquez and Minguet2019). Plants were transformed using Agrobacterium-mediated floral dip with Agrobacterium resuspended in 5% sucrose and 0.02% (v/v) Silwet L-77. Genotyping of edited plants was carried out by PCR using the following primers (VEL1-forward: AGGTCTTGCAGGCATGGAGC; VEL1-reverse: AGAGGACCCACATCAGCTTCAAG; VEL2.vel2-1-forward: TGATCTGAAATGAAGTTTGG; VEL2.vel2-1 reverse: TAGCTTTAGCTAATGGAGTC; VEL2.others forward: GTTCTGCGGTTGTAGTTGAGGT; VEL2.others-reverse: GCAGGTTAACCAGAGACTAGGATCT; VIN3-forward: GCCTGAAGAAGCTTCGGAGC; VIN3-reverse: GAGACCACTTTGTCCAGAGCTTCC; VRN5-forward: CCTCCGTAATCTCAATCACCGAAC; VRN5-reverse: GGCTGACAACTCCCCTGCC). A summary of the construct design, gRNA positioning and resulting alleles is given in Fig. 1.
Gene editing strategy for targeted chromosomal indels in VEL family genes in Arabidopsis. (A) Schematic representation of the final CRISPR construct. In brief, DNA oligonucleotides encoding the sequences of 16 single-guide RNAs (sgRNAs) were used to generate double-stranded DNA fragments, which were subsequently integrated into the final construct with an intron-optimized Cas9 and DsRED tagged selection via Golden Gate assembly, mediated by Type IIS restriction enzymes BsaI/BpiI. This enzymatic system facilitates the efficient assembly of large and complex constructs from standardized constitutive parts in a minimal number of reactions. Arabidopsis plants were transformed via floral dipping method with Agrobacterium tumefaciens carrying the final construct. Fluorescent DsRED-positive seeds were germinated, and plants were screened for somatic indels. Selected plants with indels produced DsRED-negative T1 seeds, which were further selected. T2 plants were grown, and homozygous mutants were screened through T4. (B) Sequence diagram of the VEL proteins (VEL1, VEL2, VIN3 and VRN5). Main domains of the VEL proteins (plant homeodomain, PHD; fibronectin type III, FIBIII and VERNALIZATION-INSENSITIVE3-like, VEL) are shown in orange boxes. Asterisks indicate the location of the first premature stop codon resulting from the frameshift mutation of each allele. VelT refers to vel2-5 vin3-4 vrn5-10 triple mutant. VelQ refers to vel1-3 vel2-5 vin3-5 vrn5-10 quadruple mutant.

Figure 1 Long description
The first diagram illustrates the CRISPR construct used for gene editing in Arabidopsis plants. It includes components such as rbcS-E9t, Cas9 with introns, AtUbi10p, U626c and multiple sgRNAs targeting VRN5, VIN3, VEL1 and VEL2 genes. The process involves Cas9 transformation, DsRED positive seed selection, screening for somatic indels and selection of homozygous mutants through T4 generation. Arabidopsis plants are shown at different stages: T subscript 0, T subscript 1, T subscript 2 and T subscript 3/T subscript 4. The second diagram presents sequence diagrams of VEL proteins (VEL1, VEL2, VIN3, VRN5) with domains labeled as Plant Homeodomain (PHD), Fibronectin type III (FIBIII) and VERNALIZATION-INSENSITIVE3-like (VEL). Asterisks indicate the location of premature stop codons resulting from frameshift mutations in each allele, with labels for wild type and mutant alleles such as vel superscript 1-2 vel superscript 2 vel superscript 3-5 vel superscript 1-3 vel superscript 2-5 vin superscript 3-5 vrn superscript 5-10.
RNA-seq and qPCR
RNA-seq was performed as described in Chen et al. (Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). In short, RNA was extracted from mature endosperm by first imbibing 0.05 g of seeds for 1 h. Following Iwasaki and Lopez-Molina (Reference Iwasaki and Lopez-Molina2021), endosperm-enriched tissues were then isolated using a glass slide to squeeze the seeds and centrifuged twice at maximum speed (16,000 rpm) with 40% sucrose. After washing three times with water, the samples were stored at −80°C. RNA extraction was performed using the borate method (Penfield et al., Reference Penfield, Josse, Kannangara, Gilday, Halliday and Graham2005). The RNA pellet was dissolved in water and further purified using a Qiagen RNeasy Plant Mini Kit, including on-column DNase I digestion. RNA sequencing was performed by Novogene using a HiSeq 4000 system, generating 150 bp paired-end sequences with a minimum of 25 million reads per sample. Three biological replicates were prepared. Raw RNA-seq reads were trimmed using Cutadapt (Martin, Reference Martin2011) and mapped to the A. thaliana TAIR10 genome with STAR (Dobin et al., Reference Dobin, Davis, Schlesinger, Drenkow, Zaleski, Jha, Batut, Chaisson and Gingeras2013). Read counts for each gene were obtained using featureCounts (Liao et al., Reference Liao, Smyth and Shi2014), and only sense reads were analysed further. Differentially expressed genes (DEGs) were identified with edgeR (Robinson et al., Reference Robinson, McCarthy and Smyth2010), using FDR (False Discovery Rate) < 0.05 and a fold-change ≥ 2. For qPCR, three seedlings were pooled as one biological replicate, and RNA was extracted using the RNeasy Plant Mini Kit (Qiagen, 74904). cDNA samples were prepared using the Quantitect reverse transcription kit (Qiagen, 205311). qPCR was then performed on the VEL2 gene (forward: TCCAAGACGAAGAAGTGGCAGC; reverse: CACTCTTGACCTGACCTGAGCAC) with the SYBR Green master mix (Bio-Rad, 1725271). ACT11 (forward: GGAACAGTGTGACTCACACCATC; reverse: AAGCTGTTCTTTCCCTCTACGC) and PP2A (forward: TAACGTGGCCAAAATGATGC; reverse: GTTCTCCACAACCGCTTGGT) were used as the reference genes.
Results
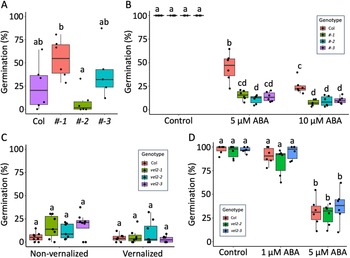
Previously, we established a role for VEL3 in seed dormancy. In this paper, this function is being re-examined while also investigating the roles of other members of the VEL family. Although VEL2 is highly expressed in seeds (Klepikova et al., Reference Klepikova, Kasianov, Gerasimov, Logacheva and Penin2016 ), the role of VEL2 remained uncharacterized because we could not isolate vel2 T-DNA mutants for analysis (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). To elucidate VEL2 function, we first generated Col-0 lines expressing GFP-tagged VEL2 under the cauliflower mosaic virus 35S promoter (Supplementary Figure 1). To induce seed dormancy, seeds from the respective lines were matured at 16°C, and germination was evaluated in the absence of stratification. Notably, the three transgenic 35S:GFP-VEL2 lines did not exhibit statistically significant differences in dormancy levels relative to the Col-0 control (Fig. 2A). Subsequently, seeds were matured at 22°C to generate non-dormant populations for ABA sensitivity assays. Under these conditions, the 35S:GFP-VEL2 lines displayed slightly increased sensitivity to ABA compared to Col-0 (Fig. 2B) but were otherwise phenotypically normal.
VEL2 does not play a role in seed dormancy. (A) Germination of Col-0, 35S::GFP-VEL2-1 (#-1), 35S::GFP-VEL2-2 (#-2) and 35S::GFP-VEL2-3 (#-3) seeds set at 16°C. (B) Germination of Col-0, OE-1, OE-2 and OE-3 seeds set at 22°C in the presence of exogenous ABA (5 or 10 µM). (C) Germination of Col, vel2-1, vel2-2 and vel2-3 seeds set at 15°C. Non-vernalized seedlings were grown continuously at 22°C until bolting, while vernalized seedlings had a 4°C treatment for 4 weeks before going back to 22°C until bolting. (D) Germination of Col-0, vel2-2 and vel2-3 seeds to 22°C in the presence of exogenous ABA (1 or 5 µM). Significant differences are shown using ANOVA with Tukey’s post hoc test (P < 0.05; n = 6–8).

Figure 2 Long description
The image contains four box plots labeled A, B, C and D. A shows germination percentages for genotypes Col, number 1, number 2 and number 3. B displays germination under control, 5 micromolar ABA and 10 micromolar ABA conditions for genotypes Col, number 1, number 2 and number 3. C illustrates non-vernalized and vernalized conditions for genotypes Col, vel2-1, vel2-2 and vel2-3. D presents control, 1 micromolar ABA and 5 micromolar ABA conditions for genotypes Col, vel2-1, vel2-2 and vel2-3. Each plot includes data points and statistical annotations such as a, b, c and d indicating significant differences.
To obtain various knockout mutants of VEL2, we used a CRISPR-Cas9 approach. We designed six RNA guides to target different portions of the VEL2 gene and obtained three independent vel2 alleles with frameshift mutations predicted to cause premature termination of translation and the production of truncated VEL2 proteins (Fig. 1). To induce dormancy, seeds from these vel2 single mutant lines were matured at 15°C, and germination was assessed in freshly harvested seeds. Dormancy of vel2 mutants remained indistinguishable from WT (Fig. 2C). Because VEL2 expression is induced by vernalization (Kim and Sung, Reference Kim and Sung2013), we vernalized Col-0 and vel2 seedlings for 4 weeks at 4°C to test whether maternal vernalization had any impact on subsequent seed dormancy in vel2 mutants. Again, no significant differences were found between genotypes (Fig. 2C). Next, we set seeds at 22°C to produce non-dormant seed lots and assess ABA sensitivity. Neither 1 µM nor 5 µM of exogenous ABA revealed any differences in seed germination between WT and vel2 mutants (Fig. 2D). Together, these results indicate that VEL2 either does not function in processes affecting seed dormancy or that VEL2 functions redundantly with other VEL family members in the endosperm.
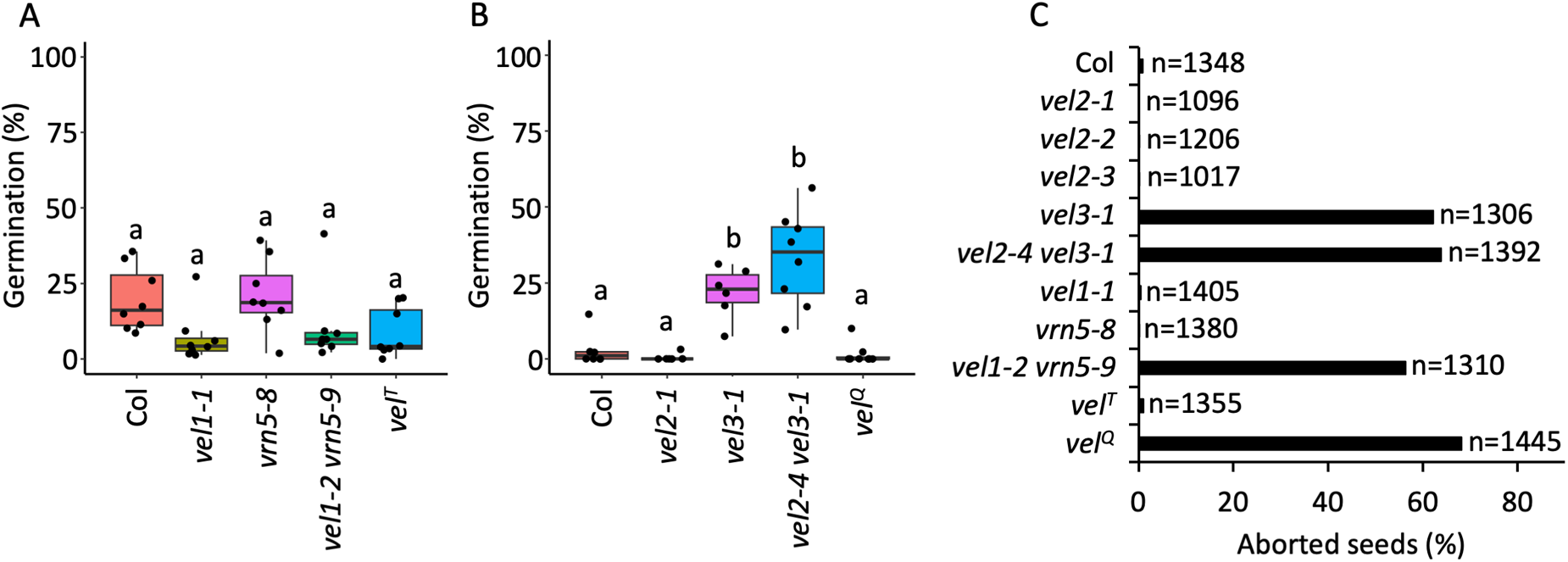
To test whether other VEL family members contribute redundantly to endosperm development or seed dormancy, we next targeted VEL1, VEL2, VIN3 and VRN5 together using CRISPR-Cas9 with the aim of generating lines with loss-of-function mutants for different combinations of VEL genes. For each gene, we designed four gRNAs, assembled into a single construct and obtained double, triple and quadruple mutants (Fig. 1). We matured seeds at 15°C from the following previously characterized mutants: vel1-1, vel2-1, vel3-1 and vrn5-8, then from newly isolated vel1-2 vrn5-9 and vel2-4 vel3-1 double mutant lines; vel2-5 vin3-4 vrn5-10 triple mutant line and vel1-3 vel2-5 vin3-5 vrn5-10 quadruple mutant line. Strikingly, only seeds with the vel3-1 background displayed an altered dormancy phenotype (Fig. 3A, B), suggesting that if these mutations cause loss of gene function as predicted, then VEL3 alone has a role in seed dormancy.
While VEL1, VEL2, VIN3 and VRN5 do not affect seed dormancy, VEL1 and VRN5 are essential to prevent seed abortion. (A) Germination of Col, vel1-1, vrn5-8, vel1-2 vrn5-9 double mutant and vel2-5 vin3-4 vrn5-10 triple (velT) mutant seeds set at 15°C. (B) Germination of Col, vel2-1, vel3-1, vel2-4 vel3-1 double mutant and vel1-3 vel2-5 vin3-5 vrn5-10 quadruple (velQ) mutant seeds set at 15°C. Significant differences are shown using ANOVA with Tukey’s post hoc test (P < 0.05; n = 6–8). (C) Seed abortion frequency (in percentage) in Col, vel2-1, vel2-2, vel2-3, vel3-1, vel2-4 vel3-1, vel1-1, vrn5-8, vel1-2 vrn5-9, vel2-5 vin3-4 vrn5-10 (velT) and vel1-3 vel2-5 vin3-5 vrn5-10 (velQ). The total number of seeds analysed for each genotype is shown.

Figure 3 Long description
The image consists of three graphs labeled A, B and C. Graph A shows germination percentage on the y-axis for Col, vel1-1, vrn5-8, vel1-2 vrn5-9 and vel superscript T genotypes. Col has the highest germination, while vel superscript T has the lowest. Graph B shows germination percentage for Col, vel2-1, vel2-4 vel3-1 and vel3-1 genotypes. Vel3-1 has the highest germination, while Col has the lowest. Graph C shows the percentage of aborted seeds on the x-axis for various genotypes, with Col having the lowest and vel superscript Q having the highest percentage. Each graph includes data points and error bars.
We previously found that vel3-1 mutants exhibit high-frequency seed abortion accompanied by ectopic gene expression in endosperm closely resembling that of maternal PRC2 mutants and activation of the so-called ‘triploid block’ (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). After testing single and multiple mutant lines, we found that vel1-2 vrn5-9 double mutant and vel1-3 vel2-5 vin3-5 vrn5-10 quadruple mutant also showed high frequency seed abortion (Fig. 3C). The normal seed development of vel1 and vrn5 single mutants strongly suggested that VEL1 and VRN5 function redundantly alongside VEL3 to facilitate PRC2 activity in the central cell and/or endosperm. Of interest here is that seed abortion increases without compromising the establishment of dormancy in the vel1-2 vrn5-9 double and vel1-3 vel2-5 vin3-5 vrn5-10 quadruple mutants, whereas the vel3-1 mutant disrupts both phenotypes, suggesting that VEL3 function in seed dormancy is independent of triploid block activation and possibly PRC2 activity.
We then aimed to isolate gene expression changes in vel3-1 specifically associated with alteration of seed dormancy by comparing the vel3-1 endosperm transcriptome with that of vel1 vrn5, performing RNA sequencing on mature endosperm from vel1-2 vrn5-9 double mutants and comparing the resulting transcriptome to our previously published vel3-1 dataset (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). Comparison between vel1-2 vrn5-9 and vel3-1 revealed 66 shared DEGs presumably associated with triploid block activation or another PRC2 function, including NAC046 (NAC domain-containing protein 46), WRKY75 (WRKY DNA‐binding protein 75) and ORESARA1 (ORE1) (Fig. 4A, B; Table S1). These genes were upregulated in both genotypes and have established roles in plant senescence (Balazadeh et al., Reference Balazadeh, Riaño-Pachón and Mueller-Roeber2010; Guo et al., Reference Guo, Li, Huang, Li, Fang, Chu and Guo2017; Huysmans et al., Reference Huysmans, Lemaire and De Veylder2018), with NAC046 and ORE1 specifically implicated in endosperm elimination via programmed cell death (Doll et al., Reference Doll, Van Hautegem, Schilling, De Rycke, De Winter, Fendrych and Nowack2023).
VEL1 and VRN5 are involved in the repression of senescence and programmed cell death-associated gene expression in the endosperm. (A) Comparison of differentially expressed genes (DEGs) in vel3-1 and vel1-2 vrn5-9 in the mature endosperm, genes in bold (NAC046 and ORE1) regulate the pace of endosperm elimination by programmed cell death, after Doll et al. (Reference Doll, Van Hautegem, Schilling, De Rycke, De Winter, Fendrych and Nowack2023). Comparison of up- (B) and downregulated (C) genes in vel3-1 and vel1-2 vrn5-9 in the mature endosperm. (D) GO-term analysis of up- (left, in blue) and downregulated (right, in green) genes between vel3-1 and vel1-2 vrn5-9 mature endosperms, showing gene function category, p-values and number of genes in each category.

Figure 4 Long description
The image consists of four parts. A shows a Venn diagram of differentially expressed genes (DEGs) with 199 unique to vel3-1, 66 shared and 396 unique to vel1-2 vrn5-9. Highlighted genes include WRKY75, NAC046 and ORE1. B shows up-regulated genes with 115 unique to vel3-1, 58 shared and 385 unique to vel1-2 vrn5-9. C shows down-regulated genes with 87 unique to vel3-1, 5 shared and 14 unique to vel1-2 vrn5-9. D presents a bar chart of gene functions, including responses to salt stress, hypoxia, heat, hydrogen peroxide, protein folding and more, with gene ratios and p-values indicated for each function category.
We next analysed DEGs unique to vel3-1, splitting them into upregulated and downregulated subsets. We identified 115 genes upregulated in vel3-1 but not vel1-2 vrn5-9 (Fig. 4B; Table S1), and Gene Ontology (GO) analysis highlighted stress‐related categories such as salt stress, hypoxia, heat and hydrogen peroxide response (Fig. 4D). In our experience, these gene functions tend to be enriched in seed transcriptomes in any experimental situation, and we did not identify any strong candidate genes for vel3-specific dormancy regulators. Interestingly, 95% of DEGs in vel1-2 vrn5-9 were upregulated relative to Col-0, whereas DEGs in vel3-1 displayed an approximately equal number between induction and repression (Fig. 4B, C; Supplementary Table 1). When comparing downregulated genes between vel1-2 vrn5-9 and vel3-1, we found 87 genes specifically repressed in vel3-1 (Fig. 4C; Table S1). GO analysis revealed genes functioning at multiple levels of gene expression, including chromatin accessibility (chromatin remodelling), transcription initiation (DNA‐templated transcription), RNA maturation (splicing) and organelle protein targeting (chloroplast import) (Fig. 4D), but no clear dormancy regulators were identified.
Discussion
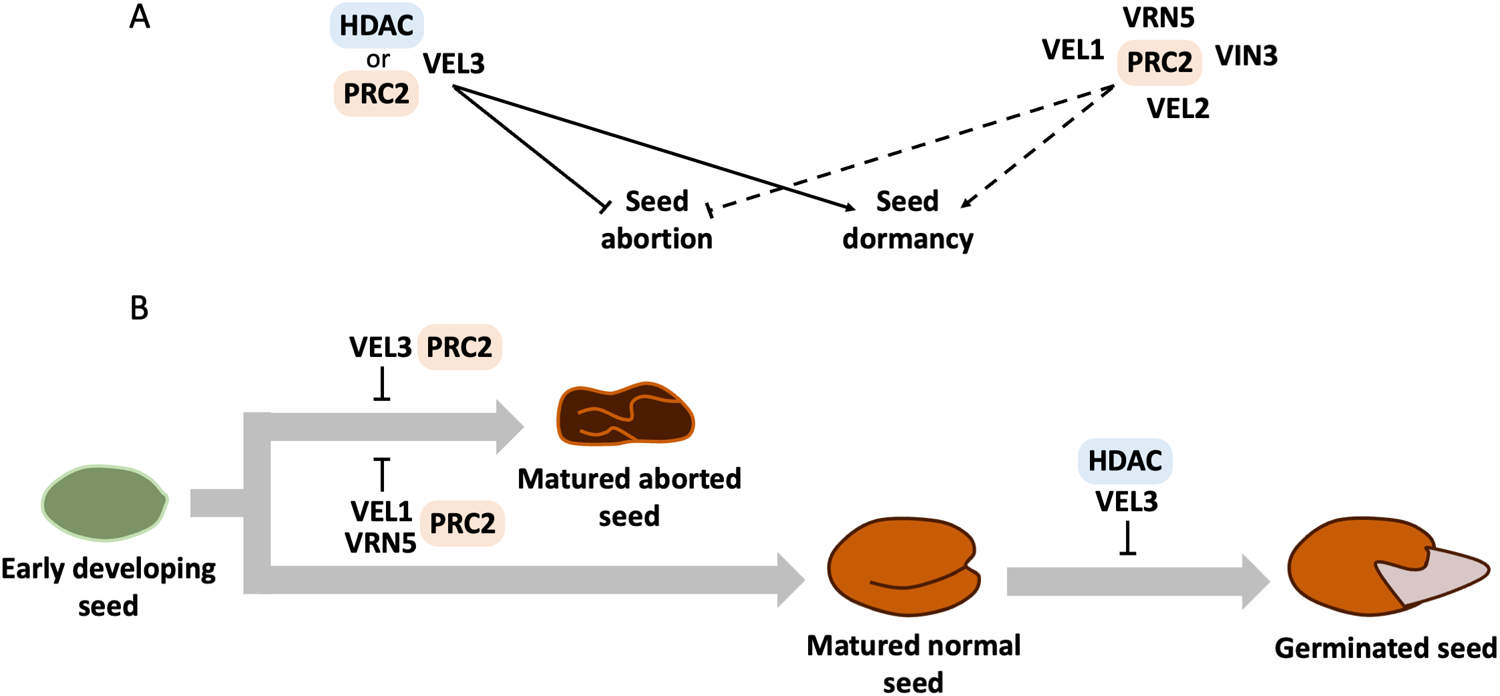
In this study, we dissect the distinct roles of the VEL family members in seeds (Fig. 5A), uncovering VEL3 as the main regulator of seed dormancy and abortion processes, while VEL1 and VRN5 assume more specialized with redundant functions in controlling seed abortion. Genetic analyses confirm that only VEL3 plays a role in the depth of primary dormancy (Fig. 3B). Although VEL2 is highly expressed in the endosperm of developing seeds (Belmonte et al., Reference Belmonte, Kirkbride, Stone, Pelletier, Bui, Yeung, Hashimoto, Fei, Harada, Munoz, Le, Drews, Brady, Goldberg and Harada2013), neither vel2 single mutants nor higher‐order combinations lacking VEL1, VEL2, VIN3 or VRN5 affect dormancy (Fig. 2C Fig. 3A, B), thereby underscoring the unique, specific and non-redundant role of VEL3.
Hypotheses, experimental findings and proposed regulatory model. (A) VEL3, in association with HDAC and possibly PRC2, promotes seed dormancy and suppresses seed abortion in Arabidopsis (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). Based on this, we hypothesized that other VEL family members (VEL1, VEL2, VIN3 and VRN5) may exert similar functions through PRC2. Solid arrows indicate previously established regulatory effects, while dotted arrows represent putative control. (B) Experimental results revealed that VEL2 and VIN3 had no detectable effect on seed abortion or dormancy. In contrast, knockout mutations in VEL1 together with VRN5 suppressed seed abortion at levels comparable to vel3-1, suggesting the involvement of a VEL–PRC2 complex in abortion control. Only VEL3, however, contributed to promoting seed dormancy, consistent with a role for the VEL3–HDAC complex in dormancy established in Chen et al. (Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). Grey thick arrows depict the timeline from early seed development through maturation to germination.

Figure 5 Long description
The image consists of two diagrams labeled A and B. Diagram A illustrates the regulatory pathways involving VEL family proteins, HDAC and PRC2. Solid arrows indicate established effects, while dotted arrows represent putative control. VEL3 is associated with HDAC or PRC2, influencing seed abortion and dormancy. VEL1, VRN5, PRC2, VIN3 and VEL2 are also shown in the network. Diagram B depicts the seed development process from early developing seed to germinated seed. VEL3 and PRC2 lead to matured aborted seeds, while VEL1 and VRN5 with PRC2 result in matured normal seeds. HDAC and VEL3 are involved in the transition from matured normal seed to germinated seed.
Strikingly, loss of VEL1 and VRN5 causes extensive seed abortion without altering dormancy (Fig. 2A, C). Like VIN3, these VEL proteins are known to fine-tune deposition of H3K27me3 together with PRC2 (VRN-PRC2), most notably at FLC during vernalization (De Lucia et al., Reference De Lucia, Crevillén, Jones, Greb and Dean2008). We have recently shown that VEL3 similarly guides PRC2 to silence maternally derived PEGs (Paternally Expressed Genes) in the endosperm (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). Studies of triploid‐block and hybridization barriers demonstrate that parental‐genome dosage imbalances involve PRC2‐mediated H3K27me3 to control seed abortion. Indeed, the FIS–PRC2 complex is dedicated to reproductive development, controlling female gametophyte differentiation, endosperm cellularization and seed maturation (Grossniklaus and Paro, Reference Grossniklaus and Paro2014; Hugues et al., Reference Hugues, Jacobs and Roudier2020). These findings implicate a role for VRN-PRC2 in endosperm cellularization and control of seed abortion but suggest that this process is unrelated to dormancy phenotypes of vel3 mutants.
Comparative transcriptome profiling of vel1-2 vrn5-9 and vel3-1 matured endosperms reveals a shared deregulation of programmed cell‐death genes that drive endosperm elimination (Fig. 4). Together, these findings suggest that VEL1, VRN5 and VEL3 act in parallel pathways with partially overlapping but also non-identical targets, and that PRC2 functional disruption does not necessarily result in a seed dormancy phenotype (Fig. 5B). However, we have previously found that upregulation of ORE1 partially accounts for the low dormancy of vel3-1 (Chen et al., Reference Chen, MacGregor, Stefanato, Zhang, Barros-Galvão and Penfield2023). Thus, we cannot rule out that differences in the timing of ORE1 activation, effects on seed survival to maturity (which is necessary for dormancy testing) or other subtle differences account for the dormancy differences between vel3-1 and vel1 vrn5 double mutants.
Together with prior evidence for a partially redundant VEL1 and VRN5 mode of action in controlling FLC (De Lucia et al., Reference De Lucia, Crevillén, Jones, Greb and Dean2008), our results support a hypothesis in which VEL1 and VRN5 proteins recruit or stabilize PRC2 at specific endosperm loci (Fig. 5B). This functional separation underscores a modular chromatin framework in seed development, where distinct components separate seed viability from dormancy programmes.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0960258526100130.
Acknowledgements
We thank Caroline Dean for providing the vil1-2 (vel1-1) and vrn5-8 mutants.
Author contributions
TB-G and SP conceived the study; TB-G, XC and FS performed experiments; TB-G and SP analysed the data; TB-G and SP wrote the manuscript.
Funding statement
This work was supported by Biotechnology and Biological Sciences Research Council Grants BB/T003030/1 to SP.
Competing interests
The authors declare no competing interests.
Data availability
RNA-seq data have been deposited at European Nucleotide Archive under accession PRJEB100848.





 Open access
Open access